Research Article |
|
Corresponding author: Robin Casalla ( casallar@uninorte.edu.co ) Academic editor: Eliana Cancello
© 2021 Robin Casalla, Rudolf H. Scheffrahn, Judith Korb.
This is an open access article distributed under the terms of the Creative Commons Attribution License (CC BY 4.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Citation:
Casalla R, Scheffrahn RH, Korb J (2021) Rugitermes ursulae (Isoptera, Kalotermitidae), a new drywood termite from the Caribbean coast of Colombia. ZooKeys 1057: 23-36. https://doi.org/10.3897/zookeys.1057.65877
|
Abstract
Rugitermes ursulae sp. nov. is described from a sample collected inside a dead branch in a tropical dry forest of Colombia’s Caribbean coast using molecular information and external morphological characters of the imago and soldier castes. Rugitermes ursulae sp. nov. soldiers and imagoes are the smallest among all described Rugitermes species. The imago’s head capsule coloration is dark castaneous, while the pronotum is contrastingly pale yellow. Our description includes soldier characters, such as subflangular elevation and shape of the antennal sockets, that can help in identification of samples lacking imagoes.
Keywords
DNA barcoding, imago, northern Colombian coast, soldier, taxonomy, tropical dry forest
Introduction
Records of termites from Colombia have increased in recent years (

Rugitermes Holmgren, 1911, is a genus from the family Kalotermitidae, mainly found in the Neotropics (
Comparison of head measurements (mm) and coloration of soldiers and imagoes of different Rugitermes species.
| No. | Species | Distribution* | Soldier | Imago | Reference | ||||
|---|---|---|---|---|---|---|---|---|---|
| Head width | Mean | Head width | Mean | Head color | Pronotum color | ||||
| 1 | R. ursulae sp. nov. | SA (Colombia) | 1.12–1.22 | 1.17 | 1.10–1.20 | 1.15 | Dark castaneous | Pale yellow | This publication |
| 2 | R. flavicinctus | SA (Guyana) | 1.19–1.35 | 1.27 | 1.19 | 1.19 | Black | Yellow |
|
| 3 | R. rufus | SA (Bolivia) | 1.16–1.56 | 1.42 | 1.19–1.26 | 1.23 | Reddish |
|
|
| 4 | R. magninotus | SA (Guyana, Peru) | 1.38–1.48 | 1.43 | 1.25 | 1.25 | Black | Yellow |
|
| 5 | R. volcanensis | SA (Bolivia) | 1.24–1.88 | 1.54 | 1.21 | 1.21 | Black |
|
|
| 6 | R. athertoni | OC (Polynesia) | 1.47–1.73 | 1.60 | 1.26 | 1.26 | Brown/Black |
|
|
| 7 | R. aridus | SA (Peru) | 1.44–1.72 | 1.60 | NA | NA | Black | Yellow |
|
| 8 | R. niger | SA (Brazil) | 1.50–1.80 | 1.65 | 1.26–1.34 | 1.30 | Black |
|
|
| 9 | R. kirbyi | CA (Costa Rica, Panama) | 1.65 | 1.65 | 1.60 | 1.60 | Black | Yellow |
|
| 10 | R. tinto | SA (Colombia) | 1.52–1.90 | 1.72 | 1.31 | 1.31 | Black | Yellow |
|
| 11 | R. unicolor | CA (Honduras) | 1.80 | 1.80 | 1.60 | 1.60 | Yellow-brown | Yellow |
|
| 12 | R. panamae | CA (Panamá) | 1.8–2.0 | 1.90 | 1.4 | 1.40 | Black | Yellow |
|
| 13 | R. bicolor | SA (Guyana) | 1.80–2.07 | 1.94 | 1.51 | 1.51 | Black | Yellow |
|
| 14 | R. occidentalis | SA (Argentina) | 2.00 | 2.00 | 1.35 | 1.35 | Black | Brown |
|
| 15 | R. laticollis | SA (Bolivia, Ecuador) | 1.83–2.43 | 2.06 | 1.5–2.00 | 1.68 | Black |
|
|
| 16 | R. costaricensis | CA (Costa Rica) | 2.10 | 2.10 | 1.80 | 1.80 | Yellow-brown | Yellow |
|
| 17 | R. nodulosus | SA (Brazil) | NA | NA | NA | NA | Black | Yellow |
|
| 18 | R. rugosus | SA (Brazil) | 1.21 (1.7?) | NA | NA | NA | Black |
|
|
NA = Not available
* CA = Central America, OC = Oceania, SA = South America
Soldier’s scale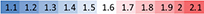
Imago’s scale
Rugitermes species have few species-specific diagnostic characters. The dorsal antennal ridge and the anterolateral corner of the frontal ridge of the soldier head, the size of the eyes of imagoes and soldiers as well as the imago’s head shape can provide useful information to describe a new species (Krishna et al. 1961;
Here, we describe the soldiers and imagoes of Rugitermes ursulae sp. nov. from a sample collected inside a dead branch in the tropical dry forest of Colombia’s Caribbean coast. In addition, we performed molecular analyses based on the marker COII (cytochrome oxidase II) and including representatives of other genera of Kalotermitidae to support species description.
Materials and methods
Study sites and sampling
In July 2014, a survey was done in the tropical dry forest of Colombia’s Caribbean coast (
Identification and genetic analysis
Rugitermes ursulae sp. nov. was compared with Rugitermes samples from the University of Florida Termite Collection (UFTC), USA, and with descriptions and measurements from the literature (
We considered COII sequences for 26 species of Kalotermitidae (if available three species per genus) and the woodroach, Cryptocercus punctulatus, as an outgroup (Suppl. material
We inferred a phylogenetic tree based on the maximum likelihood (ML) approach. We selected the best fitting model using ModelFinder (
We calculated p-distances between COII sequences with MEGA X v.10.1.8 (
Imaging and measurements
Specimens were suspended in hand sanitizer and images were taken with a Leica M205 C stereomicroscope coupled to a Leica MC190 HD digital camera. Helicon Focus software was used to stack pictures. Measurements were done following
Results
Taxonomy
Rugitermes ursulae , sp. nov.
Material examined
Holotype
soldier. Colombia: Colosó, Sucre (9.5435, -75.34884), 400 meters a.s.l., 11.JUL.2014, R. Casalla, ADD-2014-10A. Paratypes. One additional soldier, 12 pseudergates, a pair of reproductive, same colony sample as holotype ADD-2014-10B. Voucher specimen are held at the Universidad del Norte, Colombia. Holotype soldier (ADD-2014-10A) and one reproductive paratype of Rugitermes ursulae sp. nov. (ADD-2014-10B-1) will be deposited at the Natural History Museum of the Alexander von Humboldt Institute of Bogotá (MIAvH, Colombia) and a paratype soldier (ADD-2014-10B-2) and reproductive (ADD-2014-10B-3) at the collection of the American Museum of Natural History, New York. (
Diagnosis
The soldier of R. ursulae sp. nov. is the smallest of all congeneric soldiers (Fig.
| Character | Holotype | Measurements |
|---|---|---|
| Head length with mandibles | 2.81 | 2.81, 2.90 |
| Head length to lateral base of mandibles | 1.87 | 1.87, 1.99 |
| Head width max. | 1.12 | 1.12, 1.22 |
| Head height with postmentum | 1.06 | 1.06, 1.20 |
| Postmentum width min. | 0.22 | 0.22, 0.24 |
| Postmentum length | 1.34 | 1.34, 1.38 |
| Pronotum width | 1.27 | 1.27, 1.39 |
| Pronotum length | 0.63 | 0.63, 0.70 |
| Third antennal article length | 0.16 | 0.16, 0.17 |
| Left mandible length (from dorsal condyle) | 1.17 | 1.17, 1.22 |
| Frontal angle (° degrees) | 34 | 29, 34 |

The imago of R. ursulae sp. nov. is also the smallest of all Rugitermes species (Fig.
Measurements (mm) of Rugitermes ursulae sp. nov. dealated imagoes (N = 2).
| Character | Measurements |
|---|---|
| Eye diameter max. | 0.30, 032 |
| Ocellus diameter max. | 0.09, 0.10 |
| Head width (max. with eyes) | 1.10, 1.20 |
| Head length to tip | 1.44, 1.49 |
| Head height | 0.78, 0.87 |
| Pronotum width max. | 1.25, 1.27 |
| Pronotum length min. | 0.64, 0.70 |
Type locality
“Los primates” Colosó, Sucre, Colombia (Fig.
Description
Soldier
(Fig.
Imago
(Fig.

Distribution and biological observations
‘Los Primates’ is located in the mountains of the municipality of Colosó, Sucre. It is a regional forest reserve created in 1983, containing primary and secondary tropical dry forest. The mean annual temperature is 26.7 °C (min: 25.8 °C; max: 27.8 °C) with an annual precipitation of around 1337 mm (INDERENA, 1983;
Etymology
“Ursulae” derived from a diminutive of the Latin ursa, which means “little bear”, in line with the small size of the species. Ursula is also the name of José Arcadio Buendía’s wife in the novel “One hundred years of solitude” written by Gabriel García Márquez and represents an apology/symbolism for the spiritual engine, entrepreneurship, and hard and silent work of many women around the world.
Molecular analysis of the COII fragment
The topology and splits inferred from the multiple sequence alignment of the COII fragment for all Kalotermitidae genera available in NCBI, and including our new species, revealed a COII ML gene tree that clearly separated R. ursulae sp. nov. from the two other Rugitermes species with maximal BS support. Furthermore, it suggests that the genus Rugitermes is monophyletic (maximal BS support) and that it is the sister taxon of Postelectrotermes, however support values for the latter are low (19% BS; Fig.

Maximum likelihood (ML) tree inferred from COII mtDNA gene sequences of 30 species of Kalotermitidae, including the woodroach, Cryptocercus punctulatus, as outgroup. Numbers before the splits show statistical bootstrap support (BS). Terminals are labeled with the respective NCBI access numbers and genus or species name. Rugitermes ursulae sp. nov. is shown in bold.
The p-distance analyses revealed that the barcode sequences most similar to R. ursulae (accession number: MW600961) belonged to Rugitermes sp. A TB-2014 (accession number: KP026284.1) and Rugitermes ADD 2015-29 (accession number: MW600962); they shared 87% and 86% sequence similarity (p-distance), respectively (Suppl. material
Key to Rugitermes from Colombia
| 1 | Soldier with dark coloration of anterior head capsule. Anterolateral corners of frontal ridges project to form acute angles. Maximum soldier head width (mean) 1.72 mm; imago with a black head capsule; pronotum brownish orange, compound eye small, nearly circular. Ocellus very small; maximum head width (mean) 1.31 mm | Rugitermes tinto |
| – | Soldier with light yellowish orange of anterior head capsule. Anterolateral corners poorly developed. Maximum soldier head width (mean) 1.17 mm; imago with a dark-castaneous head capsule; pronotum pale yellowish, compound eye large, nearly circular. Ocellus small, oval shape; head width (mean) 1.15 mm | Rugitermes ursulae sp. nov. |
Discussion
The Caribbean Region of Colombia is rich in Kalotermitidae and the tropical dry forest supports a high species diversity for this family (
The phylogenetic relationships within the Kalotermitidae are not clearly resolved among the most recently discovered genera (
Molecular markers often allow a clear separation between Rugitermes congeners (
Our study shows the importance of further surveys at isolated sites in the tropics as they continue to reveal many new species. This is also essential for phylogenetic studies to infer the evolutionary history of the Kalotermitidae, and any taxonomic lineage in a broad way.
Acknowledgments
We thank the Universidad del Norte Barranquilla, Colombia, for a research grant to RC as part of its Strategic Research Area in Biodiversity, Ecosystem Services and Human Well-being, the University of Freiburg, and COLCIENCIAS-Colfuturo for financial support. We are also grateful to the National Agency of Environmental Licenses for research permit no. 739/ANLA/MADS (8 July 2014). We thanks to John Warner for English editing, Karen Meusemann for phylogenetic analyses and discussions, and two reviewers and the editor for helpful comments and careful review of the manuscript.
References
- Bourguignon T, Lo N, Cameron SL, Šobotník J, Hayashi Y, Shigenobu S, Watanabe D, Roisin Y, Miura T, Evans TA (2015) The evolutionary history of termites as inferred from 66 mitochondrial genomes. Molecular Biology and Evolution 32: 40–621. https://doi.org/10.1093/molbev/msu308
- Bucek A, Šobotník J, He S, Shi M, McMahon D, Holmes E, Roisin Y, Lo N, Bourguignon T (2019) Evolution of Termite Symbiosis Informed by Transcriptome-Based Phylogenies. Current Biology 29: 3728–3734. https://doi.org/10.1016/j.cub.2019.08.076
- Casalla R, Scheffrahn RH, Korb J (2016a) Cryptotermes colombianus a new drywood termite and distribution record of Cryptotermes in Colombia. ZooKeys 596: 39–52. https://doi.org/10.3897/zookeys.596.9080
- Casalla R, Scheffrahn RH, Korb J (2016b) Proneotermes macondianus, a new drywood termite from Colombia and expanded distribution of Proneotermes in the Neotropics (Isoptera, Kalotermitidae). ZooKeys 623: 43–60. https://doi.org/10.3897/zookeys.623.9677
- Casalla R, Korb J (2019) Termite diversity in Neotropical dry forests of Colombia and the potential role of rainfall in structuring termite diversity. Biotropica 51: 165–177. https://doi.org/10.1111/btp.12626
- Castro D, Scheffrahn RH, Carrijo T (2018) Echinotermes biriba, a new genus and species of soldierless termite from the Colombian and Peruvian Amazon (Termitidae, Apicotermitinae). ZooKeys 748: 21–30. https://doi.org/10.3897/zookeys.748.24253
- Castro D, Scheffrahn RH (2019) A new species of Acorhinotermes Emerson, 1949 (Blattodea, Isoptera, Rhinotermitidae) from Colombia, with a key to Neotropical Rhinotermitinae species based on minor soldiers. ZooKeys 891: 61–70. https://doi.org/10.3897/zookeys.891.37523
- Emerson AE (1925) The termites of Kartabo, Bartica District, British Guiana. Zoologica (New York) 6: 291–459.
- Fuchs A, Heinze J, Reber-Funk C, Korb J (2003) Isolation and characterization of six microsatellite loci in the drywood termite Cryptotermes secundus (Kalotermitidae). Molecular Ecology Notes 3: 355–357. https://doi.org/10.1046/j.1471-8286.2003.00448.x
- Gouy M, Guindon S, Gascuel O (2010) SeaView version 4: a multiplatform graphical user interface for sequence alignment and phylogenetic tree building. Molecular Biology and Evolution 27: 221–224. https://doi.org/10.1093/molbev/msp259
- Hausberger B, Kimpel D, Neer A, Korb J (2011) Uncovering cryptic species diversity of a termite community in a West African savanna. Molecular Phylogenetics and Evolution 61: 964–969. https://doi.org/10.1016/j.ympev.2011.08.015
- Hagen H (1858) Monographie der Termiten. Linnaea Entomologica 12: [i-iii +] 4–342. [+ 459.]
- Hijmans R, Cameron S, Parra J, Jones P, Jarvis A (2005) Very high-resolution interpolated climate surfaces for global land areas. International Journal of Climatology 25: 1965–1978. https://doi.org/10.1002/joc.1276
- Holmgren N (1911) Termitenstudien. 2. Systematik der Termiten. Die Familien Mastotermitidae, Protermitidae und Mesotermitidae. Kungliga Svenska Vetenskaps-Akademiens Handlingar 46: 1–86. [+ 6 pls]
- INDERENA (1983) Acuerdo 0028 del 6 de julio 1983. Por el cual se declara área de reserva forestal protectora, la Serranía de Coraza y Montes de María (Serranía de San Jacinto), ubicada en jurisdicción de los Municipios de Toluviejo, Colosó y Chalán (Departamento de Sucre). Bogotá, Colombia, 6 pp.
- Inward D, Vogler A, Eggleton P (2007) A comprehensive phylogenetic analysis of termites (Isoptera) illuminates key aspects of their evolutionary biology. Molecular Phylogenetic and Evolution 44: 953–967. https://doi.org/10.1016/j.ympev.2007.05.014
- Kalyaanamoorthy S, Minh BQ, Wong TKF, von Haeseler A, Jermiin LS (2017) ModelFinder: fast model selection for accurate phylogenetic estimates. Nature Methods 14: 587–589. https://doi.org/10.1038/nmeth.4285
- Krishna K (1961) A generic revision and phylogenetic study of the family Kalotermitidae (Isoptera). Bulletin of the American Museum of Natural History 122: 303–408.
- Krishna K, Grimaldi DA, Krishna V, Engel MS (2013) Treatise on the Isoptera of the world. Vol. 2 Basal Families. American Museum of Natural History Bulletin 377: 201–623. https://doi.org/10.1206/377.2
- Kumar S, Stecher G, Li M, Knyaz C, Tamura K (2018) MEGA X: Molecular Evolutionary Genetics Analysis across computing platforms. Molecular Biology and Evolution 35: 1547–1549. https://doi.org/10.1093/molbev/msy096
- Light S (1932) Termites of the Marquesas Islands. Bulletin of the Bernice Pauahi Bishop Museum 98: 73–86.
- Misof B, Liu S, Meusemann K, Peters RS, Donath A, Mayer C, Frandsen PB, Ware J, Flouri T, Beutel RG, Niehuis O, Petersen M, Izquierdo-Carrasco F, Wappler T, Rust J, Aberer AJ, Aspöck U, Aspöck H, Bartel D, Blanke A, Berger S, Böhm A, Buckley TR, Calcott B, Chen J, Friedrich F, Fukui M, Fujita M, Greve C, Grobe P, Gu S, Huang Y, Jermiin LS, Kawahara AY, Krogmann L, Kubiak M, Lanfear R, Letsch H, Li Y, Li Z, Li J, Lu H, Machida R, Mashimo Y, Kapli P, McKenna DD, Meng G, Nakagaki Y, Navarrete-Heredia JL, Ott M, Ou Y, Pass G, Podsiadlowski L, Pohl H, von Reumont BM, Schütte K, Sekiya K, Shimizu S, Slipinski A, Stamatakis A, Song W, Su X, Szucsich NU, Tan M, Tan X, Tang M, Tang J, Timelthaler G, Tomizuka S, Trautwein M, Tong X, Uchifune T, Walzl MG, Wiegmann BM, Wilbrandt J, Wipfler B, Wong TKF, Wu Q, Wu G, Xie Y, Yang S, Yang Q, Yeates DK, Yoshizawa K, Zhang Q, Zhang R, Zhang W, Zhang Y, Zhao J, Zhou C, Zhou L, Ziesmann T, Zou S, Li Y, Xu X, Zhang Y, Yang H, Wang J, Wang J, Kjer KM, Zhou X (2014) Phylogenomics resolves the timing and pattern of insect evolution. Science 346: 763–767. https://doi.org/10.1126/science.1257570
- Nguyen L, Schmidt H, Haeseler A, Quang B (2015) IQ-TREE: A Fast and Effective Stochastic Algorithm for Estimating Maximum-Likelihood Phylogenies, Molecular Biology and Evolution 32: 268–274. https://doi.org/10.1093/molbev/msu300
- Oliveira G (1979) Rugitermes niger (Isoptera, Kalotermitidae), nova espécie de térmita do sul do Brasil. Dusenia 11: 9–14.
- Pattengale N, Alipour M, Bininda-Emonds O, Moret B, Stamatakis A (2009) How Many Bootstrap Replicates Are Necessary?. In: Batzoglou S. (eds) Research in Computational Molecular Biology. RECOMB 2009. Lecture Notes in Computer Science, vol 5541. Springer, Berlin. https://doi.org/10.1007/978-3-642-02008-7_13
- Pinzón O, Castro D (2018) New records of termites (Blattodea: Termitidae: Syntermitinae) from Colombia. Journal of Threatened Taxa 10: 12218–12225. https://doi.org/10.11609/jott.3909.10.9.12218-12225
- Pinzón O, Casalla R, Castro D, Vargas-Niño A (in press) Termitas de Colombia, 35 pp.
- Pizano C, García H (2014) El Bosque Seco Tropical en Colombia. Instituto de Investigación de Recursos Biológicos Alexander von Humboldt (IAvH). Bogotá, Colombia, 354 pp.
- Postle A, Scheffrahn RH (2016) A new termite (Isoptera, Termitidae, Syntermitinae, Macuxitermes) from Colombia. ZooKeys 587: 21–35. https://doi.org/10.3897/zookeys.587.7557
- Ronquist F, Huelsenbeck JP (2003) MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 19: 1572–1574. https://doi.org/10.1093/bioinformatics/btg180
- Roonwal M (1969) Measurement of termites (Isoptera) for taxonomic purposes. Journal of the Zoological Society of India 21: 9–66.
- Scheffrahn RH (2010) An extraordinary new termite (Isoptera: Termitidae: Syntermitinae: Rhynchotermes) from the pasturelands of northern Colombia. Zootaxa 2387: 63–68. https://doi.org/10.11646/zootaxa.2387.1.6
- Scheffrahn RH (2015) Global elevational, latitudinal, and climatic limits for termites and the redescription of Rugitermes laticollis Snyder (Isoptera: Kalotermitidae) from the Andean Highlands. Sociobiology 62: 426–438. https://doi.org/10.13102/sociobiology.v62i3.793
- Scheffrahn RH (2019) Expanded New World distributions of genera in the termite family Kalotermitidae. Sociobiology 66: 136–153. https://doi.org/10.13102/sociobiology.v66i1.3492
- Scheffrahn RH (2019) Rhynchotermes armatus, a new mandibulate nasute termite (Isoptera, Termitidae, Syntermitinae) from Colombia. ZooKeys 892: 135–142. https://doi.org/10.3897/zookeys.892.38743
- Scheffrahn RH, Carrijo TF (2020) Three new species of Rugitermes (Isoptera, Kalotermitidae) from Peru and Bolivia. ZooKeys 1000: 31–44. hhttps://doi.org/10.3897/zookeys.1000.59219
- Scheffrahn RH, Pinzón OP (2020) Rugitermes tinto: A new termite (Isoptera, Kalotermitidae) from the Andean region of Colombia. ZooKeys 963: 37–44. https://doi.org/10.3897/zookeys.963.55843
- Silvestri F (1901) Nota preliminare sui Termitidi sud-americani. Bollettino dei Musei di Zoologia ed Anatomia Comparata della Reale Università di Torino 16: 1–8. https://www.biodiversitylibrary.org/page/11675026
- Snyder T (1925) A new Rugitermes from Panama. Journal of the Washington Academy of Sciences 15: 197–200.
- Snyder T (1926) Five new termites from Panama and Costa Rica. Proceedings of the Entomological Society of Washington 28: 7–16.
- Snyder T (1952) A new Rugitermes from Guatemala. Proceedings of the Entomological Society of Washington 54: 303–305.
- Snyder T (1957) A new Rugitermes from Bolivia (Isoptera, Kalotermitidae). Proceedings of the Entomological Society of Washington 59: 81–82.
- Stamatakis A (2014) RAxML version 8: a tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 30: 1312–3. https://doi.org/10.1093/bioinformatics/btu033
- Thompson G, Miller L, Lenz M, Crozier R (2000) Phylogenetic analysis and trait evolution in Australian lineages of drywood termites (Isoptera, Kalotermitidae). Molecular Phylogenetics and Evolution 17: 419–429. https://doi.org/10.1006/mpev.2000.0852
Supplementary materials
Tables S1, S2
Data type: Sequence accession identifier and Measures of genetic distance
Explanation note: Table S1. GenBank accession numbers for COII gene sequences of Kalotermitidae used in this study. Table S2. Nucleotide p-distances for COII sequences between Rugitermes ursulae sp. nov. (bold) and other species belonging to Kalotermitidae. Cryptocercus punctulatus used as outgroup.
Figure S1
Data type: Multimedia: Photo
Explanation note: Biome for Rugitermes ursulae sp. nov. (Tropical dry forest, Colosó, Colombia, July 2014).
Supplementary references
Data type: References