Research Article |
|
Corresponding author: Cornelis van Achterberg ( kees@vanachterberg.org ) Academic editor: Andreas Köhler
© 2023 Cornelis van Achterberg, John T. Smit, Toshko Ljubomirov.
This is an open access article distributed under the terms of the Creative Commons Attribution License (CC BY 4.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Citation:
van Achterberg C, Smit JT, Ljubomirov T (2023) Review of the European Eumenes Latreille (Hymenoptera, Vespidae) using morphology and DNA barcodes, with an illustrated key to species. ZooKeys 1143: 93-163. https://doi.org/10.3897/zookeys.1143.94951
|
Abstract
The European species of the potter wasp genus Eumenes Latreille, 1802 (Vespidae, Eumeninae) are illustrated and a new illustrated key to the 13 recognised species is presented. Eumenes mediterraneus aemilianus Guiglia, 1951 is synonymised with E. papillarius (Christ, 1791) (syn. nov.), E. obscurus André, 1884 and E. andrei Dalla Torre, 1894 with E. pedunculatus (Panzer, 1799) (syn. nov.) and E. crimensis Blüthgen, 1938 with E. sareptanus André, 1884 (syn. nov.).
Keywords
Biology, COI barcode, Eumeninae, new synonymy, potter wasp, systematics, taxonomy, variation
Introduction
The potter wasp genus Eumenes Latreille, 1802 (Vespidae, Eumeninae) is distributed nearly worldwide and one of the most common genera of Eumeninae foraging on small flowers with easily accessible nectar in southern Europe. The genus currently includes ca. 106 described species (plus 46 subspecies) divided into two subgenera: subgenus Eumenes Latreille (including all European species) and the small Neotropical subgenus Zeteumenoides Giordani Soika, 1972 with very slender first metasomal tergite (
The setosity of body parts is used extensively in existing literature. However, this is a variable and, therefore, rather problematic character, augmented by wear of the setae in aged specimens and concealed setae in wet and/or dirty specimens. In one species (E. pomiformis) the regularity of the setosity on the propleuron remains essential for separating it from the very similar E. subpomiformis. For all other species additional, though also variable, characters are presented. In most cases a reliable identification will be possible when several complete specimens of each sex of a population are available.
Females build nests consisting of one or several jug like mud cells (Fig.
Materials and methods
Identified material was used from the following collections: Naturalis Biodiversity Center (Leiden;

DNA extraction was conducted on single legs, using the NucleoMag 96 Tissue kit by Macherey-Nagel on a Thermo Scientific KingFisher Flex magnetic bead extraction robot, with a final elution volume of 150 µl. The standard COI barcoding fragment (

Sampled specimens from
| Taxon | ID number | BOLD accession number | Country |
|---|---|---|---|
| Eumenes coronatus |
|
NLHYM109-12 | Netherlands |
| Eumenes “coarctatus” |
|
NLHYM206-12 | Netherlands |
| Eumenes coarctatus |
|
NLHYM394-12 | Netherlands |
| Eumenes “coarctatus” |
|
NLHYM457-12 | Netherlands |
| Eumenes coronatus |
|
NLHYM608-12 | Netherlands |
| Eumenes pedunculatus |
|
NLHYM515-12 | Netherlands |
| Eumenes coarctatus |
|
NLHYM969-22 | Netherlands |
| Eumenes coarctatus |
|
NLHYM970-22 | Netherlands |
| Eumenes pedunculatus |
|
NLHYM971-22 | Netherlands |
| Eumenes lunulatus |
|
NLHYM973-22 | Bulgaria |
| Eumenes mediterraneus |
|
NLHYM974-22 | Bulgaria |
| Eumenes pomiformis |
|
NLHYM975-22 | Bulgaria |
| Eumenes papillarius |
|
NLHYM976-22 | Bulgaria |
| Eumenes papillarius |
|
NLHYM977-22 | Bulgaria |
| Eumenes coronatus |
|
NLHYM978-22 | Bulgaria |
| Eumenes coronatus |
|
NLHYM979-22 | Bulgaria |
| Eumenes coronatus |
|
NLHYM980-22 | Netherlands |
| Eumenes mediterraneus |
|
NLHYM981-22 | France |
| Eumenes mediterraneus |
|
NLHYM982-22 | Greece |
| Eumenes mediterraneus |
|
NLHYM983-22 | Bulgaria |
| Eumenes subpomiformis |
|
NLHYM984-22 | Bulgaria |
| Eumenes pomiformis |
|
NLHYM985-22 | Greece |
| Eumenes coarctatus | MZ626999 | ACUFIN803-13 | Finland |
| Eumenes coarctatus | MZ627515 | ACUFIN804-13 | Finland |
| Ancistrocerus trifasciatus |
|
NLHYM034-12 | Netherlands |
Results
Molecular data
In the Neighbour-Joining tree (using COI sequences) newly barcoded specimens from Bulgaria, France, Greece, and Netherlands are combined with unpublished older sequences from Finland and Netherlands (Fig.
First of all, it is clear that E. mediterraneus is a species complex and the characters used for its recognition seems to be insufficient. The lectotype of E. mediterraneus originates from Croatia (Dalmatia) and is most likely the same species as the sampled specimens from Bulgaria. An extensive survey is necessary to find out what the position of the taxa within this complex is.

Neighbour-Joining tree for barcoded European Eumenes species. Both E. coarctatus specimens (sampled 5 y ago for another project and not available for examination) among E. coronatus are obviously misidentified and, therefore, in quotation marks. The numbers are
The position of the E. coarctatus lunulatus specimen from Bulgaria in the NJ tree indicates that it is different from the sampled N and NW European specimens of E. c. coarctatus (whereas the two colour forms of the latter obviously belong together; Fig.
Illustrated key to European species of Eumenes
N.B. Species can only be reliably identified by a combination of characters. Setosity may be worn off in aged specimens or hardly visible in dirty specimens; therefore, aged or dirty specimens are easily misidentified. In addition, there is a considerable intraspecific variation.
| 1 | Females: antenna without terminal hook (a); clypeus often partly or entirely black (b); inner tooth of hind tarsal claw widened and apically more or less truncate (c), but intermediate in E. coronatus; [mandibles somewhat larger than in ♂] | 2 |
 |
||
| – | Males: antenna with terminal hook (aa); clypeus nearly always entirely yellow (bb); inner tooth of hind tarsal claw comparatively slender and apically acute (cc) | 16 |
 |
||
| 2 | Setae of occiput short to medium-sized (a); second and third antennal segments reddish or brownish ventrally (b), rarely entirely black in SW Europe; [pronotum often broadly yellow laterally] | 3 |
 |
||
| – | Setae of occiput long (aa; but sometimes worn off); second and third antennal segments black ventrally (bb) | 5 |
 |
||
| 3 | First metasomal segment tricoloured ventrally (a); clypeus entirely yellow (b) or with small medial black patch; parategula yellow (c); [basal half of antenna usually extensively yellowish brown ventrally; pronotum broadly yellow posteriorly] | E. cyrenaicus Blüthgen, 1938 |
 |
||
| – | First segment bicoloured ventrally (aa); clypeus partly black (bb); parategula black (cc) | 4 |
 |
||
| 4 | Apical third of clypeus broadly yellow laterally (a); setae of mesoscutum comparatively short anteriorly (b); clypeus less convex compared to face in lateral view (c) | E. dubius de Saussure, 1852 |
 |
||
| – | Apical third of clypeus broadly black laterally (aa); setae of mesoscutum comparatively long anteriorly (bb); clypeus more convex compared to face in lateral view (cc) | E. sareptanus André, 1884 |
 |
||
| 5 | Posterior part of first metasomal tergite comparatively slender in ventral view (a) and weakly convex dorsally in lateral view (b); apical lamella of second tergite often subhyaline or pale yellow (c), if dark brown then setae of second tergite medium-sized (d) to long (dd) in lateral view; [hind basitarsus often dark brown basally] | 6 |
 |
||
| – | Posterior part of first tergite comparatively robust in ventral view (aa) and more convex dorsally in lateral view (bb); apical lamella of second tergite dark brown or blackish (cc), rarely yellowish; setae of second tergite either long (dd) or inconspicuous in lateral view (ddd) | 10 |
 |
||
| 6 | Second metasomal tergite with three large (more or less separated) black spots medially (a); first metasomal segment largely orange or yellow (b); clypeus entirely yellow (c); mesoscutum with medium-sized to large yellow or orange patch laterally (d); [propleuron with short setae] | E. tripunctatus (Christ, 1791) |
 |
||
| – | Second metasomal tergite with wide black band medially (aa); first segment mainly black (bb); clypeus at least medially partly black (cc); mesoscutum usually entirely black (dd) or with pair of small to large patches (large specimens of E. papillarius) | 7 |
 |
||
| 7 | Second metasomal sternite with long setae (a), rarely intermediate; scape entirely black anteriorly (b), rarely with short yellow stripe; inner tooth of hind tarsal claw narrower and rather acute (c); clypeus comparatively deeply emarginate medio-apically (d); [hind tarsus dark brown; dorsal setae of scape often long; second tergite rather remotely punctate and bristly setose] | E. coronatus (Panzer, 1799) |
 |
||
| – | Second sternite with short to medium-sized setae (aa), rarely intermediate (Iberian Peninsula); anteriorly scape (except sometimes apical third or half) yellow (bb); inner tooth of hind tarsal claw wider and distinctly truncate (cc); clypeus less emarginate medio-apically (dd); but comparatively deep in E. mediterraneus (ddd) | 8 |
 |
||
| 8 | Apical lamella of second tergite subhyaline or pale yellowish (a); ventral corners of clypeus narrower because of deeper medio-apical emargination (b); propodeum usually with medium-sized smooth interspaces (c) and anterior half of median groove distinct (d); second tergite comparatively convex medially and with short setae (e); first tergite slender in dorsal view (f); parategula frequently more or less yellow (g); labrum partly or entirely yellow (h); [hind basitarsus often brown basally] | E. mediterraneus Kriechbaumer, 1879 |
 |
||
| – | Apical lamella of second tergite brownish or blackish (aa), rarely pale brown or pale yellowish; ventral corners of clypeus wider apically because of shallow medio-apical emargination (bb); propodeum usually without smooth interspaces (cc) and anterior half of median groove largely reduced (dd); second tergite less convex medially and with medium-sized to long setae (ee); first tergite less slender in dorsal view (ff); parategula usually black (gg) or largely so; labrum entirely dark brown or black (hh) | 9 |
 |
||
| 9 | Third tergite and sternite partly yellow posteriorly (a); hind basitarsus more or less darkened basally and outer apex of hind tibia with blackish patch dorsally (b); first metasomal tergite elongate in lateral view (c); first tergite usually less densely punctate subposteriorly (d); second tergite more shiny and usually less densely sculptured laterally (e); [large specimens (fore wing about 10 mm) have mesoscutum frequently with a pair of yellow patches antero-laterally; if hind tibia apically and basitarsus basally yellow and second tergite with satin sheen laterally, go to 10] | E. papillarius (Christ, 1791) |
 |
||
| – | Third tergite and sternite black posteriorly (aa); hind basitarsus and outer apex of hind tibia entirely yellow (bb); first tergite less elongate in lateral view (cc); first tergite more densely punctate subposteriorly (dd); second tergite more densely sculptured dorsally (ee) | E. sardous Guiglia, 1951 |
 |
||
| 10 | Middle of propodeum with small interspaces between coarse punctures micro-sculptured (a); mesoscutum with pair of yellow (and often large) patches antero-laterally (b); clypeus with coarser punctures, especially apically (c); length of fore wing 10–13 mm | E. punctaticlypeus Giordani Soika, 1943 |
 |
||
| – | Middle of propodeum either without interspaces between punctures (aa) or small interspaces present and smooth (aaa); mesoscutum entirely black or with smaller linear patches antero-laterally (bb); clypeus often only punctulate or with less coarse punctation, especially apically (cc); length of fore wing 7–11 mm | 11 |
 |
||
| 11 | Apical half of scape anteriorly (a) and clypeus (b) black or largely so; second sternite with narrow yellow band apically (c) | 12 |
 |
||
| – | Apical half of scape anteriorly (aa) usually and clypeus (bb) partly yellow; second sternite with wider yellow band apically (cc) | 13 |
 |
||
| 12 | Pronotum flattened medio-posteriorly and narrowly yellow (a); apical corners of clypeus more protruding (b); third metasomal tergite entirely black (c); inner tooth of hind tarsal claws shorter and wider (d) and claw curved (e); [if second sternite with some long setae and first tergite comparatively slender, see E. coronatus] | E. coarctatus coarctatus (Linnaeus, 1758) |
 |
||
| – | Pronotum convex medio-posteriorly and wider yellow (aa); apical corners of clypeus less protruding (bb); third tergite partly yellow (cc); inner tooth of hind tarsal claws slightly longer and narrower (dd) and claw less curved (ee); [if outer side of hind coxa mostly with short setae, shape of hind claws different, second tergite more punctate and fore tibia entirely yellow, compare with very similar E. coarctatus lunulatus with darkened scape] | E. pedunculatus (Panzer, 1799) |
 |
||
| 13 | Clypeus yellow medio-dorsally (a) and apical half largely black (b); posterior half of first tergite usually more robust in dorsal view (c); medio-posteriorly third sternite yellow (d); apical corners of clypeus slightly less protruding (e) | 14 |
 |
||
| – | Clypeus black medio-dorsally (aa), if yellow (aaa) then apical half of clypeus also largely yellow (bb); posterior half of first tergite less robust in dorsal view (cc); medio-posteriorly third sternite usually black (dd) or yellow band reduced; apical corners of clypeus slightly more protruding (ee) | 15 |
 |
||
| 14 | Medio-posteriorly third metasomal sternite broadly black or dark brown (a); outer side of hind coxa mostly with very long setae in dorsal view (b); first tergite comparatively convex in lateral view (c); head conspicuously long setose (d); fore tibia often with dark brown or blackish patch medio-posteriorly (e); second tergite finely punctate (f); [yellow dorsal part of clypeus transverse | E. coarctatus coarctatus (Linnaeus, 1758) |
 |
||
| – | Medio-posteriorly third sternite yellow (aa) or narrowly interrupted; hind coxa with short to medium-sized setae in dorsal view (bb); first tergite less convex in lateral view (cc); head less conspicuously setose (dd); fore tibia entirely yellow (ee) or with blackish patch medio-posteriorly; second tergite more coarsely punctate (ff) | E. coarctatus lunulatus Fabricius, 1804 |
 |
||
| 15 | Laterally propleuron regularly short setose in lateral view, setae curved and 0.1–0.3 times as long as occipital setae (a); hind tarsus (except dark brown telotarsus) brownish yellow (b); [apical lamellae of propodeum more or less darkened] | E. pomiformis (Fabricius, 1781) |
 |
||
| – | Laterally propleuron irregularly medium-sized setose in lateral view, setae straight (aa) or adpressed; hind tarsus dark brown (bb) or largely so | E. subpomiformis Blüthgen, 1938 |
 |
Males
| 16 | Setae of occiput short to medium-sized (a); second and third antennal segments reddish or brownish ventrally (b); [antennal hook distinctly bent; laterally pronotum often broadly yellow] | 17 |
 |
||
| – | Setae of occiput long (aa; but sometimes worn off); second and third antennal segments black ventrally (bb) | 19 |
 |
||
| 17 | Antennal hook robust (a, b) and in lateral view claw-like (b); ventro-lateral corners of clypeus rather acute (c); [first tergite bicoloured ventrally; mesoscutal setosity short] | E. dubius de Saussure, 1852 |
 |
||
| – | Antennal hook more slender (aa, bb) and less claw-like in lateral view (bb), but sometimes intermediate in SW Europe; ventro-lateral corners of clypeus slightly more obtuse (cc); [first tergite tri- or bicoloured ventrally] | 18 |
 |
||
| 18 | First metasomal segment tricoloured ventrally (a); antennal hook more slender in lateral view (b; sometimes less than shown); setae of eye incision at most half as long as apical width of scape in lateral view (c); parategula largely yellow (d), rarely dark brown; [antenna extensively yellowish brown ventrally as in E. dubius palaestinensis Blüthgen, 1938] | E. cyrenaicus Blüthgen, 1938 |
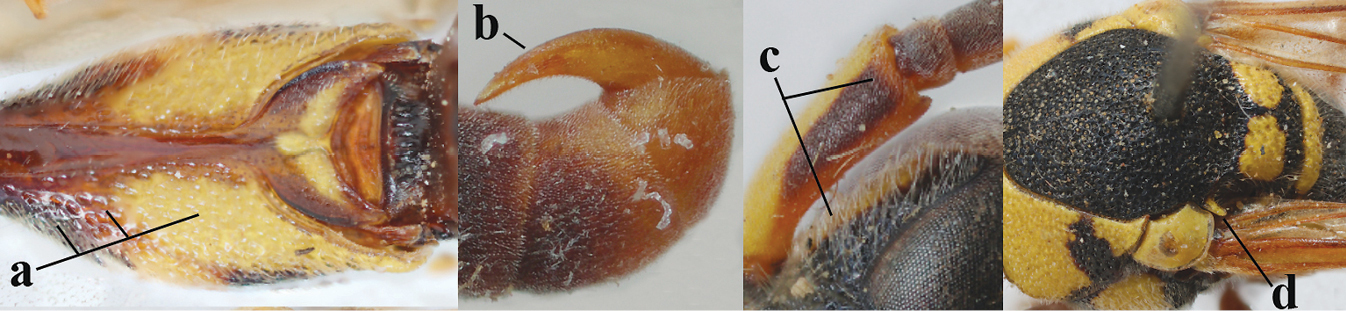 |
||
| – | First segment bicoloured ventrally (aa); antennal hook less slender in lateral view (bb); setae of eye incision about as long as apical width of scape in lateral view (cc); parategula black (dd); [if antennal hook minute and nearly straight, cf. E. pomiformis with short occipital setae] | E. sareptanus André, 1884 |
 |
||
| 19 | Apical half of first metasomal segment largely orange or yellow dorsally (a); mesoscutum with medium-sized to large yellow or orange patch laterally (b); [propleuron with short setae] | E. tripunctatus (Christ, 1791) |
 |
||
| – | Apical half of first segment largely black dorsally (aa); mesopleuron mainly or entirely black anteriorly (bb); mesoscutum black (cc) or with small transverse patch laterally (ccc), rarely larger | 20 |
 |
||
| 20 | Middle of propodeum with matt, micro-sculptured interspaces (a); mesoscutum with pair of transverse yellow patches antero-laterally (b), but sometimes reduced; apical antennal hook robust apically and dark brown (c); base of mandible often partly pale yellowish (d); length of fore wing 9–13 mm; [posterior half of first tergite very robust] | E. punctaticlypeus Giordani Soika, 1943 |
 |
||
| – | Middle of propodeum without micro-sculptured interspaces (aa); mesoscutum entirely black (bb); apical antennal hook slender apically and yellow (cc), rarely infuscate or dark brown (E. subpomiformis); base of mandible black (dd), rarely with small yellow patches; length of fore wing 7–11 mm | 21 |
 |
||
| 21 | Second metasomal sternite with some long setae (a), rarely medium-sized; posterior part of first tergite flattened in lateral view (b); scape black anteriorly or largely so (NW Europe; c), but more or less yellow in S Europe; ventro-apical corners of clypeus narrower (d); first tergite without pair of yellow spots (e) or spots minute; [second tergite conspicuously setose and remotely sculptured] | 22 |
 |
||
| – | Second sternite with short or medium-sized setae only (aa); posterior part of first tergite convex in lateral view (bb); scape yellow anteriorly (c) or largely so; ventro-apical corners of clypeus wider (d); usually first tergite with pair of yellow spots (ee), but often reduced in E. papillarius | 23 |
 |
||
| 22 | Second tergite coarser punctate (a); first tergite slightly less robust in ventral view (b); third tergite more extensively yellow (c); setosity of scape either long dorsally (d) or (Balkan Peninsula) short | E. coronatus (Panzer, 1799) |
 |
||
| – | Second tergite comparatively finely punctate (aa); first tergite slightly more robust in ventral view (bb); third tergite less extensively yellow (cc); scape short setose dorsally (dd); [dark form with dark scape, more or less partly long setose second sternite and finely punctate second tergite] | E. c. coarctatus (Linnaeus, 1758) |
 |
||
| 23 | Antennal hook short, bent and slightly wider medially (a); apical lamella of second tergite subhyaline or pale yellow (b); second tergite more or less concave medio-posteriorly in lateral view (c); clypeus slightly more emarginated medio-apically (d) | E. mediterraneus Kriechbaumer, 1879 |
 |
||
| – | Antennal hook longer, nearly straight (but medio-dorsally slightly depressed) and medially more slender (aa), but intermediate in E. subpomiformis; apical lamella of second tergite dark brown or blackish (bb), but yellow in E. sardous; second tergite flat or weakly concave medio-posteriorly in lateral view (cc); clypeus less emarginated medio-apically (dd) | 24 |
 |
||
| 24 | Outer side of hind tibia with black or dark brown patch dorso-apically, contrasting with mainly pale hind basitarsus (a); first tergite comparatively slender in ventral (b) and dorsal (c) view; apical half of antennal hook with more or less dark brown keel dorsally and relatively slender (d); [second metasomal tergite shiny apico-laterally] | E. papillarius (Christ, 1791) |
 |
||
| – | Outer side of hind tibia dorso-apically and hind basitarsus yellow (aa); first tergite usually more robust in ventral (bb) and dorsal (cc) view; apical half of antennal hook without dark brown keel dorsally (dd), if slightly developed then less slender; [second tergite usually with satin sheen apico-laterally] | 25 |
 |
||
| 25 | Third and fourth metasomal tergites black (a), rarely third tergite narrowly yellow; mesoscutum scrobiculate-reticulate (b); apical lamella of second tergite yellow or brownish (c); [antennal hook with submedial depression] | E. sardous Guiglia, 1951 |
 |
||
| – | Third and fourth tergites partly yellow (aa); mesoscutum coarsely punctate or punctate-reticulate (bb); apical lamella of second tergite dark brown or blackish (cc) | 26 |
 |
||
| 26 | Setae of propleuron regular and 0.1–0.3 times as long as occipital setae (a); antennal hook nearly straight (a; sometimes slightly more curved than illustrated) | E. pomiformis (Fabricius, 1781) |
 |
||
| – | Setae of propleuron irregular and mainly straight and 0.3–0.7 times as long as occipital setae (aa); antennal hook more curved (bb) | 26 |
 |
||
| 27 | Antennal hook largely dark brown or brown (a), more slender and slightly bent (b); penultimate antennal segment dark brown ventrally (c); second metasomal tergite usually rather bristly setose (d) | E. subpomiformis Blüthgen, 1938 |
 |
||
| – | Antennal hook yellow (aa) or largely so, less slender and more bent (bb); penultimate antennal segment yellowish brown ventrally (cc); second metasomal tergite usually only short setose (dd), but bristly in typical E. coarctatus (ddd) | 27 |
 |
||
| 28 | Outer side of hind coxa with short to medium-sized setae in dorsal view (a); first metasomal tergite usually slightly less convex in lateral view (b); head moderately setose (c); apical yellow band of second tergite less widened dorsally (d) | E. coarctatus lunulatus Fabricius, 1804 |
 |
||
| – | Outer side of hind coxa with very long setae in dorsal view (aa); first tergite comparatively convex in lateral view (bb); head conspicuously long setose (cc); apical yellow band of second tergite often comparatively wide dorsally (dd) | 29 |
 |
||
| 29 | Second metasomal tergite moderately to coarsely punctate (a) and usually rather dull (b); inner side of apical hook with small setae (c); [pale form of E. coarctatus with more or less coarsely punctate second tergite, long setose hind coxae and short setose second sternite; f. barbatulus Blüthgen, 1943] | E. coarctatus coarctatus (Linnaeus, 1758) |
 |
||
| – | Second metasomal tergite sparsely punctate or punctulate (aa) and shiny (bb); inner side of apical hook without distinct setae (cc) | E. pedunculatus (Panzer, 1799) |
 |
Species accounts
Eumenes c. coarctatus
Vespa coarctata Linnaeus, 1758: 573.
Eumenes coarctatus coarctatus; Gusenleitner, 1972: 75, 1999: 569;
Eumenes coarctatus;
Eumenes (Eumenes) coarctatus;
Eumenes (Eumenes) coarctatus coarctatus;
Eumenes pomiformis dernaensis
Blüthgen, 1938: 494;
Eumenes lunulatus dernaensis;
Eumenes coarctatus dernansis
(!);
Eumenes coarctatus dernaensis;
Eumenes pomiformis barbatulus
Blüthgen, 1943: 303;
Eumenes pedunculata var. punctata
Hellén, 1944: 11;
Eumenes pedunculatus var. turaniformis
Blüthgen, 1959: 13;
Eumenes coarctatus turaniformis;
Eumenes (Eumenes) coarctatus turaniformis;
Eumenes coarctatus corsicus
Gusenleitner, 1972: 77, 1999: 569 (as synonym of E. coarctatus);
Eumenes coarctatus maroccanus
Gusenleitner, 1972: 76–77, 1999: 569, 2013: 26;
Eumenes (Eumenes) coarctatus maroccanus;
Eumenes coarctatus nugaricus
Giordani Soika, 1986: 123;
Eumenes (Eumenes) coarctatus nuragicus;
Notes
The holotype female of E. coarctatus is heavily damaged (e.g., antenna completely missing) but the metasoma is preserved showing the robust first tergite in lateral view (https://linnean-online.org/16751/) and the second sternite lacks long setae (see
Distribution
The dark typical form is mostly boreo-alpine of distribution and occurs in Scandinavia (up to S Finland and SE Sweden), UK (England and Wales) and mountainous regions in Central Europe (reaching 1550 m altitude in Switzerland). The pale typical form (= f. barbatulus Blüthgen, 1943) occurs in mainly C and SW Europe and is often the most common species. Outside Europe known from N Africa and the East Palaearctic region up to China and Japan. Only breeding species in UK where it is considered a most threatened species and is known as the Heath Potter Wasp (https://naturebftb.co.uk/artwork/heath-tiger-beetle-alex-hyde/pots-of-the-heath-potter-wasp-eumenes-coarctatus/).
Eumenes coarctatus lunulatus
Eumenes lunulata
Fabricius, 1804: 290;
Eumenes lunulatus;
Eumenes (Eumenes) lunulatus lunulatus;
Eumenes lunulatus lunulatus;
Eumenes coarctatus lunulatus;
Eumenes pomiformis ordubadensis
Blüthgen, 1938: 493;
Eumenes (Eumenes) lunulatus ordubadensis;
Eumenes lunulatus ordubadensis;
Eumenes coarctatus ordubadensis;
Eumenes (Eumenes) coarctatus ordubadensis;
Eumenes pomiformis limissicus
Blüthgen, 1938: 493;
Eumenes lunulatus limissicus; Gusenleitner 1970: 163, 1972: 80–81.
Eumenes (Eumenes) lunulatus limissicus;
Eumenes coarctatus limissicus;
Eumenes (Eumenes) coarctatus limissicus;
Eumenes lunulatus var. tenebricosus
Gusenleitner, 1972: 79;
Eumenes lunulatus tenebricosus;
Eumenes lunulatus var. balcanicus
Gusenleitner, 1972: 79;
Eumenes coarctatus;
Notes
The holotype female of E. lunulatus was digitally examined by using photographs kindly supplied by Sree Gayathree Selvantharan and Lars Vilhelmsen (NHMD). Unfortunately, the head is missing, but the remaining body parts agree with the current interpretation.

Eumenes coarctatus coarctatus (Linnaeus), Netherlands (Otterloo), female 4 metasoma lateral 5 first metasomal tergite dorsal 6 first tergite ventral 7 mesosoma dorsal 8 second metasomal tergite latero-dorsal 9 head anterior 10 head and propleuron lateral 11 antenna 12 hind tarsal claw.

Eumenes coarctatus coarctatus (Linnaeus), Netherlands (Otterloo), male 13 metasoma lateral 14 first metasomal tergite dorsal 15 first tergite ventral 16 mesosoma dorsal 17 second metasomal tergite dorsal 18 head anterior 19 head and propleuron lateral 11 apical half of antenna 12 apical hook of antenna lateral.


Eumenes coarctatus lunulatus Fabricius, Bulgaria, male 31 metasoma lateral 32 first metasomal tergite dorsal 33 first tergite ventral 34 head and mesosoma dorsal 35 second metasomal tergite dorso-lateral 36 head anterior 37 head and propleuron lateral 38 apical hook of antenna lateral 39 hind coxa dorsal.
Distribution
SE and C Europe, NW Asia. Examined specimens originating from Austria (type locality), Hungary, Czech Republic, and Slovakia (but most specimens from these countries in collections belong to E. coarctatus sensu stricto), Italy, Bulgaria, Turkey, Greece (in Peloponnesus the most common species according to
Eumenes coronatus
Vespa coronata Panzer, 1799: (6) 64: 12, pl. 12.
Eumenes (Eumenes) coronatus coronatus;
Eumenes coronatus;
Eumenes (Eumenes) coronatus;
Eumenes coronatus coronatus;
Eumenes atricornis
Fabricius, 1804: 289;
Eumenes costata
(!) Lucciani, 1845: CX;
Eumenis
(!) mediterranea var. neesi Kriechbaumer, 1879: 88;
Eumenes coarctatus var. opulenta
Blüthgen, 1938: 482–483;
Eumenes coarctatus detonsus
Blüthgen, 1943: 297;
Eumenes (Eumenes) coronatus detonsus;
Eumenes coarctatus ab. nigrotibia
Hellén, 1944: 11;
Eumenes coarctatus var. niger
Szulczewski, 1950: 8 (invalid homonym; as synonym of E. coronatus);
Eumenes coarctatus ibericus
Blüthgen, 1956: 2;
Eumenes coronatus corruetus
Gusenleitner, 1972: 87, 1999: 570–571, 2013: 26;
Eumenes (Eumenes) coronatus corruetus;
Notes
Males from the Balkan Peninsula have the apical half of antennal hook distinctly flattened, different from the wider apical half in Central European males (Fig.

Distribution
Rather common in most of Europe (including southern Scandinavia;


Eumenes cyrenaicus Blüthgen, Italy (Sardinia), female 55 metasoma lateral 56 first metasomal tergite dorsal 57 first tergite ventral 58 mesosoma dorsal 59 mesoscutum and scutellum lateral 60 head anterior 61 head and propleuron lateral 62 hind tarsal claw 63 antenna 64 second metasomal tergite dorso-lateral.

Eumenes cyrenaicus Blüthgen, Morocco, male 65 metasoma lateral 66 first metasomal tergite dorsal 67 first tergite ventral 68 head and mesosoma dorsal 69 head posteriorly and propleuron lateral 70 mesoscutum and scutellum lateral 71 head anterior 72 head and mesosoma lateral 73 antenna 74 apical hook of antenna lateral.
Eumenes cyrenaicus
Eumenes dubius cyrenaicus Blüthgen, 1938: 464, 468.
Eumenes (Eumenes) dubius cyrenaicus;
Eumenes (Eumenes) cyrenaicus cyrenaicus;
Eumenes dubius cyrenaicus var. congruens
Blüthgen, 1938: 464;
Eumenes dubius dubius var. pseudogermanica Blüthgen, 1938: 464.
Eumenes (Eumenes) dubius pseudogermanicus;
Eumenes cyrenaicus pseudogermanicus;
Eumenes (Eumenes) cyrenaicus pseudogermanicus;
Notes
Eumenes cyrenaicus is similarly coloured as E. dubius f. palaestinensis Blüthgen, 1938 from Asia Minor, but E. cyrenaicus has the yellow stripe of the eye incision narrow or absent (wide in E. dubius f. palaestinensis) and the clypeus sparser setose (densely silvery setose in E. dubius f. palaestinensis). Males can be separated by the shape of the antennal hook (in ventral view normal in E. cyrenaicus and widened in E. dubius f. palaestinensis) and sculpture of fifth sternite (distinctly punctate in E. cyrenaicus and punctulate in E. dubius f. palaestinensis). Typical E. dubius f. palaestinensis has the apical lamella of the second tergite yellow and in E. cyrenaicus light brown or yellowish (Fig.
Distribution
North Africa, South Europe (*Spain, *Portugal, Italy (Sardinia, Sicily), *N Macedonia, *Bulgaria, *Greece) and *Turkey.
Eumenes dubius
Eumenes dubia de Saussure, 1852: 32 (depository of type series unknown).
Eumenes dubius;
Eumenes dubius dubius;
Eumenes (Eumenes) dubius dubius;
Eumenes (Eumenes) dubius;
Eumenes (Eumenes) dubius dubius var. palaestinensis
Blüthgen, 1938: 467;
Eumenes (Eumenes) dubius palaestinensis;
Eumenes (Eumenes) dubius dubius var. macedonica
Blüthgen, 1952: 5, 15;
Eumenes (Eumenes) dubius macedonicus;
Notes
Rarely collected species in C and S Europe, but common in Spain and Portugal (
Eumenes dubius de Saussure, Bulgaria, female 75 metasoma lateral 76 first metasomal tergite dorsal 77 first tergite ventral 78 mesosoma dorsal 79 second metasomal tergite latero-dorsal 80 head anterior 81 head and propleuron lateral 82 antenna 83 hind tarsal claws 84 mesoscutum and scutellum lateral.

Eumenes dubius de Saussure, Bulgaria, male 85 metasoma lateral 86 first metasomal tergite dorsal 87 first tergite ventral 88 head and mesosoma dorsal 89 apical hook of antenna lateral 90 head anterior 91 head and propleuron lateral 92 apical hook of antenna latero-ventral 93 mesoscutum and scutellum lateral.
Distribution
Asia, Central and South Europe. Absent in Switzerland according to
Eumenes mediterraneus
Eumenis (sic!) mediterraneus Kriechbaumer, 1879: 85.
Eumenes (Eumenes) mediterraneus mediterraneus;
Eumenes (Eumenes) mediterraneus;
Eumenes mediterraneus;
Eumenes mediterraneus mediterraneus;
Eumenes affinissima race quettaensis
Cameron, 1907: 132–133;
Eumenes (Eumenes) mediterraneus quettaensis;
Eumenes (Eumenes) mediterraneus quettaensis;
Labus superbus
Meade-Waldo, 1910: 36;
Eumenes (Eumenes) mediterraneus superbus;
Eumenes mediterraneus bengasinus
Blüthgen, 1938: 487;
Eumenes (Eumenes) mediterraneus bengasinus;
Eumenes mediterraneus cypricus
Blüthgen, 1938: 488;
Eumenes (Eumenes) mediterraneus cypricus;
Eumenes (Eumenes) houskai
Giordani Soika, 1952a: 17;
Eumenes (Eumenes) mediterraneus anatolicus
Giordani Soika, 1952b: 376;
Eumenes mediterraneus manchurianus
Giordani Soika, 1971: 70;
Eumenes (Eumenes) mediterraneus manchurianus;
Eumenes mediterraneus var. opacus
Gusenleitner, 1972: 92;
Eumenes mediterraneus filitosa
Gereys, 2011: 224–225, 2016: 132;
Notes
This species is in need of a critical revision; the few molecular data indicate that several cryptic species may be included under E. mediterraneus (Fig.


Eumenes mediterraneus Kriechbaumer, Bulgaria, male 103 metasoma lateral 104 first metasomal tergite dorsal 105 first tergite ventral 106 head and mesosoma dorsal 107 propodeum dorsal 108 head anterior 109 head and mesosoma lateral 110 antenna anterior 111 apical hook of antenna lateral.
Distribution
Mediterranean, Balkan Peninsula, rarely in Central Europe (e.g., Switzerland only in Ticino and Valais and late in season (July–October;
Eumenes papillarius
Sphex papillarius Christ, 1791: 325.
Eumenes papillarius;
Eumenes (Eumenes) papillarius papillarius;
Eumenes papillarius papillarius;
Eumenes (Eumenes) papillarius;
Eumenes bipunctis
de Saussure, 1852: 33;
Eumenes bimaculatus
André, 1884: 645;
Eumenes papillarius var. baltica
Blüthgen, 1938: 485;
Eumenes (Eumenes) papillarius balticus;
Eumenes papillarius balticus;
Eumenes mediterraneus aemilianus Guiglia, 1951: 28. Syn. nov.
Eumenes (Eumenes) mediterraneus aemilianus;
Eumenes aemilianus;
Eumenes papillarius monticola
Blüthgen, 1956: 2;
Eumenes papillarius rubricornis
Giordani Soika (in Gusenleitner), 1972: 90;
Eumenes (Eumenes) papillarius rubricornis;
Notes
Large specimens (fore wing length about 10 mm) have frequently a pair of yellow patches on the mesoscutum antero-laterally; the patches vary from minute to large. The photographs of the female holotype kindly supplied by Roberto Poggi (

Eumenes papillarius (Christ), France, female 112 metasoma lateral 113 first metasomal tergite dorsal 114 first tergite ventral 115 head and mesosoma dorsal 116 second tergite latero-dorsal 117 head anterior 118 head and pronotum lateral 119 antenna anterior 120 hind tarsal claw 121 hind tibia lateral.

Eumenes papillarius (Christ), France, male 122 metasoma lateral 123 first metasomal tergite dorsal 124 first tergite ventral 125 head and mesosoma dorsal 126 second tergite dorsal 127 hind tibia lateral 128 head anterior 129 head and mesosoma lateral 130 antenna anterior 131 apical hook of antenna lateral.
Distribution
Widespread in most of Europe, but considered absent from UK (only known as vagrant in England;
Eumenes pedunculatus
Vespa pedunculata Panzer, 1799: (6) 63: 8, pl. 8.
Eumenes (Eumenes) pedunculatus pedunculatus;
Eumenes pedunculatus;
Eumenes pedunculatus pedunculatus;
? Eumenes marginella Herrich-Schäffer, 1841: 44, pl. 179-8.
? Eumenes (Eumenes) marginellus;
Eumenes obscurus André, 1884: 636–637. Syn. nov.
Eumenes andrei
Dalla Torre, 1894: 17 (new name for junior homonym E. obscurus André);
Eumenes eburneopictus
Giordani Soika, 1940: 97;
Eumenes (Eumenes) eburneopictus;
Eumenes pedunculatus eburneopictus;
Eumenes pedunculatus turanus
Blüthgen, 1943: 302;
Eumenes (Eumenes) pedunculatus turanus;
Eumenes pedunculata var. lapponica
Hellén, 1944: 11;
Eumenes karafutonis
Yamane, 1977: 61–62;
Notes
The depository of the female holotype of E. obscurus André (= E. andrei Dalla Torre) is unknown, but the extensive description allows identification. The robust posterior part of the first tergite (in dorsal view campaniform), the entirely dark antenna, the black clypeus except for a yellow dorsal linear patch and the shiny and very finely punctate second tergite points to E. pedunculatus (Panzer). The type series of E. marginellus is lost; the more or less yellow scape, the black scutellum, the narrow yellow patch of the pronotum and narrow yellow posterior patch of the first tergite are similar to some examined specimens of E. pedunculatus.
Distribution
Widely distributed in Europe but relatively rare in collections from NW and S Europe (e.g., only Eumenes sp. known from Norway, absent from UK and Corsica, in S Europe rare and restricted to montane habitats (


Eumenes pedunculatus (Panzer), Netherlands (Helenaveen), male 140 metasoma lateral 141 first metasomal tergite dorsal 142 first tergite ventral 143 head and mesosoma dorsal 144 second tergite dorsal 145 scape anterior 146 head anterior 147 head and mesosoma lateral 148 antenna anterior 149 apical hook of antenna lateral.
Eumenes pomiformis
Vespa pomiformis Fabricius, 1781: 467.
Eumenes pomiformis pomiformis;
Eumenes pomiformis;
Eumenes (Eumenes) pomiformis;
? Vespa histrio de Villers, 1789: 282–283. Type series lost.
? Eumenes (Eumenes) histrio;
Eumenis
(sic!) mediterranea var. heri Kriechbaumer, 1879: 88;
Eumenes fastidiosissimus
Giordani Soika, 1943: 29;
Eumenes pomiformis turcicus
Giordani Soika, 1952: 367;
Notes
The female lectotype of E. pomiformis was examined digitally by photographs kindly supplied by Sree Gayathree Selvantharan and Lars Vilhelmsen (NHMD), as the male holotype of E. heri (photographs kindly supplied by Stephan and Olga Schmidt (
Distribution
One of the common species in S Europe, reaching Germany (but very rarely collected) and Belarus. Known from Corsica, Sardinia, Sicily, and Malta (


Eumenes pomiformis (Fabricius), Italy, male 159 metasoma lateral 160 first metasomal tergite dorsal 161 first tergite ventral 162 head and mesosoma lateral 163 second tergite dorso-lateral 164 antenna 165 head anterior 166 head and mesosoma lateral 167 head and mesosoma dorsal 168 hind femur and tibia lateral 169 apical hook of antenna lateral.
Eumenes punctaticlypeus
Eumenes robusta Kostylev, 1940: 141 (primary homonym; not E. robustus Isely, 1917).
Eumenes (Eumenes) robustus;
Eumenes (Eumenes) punctaticlypeus
Giordani Soika, 1943: 29;
Eumenes punctaticlypeus punctaticlypeus; Gusenleitner, 1999: 573.
Eumenes punctaticlypeus;
Eumenes (Eumenes) calabricus
Giordani Soika, 1943: 31; Giordani Soika 1956: 316;
Eumenes kostylevi
Kurzenko, 1976: 437 (replacement name for E. robusta);
Eumenes kostylevi kostylevi;
Eumenes kostylevi punctaticlypeus;
Eumenes punctaticlypeus kostylevi;
Notes
As shown by the short setae of the hind coxa, the robust posterior part of the first metasomal tergite and frequently present moon-shaped yellow patch of the clypeus in females of both E. punctaticlypeus and E. lunulatus, the first one could be considered a large form of the latter. We recognise E. punctaticlypeus as a separate species because of the dark brown antennal hook of the males (yellow in E. lunulatus), differences in sculpture (but part may be the result of the larger body size) and the presence of a pair of large yellow spots on the mesoscutum of females (but the latter is variable in E. papillarius and this may be also the case in this species).
Distribution
Examined specimens are from Spain, France, Bulgaria, and Turkey. This rarely collected species is also reported from Albania, Italy (type series), Greece and Ukraine (Crimea).
Eumenes sardous
Eumenes sardous
Guiglia, 1951: 27;
Eumenes (Eumenes) sardous;
Notes
Similar to E. subpomiformis according to

Eumenes punctaticlypeus Giordani Soika, France, female 170 metasoma lateral 171 first metasomal tergite dorsal 172 first tergite latero-ventral 173 head and mesosoma dorsal 174 second tergite latero-dorsal 175 hind tarsal claw 176 head anterior 177 head and pronotum lateral 178 antenna anterior 179 propodeum dorsal.


Eumenes sardous Guiglia, France (Corsica), female 189 metasoma lateral 190 first metasomal tergite dorsal 191 first tergite ventral 192 head and mesosoma dorsal 193 second tergite latero-dorsal 194 head anterior 195 head and mesosoma lateral 196 antenna anterior 197 hind tarsal claw.

Eumenes sardous Guiglia, France (Corsica), male 198 metasoma lateral 199 first metasomal tergite dorsal 200 first tergite ventral 201 head and mesosoma dorsal 202 second tergite dorso-lateral 203 head anterior 204 head and mesosoma lateral 205 antenna anterior 206 apical hook of antenna lateral 207 id. of other antenna.
Distribution
An endemic species of Sardinia (Italy) and Corsica (France), occurring from sea level up to 1600 m altitude in Corsica (
Eumenes sareptanus
Eumenes sareptanus
André, 1884: 638;
Eumenes (Eumenes) sareptanus sareptanus;
Eumenes sareptanus sareptanus;
Eumenes (Eumenes) sareptanus;
Eumenes pomiformis f. insolata
Müller, 1923: 627;
Eumenes (Eumenes) sareptanus insolatus;
Eumenes sareptanus insolatus;
Eumenes dubius sareptanus var. germanica
Blüthgen, 1938: 469, 474, 495;
Eumenes dubius crimensis
Blüthgen, 1938: 468–469;
Eumenes (Eumenes) crimensis;
Eumenes sareptanus scabrosus;
Notes


Eumenes sareptanus André, Bulgaria, male 216 metasoma lateral 217 first metasomal tergite dorsal 218 first tergite ventral 219 mesosoma dorsal 220 second tergite latero-dorsal 221 mesoscutum and scutellum lateral 222 head anterior 223 head and mesosoma lateral 224 apical hook of antenna ventral 225 setosity of head latero-dorsal 226 antennal hook lateral.
Unfortunately, the original description does not include any remarks on the shape of the apical antennal segment of the male. The males should have the apical antennal segment narrower basally and less curved than in typical E. dubius according to Gusenleitner (1972). Michael Greeff (
Distribution
A comparatively rarely collected species in C and S Europe as well in NW Asia. The typical form occurs in southern European Russia up to western Siberia. In Switzerland occurring between 255 and 1250 m altitude (
Eumenes subpomiformis
Eumenes subpomiformis
Blüthgen, 1938: 480, 496;
Eumenes (Eumenes) subpomiformis;
Eumenes subpomiformis subpomiformis;
Eumenes subpomiformis crassipunctatus
Blüthgen, 1956: 3;
Notes
As pointed out by
Specimens in


Distribution
C and S Europe, but unknown from Sardinia (
Eumenes tripunctatus
Sphex tripunctatus Christ, 1791: 317 (type series lost).
Eumenes (Eumenes) tripunctatus;
Eumenes tripunctatus;
Vespa trimaculata
Lichtenstein, 1796: 202;
Eumenes venusta
Fischer-Waldheim, 1843: 1, pl. 122;
Note
Conspicuous orange species only recently known to occur in Europe (
Distribution
Central Asia, European Russia, Ukraine (Crimea).

Eumenes tripunctatus (Christ), Kazakhstan, female 244 metasoma lateral 245 first metasomal tergite dorsal 246 first tergite ventral 247 head and mesosoma dorsal 248 second metasomal tergite dorso-lateral 249 head anterior 250 head and mesosoma lateral 251 detail head and propleuron lateral 252 hind tarsal claw.

Eumenes tripunctatus (Christ), Kazakhstan, male 253 metasoma lateral 254 first metasomal tergite dorsal 255 first tergite ventral 256 head and mesosoma lateral 257 apical hook of antenna lateral 258 head anterior 259 head and propleuron lateral. 260 second metasomal tergite dorso-lateral 261 antenna.

Acknowledgements
We are very grateful for photographs of Fabricius, Kriechbaumer, Blüthgen and Guiglia types kindly supplied by Lars Vilhelmsen and Sree Gayathree Selvantharan (Natural History Museum of Denmark, Copenhagen), Stephan and Olga Schmidt (Zoologische Staatssammlung, München), Michael Greeff (Eidgenössische Technische Hochschule, Zürich) and Roberto Poggi (Museo Civico di Storia Naturale Giacomo Doria, Genoa), respectively.
We thank Frederique Bakker (
References
- Abenius J (2012) Vespidae, Eumeninae - solitäre getingar. In: Douwes P, Abenius J, Cederberg B, Wahlstedt U (Eds) Steklar: Myror-getingar. Hymenoptera: Formicidae – Vespidae. Nationalnyckeln till Sveriges flora och fauna. ArtDatabanken, SLU, Uppsala, 213–275.
- André E[dmond] (1884) Les Guèpes. Species des hyménoptères d’Europe & d’Algérie 2: 404–910.
- Archer ME (2003) The British Potter and Mason Wasps. A handbook. Vespid Studies: 1–96. [BWARS]
- Archer ME (2014) The vespoid wasps (Tiphiidae, Mutillidae, Sapygidae, Scoliidae and Vespidae) of the British Isles. Handbooks for the identification of British Insects 6(6): [i–vi +] 1–82.
- Arens W (2012) Die solitären Faltenwespen der Peloponnes (Hymenoptera: Vespidae: Raphiglossinae, Eumeninae) 1. Teil. Linzer Biologische Beitrage 44(1): 481–522.
- Baldock D, Castro L, Cross I, Schmid-Egger C, Smit J, Wood TJ (2020) The vespiform wasps of Portugal (Hymenoptera: Scolioidea, Tiphioidea, Pompiloidea & Vespoidea). Monografìas Sociedad Entomológiva Aragonesa 34: 1–73.
- Blüthgen P (1938) Beiträge zur Kenntnis der paläarktischen Eumeniden (Hym. Vespidae). Deutsche Entomologische Zeitschrift 1938: 434–496. https://doi.org/10.1002/mmnd.48019380208
- Blüthgen P (1943) Neue paläarktische Eumenes-Arten oder -Rassen (Hym., Vespidae, Eumenidinae). Mitteilungen der Münchener Entomologischen Gesellschaft 38: 297–306.
- Blüthgen P (1952) Weitere neue oder bemerkenswerte paläarktische Faltenwespen aus der Zoologischen Staatssammlung in München (Hym. Eumenidae, Masaridae). Mitteilungen der Münchener Entomologischen Gesellschaft 42: 1–19.
- Blüthgen P (1954) Über Eumenes sardous Guigl. (1951). Bollettino di Societa de Entomologia Italiano 84: 117–199.
- Blüthgen P (1956) Portuguese and Spanish wasps: first supplement (Hymenoptera, Vespoidea). Memórias e estudos do Museo Zoológico da Universidade de Coimbra 240: 1–22.
- Blüthgen P (1959) Über zwei Eumenidae aus Schweden (Hym.). Opuscula Entomologica 24: 13–14.
- Borsato W (2006) Contributo alla conoscenza degli Eumenidae di Sardegna e note sulla fauna corsa (Hymenoptera, Vespoidea). Bollettino del Museo Civico di Storia Naturale di Venezia 57: 123–147.
- Borsato W, Turrisi GF (2004) Contributo alla conoscenza degli Eumenidae di Sicilia (Hymenoptera, Vespoidea). Bollettino del Museo Civico di Storia Naturale di Venezia 55: 127–150.
- Cameron P (1907) On a new genus and some new species of Aculeate Hymenoptera collected by Lieut.-Col. C. G. Nurse in Baluchistan. Journal of the Bombay Natural History Society 18: 130–136.
- Cassar T, Mifsud D, Selis M (2022) The potter wasps of the Maltese Islands (Hymenoptera, Vespidae, Eumeninae). Journal of Hymenoptera Research 90: 201–212. https://doi.org/10.3897/jhr.90.79373
- Castro L (1992) Sobre los euménidos (Hym., Vespoidea) del valle medio del Ebro. Zapateri, Revista aragonesa de entomología 2: 21–34.
- Castro L (1997) Familia Vespidae: Subfamilia Eumeninae. Catalogus de la entomofauna aragonesa 16: 3–8.
- Castro L, Sanza F (2009) Aportación al conocimiento de los Vespidae (Hymenoptera) de Sierra Nevada (España), con algunos comentarios taxonómicos. Boletin de la SEA 45: 259–278.
- Christ JL (1791) Naturgeschichte, Klassification und Nomenclatur der Insekten vom Bienen, Wespen und Ameisengeschlecht: als der fünften Klasse fünfte Ordnung des Linneischen Natursystems von den Insekten, Hymenoptera: mit häutigen Flügeln, p. 1–535. Frankfurt am Main. https://doi.org/10.5962/bhl.title.87724
- Dal Pos D, Carpenter JM, Uliana M (2022) The Giordani Soika collection of Eumeninae at the Natural History Museum of Venice Giancarlo Ligabue: Catalogue of species and type specimens (Insecta, Hymenoptera, Vespidae). Zootaxa 5137(1): 1–111. https://doi.org/10.11646/zootaxa.5137.1.1
- Dalla Torre KW (1894) Catalogus Hymenopterorum hucusque descriptorum systematicus et synonymicus. Vespidae (Diploptera) 9: 1–181. [Leipzig] https://doi.org/10.5962/bhl.title.10348
- Day MC (1979) The species of Hymenoptera described by Linnaeus in the genera Sphex, Chrysis, Vespa, Apis and Mutilla. Biological Journal of the Linnean Society. Linnean Society of London 12(1): 45–84. https://doi.org/10.1111/j.1095-8312.1979.tb00049.x
- de Saussure HF (1852) Etudes sur la famille des vespides 1. Monographie des Guepes Solitaires ou de la tribe des Eumeniens. V: [i–ixlx +] 1–286 [pp. 1–128, pls. 2–5 + 7 + 10 + 14 (1852), second part in 1853].
- de Villers C (1789) Caroli Linnaei entomologia, faunae suecicae descriptionibus aucta 3: 1–657. Piestre & Delamolliere, Lugduni.
- Fabricius JC (1781) Species insectorum, exhibentes eorum differentias specificas, synonyma auctorum, loca natalia, metamorphosin, adjectis observationibus, descriptionibus 1: 1–552. Hamburgi & Kilonii. https://doi.org/10.5962/bhl.title.36509
- Fabricius JC (1804) Systema Piezatorum secundum ordines, genera, species, adjectis synonymis, locis, observationibus, descriptionibus: 1–439. Brunsvigae. https://doi.org/10.5962/bhl.title.10490
- Fateryga AV (2010) Landscape distribution of potter wasps of the subfamily Eumeninae (Hymenoptera: Vespidae) of Crimea. Proceedings of the Russian Entomological Society 81(2): 74–82.
- Fateryga AV (2017) Subfamily Eumeninae – Potter wasps, 178–192. In: Belokobylskij SA, Lelej AS (Eds) Annotated catalogue of the Hymenoptera of Russia. Symphyta and Apocrita: Aculeata 1: 1–476.
- Fateryga AV (2018) Wasps of the family Vespidae (Hymenoptera) of the Crimean Peninsula Entomofauna 39: 193–233.
- Fateryga AV, Matushkina NA (2010) The first nest records of the wasp Eumenes punctaticlypeus kostylevi Kurzenko, 1976 (Hymenoptera, Vespidae, Eumeninae) and notes on its distribution in Crimea. Vestnik Zoologii 44(4): 377–382. https://doi.org/10.2478/v10058-010-0024-8
- Fateryga AV, Proshchalykin MY, Kochetkov DN, Buyanjargal B (2020) New records of eumenine wasps (Hymenoptera, Vespidae, Eumeninae) from Russia, with description of a new species of Stenodynerus de Saussure, 1863. Journal of Hymenoptera Research 79: 89–109. https://doi.org/10.3897/jhr.79.57887
- Fischer-Waldheim G (1843) Observata quaedem de Hymenopteris Rossicis. Magasin de zoologie, d’anatomie compare et de palaeontologie, Ser. 2, 5(1): 1–4[, pl. 122].
- Folmer O, Black M, Hoeh W, Lutz R, Vrijenhoek R (1994) DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Molecular Marine Biology and Biotechnology 3(5): 294–299.
- Frommer U (2012) Mediterrane Stechimmen (Hymenoptera, Aculeata) in Deutschland und angrenzenden Gebieten nach 1990. Eine Übersicht anlässlich des aktuellen Nachweises der mediterranen Töpferwespe Eumenes m. mediterraneus Kriechbaumer, 1879 (Vespidae, Eumeninae). Mitteilungen des Internationalen Entomologischen Vereins 37: 175–197.
- Frommer U (2013) Hinweise zur Trennung der solitären Faltenwespen Eumenes c. coarctatus (Linnaeus, 1758) und Eumenes p. pedunculatus (Panzer, 1799) (Hymenoptera, Vespidae, Eumeninae). Bembix - Zeitschrift für Hymenopterologie 36: 18–21.
- Gereys B (2006) Mise à jour de la nomenclature des Vespidae (Hymenoptera) de France métropolitaine et premier inventaire du département des Alpes-de-Haute-Provence (France) (Deuxième partie). Bulletin de la Société Linnéenne de Lyon 75(10): 383–392. https://doi.org/10.3406/linly.2006.13644
- Gereys B (2011) Eumenes mediterraneus filitosa nov. ssp. de Corse (Hymenoptera Vespidae). Bulletin Mensuel de la Societe Linneenne de Lyon 80(9–10): 221–225. https://doi.org/10.3406/linly.2011.13812
- Gereys B (2012) Notes sur deux Eumeninae d’Afrique du Nord (Hymenoptera Vespidae Eumeninae). Entomologiste 68(4): 199–202.
- Gereys B (2016) Vespidae solitaire de France métropolitaine (Hymenoptera: Eumeninae, Masarinae). Faune de France 98: 1–330.
- Giordani Soika A (1940) Diagnosi di nuovi Eumenes dell’ Asia nord-orientale (Hym. Vespidae) Bollettino della Società veneziana di storia naturale e del Museo civico di storia naturale 2: 96–98.
- Giordani Soika A (1943) Nuovi Vespidi Mediterranei ed Ethiopici appartenenti al genera Eumenes, Leptomenes, Odynerus e Pterochilus. Bollettino della Società veneziana di storia naturale e del Museo civico di storia naturale 3(1): 29–42.
- Giordani Soika A (1952a) Sulle caratteristiche biogeografiche della Palestina, Arabia ed Egitto, con un contributo alla conoscenza degli Zethini ed Eumenini della Palestina. Bollettino della Societa Veneziana di Storia Naturale e del Museo Civico di Storia Naturale 6: 5–62.
- Giordani Soika A (1952b) Risultati della spedizione scientifica zoologica in Turchia del Museo Nazionale di Praha. Acta Entomologica Musei Nationalis Pragae 27(1951): 375–386
- Giordani Soika A (1973) Designazione di lectotipi ed elenco dei tipi di Eumenidi, Vespidi e Masaridi da me descritti negli anni 1934–1960. Bollettino del Museo Civico di Storia Naturale di Venezia 24: 7–53.
- Giordani Soika A (1986) Eumenidi paleartici nuovi o poco noti. Bollettino del Museo Civico di Storia Naturale di Venezia 35(1984): 91–162.
- Giordani Soika A, Borsato W (1995) Hymenoptera Vespoidea. In: Minelli A, Ruffo S, La Posta S (Eds) Checklist delle specie della fauna Italiana 103: 1–9. [Calderini (Bologna)]
- Gogala A (2022) Ose družine Vespidae v Sloveniji (Hymenoptera: Vespoidea: Vespidae) / Vespid wasps of Slovenia (Hymenoptera: Vespoidea: Vespidae). Scopolia 102: 1–79.
- Grandinete YC, Noll FB, Carpenter J (2018) Taxonomic review of Eumenes Latreille 1802 (Hymenoptera, Vespidae, Eumeninae) from the New World. Zootaxa 4459(1): 1–52. https://doi.org/10.11646/zootaxa.4459.1.1
- Guiglia D (1951) Nuovi Eumenes Italiani (Hymen. Eumenidae). Bollettino della Società Entomologica Italiana 81: 27–30.
- Gusenleitner J (1972) Übersicht über die derzeit bekannten westpaläarktischen Arten der Gattung Eumenes Latr. 1802 (Hym. Vespoidea). Bollettino del Museo civico di storia naturale di Venezia XXII–XXIII: 67–117.
- Gusenleitner J (1999) Bestimmungstabellen mittel- und südeuropäischer Eumeniden (Vespoidea, Hymenoptera) Teil 11. Die Gattungen Discoelius Latreille 1809, Eumenes Latreille 1802, Katamenes Meade-Waldo 1910, Delta Saussure 1855, Ischnogasteroides Magretti 1884 und Pareumenes Saussure 1855. Linzer Biologische Beitrage 31(2): 561–584.
- Gusenleitner J (2013) Die Gattungen der Eumeninae im Nahen Osten, in Nordafrika und in Arabien (Hymenoptera: Vespidae: Eumeninae). Linzer Biologische Beitrage 45(1): 5–107.
- Hebert PDN, Cywinska A, Ball SL, deWaard JR (2003) Biological identifications through DNA barcodes. Proceedings of the Royal Society of London: Series B, Biological Sciences 270(1512): 313–321. https://doi.org/10.1098/rspb.2002.2218
- Hebert PDN, Penton EH, Burns JM, Janzen DH, Hallwachs W (2004) Ten species in one: DNA barcoding reveals cryptic species in the neotropical skipper butterfly Astraptes fulgerator. Proceedings of the National Academy of Sciences of the United States of America 101(41): 14812–14817. https://doi.org/10.1073/pnas.0406166101
- Hellén W (1944) Die in Finnland gefundenen Formen der Gattung Eumenes F. Notulae Entomologicae 24: 10–11.
- Hensen R (1985) De plooivleugelwespen. Jeugdbondsuitgeverij, ‘s Graveland, 60 pp.
- Herrich-Schäffer GAW (1841) Favnae insectorvm Germanicae initia, oder, Deutschlands Insecten 44: pl. 179-8.
- Kostylev G (1940) Espèces nonvelles et pen counues de Vespides, Eumenides et de Masarides paléarctiques (Hymenoptera). I. Bulletin de la Société des naturalistes de Moscou, Section biologique (N.S.) 49 (3/4): 137–154.
- Kriechbaumer J (1879) Eumeniden-Studien. Entomologische Nachrichten 5: 57–59, 85–89, 201–204, 309–312.
- Kumar GP, Carpenter JM, Castro L, Sureshan PM (2017) A taxonomic review of the Indian species of the genus Eumenes Latreille (Hymenoptera: Vespidae: Eumeninae). Zootaxa 4317(3): 469–498. https://doi.org/10.11646/zootaxa.4317.3.3
- Kurzenko NV (1976) New species of vespoid solitary wasps (Hymenoptera, Eumenidae) in the fauna of the USSR. Entomologicheskoe Obozrenie 55(2): 434–437. [in Russian]
- Lichtenstein AAH (1796) . Catalogus Museum Holth. 3: 202. [only auction catalogue; newly prosed names probably invalid]
- Linnaeus C (1758) Systema naturae per regna tria naturae: secundum classes, ordines, genera, species, cum characteribus, differentiis, synonymis, locis 1: 1–824. 10th ed., Salvii, Holmiae. https://doi.org/10.5962/bhl.title.542
- Lucciani M (1845) Lettre sur les moeurs des Eumeniens. Annales de la Société entomologique de France. Deuxieme Serie 3: CX–CXII.
- Meade-Waldo G (1910) New species of Diploptera in the collection of the British Museum. Part I. Annals and Magazine of Natural History, 8, 5(25): 30–51. https://doi.org/10.1080/00222931008692723
- Müller M (1923) Unsere märkische Pillenwespe (Eumenes F.). Deutsche Entomologische Zeitschrift 1923: 624–632.
- Neumeyer R (2014) Kommentierte liste der in der Schweiz vorkommenden Faltenwespen (Hymenoptera: Vespidae). Mitteilungen der Schweizerischen entomologischen Gesellschaft 87: 359–369.
- Neumeyer R (2019) Vespidae. Fauna Helvetica 31: 1–381.
- Neumeyer R, Praz C (2015) DNA barcoding of the potter wasp genus Eumenes Latreille, 1802 (Hymenoptera: Vespidae: Eumeninae) in Switzerland. Mitteilungen der Schweizerischen entomologischen Gesellschaft 88: 307–320.
- Panzer GWF (1799) Favnae insectorvm Germanicae initia, oder, Deutschlands Insecten (6) 63: 1–24 & (6) 64: 1–24.
- Reder G (2022) Die Topferwespe Eumenes mediterraneus Kriechbaumer, 1879 in Rheinland-Pfalz (Vespidae, Eumeninae) – ein Hinweis zur Bodenstandigkeit. Ampulex 13: 72–74.
- Richards OW (1980) Scolioidea, Vespoidea and Sphecoidea. Hymenoptera. Aculeata 6(3b): 1–118.
- Sanza F (1997) Revisión de los euménidos de la Península Ibérica (Hymenoptera: Eumenidae) (tesis doctoral, inédita). Unpublished thesis, Dept de Biología Animal, Ecología, Parasitología, Edafología y Química Agrícola de la Universidad de Salamanca, Salamanca, i–vii + 1–541.
- Schmid-Egger C (2004) Bestimmungsschlüssel für die deutschen Arten der solitären Faltenwespen (Hymenoptera: Eumeninae). Deutscher Jugendbund für Naturbeobachtung Hamburg, 54–102. 3. überarbeitete Auflage. Hamburg.
- Schmid-Egger C (2010) Rote Liste der Wespen Deutschlands. Hymenoptera Aculeata: Grabwespen (Ampulicidae, Crabronidae, Sphecidae), Wegwespen (Pompilidae), Goldwespen (Chrysididae), Faltenwespen (Vespidae), Spinnenameisen (Mutillidae), Dolchwespen (Scoliidae), Rollwespen (Tiphiidae) und Keulhornwespen (Sapygidae). Ampulex 1: 5–40.
- Schmid-Egger C (2011) Hymenoptera Aculeata from “Parc national du Mercantour” (France) and “Parco delle Alpi Marittime” (Italy) in the south-western Alps. Ampulex 3: 13–50. [Berlin.]
- Schmid-Egger C, Schmidt K (2002) Kritisches Verzeichnis der deutschen Arten der Faltenwespen (Hymenoptera, Vespidae). Bembix 16: 13–28.
- Schmid-Egger C, Schmidt S (2021) Unexpected diversity in Central European Vespoidea (Hymenoptera, Mutillidae, Myrmosidae, Sapygidae, Scoliidae, Tiphiidae, Thynnidae, Vespidae), with description of two species of Smicromyrme Thomson, 1870. ZooKeys 1062: 49–72. https://doi.org/10.3897/zookeys.1062.70763
- Smit J, Peeters TMJ, Lefeber V (2004) Vespidae – plooivleugelwespen, 321–344. In: Peeters TMJ, van Achterberg C, Heitmans WRB, Klein WF, Lefeber V, van Loon AJ, Mabelis AA, Nieuwenhuijsen H, Reemer M, de Rond J, Smit J, Velthuis HHW (Eds) De wespen en mieren van Nederland (Hymenoptera: Aculeata). Nederlandse Fauna 6: 1–507.
- Szulczewski JW (1950) [Hymenoptera of the National Park of Great Poland] IV Aculeata. Pr. Monografia Przyrody Wielkopolski Parku narodowy. Posnan 2(4): 1–20.
- Tobias VI, Kurzenko NV (1978) Sem. 2. Eumenidae, 152–173. In: Medvedev G (Ed.) Keys to the identification of insects of European USSR. Hymenoptera 3(1), 1–584. [In Russian]
- van der Vecht J (1968) Notes on some European Eumenes-species Hymenoptera, Eumenidae). Entomologische Berichten (Amsterdam) 28: 68–80.
- van der Vecht J, Fischer FCJ (1972) Palaearctic Eumenidae. Hymenopterorum catalogus nova editio 8: 1–200.
- Vergés Serra F (1985) Estudio monográfico de los Eumenes de España (Hymenoptera, Eumenidae). Graellsia 41: 137–152.
- Woydak H-G (2006) Hymenoptera Aculeata Westfalica. Die Faltenwespen von Nordrhein-Westfalen (Hymenoptera, Vespoidea; Vespidae und Eumenidae) (Soziale Papier und Lehmwespen). Abhandlungen aus dem Westfalischen Museum fur Naturkunde 68(1): 1–133.
- Yamane Sk (1977) Notes on eumenid wasps from Japan and adjacent regions (Hymenoptera: Vespidae) 2. New Entomologist 26: 59–64. [In Japanese]
- Yildirim E, Özbek H (1996) Eumenidae (Hymenoptera, Vespoidea) of Turkey. Turkish Journal of Zoology 20(2): 189–209. https://doi.org/10.55730/1300-0179.3031