Research Article |
|
Corresponding author: Valentin Moser ( valentinmoser@hotmail.com ) Corresponding author: Hannes Baur ( hannes.baur@nmbe.ch ) Academic editor: Tony Robillard
© 2021 Valentin Moser, Hannes Baur, Arne W. Lehmann, Gerlind U. C. Lehmann.
This is an open access article distributed under the terms of the Creative Commons Attribution License (CC BY 4.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Citation:
Moser V, Baur H, Lehmann AW, Lehmann GUC (2021) Two species? – Limits of the species concepts in the pygmy grasshoppers of the Tetrix bipunctata complex (Orthoptera, Tetrigidae). ZooKeys 1043: 33-59. https://doi.org/10.3897/zookeys.1043.68316
|
Abstract
Today, integrative taxonomy is often considered the gold standard when it comes to species recognition and delimitation. Using the Tetrix bipunctata complex, we here present a case where even integrative taxonomy may reach its limits. The Tetrix bipunctata complex consists of two morphs, bipunctata and kraussi, which are easily distinguished by a single character, the length of the hind wing. Both morphs are widely distributed in Europe and reported to occur over a large area in sympatry, where they occasionally may live also in syntopy. The pattern has led to disparate classifications, as on the one extreme, the morphs were treated merely as forms or subspecies of a single species, on the other, as separate species. For this paper, we re-visited the morphology by using multivariate ratio analysis (MRA) of 17 distance measurements, checked the distributional data based on verified specimens and examined micro-habitat use. We were able to confirm that hind wing length is, indeed, the only morphological difference between bipunctata and kraussi. We were also able to exclude a mere allometric scaling. The morphs are, furthermore, largely sympatrically distributed, with syntopy occurring regularly. However, a microhabitat niche difference can be observed. Ecological measurements in a shared habitat confirm that kraussi prefers a drier and hotter microhabitat, which possibly also explains the generally lower altitudinal distribution. Based on these results, we can exclude classification as subspecies, but the taxonomic classification as species remains unclear. Even with different approaches to classify the Tetrix bipunctata complex, this case is, therefore, not settled. We recommend continuing to record kraussi and bipunctata separately.
Keywords
Allometry, integrative taxonomy, morphometry, Orthoptera, species delimitation, Tetrigidae, Tetrix
Introduction
Species concepts shape the way we see an individual from a given population. Species are the fundamental unit in evolutionary biology (
The Tetrix bipunctata complex is an intriguing case: T. bipunctata (Linnaeus, 1758) and T. kraussi Saulcy, 1888 (see

The status of the two morphs has always been controversial.
Some authors have suggested that there are further morphological characters besides the hind wing that would allow us to distinguish the two morphs. Koch and Meineke (in
No genetic differences have been found so far, as the two morphs form a single cluster when compared using COI barcoding (
In this study, we examine the morphs bipunctata and kraussi and discuss their status, based on new data from: (1) multivariate morphometry, (2) biogeography in Central Europe and (3) microhabitat niche use in syntopy.
- 1) Concerning morphological characters, we address the following questions:
- – Are further characters – besides wing length – important for the separation of bipunctata and kraussi and to what extent? Some authors claim that body proportions seem to differ; however, nobody has ever tried to quantify those traits.
- – What are the best shape characters for separating bipunctata and kraussi? As mentioned before, so far only a single ratio, hind wing length to tegmen length (either by taking into account the entire hind wing length or just the part projecting beyond the tegmen), has been used regularly. A morphometric analysis thus might reveal some more reliable ratios.
- – Despite the evidence for two distinct morphs (
Fischer 1948 ;Schulte 2003 ), specimens with intermediate wing ratios have been reported byNadig (1991) . Therefore, we re-examined Nadig’s collection including the specimens in question. - – How much allometry is present? Size-dependent variation in the adult stage (static allometry, see
Gould 1966 ;Klingenberg 2008 ,2016 ;Anichini et al. 2017 ;Rebrina et al. 2020 ) plays a major role in such investigations, but so far, it has been neglected in this complex. Here, we analyse which characters and character ratios correlate with body size.
- 2) Biogeography
Due to the uncertain taxonomic situation, the distribution is far from settled, as many authors have not differentiated between bipunctata and kraussi. Furthermore, a substantial number of misidentifications have been published for Tetrigidae (own results, compare
- 3) Ecology of habitat use at a syntopic population in Brandenburg
The segregated distribution of bipunctata and kraussi is interpreted as an ecological separation (
Materials and methods
Identification of specimens
Below, we consistently refer to the morphs as “bipunctata” and “kraussi” and treat them in the sense of operational taxonomic units. For the assignment of specimens to morphs, we adopted the identifications found on the labels in the Swiss collections. This was mainly the case for specimens in Nadig’s collection, also with respect to what he considered as intermediate specimens. In all other instances we followed current practice (
1) Morphometry
Character measurements
We measured 20 characters from all over the body to cover the most relevant variation in size and shape between bipunctata and kraussi. The selection of characters was based on
Abbreviation, name, definition and magnification (on Keyence digital microscope) of the 20 measurements used for the morphometric analyses of Tetrix bipunctata complex females. General morphology follows
| No. | Abbrev. | Character name | Character definition | Magnification |
|---|---|---|---|---|
| 1 | bt3.l | Basitarsus length | Length of basitarsus of hind tarsus, from proximal expansion to apex, outer aspect along ventral side | 150 |
| 2 | eye.b | Eye breadth | Greatest breadth of eye, lateral view | 150 |
| 3 | eye.h | Eye height | Greatest height of eye, lateral view | 150 |
| 4 | fl5.b | 5th flagellomere breadth | Greatest breadth of 5th flagellomere, dorsal (inner) aspect | 150 |
| 5 | fl5.l | 5th flagellomere length | Greatest length of 5th flagellomere, dorsal (inner) aspect | 150 |
| 6 | fm2.b | Mid-femur breadth | Greatest breadth of mid-femur, lateral view | 100 |
| 7 | fm2.l | Mid-femur length | Length of mid-femur, from proximal emargination of trochanter to emargination of knee, lateral view | 100 |
| 8 | fm3.b | Hind femur breadth | Greatest breadth of hind femur, lateral view | 30 |
| 9 | fm3.l | Hind femur length | Length of hind femur, from proximal edge to tip of knee disc, lateral view | 30 |
| 10 | fro.h | Frons height | Height of frons, from lower margin of clypeus to lower margin of eye orbit, frontal view | 100 |
| 11 | hea.b | Head breadth | Greatest breadth of head, dorsal view | 100 |
| 12 | hwi.l | Hind wing length | Length of hind wing, from proximal edge of tegmen to tip of hind wing, in situ. Remark: Very often, only the part protruding below the tegmen has been considered. Unfortunately, the measurement is then critically dependent on the position of the tegmen, which is often displaced relative to the hind wing. We, therefore, preferred the entire hind wing length, which can be measured rather more reliably | 30 |
| 13 | prn.b | Pronotum breadth | Greatest breadth of pronotum, dorsal view | 30 |
| 14* | prn.h | Pronotum height | Greatest height of pronotum, from carina humeralis at level of proximal edge of tegmen to highest point of carina medialis, exact lateral view | 30 |
| 15 | prn.l | Pronotum length | Length of pronotum, from anterior margin to the tip of the posterior pronotal process, dorsal view along carina medialis | 30 |
| 16* | pu2.l | 2nd pulvillus length | Length of 2nd pulvillus on basitarsus of hind tarsus, from its proximal notch to distal notch, outer aspect | 150 |
| 17* | pu3.l | 3rd pulvillus length | Length of 3rd pulvillus on basitarsus of hind tarsus, from its proximal notch to distal notch, outer aspect | 150 |
| 18 | teg.b | Tegmen breadth | Greatest breadth of sclerotised part of tegmen, outer aspect | 100 |
| 19 | teg.l | Tegmen length | Length of fore wing, from proximal edge of tegmen to tip of fore wing, outer aspect | 100 |
| 20 | vrt.b | Vertex breadth | Shortest breadth of vertex, dorsal view. Together with head breath, this covers also potential differences in eye breath. | 100 |
Overview on Tetrix bipunctata complex populations (females only) included in the morphometric analyses. Most specimens are from the Nadig collection in
| Country | Population |
|---|---|
| AT | Kärnten |
| CH | BE Beatenberg |
| CH | BE/JU Jura |
| CH | GR Oberengadin |
| CH | GR Schams |
| CH | GR Unterengadin |
| CH | UR Urnerboden |
| DE | S-Bayern |
| DE | Schwarzwald |
| IT | Chiavenna |
| IT | Como |
| IT | Gardasee |
| IT | S-Tirol E/Mittenwald |
| IT | Trentino |
Each character was photographed with a Keyence VHX 2000 digital microscope and a VH-Z20R/W zoom lens at different magnification, depending on the size of the body part (see Table
Multivariate ratio analysis of the body measurements
For the data analysis, we applied multivariate ratio analysis (MRA) (
Very often shape correlates with size, which corresponds to the well-known phenomenon of allometry. In the case of specimens belonging to the same stage, in our case adults, we are talking of static allometry (
For a sensible interpretation of morphometric results, it is therefore essential to consider allometric variation. In many studies, such variation is simply removed from the data by various “correction” procedures (
We first performed a series of shape PCAs to see how well the morphs were supported by variation in shape. A shape PCA shows in very few axes (usually just the first one or two shape PCs are important) the unconstrained pattern of variation in the data. A PCA type of analysis is convenient here, as it does not require a priori assignment of specimens to a particular group, but assumes that all belong to a single group. We could thus avoid bias with respect to groupings (
We, furthermore, employed the PCA ratio spectrum that allows an easy interpretation of shape PCs in terms of body ratios. In a PCA ratio spectrum, the eigenvector coefficients of all variables are arranged along a vertical line. Ratios calculated from variables lying at the opposite ends of the spectrum have the largest influence on a particular shape PCA; ratios from variables lying close to each other or in the middle of the graph are negligible (
The situation changes once we specifically ask for differences between groups. For this question, we use a method where the groups are specified a priori. In the morphometry of distance measurements, such methods are usually based on linear discriminant analysis (LDA) (e.g.
We used the R language and environment for statistical computing for data analysis, version 4.0.3 (R Core Team 2020). For MRA, we employed the R-scripts provided by
Raw data in millimetres and the complete set of photographs with measurements, as well as the R-scripts used for the analyses, are available in a data repository on Zenodo (
2) Biogeography
Given the high level of erroneous Tetrigidae determinations in collections, we refrain from incorporating published records. Instead, we concentrate on specimens studied by ourselves from several European Museums and private collections (Table
List of Museums and private collections with material of bipunctata and kraussi studied for the biogeography pattern. Museum codes are unified using the NCBI database (https://www.ncbi.nlm.nih.gov/biocollections/), see also
| Code | Institution |
|---|---|
|
|
Senckenberg Deutsches Entomologisches Institut |
|
|
Muséum d'Histoire Naturelle, Geneva |
|
|
Muséum National d’Histoire Naturelle (Paris) |
|
|
Naturhistorisches Museum Bern |
| NHMV | Müritzeum / Naturhistorische Landessammlungen für Mecklenburg-Vorpommern |
|
|
Naturhistorisches Museum Wien |
| NKML | Naturkundemuseum Leipzig |
|
|
Senckenberg Museum für Naturkunde Görlitz |
|
|
Universiteit van Amsterdam, Zoologisch Museum |
|
|
Museum für Naturkunde Berlin |
|
|
Zoologische Staatssammlung München |
| Collectio Gatz | Katharina Gatz, Berlin, Germany |
| Collectio Gomboc | Stanislav Gomboc, Kranj,Slovenia |
| Collectio Hochkirch | Prof. Axel Hochkirch, Trier, Germany |
| Collectio Karle-Fendt | Alfred Karle-Fendt, Sonthofen, Germany |
| Collectio Landeck | Ingmar Landeck, Finsterwalde, Germany |
| Collectio Lehmann | Dr. Arne Lehmann, Stahnsdorf, Germany |
| Collectio Muth | Martin Muth, Kempten, Germany |
Specimens were assigned to each morph by calculating the standard ratio (see above). After eliminating erroneous determinations by our precursors, nymphs and a single specimen of the f. macroptera which cannot be associated with either bipunctata or kraussi so far, we were able to include 660 specimens from the six Central Europe countries Germany, Netherlands, Switzerland, Austria, Italy and Slovenia (Suppl. material
For the generation of the map, we used QGis 3.10.13-A Coruna and the Natural Earth Data (https://www.naturalearthdata.com/about/terms-of-use/, https://www.openstreetmap.org/copyright, OpenStreetMap contributors).
3) Microhabitat niches
In a syntopic population in Brandenburg (2.5 km E of Theisa 51.542°N, 13.503°E), the microhabitat use was studied for four months from May to August 2015 by Katharina Gatz, supervised by G.U.C. Lehmann. By slowly walking through the habitat, individuals were located either sitting or jumping from a retraceable spot. At the point of origin, a little flag was placed and the animal afterwards caught with the help of a 200 ml plastic vial (Greiner BioOne) (Fig.
Results
Measurement data
Appendix
Analysis using shape PCA
We first performed a series of shape PCAs to see how well the morphs were supported by variation in shape and which body ratios were responsible for separation (Fig.

Shape principal component analysis (shape PCA) of 273 females of Tetrix bipunctata and kraussi A analysis including 17 variables, scatterplot of first against second shape PC; in parentheses the variance explained by each shape PC B PCA ratio spectrum for first shape PC C PCA ratio spectrum for second shape PC. Horizontal bars in the ratio spectra represent 68% bootstrap confidence intervals, based on 1000 replicates; only the most important characters are indicated in ratio spectra.
In the scatterplot of the first against second shape PC, the individuals were almost perfectly separated along the first shape PC, but entirely overlapping along the second (Fig.
With respect to the second shape PC, the situation is quite different as there is broad overlap between bipunctata and kraussi. According to its PCA ratio spectrum (Fig.
Allometry
Plotting isosize against the first shape PC revealed that intraspecific allometry was weak in bipunctata and moderate in kraussi (Fig.

Extracting best ratios
The LDA ratio extractor found hind wing length to mid-femur length as the best ratio for separating bipunctata from kraussi. This ratio was indeed more powerful than the standard ratio (compare Fig.

Boxplots of body ratios of 273 females of Tetrix bipunctata and kraussi A hind wing length to mid-femur length, the ratio selected by the LDA ratio extractor as the best ratio for separating the morphs B hind wing length to tegmen length, the standard ratio used for discrimination C tegmen length to hind femur length, the second best ratio found by the LDA ratio extractor (actually the best ratio when hind wing length is omitted). Means in all plots significantly different (ANOVA, p < 0.001).
The specimens considered as “Nadig intermediates” (“Zwischenformen”) are found in both groups. In the plot with the best ratio (Fig.

Scatterplots of isosize against body ratios of 273 females of Tetrix bipunctata and kraussi, showing the position of intermediate specimens A isosize against ratio of hind wing length to mid-femur length, the best ratio for separation of morphs B isosize against ratio of hind wing length to tegmen length, the standard ratio for discrimination (see Fig.
Biogeography
In total, 660 specimens from 286 localities could be included into our biogeographic analysis (Suppl. material

Distribution of 260 localities with records of Tetrix bipunctata (green dots), kraussi (orange dots) and syntopic populations (purple dots), mapped for six central European countries. Map generated using Natural Earth Data https://www.naturalearthdata.com/about/terms-of-use/.

Altitudinal distribution (mean ± SD) of 286 populations of Tetrix bipunctata (green) and kraussi (orange) segmented for five Central European countries and eight Federal States in Germany. Regions are grouped along the north-south axis, NL = The Netherlands, DE = Germany: DE MV = Mecklenburg-Vorpommern, DE BB = Brandenburg, DE ST = Sachsen-Anhalt, DE SN = Sachsen, DE TH = Thüringen, DE HE = Hessen, DE BW = Baden-Württemberg, DE BY = Bayern, AT = Austria, CH = Switzerland, IT = Italy, SL = Slovenia.
Microhabitat niches
In the syntopic population in Brandenburg, adults of bipunctata and kraussi show separated microhabitat niche use. While bipunctata adults preferentially inhabit denser vegetation with higher plants (Fig.

Microhabitats of bipunctata had a mean vegetation cover of 70 ± 18%, nearly twice as dense as the vegetation at kraussi spots (40 ± 7%) (Fig.

The vegetation at sites inhabited by bipunctata adults was on average 27 cm ± 12 cm tall, nearly twice as high as the plants at patches with kraussi occurrence (16 cm ± 4 cm) (Fig.

Discussion
The morphometric analyses revealed that the morphs are merely separated by hind wing length or hind wing length in combination with any other character as a shape ratio. It was thus, by far, the most important character (Figs
Isometric size between the morphs is widely overlapping with bipunctata being slightly larger on average (Fig.
In conclusion, we did find clear morphometric differences between bipunctata and kraussi only in hind wing length and all ratios including this variable. This is in agreement with results by
Nadig’s intermediate specimens and the subspecies hypothesis
Our analyis shows that the specimens from the Engadin, determined as intermediates (“Zwischenformen”) by
Based on his observation,
Habitat differentiation
The morphs show a preference for slightly different habitats, with kraussi preferring shorter and less dense vegetation cover (Fig.
The question whether kraussi and bipunctata represent different species or should be interpreted as infraspecific morphs is still open. The lack of genetic differentiation (see
More research is needed to distinguish between the two possibilities that bipunctata and kraussi are genetically young species or infraspecific ecomorphs. However, this is a prime example how even modern species concepts can reach their limits. What we can exclude is their status as subspecies. Missing evidence concerns the genetic and developmental mechanisms behind the wing length. Crossing experiments could, furthermore, be informative to study reproductive barriers and hybrid disadvantage. We recommend that bipunctata and kraussi are considered as separate units until the species question can be answered more precisely.
Acknowledgements
We thank Elsa Obrecht (
References
- Ali RF, Neiber MT, Walther F, Hausdorf B (2016) Morphological and genetic differentiation of Eremina desertorum (Gastropoda, Pulmonata, Helicidae) in Egypt. Zoologica Scripta 45: 48–61. https://doi.org/10.1111/zsc.12134
- Anichini M, Kuchenreuther S, Lehmann GUC (2017) Allometry of male sound-producing structures indicates sexual selection on wing size and stridulatory teeth density in a bushcricket. Journal of Zoology 301: 271–279. https://doi.org/10.1111/jzo.12419
- Bartels PJ, Nelson DR, Exline RP (2011) Allometry and the removal of body size effects in the morphometric analysis of tardigrades. Journal of Zoological Systematics and Evolutionary Research 49: 17–25. https://doi.org/10.1111/j.1439-0469.2010.00593.x
- Bartlett JW, Frost C (2008) Reliability, repeatability and reproducibility: analysis of measurement errors in continuous variables. Ultrasound in Obstetrics and Gynecology 31: 466–475. https://doi.org/10.1002/uog.5256
- Baur B, Baur H, Roesti C, Roesti D (2006) Die Heuschrecken der Schweiz. 1. Haupt, Bern, 352 pp. https://books.google.ch/books/about/Die_Heuschrecken_der_Schweiz.html?id=e8NJAAAACAAJ&redir_esc=y
- Baur H, Leuenberger C (2011) Analysis of ratios in multivariate morphometry. Systematic Biology 60: 813–825. https://doi.org/10.1093/sysbio/syr061
- Baur H, Leuenberger C (2020) Multivariate Ratio Analysis (MRA): R-scripts and tutorials for calculating Shape PCA, Ratio Spectra and LDA Ratio Extractor. Zenodo. https://doi.org/10.5281/zenodo.3890195
- Baur H, Kranz-Baltensperger Y, Cruaud A, Rasplus J-Y, Timokhov AV, Gokhman VE (2014) Morphometric analysis and taxonomic revision of Anisopteromalus Ruschka (Hymenoptera: Chalcidoidea: Pteromalidae)–an integrative approach. Systematic Entomology 39: 691–709. https://doi.org/10.1111/syen.12081
- Bellmann H, Rutschmann F, Roesti C, Hochkirch A (2019) Der Kosmos Heuschreckenführer. Frankh-Kosmos Verlags-GmbH, Stuttgart, 430 pp.
- Braby MF, Eastwood R, Murray N (2012) The subspecies concept in butterflies: has its application in taxonomy and conservation biology outlived its usefulness? Biological Journal of the Linnean Society 106: 699–716. https://doi.org/10.1111/j.1095-8312.2012.01909.x
- Buuren S van, Groothuis-Oudshoorn K (2011) mice: Multivariate Imputation by Chained Equations in R. Journal of Statistical Software 45: 1–67. https://doi.org/10.18637/jss.v045.i03
- Cigliano MM, Braun H, Eades DC, Otte D (2021) Orthoptera Species File. Version 5.0/5.0. [2021_03_20] http://Orthoptera.SpeciesFile.org
- Coyne JA, Orr HA (2004) Speciation. Sinauer, 480 pp.
- Dayrat B (2005) Towards integrative taxonomy. Biological Journal of the Linnean Society 85: 407–415. https://doi.org/10.1111/j.1095-8312.2005.00503.x
- De Queiroz K (2007) Species concepts and species delimitation. Systematic Biology 56: 879–886. https://doi.org/10.1080/10635150701701083
- Default B, Morichon D (2015) Criquets de France, Volume 1, fascicule A et B (Orthoptera: Caelifera). Faune de France, 738 pp. https://www.nhbs.com/faune-de-france-volume-97-criquets-de-france-volume-1-fascicule-a-et-b-orthoptera-caelifera-2-volume-set-book [January 26, 2021]
- Devriese H (1996) Bijdrage tot de systematiek, morfologie en biologie van de West-Palearktische Tetrigidae. Nieuwsbrief Saltabel 15: 1–50.
- Evenhuis NL (2002) Publication and dating of the two “Bulletins” of the Société Entomologique de France (1873–1894). Zootaxa 70: 1–32. https://doi.org/10.11646/zootaxa.70.1.1
- Fischer H (1948) Die schwäbischen Tetrix-Arten (Heuschrecken). Bericht der Naturforschenden Gesellschaft Augsburg 1: 40–87.
- Fischer J, Steinlechner D, Zehm A, Poniatowski D, Fartmann T, Beckmann A, Stettmer C (2020) Die Heuschrecken Deutschlands und Nordtirols: Bestimmen – Beobachten – Schützen. 2., korrigierte Auflage. Quelle & Meyer, Wiebelsheim, 371 pp.
- Gould SJ (1966) Allometry and size in ontogeny and phylogeny. Biological Reviews 41: 587–638. https://doi.org/10.1111/j.1469-185X.1966.tb01624.x
- Günther K (1979) Die Tetrigoidea von Afrika südlich der Sahara (Orthoptera: Caelifera). Beiträge zur Entomologie = Contributions to Entomology 29: 7–183. https://doi.org/10.21248/contrib.entomol.29.1.7-183
- Harz K (1957) Die Geradflügler Mitteleuropas. Fischer, Jena, 494 pp.
- Harz K (1975) Die Orthopteren Europas – The Orthoptera of Europe II. Dr. W. Junk B. V., The Hague, 939 pp. https://doi.org/10.1007/978-94-010-1947-7
- Hastie T, Tibshirani R, Friedman J (2009) The elements of statistical learning. 2nd edn. Springer, New York, 745 pp. http://statweb.stanford.edu/~tibs/book/preface.ps [June 26, 2017]
- Hawlitschek O, Morinière J, Lehmann GUC, Lehmann AW, Kropf M, Dunz A, Glaw F, Detcharoen M, Schmidt S, Hausmann A, Szucsich NU, Caetano-Wyler SA, Haszprunar G (2017) DNA barcoding of crickets, katydids and grasshoppers (Orthoptera) from Central Europe with focus on Austria, Germany and Switzerland. Molecular Ecology Resources 17: 1037–1053. https://doi.org/10.1111/1755-0998.12638
- Hebert PDN, Cywinska A, Ball SL, deWaard JR (2003) Biological identifications through DNA barcodes. Proceedings of the Royal Society of London. Series B: Biological Sciences 270: 313–321. https://doi.org/10.1098/rspb.2002.2218
- Huber C, Schnitter PH (2020) Nebria (Pseudonebriola) tsambagarav sp. nov., a new alpine species from the Mongolian Altai (Coleoptera, Carabidae). Alpine Entomology 4: 29–38. https://doi.org/10.3897/alpento.4.50408
- Jolicoeur P (1963) 193. Note: The Multivariate Generalization of the Allometry Equation. Biometrics 19: 497–499. https://doi.org/10.2307/2527939
- Kevan DKMcE (1953) The status of Tetrix bipunctatum (Linn.) (Orthoptera; Tetrigidae) in Britain. – Entomologist’s Gazette 4: 205–224.
- Klingenberg CP (2008) Morphological Integration and Developmental Modularity. Annual Review of Ecology, Evolution and Systematics 39: 115–132. https://doi.org/10.1146/annurev.ecolsys.37.091305.110054
- Klingenberg CP (2016) Size, shape, and form: concepts of allometry in geometric morphometrics. Development Genes and Evolution 226: 113–137. https://doi.org/10.1007/s00427-016-0539-2
- László Z, Baur H, Tóthmérész B (2013) Multivariate ratio analysis reveals Trigonoderus pedicellaris Thomson (Hymenoptera, Chalcidoidea, Pteromalidae) as a valid species. Systematic Entomology 38: 753–762. https://doi.org/10.1111/syen.12026
- Lawrence JF, Nielsen ES, Mackerras IM (1991) Skeletal anatomy and key to orders. In: Naumann ID (Ed.) Insects of Australia: A textbook for students and research workers. Division of Entomology, CSIRO, Carlton, 3–32.
- Le NH, Nahrung HF, Morgan JAT, Lawson SA (2020) Multivariate ratio analysis and DNA markers reveal a new Australian species and three synonymies in eucalypt-gall-associated Megastigmus (Hymenoptera: Megastigmidae). Bulletin of Entomological Research 110: 709–724. https://doi.org/10.1017/S000748532000022X
- Lehmann AW (2004) Die Kurzflügel-Dornschrecke Tetrix (bipunctata) kraussi Saulcy, 1888: eine missachtete (Unter-) Art. Articulata 19: 227–228.
- Lehmann AW, Landeck I (2011) Erstfund der Kurzflügel-Dornschrecke Tetrix kraussi Saulcy, 1888 im Land Brandenburg (Orthoptera: Tetrigidae). Märkische Entomologische Nachrichten 13: 227–232.
- Lehmann AW, Devriese H, Tumbrinck J, Skejo J, Lehmann GUC, Hochkirch A (2017) The importance of validated alpha taxonomy for phylogenetic and DNA barcoding studies: a comment on species identification of pygmy grasshoppers (Orthoptera, Tetrigidae). ZooKeys 679: 139–144. https://doi.org/10.3897/zookeys.679.12507
- Lehmann GUC, Marco H, Lehmann AW, Gäde G (2018) Seasonal differences in body mass and circulating metabolites in a wing-dimorphic pygmy grasshopper – implications for life history? Ecological Entomology 43: 675–682. https://doi.org/10.1111/een.12647
- Linnaeus C (1758) Systema naturae per regna tria naturæ, secundum classes, ordines, genera, species, cum characteribus, differentiis, synonymis, locis (10th edn.). Impensis direct Laurentii Salvii, Stockholm, 847 pp. https://www.biodiversitylibrary.org/page/726886
- Lougheed SC, Arnold TW, Bailey RC (1991) Measurement error of external and skeletal variables in birds and its effect on principal components. The Auk 108: 432–436.
- Mallet J (2007) Subspecies, semispecies, superspecies. Encyclopedia of Biodiversity 5: 1–5. https://doi.org/10.1016/B0-12-226865-2/00261-3
- Massa B, Fontana P, Buzzetti FM, Kleukers RMJC, Baudewijn O (2012) Orthoptera. Calderini-Edizioni, Bologna, 563 pp.
- Mayr E (1963) Animal species and evolution. Belknap Press of Harvard University Press, Cambridge, [xiv,] 797 pp.
- Moser V, Baur H (2021) Morphometric data from: Two species? – Limits of the species concepts in the pygmy grasshoppers of the Tetrix bipunctata complex (Orthoptera: Tetrigidae). Zenodo. https://doi.org/10.5281/zenodo.4818525
- Mosimann JE (1970) Size allometry size and shape variables with characterizations of the lognormal and generalized gamma distributions. Journal of the American Statistical Society 65: 930–945. https://doi.org/10.1080/01621459.1970.10481136
- Nadig A (1991) Die Verbreitung der Heuschrecken (Orthoptera:Saltatoria) auf einem Diagonalprofil durch die Alpen (Inntal-Maloja-Bregaglia-Lago di Como-Furche. Jahresbericht der Naturforschenden Gesellschaft Graubündens 102: 277–378.
- Nakagawa S, Schielzeth H (2010) Repeatability for Gaussian and non-Gaussian data: a practical guide for biologists. Biological Reviews of the Cambridge Philosophical Society 85: 935–956. https://doi.org/10.1111/j.1469-185X.2010.00141.x
- Pfeifer MA, Niehuis M, Renker C (2011) Fauna und Flora in Rheinland-Pfalz Beiheft Die Fang- und Heuschrecken in Rheinland-Pfalz. GNOR, Landau, 678 pp.
- Pimentel RA (1979) Morphometrics, the Multivariate Analysis of Biological Data. Kendall/Hunt Pub. Co, Dubuque, Iowa, 276 pp.
- R Core Team (2021) R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing, Vienna. https://www.R-project.org/
- Rebrina F, Anichini M, Reinhold K, Lehmann GUC (2020) Allometric scaling in two bushcricket species (Orthoptera: Tettigoniidae) suggests sexual selection on song-generating structures. Biological Journal of the Linnean Society 131: 521–535. https://doi.org/10.1093/biolinnean/blaa122
- Sardet E, Dehondt F, Mora F (2015a) Tetrix bipunctata (L., 1758) et Tetrix kraussi Saulcy, 1889 en France: répartition nationale, biométrie, écologie, statut et sympatrie (Orthoptera: Caelifera, Tetrigoidea, Tetrigidae). Matériaux Orthoptériques et Entomocénotiques 20: 15–24.
- Sardet É, Braud Y, Roesti C, Koch V (2015b) Cahier d’identification des Orthoptères de France, Belgique, Luxembourg & Suisse. Biotope éditions, Mèze, 304 pp.
- Saulcy F (1888) Notice sur le genre Tetrix Latreille. Bulletin de la Société Entomologique de France 6: 135–136.
- Schneider C, Rasband W, Eliceiri K (2012) NIH Image to ImageJ: 25 years of image analysis. Nature Methods 9: 671–675. https://doi.org/10.1038/nmeth.2089
- Schulte AM (2003) Taxonomie, Verbreitung und Ökologie von Tetrix bipunctata (Linnaeus 1758) und Tetrix tenuicornis (Sahlberg 1893) (Saltatoria: Tetrigidae). Articulata, Beiheft 10: 1–226.
- Seifert B (2002) How to distinguish most similar insect species – improving the stereomicroscopic and mathematical evaluation of external characters by example of ants. Journal of Applied Entomology 126: 445–454. https://doi.org/10.1046/j.1439-0418.2002.00693.x
- Selz OM, Dönz CJ, Vonlanthen P, Seehausen O (2020) A taxonomic revision of the whitefish of Lakes Brienz and Thun, Switzerland, with descriptions of four new species (Teleostei, Coregonidae). ZooKeys 989: 79–162. https://doi.org/10.3897/zookeys.989.32822
- Sharma S, Ciufo S, Starchenko E, Darji D, Chlumsky L, Karsch-Mizrachi I, Schoch CL (2018) The NCBI BioCollections Database. Database 2018: bay006. https://doi.org/10.1093/database/bay006
- Sidlauskas BL, Mol JH, Vari RP (2011) Dealing with allometry in linear and geometric morphometrics: a taxonomic case study in the Leporinus cylindriformis group (Characiformes: Anostomidae) with description of a new species from Suriname. Zoological Journal of the Linnean Society 162: 103–130. https://doi.org/10.1111/j.1096-3642.2010.00677.x
- Sites JW, Marshall JC (2004) Operational Criteria for Delimiting Species. Annual Review of Ecology, Evolution and Systematics 35: 199–227. https://doi.org/10.1146/annurev.ecolsys.35.112202.130128
- Steenman A, Lehmann AW, Lehmann GUC (2013) Morphological variation and sex-biased frequency of wing dimorphism in the pygmy grasshopper Tetrix subulata (Orthoptera: Tetrigidae). European Journal of Entomology 110: e535. https://doi.org/10.14411/eje.2013.071
- Steenman A, Lehmann AW, Lehmann GUC (2015) Life-history trade-off between macroptery and reproduction in the wing-dimorphic pygmy grasshopper Tetrix subulata (Orthoptera Tetrigidae). Ethology Ecology & Evolution 27: 93–100. https://doi.org/10.1080/03949370.2014.885466
- Tumbrinck J (2014) Taxonomic revision of the Cladonotinae (Orthoptera: Tetrigidae) from the islands of South-East Asia and from Australia, with general remarks to the classification and morphology of the Tetrigidae and descriptions of new genera and species from New Guinea and New Caledonia. In: Telnov D (Ed.) Biodiversity, biogeography and nature conservation in Wallacea and New Guinea. Riga, the Entomological Society of Latvia, 345–396.
- Wägele JW (2005) Foundations of Phylogenetic Systematics. (1st edn.). Pfeil, F, München, 365 pp.
- Warton DI, Wright IJ, Falster DS, Westoby M (2006) Bivariate line-fitting methods for allometry. Biological Reviews 81: 259–291. https://doi.org/10.1017/S1464793106007007
- West-Eberhard MJ (2003) Developmental Plasticity and Evolution. Illustrated Edition. Oxford University Press, Oxford, 816 pp. https://doi.org/10.1093/oso/9780195122343.001.0001
- Wickham H (2016) ggplot2: elegant graphics for data analysis (2nd edn.). Springer, Huston, Texas, xvi, 260 pp.
- Will KW, Mishler BD, Wheeler QD (2005) The perils of DNA barcoding and the need for integrative taxonomy. Systematic Biology 54: 844–851. https://doi.org/10.1080/10635150500354878
- Willemse L, Kleukers RMJC, Baudewijn O (2018) The Grasshoppers of Greece. EIS Kennniscentrum Insecten & Naturalis Biodiversity Center, Leiden, 439 pp. https://www.nhbs.com/the-grasshoppers-of-greece-book [January 26, 2021]
- Wranik W, Meitzner V, Martschei T (2008) Beiträge zur floristischen und faunistischen Erforschung des Landes Mecklenburg-Vorpommern (Lung M.-V.) Verbreitungsatlas der Heuschrecken Mecklenburg-Vorpommerns. Steffen GmbH, Friedland, 281 pp.
- Yeates DK, Seago A, Nelson L, Cameron SL, Joseph L, Trueman JWH (2011) Integrative taxonomy, or iterative taxonomy? Systematic Entomology 36: 209–217. https://doi.org/10.1111/j.1365-3113.2010.00558.x
- Zachos FE (2016) Species Concepts in Biology: Historical Development, Theoretical Foundations and Practical Relevance. Springer International Publishing, 220 pp. https://doi.org/10.1007/978-3-319-44966-1
- Zuna-Kratky T, Landman A, Illich I, Zechner L, Essl F, Lechner K, Ortner A, Weissmair W, Wöss G (2017) Die Heuschrecken Österreichs. Denisia 39: 1–872.
Appendix 1
Identification and removal of unreliable characters.
As mentioned under Materials and methods, we omitted three characters from all morphometric analyses presented in the results. In the following, we briefly describe the procedure that led to their removal.
Initially, we started with a shape PCA, based on all 20 characters (see Suppl. material
It is well known that a high quality of measurements is crucial in morphometric data, as low reliability may cause serious problems for multivariate data analysis (
Appendix 2
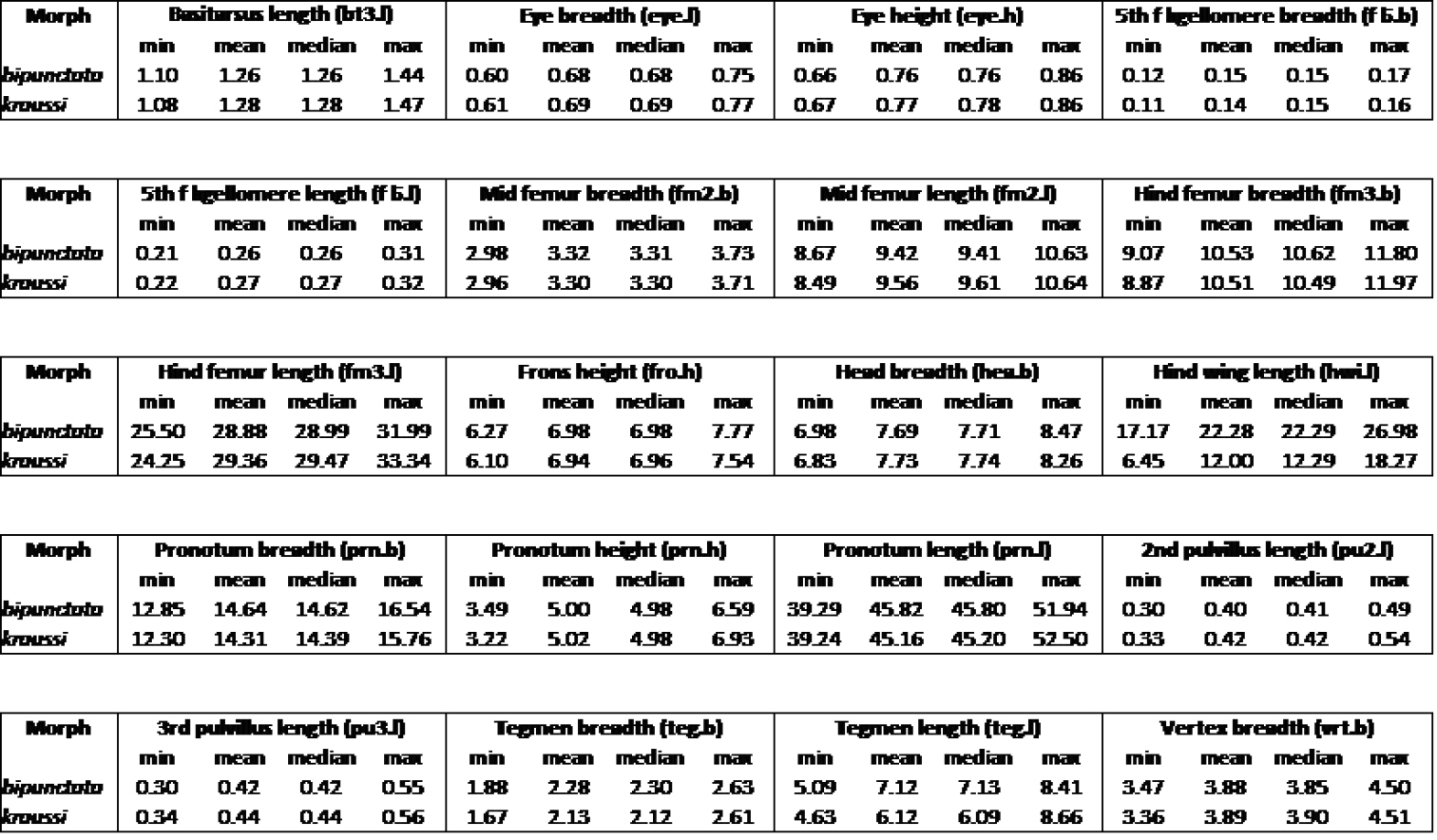
Overview of measurements of Tetrix females, showing minimum, mean, median and maximum in mm.
Supplementary materials
Table S1
Data type: table
Explanation note: Records of 660 specimen of Tetrix bipunctata and T. kraussi, based on our surveys in European Museums and private collections, see Table
The 17 rows coloured represent syntopic occurrences of Tetrix bipunctata and T. kraussi.
Species: z = Zwischenformen, specimen supposed to be intermediates by
Date: Collection date as reported on labels, in square brackets we added the unreported centuries [18] or [19] deduced from our knowledge of collectors biographies.
State: English name of the governmental province.
Bundesland / Kanton: German name of the governmental province.
Geographic coordinates and altitudes: extracted with the help of open mapping tools (https://tools.retorte.ch/map/, https://www.mapcoordinates.net).
Comments: Additional information given on labels.
First and second determination: Identifications based on label information.
Authors’ determination: Identifications based on the standard ratio of the full hind wing length to tegmen length: ≥ 2.5 = bipunctata, < 2.5 = kraussi (corresponding to the ratio of the protruding part of hind wing length to tegmen length of ≥ 1.5 and < 1.5, respectively).
Collectio: Abbreviations of European Museums and private collections with material studied. Museum codes are unified using the NCBI database (https://www.ncbi.nlm.nih. gov/biocollections/), see also
Collection number: Individual codes assigned by the Collectio Lehmann [CL], the Muséum d'Histoire Naturelle, Geneva (MHNG) or Naturhistorisches Museum Bern (NMBE).
Figure S1
Data type: (measurement/occurrence/multimedia/etc.)
Explanation note: Shape principal component analysis (shape PCA) of 273 females of Tetrix bipunctata and kraussi. A: analysis including 20 variables, scatterplot of first against second shape PC. B: PCA ratio spectrum for first shape PC. C: PCA ratio spectrum for second shape PC. Horizontal bars in the ratio spectra represent 68% bootstrap confidence intervals based on 1000 replicates.
Figure S2
Data type: (measurement/occurrence/multimedia/etc.)
Explanation note: Variation in pronotum shape (lateral view) of some Tetrix females included in the morphometric analyses. A–D: bipunctata; E–H: kraussi. The position where pronotum height was measured is indicated by a magenta line.