Research Article |
|
Corresponding author: Yuehua Song ( songyuehua@163.com ) Academic editor: Mick Webb
© 2022 Jia Jiang, Christopher H. Dietrich, Can Li, Yuehua Song.
This is an open access article distributed under the terms of the Creative Commons Attribution License (CC BY 4.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Citation:
Jiang J, Dietrich CH, Li C, Song Y (2022) Comparative morphology of part of the integumental fine structure of two Erythroneurine species: Singapora shinshana (Matsumura, 1932) and Empoascanara sipra Dworakowska, 1980 (Hemiptera, Cicadellidae, Typhlocybinae). ZooKeys 1103: 1-23. https://doi.org/10.3897/zookeys.1103.80787
|
Abstract
This study describes the fine structure of the mouthparts, antennae, forewings, and brochosomes of two leafhopper species belonging to the typhlocybine tribe Erythroneurini collected from the Karst area of Guizhou Province, southern China: Singapora shinshana, which prefers woody dicot hosts, and Empoascanara sipra, which feeds on grasses. As in other leafhoppers, the piercing-sucking mouthparts consist of a conical labrum, a cylindrical three-segmented labium, and a slender stylet fascicle. The labrum of both species has no sensilla and the labium has several common types of sensilla, but the two species differ in the numbers, types, and distribution of sensilla and in other aspects of the surface sculpture of the mouthparts. The stylet fascicle has distinctive dentition on both the maxillary and mandibular stylets. The antennae of the two species differ in several respects, including the sensilla and sculpture of the scape, pedicel, and flagellum, as well as the degree of sub-segmentation of the flagellum. Except for the variable scaly structure and rounded protrusions on the surface of S. shinshana, the fine structure of the forewing surfaces of the two species are similar to those of other leafhoppers. Only small spherical brochosomes were found on the body surface of S. shinshana and E. sipra. Similar studies of additional erythroneurine species are needed to determine whether differences in mouthpart and antennal fine structure may reflect adaptation to different host plant.
Keywords
Antennae, brochosomes, fine structure, forewings, karst, mouthparts
Introduction
Leafhoppers, the Cicadellidae, are the largest family of Hemiptera and are widely distributed in six zoogeographic regions with more than 2,600 genera and 22,000 species (
Over the course of their more than 400 million years of evolution, different insects have acquired a wide variety of integumental structures, including sensilla and sculpturing that enabled them to interact and adapt to various environmental conditions. Such structures play important roles in finding hosts, mating, and defense. Using light microscopy and scanning electron microscopy,
To date, the fine structure of the integument of Typhlocybinae remains largely unstudied.
Materials and methods
The adult specimens of S. shinshana were collected on a peach tree on the Baoshan Campus of Guizhou Normal University, Guiyang City, Guizhou Province, China (26°35'30"N, 106°43'9"E) on 21 June 2020. The temperature at the time of collection was 27 °C, and the humidity was 91%. The adult specimens of E. sipra Dworakowska, 1980 were collected on Festuca elata Keng ex E. Alexeev, 1977 in Changpoling Forest Park, Guiyang City, Guizhou Province, China (26°38'45"N, 106°39'10"E) on 27 June 2020. The temperature was 20 °C and the humidity was 99% during collection. The overall appearance of the two leafhopper species is shown in Fig.

Newly captured adult specimens were placed in a -24 °C freezer for 20 min. Then ten frozen specimens (5 males and 5 females) were selected at random and dissected under a stereo microscope (Olympus SZX16, Japan), with the head and wings removed on dry filter paper, then placed in 2.5% glutaraldehyde fixative at 4 °C for 12 hours. Specimens were subsequently transferred to phosphate buffer saline (PBS, 0.1M, pH7.2) and rinsed five times, 5 min each time. Dissected parts (except wings) were then placed in an ultrasonic cleaner for 30 s, and then dehydrated in a graded series of 30%, 50%, 70%, 90%, 95%, and 100% acetonitrile for 20 min. Thereafter, the samples were mounted on aluminum stubs with double-sided sticky copper tape and sputtered with gold/palladium in a JEOL JFC-1600 high resolution sputter coater. The samples were subsequently examined with a JSM-6490LV SEM operated at 20 kV. The measurement data were obtained by scanning electron microscope.
General terminology for the classification of sensilla follows
| Type | Features | Reference images | |
| Sensilla trichodea | S.t. I | Hair-like, slender, slightly curved, length ≥ 20 μm. |

|
| S.t. II | Relatively short. | ||
| S.t. III | Short and thin, length ≤ 10 μm. | ||
| Sensilla chaetica | S.c. | Shaped like short spines, erect or curved along the axis. | |
| sensilla basiconica | S.b. I | Upright or curved along the axis, the top is blunt, thick and short, length ≤ 10 μm. | |
| S.b. II | Relatively thick and long. | ||
| S.b. III | Thick and long, length ≥ 20 μm. | ||
| Peg sensilla | Pg.s.u. I | Peg-like, length 2.0~5.0 μm. |
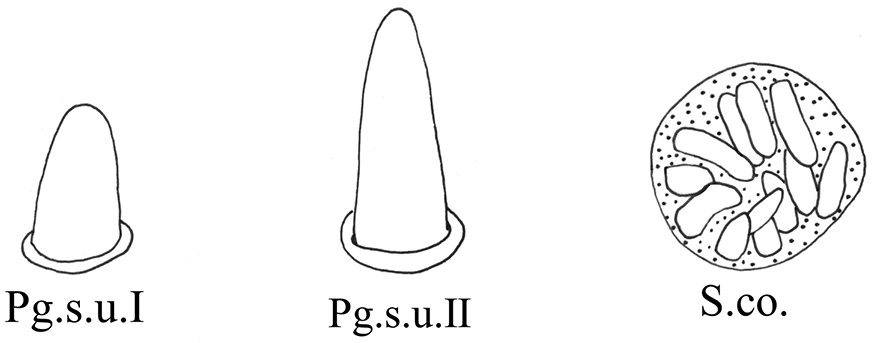
|
| Pg.s.u. II | Peg-like, length 5.0~7.0 μm. | ||
| Sensilla coeloconica | S.co. | A cluster of finger-like structures arranged in a round concavity, 6–16 finger-like protrusions. | |
| Scaly structures | Sc.s. | A scaly protrusion or a scaly structure composed of many small protrusions (non-sensilla). |

|
| Cuticular processes | C.p. | Triangular protrusions with thin and pointed ends (non-sensilla). | |
| Microtrichia | Mt. | Small rigid projections occurring singly or in groups of two or three arranged together (non-sensilla). | |
Results
The mouthparts of S. shinshana and E. sipra are typical piercing-sucking mouthparts, consisting of a labrum (Lm), labium (Lb), two mandibular stylets (Md), and two maxillary stylets (Mx) comprising the stylet fascicle (Sf) (Figs
Measurements of labrum and labium (mean ± SE) obtained from scanning electron microscopy, n = 5. Lm: labrum; Lb: labium; Lb-1: first segment of labium; Lb-2: second segment of labium; Lb-3: third segment of labium.
| Segment | Lm | Lb-1 | Lb-2 | Lb-3 | Lb total length | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| S. shinshana | E. sipra | S. shinshana | E. sipra | S. shinshana | E. sipra | S. shinshana | E. sipra | S. shinshana | E. sipra | ||
| Length (μm) | Male | 62.7±12.0 | 52.37±3.2 | 81.3±8.7 | 73.1±7.9 | 90.8±10.3 | 73.7±5.6 | 108.3±5.4 | 96.0±15.1 | 280.4±24.4 | 242.8±28.6 |
| Female | 72.7±9.8 | 69.4±10.1 | 96.1±16.1 | 78.6±5.2 | 99.7±9.2 | 84.6±3.5 | 122.1±6.4 | 114.4±7.9 | 317.9±31.7 | 275.6±16.6 | |

SEM of the mouthparts of S. shinshana A anterior view, showing labrum (Lm), mandibular stylets (Md), maxillary stylets (Mx), sensilla trichodea I (S.t. I) and sensilla trichodea II (S.t. II) B anterior view of anteclypeus and labrum (Lm), showing irregular protrusions on surface of anteclypeus, labrum, mandibular stylets (Md) and sensilla trichodea II (S.t. II) C cone-shaped labrum showing a smooth surface.

SEM of the labium of S. shinshana A anterior view of labium showing three-segmented labium (I-III), and sensilla symmetrically located on each side of the labial groove B anterior view of first segment of labium showing sensilla basiconica II (S.b.II) and cuticular processes (C.p) C sensilla coeloconica (S.co.) D the anterior view of second segment of labium showing sensilla trichodea I (S.t.I) and microtrichia (Mt.) E anterior view of third segment of labium showing sensilla trichodea I (S.t.I), sensilla basiconica II (S.b.II), sensilla chaetica (S.c.) and microtrichia (Mt.) F anterior view of labial tip showing peg sensilla I (Pg.s.I) and sensilla trichodea III (S.t.III) G dorsal view of mouthparts showing three-segmented labium (I-III) and some sensilla H dorsal view of second segment of labium showing sensilla trichodea II (S.t.II) and sensilla basiconica II (S.b.II) I junction of second and third labial segments showing spherical protrusions J dorsal view of third segment of labium showing sensilla trichodea I (S.t.I) K tip of labium, showing distribution of sensilla.
The labrum is conical in shape and connected to the apical margin of the anteclypeus. The anteclypeus has many irregular protrusions on its surface, with some sensilla trichodea I and sensilla trichodea II symmetrically distributed on its surface (Figs
The labium consists of three cylindrical segments (Figs
A few microtrichia are concentrated on oblique ridges near the median longitudinal groove on the second labial segment of S. shinshana, while a larger number of microtrichia are distributed on the second and third labial segments of E. sipra (Figs
The third labial segment is longer than other two segments, gradually tapered towards the apex, and more densely covered with sensilla, mostly symmetrically distributed. Sensilla trichodea I–III, peg sensilla (S. shinshana: peg sensilla I, ~ 3.32 μm in length; E. sipra: peg sensilla II, ~ 5.57 μm in length) and sensilla basiconica (S. shinshana: sensilla basiconica II, ~ 18.05 μm in length; E. sipra: sensilla basiconica I, ~ 9.58 μm in length) are distributed on the third labial segment of the two species; and there is a pair of peg sensilla distributed on both sides of the longitudinal groove. The labial tip surface is uneven, with many small, rounded protrusions (Figs
The stylet fascicle is composed of paired, elongated mandibular and interlocking maxillary stylets. The mandibular stylets partially sheathe the maxillary stylets laterally and are significantly shorter than the latter. They are crescent-shaped in cross-section, thus forming a deep groove enclosing the maxillary stylets. Each mandibular stylet has a row of slender tooth-like protrusions on its inner edge in the basal half, and the protrusions together form a zigzag structure (Figs

SEM of the stylet fascicle of S. shinshana A mandibular stylets (Md), showing relative position of mandibular stylets and labrum (Lm) B mandibular stylet (Md), showing serrate ridge on the convex external surface and zigzag structure on inner edge C enlarged middle of mandibular stylet (Md), showing zigzag structure on inner edge D maxillary stylets E dorsal view of middle section of maxillary stylets (Mx), showing lines indicating food canal (Fc) F lateral view of middle section of maxillary stylets (Mx), showing relatively blunt tooth-like protrusion G tip of maxillary stylet (Mx), showing salivary canal (Sc) and food canal (Fc) H tip of maxillary stylets (Mx), showing two stylets with different lengths.

SEM of the mouthparts of E. sipra A the anterior view of labrum and labium showing sensilla symmetrically located on each side of the labial groove or around the tip of the labium B one of the maxillary stylets (Mx) showing food canal (Fc) and salivary canal (Sc) C the enlarged view of the tip of maxillary stylets (Mx) which are pointed and incurred D cone-shaped labrum showing a smooth surface E mandibular stylet (Md), showing serrate ridge on the convex external surface and zigzag structure on inner edge F mandibular stylet (Md), showing the depression on the side of the mandibular stylet G dorsal view of first segment of labium showing a smooth surface H dorsal view of second segment of labium showing sensilla trichodea I (S.t.I) and sensilla trichodea II (S.t.II) I dorsal view of third segment of labium showing sensilla trichodea I (S.t.I), sensilla trichodea II (S.t.II), sensilla trichodea III (S.t.III) J tip of labium, showing sensilla basiconica I (S.b.I) and peg sensilla II (Pg.s.II) K anterior view of first segment of labium showing sensilla basiconica, II (S.b.II) and cuticular processes (C.p) L anterior view of second segment of labium showing sensilla trichodea I (S.t.I), sensilla trichodea II (S.t.II) and microtrichia (Mt.) M anterior view of third segment of labium showing sensilla trichodea II (S.t.II), sensilla chaetica (S.c.) and peg sensilla II (Pg.s.II).
The two maxillary stylets are semicircular in cross-section and tightly interlocked to form a salivary canal (Sc) and a food canal (Fc) (Figs
The antennae of the two studied species are of the typical arisoid type present in other Cicadellidae, composed of three parts: scape (Sc), pedicel (Pe) and flagellum (Fl) (Figs
Measurements of antennae (mean ± SE) obtained from scanning electron microscopy, n = 5. Sc: scape; Pe: pedicel; Fl: flagellum.
| Segment | Sc | Pe | Fl | total length | |||||
|---|---|---|---|---|---|---|---|---|---|
| S. shinshana | E. sipra | S. shinshana | E. sipra | S. shinshana | E. sipra | S. shinshana | E. sipra | ||
| Length (μm) | Male | 58.7±3.9 | 52.7±9.6 | 78.3±6.8 | 72.9±6.1 | 518.5±14.1 | 496.9±13.5 | 655.5±25.9 | 622.5±29.2 |
| Female | 59.1±2.8 | 56.4±8.1 | 80.6±9.2 | 75.6±10.4 | 548.7±25.1 | 513.3±12.3 | 688.4±37.1 | 645.3±30.8 | |
A statistical table of the sensilla and cuticular processes of the labium, antennae, and forewings. Lb-1: first segment of labium; Lb-1: second segment of labium; Lb-1: third segment of labium; Sc: scape; Pe: pedicel; Fl: flagellum; Fw: forewing. Note: The number of sensilla or cuticular processes is the average for the number of samples (n = 10); no entry indicates that the number of some sensors was not counted.
| Sensilla type | Distribution (number) | |
|---|---|---|
| S. shinshana | E. sipra | |
| S.t. I | Lb-2(12); Lb-3 | Lb-2(10); Lb-3(2) |
| S.t. II | Lb-2(4); Lb-3(4) | Lb-1(2); Lb-2(10); Lb-3 |
| S.t. III | Lb-3(2); Pe(4) | Lb-3; Sc(1); Pe(2) |
| S.c. | Lb-3(2); Sc(2); Fw | Sc(1); Lb-3(2); Fw |
| S.b. I | Lb-1(2); Lb-3(2) | |
| S.b. II | Lb-1(1); Lb-3(2) | Fl(1) |
| S.b. III | Fl(1) | |
| Pg.s.u. I | Lb-3(2) | |
| Pg.s.u. II | Lb-3(2) | |
| S.co. | Lb-1(2) | |
| Sc.s. | Pe | Sc; Pe |
| C.p. | Lb-1 | |
| Mt. | Lb-2; Lb-3; Fl; Fw | Lb-1; Lb-2; Lb-3; Pe; Fl; Fw |

SEM of the antennae of S. shinshana A antenna, composed of three parts: scape (Sc), pedicel (Pe) and three regions of flagellum (Fl) B scape, showing smooth surface, with two sensilla chaetica (S.c.) C pedicel, showing scale-like structures (Sc.s.) and sensilla trichodea III (S.t.III) D enlarged view of pedicel, showing scaly structures and sensilla trichodea III (S.t.III) E junction between pedicel and flagellum, showing microtrichia (Mt.) F first region of flagellum, showing sensilla basiconica III (S.b.III) G second region of the flagellum H junction between second and third regions of flagellum, showing change in surface protrusions I junction between first part and second regions of flagellum, showing microtrichia (Mt.) J enlarged view of second part of flagellum, showing cylindrical subsegments K enlarged view of third part of flagellum, showing brochosomes (BS).

SEM of the antennae of E. sipra A antenna, composed of three parts: scape (Sc), pedicel (Pe) and three regions of flagellum (Fl) B scape and pedicel, showing scale-like structures (Sc.s.), sensilla trichodea III (S.t.III), sensilla chaetica (S.c.), microtrichia (Mt.) C first region of flagellum, showing sensilla basiconica II (S.b.II) D junction between first and second regions of flagellum, showing change in surface protrusions and microtrichia (Mt.) E second region of the flagellum, showing spherical protrusions on the surface.
The scape is short, thick, approximately bell-shaped, with the base consisting of a flexible antennal membrane (Figs
The pedicel is connected to the recessed socket at the end of the scape (Figs
The flagellum is elongated and divided into numerous subsegments (Figs
As in other Typhlocybinae, the costal area has an elongated oval white area often referred to in previous literature as the “brochosomal area” or “wax field”, but actually consisting of a patch of brochosomes. There are numerous microtrichia and small sensilla chaetica scattered on upper forewing surface of the two erythroneurine species (Figs

SEM of the brochosomes of S. shinshana A peculiar fine structure of brochosomal area B microtrichia (Mt.) on transparent membrane of brochosomal area C sensilla chaetica (S.c.) on front edge of forewing D posterior edge of forewing, showing surface folds, scaly structure (Sc.s.) and sensilla chaetica (S.c.) E posterior edge of forewing, showing sensilla chaetica (S.c.) and microtrichia (Mt.) F brochosomes (BS) on front edge of forewing G enlarged view of brochosomes (BS) on front edge of forewing.

Small spherical brochosomes (the white powder on the forewings) were found on body surfaces of both male and female adults, with diameters of 402.00–583.10 nm (Figs
Discussion
Despite belonging to a single leafhopper tribe, the two studied species of Erythroneurini show remarkable differences in the fine structure of their mouthparts and antennae. The mouthparts of S. shinshana and E. sipra are generally similar to those of other Hemiptera in gross morphology (
The number of labium segments of Hemiptera insects varies between 1–5, but most species have 3 or 4 (
The stylet fascicle is the main tool used for feeding, and it is also an important medium for spreading plant pathogens. Singapora shinshana and E. sipra have a ridge at the apex of the feeding stylet with a serrated structure in the middle. The ridges are not connected to the serrated structure, and their shape is very similar to that of A. triangularis (
Insect antennae are variously used in insect communication, foraging for food and courtship. Leafhopper antennae are relatively simple in structure and have relatively few sensory structures compared to those of some other Auchenorrhyncha (particularly Fulgoroidea); thus they have been little studies from a comparative perspective. The antennae of S. shinshana and E. sipra generally resemble those of other leafhoppers (
The cylindrical pedicel is slightly longer than the scape. Singapora shinshana and E. sipra have scaly structures of different sizes scattered on the surface of the pedicel, but the cuticular processes that make up the scaly structure are different. The cuticular processes of S. shinshana are obviously wider than those of E. sipra. The scaly structure of E. sipra composed of micro-thorn-like cuticular processes is different from that of other leafhoppers (
The flagellum is the longest segment and has a large number of microtrichia at the end of each basal subsegment. Both S. shinshana and E. sipra have only one very long sensilla basiconica that appears on the 5th subsegment of the flagellum. Previously studied leafhopper species, such as Scaphoideus titanus Ball, 1932, Empoasca onukii Matsuda, 1952, and Chlorotettix nigromaculatus (Dai, Chen & Li, 2006), have a longer sensillum between the 3rd and 6th subsegments of the flagellum (
Brochosomes are minute protein-lipid particles with a net-like surface produced intracellularly in specialized glandular segments of the Malpighian tubules of leafhoppers. Their protein content ranges from 45–70% (
After leafhoppers molt, brochosomes are secreted and anointed onto the body surface. Leafhopper species may differ in the amount of brochosomes secreted and in the time spent anointing. Singapora shinshana secretes 19 drops during each anointing episode on average, and the anointing behavior takes 2–4 h (
Brochosomes form a hydrophobic coating of the integument that can protect leafhoppers from wetting in areas of high humidity or rainfall. The brochosome coating may also provide some protection against high temperature and solar radiation, may help prevent evaporation of body surface water, and may also help leafhoppers avoid natural enemies, diseases, and parasites (
Conclusions
SEM comparisons of the integumental fine structure of two species of erythroneurine leafhoppers representing two different genera show that, although the overall structure of the mouthparts, antennae, and forewings are highly similar, many details differ between these species in integumental sculpturing, and the numbers, types, and distribution of sensilla. Singapora shinshana feeds on the leaves of peach and related Rosaceous trees while E. sipra and other species of Empoascanara feed on grasses. Thus, some of the observed differences may reflect adaptation to the very different chemical composition and structure of the host plants of these species. Further studies of other species in this tribe are needed to determine whether particular aspects of the mouthpart and antennal structures may be more broadly correlated to particular feeding preferences.
Acknowledgements
This study was partly funded by the World Top Discipline Program of Guizhou Province: Karst Ecoenvironment Sciences (No.125 2019 Qianjiao Keyan Fa), the Guizhou Provincial Science and Technology Foundation ([2018]1411), the Innovation Group Project of Education Department of Guizhou Province ([2021]013), the Guizhou Science and Technology Support Project ([2019]2855), the Science and Technology Project of Guiyang City ([2020]7-18), the Training Program for High-level Innovative Talents of Guizhou Province ([2016]4020) and the Project for Regional Top Discipline Construction of Guizhou Province: Ecology in Guiyang University [Qian Jiao Keyan Fa [2017]85].
References
- Aljunid SF, Anderson M (1983) Ultrastructure of sensilla on the antennal pedicel of the brown planthopper Nilaparvata lugens Stal (Insecta: Homoptera). Cell and Tissue Research 228(2): 313–322. https://doi.org/10.1007/BF00204881
- Altner H, Prillinger L (1980) Ultrastructure of invertebrate chemo-, thermo-, and hygroreceptors and its functional significance. International Review of Cytology 67: 69–139. https://doi.org/10.1016/S0074-7696(08)62427-4
- Anderson WG, Heng-Moss T, Baxendale FP, Baird LM, Sarath G, Higley L (2006) Chinch Bug (Hemiptera: Blissidae) mouthpart morphology, probing frequencies, and locations on resistant and susceptible germplasm. Journal of Economic Entomology 99(1): 212–221. https://doi.org/10.1093/jee/99.1.212
- Backus EA, McLean DL (1982) The sensory systems and feeding behavior of leafhoppers. I. The aster leafhopper, Macrosteles fascifrons Stål (Homoptera, Cicadellidae). Journal of Morphology 172(3): 361–379. https://doi.org/10.1002/jmor.1051720310
- Boyd DW (2003) Digestive enzymes and stylet morphology of Deraeocoris nigritulus (Uhler) (Hemiptera: Miridae) reflect adaptations for predatory habits. Annals of the Entomological Society of America 96(5): 667–671. https://doi.org/10.1603/0013-8746(2003)096[0667:DEASMO]2.0.CO;2
- Brozek J, Bourgoin T (2013) Morphology and distribution of the external labial sensilla in Fulgoromorpha (Insecta: Hemiptera). Zoomorphology 135(1): 33–65. https://doi.org/10.1007/s00435-012-0174-z
- Brozek J, Bourgoin T, Szwedo J (2006) The interlocking mechanism of maxillae and mandibles in Fulgoroidea (Insecta: Hemiptera: Fulgoromorpha). Polskie Pismo Entomologiczne 75: 239–253.
- Dai W, Pan LX, Lu YP, Jin L, Zhang CN (2014) External morphology of the mouthparts of the white-backed planthopper Sogatella furcifera (Hemiptera: Delphacidae), with special reference to the sensilla. Micron (Oxford, England) 56: 8–16. https://doi.org/10.1016/j.micron.2013.09.005
- Davidson J (1914) On the mouthparts and mechanism of suction in Schizoneuru lanigera Hausm. Journal of the Linnean Society of London 32(218): 307–330. https://doi.org/10.1111/j.1096-3642.1914.tb01460.x
- Dietrich CH (2005) Keys to the families of Cicadomorpha and subfamilies and tribes of Cicadellidae (Hemiptera: Auchenorrhyncha). The Florida Entomologist 88(4): 502–517. https://doi.org/10.1653/0015-4040(2005)88[502:KTTFOC]2.0.CO;2
- Dong HY, Huang M (2013) Analysis of the anointing and grooming behavior of several adult insects in Typhlocybinae (Hemiptera: Cicadellidae). Journal of Insect Behavior 26(4): 540–549. https://doi.org/10.1007/s10905-012-9370-4
- Emeljanov AF (1987) Phylogeny of Cicadina (Homoptera, Cicadina) according to data on comparative morphology. Trudy Vsesoiuznogo Entomologicheskogo Obschestva 69: 19–109.
- Forbes AR (1977) The mouthparts and feeding mechanism of aphids. Aphids As Virus Vectors 35(138): 83–103. https://doi.org/10.1016/B978-0-12-327550-9.50008-2
- Ge FR, Dietrich C, Dai W (2016) Mouthpart structure in the woolly apple aphid Eriosoma lanigerum (Hausmann) (Hemiptera: Aphidoidea: Pemphigidae). Arthropod Structure & Development 45(3): 230–241. https://doi.org/10.1016/j.asd.2016.01.005
- Guo FZ, Yuan SX, Wang HR, Guo KJ (2018) Observation of the antenna and antennal sensilla of adults of the leafhopper Chlorotettix nigromaculatus (Hemiptera: Cicadellidae) with scanning electron microscope. Acta Entomologica Sinica 61(10): 1192–1201. https://doi.org/10.16380/j.kcxb.2018.10.009
- Hao Y, Dietrich CH, Dai W (2016a) Development of mouthparts in the cicada Meimuna mongolica (Distant): Successive morphological patterning and sensilla differentiation from nymph to adult. Scientific Reports 6(1): e38151. https://doi.org/10.1038/srep38151
- Hao YN, Dietrich CH, Dai W (2016b) Structure and sensilla of the mouthparts of the spotted Lanternfly Lycorma delicatula (Hemiptera: Fulgoromorpha: Fulgoridae), a polyphagous invasive planthopper. PLoS ONE 11(6): e0156640. https://doi.org/10.1371/journal.pone.0156640
- Hirao J, Inoue H (1979) Transmission characteristics of rice waika virus by the green rice leafhopper, Nephotettix cincticeps Uhler (Hemiptera: Cicadelidae). Applied Entomology and Zoology 14(1): 44–50. https://doi.org/10.1303/aez.14.44
- Humphrey EC, Dworakowska I (2002) The natural history of brochosomes in Yakuza gaunga (Hemiptera, Auchenorrhyncha, Cicadellidae, Typhlocybinae, Erythroneurini). Denisia 4: 433–454.
- Hunt RE, Nault LR (1990) Influence of life history of grasses and maize chlorotic dwarf virus on the biotic potential of the leafhopper Graminella nigrifrons (Homoptera: Cicadellidae). Environmental Entomology 19(1): 76–84. https://doi.org/10.1093/ee/19.1.76
- Leopold RA, Freeman TP, Buckner JS, Dennis RN (2003) Mouthpart morphology and stylet penetration of host plants by the glassy-winged sharpshooter, Homalodisca coagulata (Homoptera: Cicadellidae). Arthropod Structure & Development 32(2–3): 189–199. https://doi.org/10.1016/S1467-8039(03)00047-1
- Liang AP, Fletcher MJ (2002) Morphology of the antennal sensilla in four Australian spittlebug species (Hemiptera: Cercopidae) with implications for phylogeny. Australian Journal of Entomology 41(1): 39–44. https://doi.org/10.1046/j.1440-6055.2002.00266.x
- Liu W, Zhang Y, Dietrich CH, Duan Y (2020) Comparative analysis of antennal fine structure of Goniagnathus punctifer, Stirellus yeongnamensis and Stirellus indrus (Hemiptera: Cicadellidae: Deltocephalinae). Zoomorphology 139(4): 461–469. https://doi.org/10.1007/s00435-020-00501-5
- Matsumura S (1932) A revision of the Palaearctic and Oriental Typhlocybid genera with descriptions of new species and new genera. Insecta Matsumurana 6(3): 93–120.
- Mazzoni V, Ioriatti C, Trona F, Lucchi A, De Cristofaro A, Anfora G (2009) Study on the role of olfaction in host plant detection of Scaphoideus titanus (Hemiptera: Cicadellidae) nymphs. Journal of Economic Entomology 102(3): 974–980. https://doi.org/10.1603/029.102.0316
- Moulins M (1971) Ultrastructure et physiologie des organes épipharyngiens et hypopharyngiens (chimiorécepteurs cibariaux) de Blabera craniifer Burm (Insecte, Dictyoptère). Zeitschrift fur Vergleichende Physiologie 73(2): 139–166. https://doi.org/10.1007/BF00304130
- Oman PW, Knight WJ, Nielson MW (1990) Leafhoppers (Cicadellidae): A bibliography, generic check-list and index to the world literature 1956–1985. Annals of the Entomological Society of America 51(5): 53–89. https://doi.org/10.1016/0022-0965(91)90077-6
- Pan LX (2013) Comparative morphology of the mouthparts in Auchenorrhyncha (Insecta: Hemiptera). Northwest A & F University, 26–37.
- Pointeau S, Ameline A, Laurans F, Sallé A, Rahbé Y, Bankhead-Dronnet S, Lieutier F (2012) Exceptional plant penetration and feeding upon cortical parenchyma cells by the woolly poplar aphid. Journal of Insect Physiology 58(6): 857–866. https://doi.org/10.1016/j.jinsphys.2012.03.008
- Qiao L, Zhang L, Qin DZ, Li BL, Lu ZC, Li HL, Xia MC (2016) Ultramicromorphology of antennal sensilla and brochosome of adult Empoasca onukii. Xibei Nongye Xuebao 25(03): 471–476.
- Rakitov RA (1999) Secretory products of the Malpighian tubules of Cicadellidae (Hemiptera, Membracoidea): An ultrastructural study. International Journal of Insect Morphology & Embryology 28(3): 179–193. https://doi.org/10.1016/S0020-7322(99)00023-9
- Rakitov RA (2000) Secretion of brochosomes during the ontogenesis of a leafhopper, Oncometopia orbona (F.) (Insecta, Homoptera, Cicadellidae). Tissue & Cell 32(1): 28–39. https://doi.org/10.1054/tice.1999.0084
- Rakitov RA (2004) Powdering of egg nests with brochosomes and related sexual dimorphism in leafhoppers (Insecta, Hemiptera, Cicadellidae). Zoological Journal of the Linnean Society 140: 353–381. https://doi.org/10.1111/j.1096-3642.2003.00103.x
- Rakitov RA (2009) Brochosomal coatings of the integument of leafhoppers (Hemiptera, Cicadellidae). Functional Surfaces in Biology 1: 113–137. https://doi.org/10.1007/978-1-4020-6697-9_8
- Rakitov RA, Carolina G (2005) New egg-powdering sharpshooters (Hemiptera: Cicadellidae: Proconiini) from Costa Rica. Annals of the Entomological Society of America 98(4): 444–457. https://doi.org/10.1603/0013-8746(2005)098[0444:NESHCP]2.0.CO;2
- Rakitov R, Moysa AA, Kopylov AT, Moshkovskii SA, Peters RS, Meusemann K, Misof B, Dietrich CH, Johnson CH, Podsiadlowski L, Walden KKO (2018) Brochosomins and other novel proteins from brochosomes of leafhoppers (Insecta, Hemiptera, Cicadellidae). Insect Biochemistry and Molecular Biology 94: 10–17. https://doi.org/10.1016/j.ibmb.2018.01.001
- Razaq A, Kashiwazaki T, Mohammad P, Shiraishi M (2000) SEM observations on the citrus green aphid, Aphis citricola van der Goot (Homoptera: Aphididae). Pakistan Journal of Biological Sciences 3(6): 949–952. https://doi.org/10.3923/pjbs.2000.949.952
- Rice MJ (1973) Cibarial sense organs of the blowfly, Calliphora erythrocephala (Meigen) (Diptera: Calliphoridae). International Journal of Insect Morphology & Embryology 2(2): 109–116. https://doi.org/10.1016/0020-7322(73)90012-3
- Romani R, Rossi Stacconi MV, Riolo P, Isidoro N (2009) The sensory structures of the antennal flagellum in Hyalesthes obsoletus (Hemiptera: Fulgoromorpha: Ciixidae): A functional reduction? Arthropod Structure & Development 38(6): 473–483. https://doi.org/10.1016/j.asd.2009.08.002
- Silva MBA, Barbosa HS, Jurberg J, Galvão C, Carcavallo RU (2002) Comparative ultrastructural analysis of the antennae of Triatoma guazu and Triatoma jurbergi (Hemiptera: Reduviidae) during the nymphal stage development. Journal of Medical Entomology 39(5): 705–715. https://doi.org/10.1603/0022-2585-39.5.705
- Stacconi MVR, Romani R (2012) Antennal sensory structures in Scaphoideus titanus ball (Hemiptera: Cicadellidae). Microscopy Research and Technique 75(4): 458–466. https://doi.org/10.1002/jemt.21078
- Tavella L, Arzone A (1993) Comparative morphology of mouth parts of Zyginidia pullula, Empoasca vitis, and Graphocephala fennahi (Homoptera, Auchenorrhyncha). Bollettino di Zoologia 60(1): 33–39. https://doi.org/10.1080/11250009309355788
- Wang X, Li QL, Wei C (2018) Comparative morphology of antennae in Cicadoidea (Insecta: Hemiptera), with respect to functional, taxonomic and phylogenetic implications. Zoologischer Anzeiger 276: 57–70. https://doi.org/10.1016/j.jcz.2018.05.003
- Wang Y, Brożek J, Dai W (2020) Functional morphology and sexual dimorphism of antennae of the pear lace bug Stephanitis nashi (Hemiptera: Tingidae). Zoologischer Anzeiger 286: 11–19. https://doi.org/10.1016/j.jcz.2020.03.001
- Wiesenborn WD (2004) Mouth parts and alimentary canal of Opsius stactogalus Fieber (Homoptera: Cicadellidae). Journal of the Kansas Entomological Society 77(2): 152–155. https://doi.org/10.2317/0307.28.1
- Willis D (1949) The anatomy and histology of the head, gut and associated structures of Typhlocyba ulmi. Proceedings of the Zoological Society of London 118(4): 984–1001. https://doi.org/10.1111/j.1096-3642.1949.tb00416.x
- Yan L, Du Z, Wang H, Zhang S, Cao M, Wang X (2018) Identification and characterization of wheat yellow striate virus, a novel leafhopper-transmitted nucleorhabdovirus infecting wheat. Frontiers in Microbiology 9: e468. https://doi.org/10.3389/fmicb.2018.00468
- Zacharuk RY (1980) Ultrastructure and function of insect chemosensilla. Annual Review of Entomology 25(1): 27–47. https://doi.org/10.1146/annurev.en.25.010180.000331
- Zhang CN, Pan LX, Lu YP, Dietrich C, Dai W (2016) Reinvestigation of the antennal morphology of the white-backed planthopper Sogatella furcifera (Horváth) (Hemiptera: Delphacidae). Zoologischer Anzeiger 262: 20–28. https://doi.org/10.1016/j.jcz.2016.03.011
- Zhang Y, Dietrich CH, Duan Y (2020) Structure and sensilla of the mouthparts of Alobaldia tobae, Maiestas dorsalis and Stirellus indrus (Hemiptera: Cicadellidae: Deltocephalinae). Zoomorphology 139(2): 189–198. https://doi.org/10.1007/s00435-020-00478-1
- Zhao LQ, Dai W, Zhang CN, Zhang YL (2010) Morphological characterization of the mouthparts of the vector leafhopper Psammotettix striatus (L.) (Hemiptera: Cicadellidae). Micron (Oxford, England) 41(7): 754–759. https://doi.org/10.1016/j.micron.2010.06.001