Research Article |
|
Corresponding author: Anatoly Babenko ( lsdc@mail.ru ) Academic editor: Louis Deharveng
© 2018 Mikhail Potapov, Motohiro Hasegawa, Natalia Kuznetsova, Anatoly Babenko, Alexander Kuprin.
This is an open access article distributed under the terms of the Creative Commons Attribution License (CC BY 4.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Citation:
Potapov M, Hasegawa M, Kuznetsova N, Babenko A, Kuprin A (2018) Isotomidae of Japan and the Asiatic part of Russia. I. Folsomia ‘inoculata’ group. ZooKeys 750: 1-40. https://doi.org/10.3897/zookeys.750.22764
|
Abstract
The paper considers blind species of the genus Folsomia having two pairs of macrosetae on both meso- and metathorax and united in so-called ‘inoculata’ group, which is given a new, more laconic definition. Morphological characters important in the group’s taxonomy are discussed and a further division into four subgroups is proposed. Eight new species, i.e., F. amurica Potapov & Kuznetsova, sp. n., F. breviseta Potapov & Kuznetsova, sp. n., F. calcarea Potapov, sp. n., F. imparis Potapov & Hasegawa, sp. n., F. laconica Potapov & Kuznetsova, sp. n., F. tertia Potapov, sp. n., F. trisensilla Potapov, sp. n., and F. tubulata Potapov & Babenko, sp. n., are described. F. hidakana Uchida & Tamura and F. inoculata Stach are redescribed basing on new material, for the latter species the Stach’s individuals were also examined. A key to species of the group is given.
Keywords
α-taxonomy, Collembola , Far East of Russia, Japan, key
Introduction
The present revision is based on a vast material recently collected by the authors in various parts of the Eastern Palaearctic and older collections kindly provided by our colleagues. All used materials are deposited in the Tottori Prefectural Museum (Japan) and Moscow State Pedagogical University (Russia).
Traditional methods of morphological taxonomy were mainly used. Multi-dimensional scaling was also applied for variability analysis of widespread F. inoculata. Nine metric characters (ratios) were defined for 84 individuals from five large regions of the Palearctic (see the legend to Fig.
Abbreviations used:
Abd. abdominal segments;
alt. altitude;
Ant. antennal segments;
AO antennal organ;
bms basal ms on antennal segments;
Md, Mdl, Ml macrosetae in dorsal, dorso-lateral and lateral position;
ms micro s-seta(e) or ms-setae;
PAO postantennal organ;
s in the text and figures – macro s-setae or s-setae;
Th. thoracic segments;
Taxonomy
Remarks on Folsomia inoculata group
The group was firstly characterized by three basic characters, i.e., the presence of dorsal macrosetae on Th.II-III, posterior position of medial s-setae on abdominal segments, and the presence of ventral setae on Th.III (
Now it became clear that there are some important exceptions, namely three species with mid-tergal position of s-setae (breviseta sp. n., calcarea sp. n., and torpeda) and two species without ventral setae on Th.III (breviseta sp. n. and hidakana). Otherwise, all these species are obvious members of the same very characteristic East Asiatic group. This fact forces us to propose a new definition of the group: Folsomia with eyes absent, macrosetae on dorsum (= Md) of each Th. II and III present resulting in 22/333 formula, body shape tubular, head massive, PAO long and slender.
All species of the group also share several ordinary characters: four sublobal hairs on maxillary outer lobe, bifurcate maxillary palp, labral formula 4/5,5,4, not reduced edge of labrum, unguis without lateral or inner teeth, the absence of foil setae at the tip of abdomen. Therefore, we exclude all these characters from the species diagnoses given below.
Classification of the group
The group is not homogenous and its members can be classified by appearance into three types: ‘long-furcated’ common for Folsomia, ‘short-furcated’ slender cylindrical (‘tatarica’ subgroup), and stout with massive head (F. inoculata). Moreover, the group can be divided into four subgroups basing on well visible although not necessary evolutionary significant characters:
– ‘hidakana’ subgroup. It consists of the most primitive species having long furca, complete sets of s-setae and common (for the genus) number of ms-setae on body (43/22235, 10/100, as in Figs
– ‘macrochaetosa’ subgroup. Unlike the previous group, its species have ventral setae on Th.III (Fig.
– ‘laconica’ subgroup. Species with incomplete set of s-setae (33/22224) and ms-setae (10/000) on body (as in Figs
– ‘tatarica’ subgroup. The species are habitually specific due to slender body (Figs
Folsomia inoculata holds a unique position in the group due to s-pattern on Abd.V, chaetotaxy of furca, and specific appearance (for details see Remarks for the species).
The state of knowledge of the group
We believe that representatives of the ‘inoculata’ group were previously collected and recorded by other researchers in the eastern areas of Asia. In the associated regional papers (
Distribution and ecology
The known species of the group mostly inhabit the boreal zone of the Eastern Asia. If considering our unpublished materials from North America, one species (F. inoculata, together with its junior synonym F. ezoensis) has almost trans-Holarctic range. Subgroup of short-furcated species (‘tatarica’ sgr.) occupies areas close to the Ural Mts (excl. F. tubulata sp. n.) although does not penetrate to the main part of Europe. In North America (unpubl. material, coll. A. Fjellberg) the group is not so diverse and we have discovered only few, mostly new species. Representatives of the ‘inoculata’ group often predominate in litter of native forests and such ecological niche may be used as an additional difference from the ‘fimetaria’ group. The latter group prefers various disturbed habitats, organically enriched sites, etc. (F. candida, F. fimetaria, and F. litsteri, for example).
Key taxonomic characters of the group
Position of s-setae on tergites. The group show a high diversity of position of medial s-setae relatively to p-row – four patterns can be discriminated (Table
| Th.II–III | Abd.I–II | Abd.III | Species |
|---|---|---|---|
| in p-row | in p-row | in p-row | F. amurica sp. n., F. imparis sp. n., F. laconica sp. n., F. macrochaetosa, F. setifrontalis, F. tertia sp. n., F. trisensilla sp. n., F. baida, F. tubulata sp. n. |
| in front of p-row | in p-row | in p-row | F. bashkira, F. brevisensilla, F. hidakana, F. inoculata |
| in p-row | in p-row | in front of p-row | F. tatarica |
| in front of p-row | in front of p-row | in front of p-row | F. breviseta sp. n., F. calcarea sp. n., F. torpeda |
S-pattern on Abd.IV and V. Position and differentiation of s-setae on Abd.V is one of the keys to understanding of evolution of the genus Folsomia (
Front setae on coxa of leg I. Species of the ‘tatarica’ subgroup, F. inoculata, F. brevisensilla, F. breviseta sp. n., and F. tertia sp. n. have two such setae, F. laconica sp. n., F. trisensilla sp. n., and species of the ‘macrochaetosa’ subgroup – three, and F. hidakana – four (Fig.
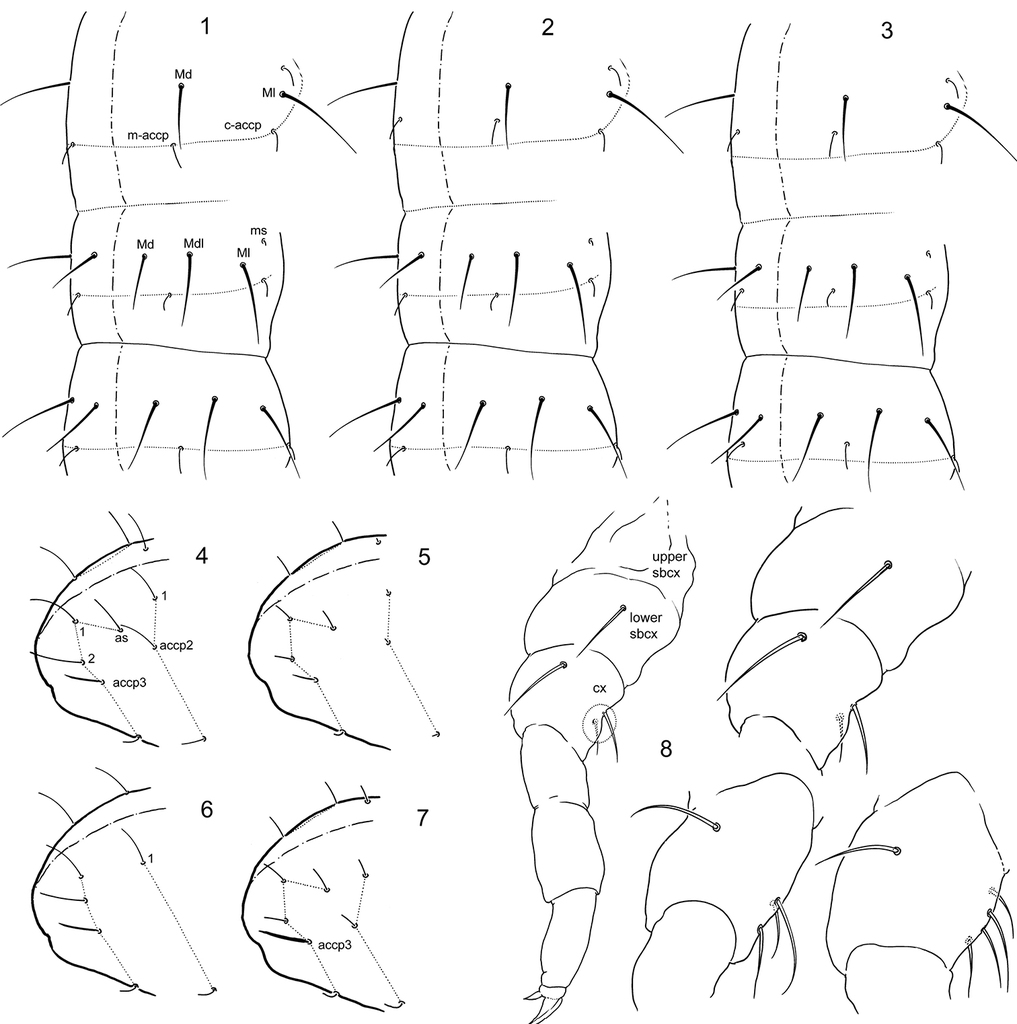
. Folsomia spp. 1–3 Variants of position of medial s-setae on Th.III, Abd.I, and Abd.II 4–7 Sensillar patterns at the end of abdomen in F. amurica sp. n. (4), F. bashkira (5), F. laconica sp. n. (6), and F. inoculata (7) 8 Position and different number (two, three, and four) of frontal setae on coxa of leg I (encircled). Abbreviations: Md, Mdl, Ml–dorsal, dorso-lateral and lateral macroseta, m-accp, c-accp–medial and corner accp-s-setae, ms–ms-seta, as–as-s-seta, accp2, accp2 –accp-s-seta, upper sbcx, lower sbcx–upper and lower subcoxae, cx–coxa.
Setae on ventrum of metathorax. The species of the ‘inoculata’ group normally have three ventral setae on each side of Th.III, one of which is long and two are short (Fig.
Characters of lower taxonomic value
– PAO in all ‘inoculata’ species is slender, with parallel edges, and more than 1.3 times longer than width of Ant.I (Figs
– All species have three basal ms-setae on Ant.I: one ventral and two dorsal. Two dorsal bms-setae are arranged in a longitudinal line, proximal bms is longer (Figs
– The members of ‘hidakana’, ‘macrochaetosa’, and ‘laconica’ subgroups have minute subapical setae on posterior side of dens. The size of the seta varies depending on specimens and often hardly detectable. Small wrinkle in which this seta set in is always visible.

Folsomia spp. 9–10 Ventrum of Th.II, Th.III, and Abd.I in F. imparis sp. n. (9), setae on Th.III present and F. hidakana (10), setae absent 11–12, 14 F. breviseta sp. n., furca, lateral view (11), appearance (12), PAO and Ant.I (14) 13 Appearance of F. hidakana. Abbreviation: vm–ventro-medial setae.
Descriptions of species
Subgroup ‘hidakana’
Folsomia breviseta , sp. n.
Type material
Holotype, female, Russia, NE Yakutia, Middle Indigirka, near Ust-Nera, 700 m alt., larch forest with thick lichen cover, litter, 22.vii.1992. Ten paratypes from the same biotope, five paratypes from mountain tundra (1500 m alt.) with bushes of Betula nana and one paratype from mosses in stony niche on northern slope (1600 m alt.) in the same location. All collected by N. Kuznetsova and M. Potapov. Deposited in
Diagnosis
Blind. Dorsal macrosetae (Md) present on both Th.II and Th.III. Sensillary formula complete (43/22235; 10/100). Medial s-setae on body tergites short, set in anterior position. Macrosetae short. Ventral setae on Th.III absent. Anterior side of manubrium normally with 2+2 setae, dens with 18–22 anterior setae. Mucro bidentate.
Description
Body size from 1.0 (one adult male) to 1.7 mm. Body shape relatively tubular, general appearance not typical of ‘inoculata’ group due to short macrosetae (Fig.
Common setae short. Sensillary formula as 43/22235 (s) and 10/100 (ms). S-setae short, four s-setae on dorsal side of Abd.V longer. Medial s-setae on Th.II–Abd.III situated in front position, on Abd.I–III between Md and Mdl (Figs
Empodial appendage as long as 0.5–0.6 unguis. Tibiotarsi with 24–27 setae on legs I–II, and 28–32 on leg III. Upper and lower subcoxae of legs I–III with 0,1/3,6–8/5–7,7–9 setae, respectively. Coxae of leg I with two front setae. Ventral tube with 4+4 latero-distal and 5–7 posterior setae (two in distal transversal row and 3–5 in more proximal position), anteriorly without setae. Tenaculum with 4+4 teeth and a seta. Anterior furcal subcoxae with 12–15, posterior one with six setae. Anterior side of manubrium normally with 2+2 setae at distal edge, arranged in two longitudinal lines (Fig.

F. breviseta sp. n. 15Ant.III (area of common position of lateral s-setae marked) 16–17 Position of macrosetae, setae of p-row, and s-setae on posterior (16) and anterior (17) half of corpus 18 Chaetotaxy of Abd. IV–VI 19–20 Chaetotaxy of anterior side of manubrium, variations. bms–basal ms.
Remarks
Folsomia breviseta sp. n. combines several characters rare for the group: short macrosetae and s-setae, anterior position of medial s-setae on body tergites, the absence of ventral setae on Th.III. A loss of lateral s-setae on Ant.III is a very peculiar character while the nature of this character is not easy to interpret since all species of the family Isotomidae, if not all Collembola, have these s-setae. The absence of s-setae was confirmed in all available specimens. Beyond ‘inoculata’ group several blind species with two pairs of setae on manubrium can be confused with F. breviseta sp. n., for example, F. bisetosa Gisin, F. cephalota Bu et al., and F. sensibilis Kseneman. All these species belong to other groups and therefore differ essentially by characters of high rank.
Distribution and ecology
The species is known only from the type locality where it inhabits mountain forests and tundra.
Derivatio nominis
The new species name reflects short setae on body.
Folsomia hidakana
Material
Japan, Honshu Island, Nobeyama, Nagano, 10.x.2012, coll. M. Hasegawa; Kitaibaraki, 2.xi.2011, coll. M. Hasegawa; Hokkaido Island. Shore of Harutori Lake, Kushiro city, 14.v.2014 and 19.viii.2014, coll. Y. Suma; Shiretoko Peninsula, surroundings of Utoro, litter of mixed forest, 90 m alt., 20.viii.2016, 44.1006°N, 145.0584°E, coll. M. Potapov and N. Kuznetsova; Shiretoko Peninsula, trail to Rausu Mount, ~200 m alt., 29.ix.2013, coll. R. Kitagawa and S. Fujii; ibidem, trail to Rausu Mount, ~1000 m alt, oak litter, coll. M. Potapov and N. Kuznetsova.
North Korea. Hamgyong-namdo Province (= South Hamgyong), valley N from Song-riong, 03.vi.1987, and SW from Tanchon, 30.v.1987, coll. A. Szeptycki.
Far East of Russia, Primorsky Krai, Lazovsky District, nearby Preobrazheniye, litter and rotten wood in deciduous and coniferous forests, 400–700 m alt., 21.ix.2011, coll. M. Potapov, Y. Bu, H. Chen-Wang; Khasansky District, Peschany Peninsula, near Beregovoye, litter under dogrose, 09.ix.2004, coll. M. Potapov, L. Deharveng, R. Pomorski, and A. Bedos, Khasansky District, ~15 km S Kraskino, Mramorny Cape, oak wood litter and soil under coastal reed, 28.ix.2004, coll. M. Potapov, L. Deharveng, R. Pomorski, and A. Bedos; Khasansky District, ~15 km W Kraskino, Mayachnoye (Chertova Gorka), forest litter, 28.ix.2004, coll. M. Potapov, L. Deharveng, R. Pomorski, and A. Bedos; Khasansky District, Krabbe Peninsula, Astafyeva Cape, deciduous litter, v.2007, coll. E. Sokolova; Khasansky District, “Kedrovaya Pad”, mixed forest, litter, 29.vii.2016, coll. M. Potapov and N. Kuznetsova; ibidem, coniferous and deciduos litter, 29.ix.2004, coll. M. Potapov, L. Deharveng, R. Pomorski, and A. Bedos; ibidem, 03.x.2009, coll. O. Smirnova, ibidem, v.2015, coll. A. Matalin; Khasansky District, near Barabash, oak wood on slope, litter, 27.ix.2004, coll. M. Potapov, L. Deharveng, R. Pomorski, and A. Bedos; Partizansky District, vicinities of Ekaterinovka, Chondalaz (= Lazovy) Range, oak litter, 26.ix.2004, coll. M. Potapov, L. Deharveng, R. Pomorski, and A. Bedos; Ussuriyski District, Ussuriyski Reserve, decaying wood, 5.x.2004, coll. M. Potapov, L. Deharveng, R. Pomorski, and A. Bedos; Shkotovsky District, Khualaza Mount, 2.x.2004, coll. R. Pomorski; between Vladivostok and Artem, botanical garden, litter of mixed forest, ix.2012 and 04.x.2009, coll. O. Smirnova; Terneysky District, Sikhote-Alimski Reserve, Kabany station, forest litter, 08.viii.2017, coll. N. Kuznetsova, A. Geras’kina, A. Kuprin; Kavalerovski District, road Kavalerovo-Dal’negorsk, 44.3844°N, 135.3639°E, mossy larch forest with Rhododendron, 09.viii.2017, coll. N. Kuznetsova, A. Geras’kina, A. Kuprin; Khabarovsky Krai, Vaninsky District, nearby Datta, coastal larch-wood, 28.ix.2011, coll. M. Potapov; Vaninsky District, five km N Vysokogorny, valley of Mulinka River, larch-forest litter, ~ 600 m alt., 29.ix.2011, coll. M. Potapov.
Description
Body size from 1.2 to 1.7 mm. Body shape as common for the group, not slender (Fig.

F. hidakana 21–22 Position of macrosetae, setae of p-row, and s-setae on anterior (21) and posterior (22) half of corpus 23Ant.I (S Primorye: Far East of Russia) 24PAO and Ant.I (Honshu: Japan) 25Ant.III 26–27 Manubrium, anterior (26) and posterior (27) views. Abbreviations: bms–basal ms, ls–lateral s.
Common setae middle-sized. Setae covering polychaetotic: Abd.IV with 7–9 p-setae between medial accp-s, Abd.V with m1-setae (marked on Fig.
Empodial appendage approximately half as long as unguis. All tibiotarsi with many additional setae: 29–33 on legs I–II, ~38–42 on leg III. Upper and lower subcoxae of legs I–III with 0,1/4–6,8–11/8–12,8–11 setae, respectively. Coxae of leg I with four (rarely five) front setae. Ventral tube with 5–6+5–6 latero-distal and 6–8 posterior setae, anteriorly without setae. Tenaculum with 4+4 teeth and a seta. Anterior furcal subcoxae with 11–14, posterior one with four setae. Anterior side of manubrium with 4–6+4–6 pair setae and usually with two unpaired axial setae (Fig.
Remarks
Our specimens fit to the original description of F. hidakana in all significant features.

Folsomia spp. 28 F. hidakana, end of abdomen, dorsal view 29–31 F. amurica sp. n., end of abdomen, specimens from Amurskaya Region, dorsal (29) and lateral views (31), specimen from Inner Mongolia Province, lateral view (30) 32 F. imparis sp. n., end of abdomen 33–35 F. amurica sp. n., chaetotaxy of anterior side of manubrium, variations.
Specimens collected in Japan, both in Honshu and Hokkaido, differ from specimens from Russia by three (vs two) s-setae on Ant.I. In Japanese populations the individuals with two s rarely occur so we keep both variants within diagnosis of F. hidakana.
Distribution and ecology
Species was described from Hokkaido (Hidaka-Mombetsu) and subsequently listed in catalogues of Japanese Collembola (
As a whole, distributional range of the species appears to cover southern area of the Russian Far East, North Korea, and northern half of Japan (Fig.
Subgroup ‘macrochaetosa’
Folsomia amurica , sp. n.
Type material
Holotype, female, Far East of Russia, Amurskaya Region, Zeysky Reserve, ~50 km W Zeya, near “Gol’tsy” station, subalpine dwarf wood (Pinus pumila), ~ 1300 m alt., coniferous litter, 20.viii.2014., Ten paratypes from the same biotope and five paratypes from the same location, litter of mixed forest at 700 m alt., 21.viii.2014, coll. M. Potapov and N. Kuznetsova. Deposited in
Other material
Far East of Russia, Amurskaya Region, various biotopes nearby type locality: litter and rotten wood in mountain tundra, spruce and larch forests at different altitudes (from 400 to 1400 m alt.); Amurskaya Region, ~ three km W Arkhara, oak-forest, litter, 17.viii.2014, coll. M. Potapov and N. Kuznetsova.
China, Inner Mongolia Province, Da Hinngan Ling Mts, ~ 25 km W BaLin, young tussocky wet birch forest, litter, 12.viii.2014, coll. M. Potapov.
Diagnosis
Blind. Dorsal macrosetae (Md) present on both Th.II and Th.III. Sensillary formula complete (43/22235; 10/100). Medial s-setae on body tergites long, set in p-row. Ventral setae on Th.III present. Manubrium on anterior side with 4–6+4–6 setae, no unpaired axial setae, dens with 23–27 anterior setae. Mucro bidentate.
Description
Body size from 1.0 to 1.5 mm. Body without pigmentation, relatively tubular (Fig.
Common setae long. Sensillary formula as 43/22235 (s), 10/100 (ms) (Figs
Unguis of normal shape, without lateral and inner teeth. Empodial appendage as long as 0.5–0.6 unguis. All tibiotarsi with additional setae: 26–29 on legs I–II and > 35 on leg III. Upper and lower subcoxae of legs I–III with 0,1/3,8–9/5–6,8–10 setae, respectively. Coxae of leg I with three front setae. Tibiotarsal tenent setae pointed, some setae on distal half of tibiotarsi thickened. Ventral tube with 4+4 latero-distal and 7–8 posterior setae (four in distal transversal row and 3–4 in more proximal position), anteriorly without setae. Tenaculum with 4+4 teeth and a seta. Anterior furcal subcoxae with 8–9, posterior one with five setae. Anterior side of manubrium with 4–6+4–6 setae, their position vary, unpaired setae absent (Figs

Remarks
The species most resembles F. macrochaetosa and F. imparis sp. n. and is characterized by the absence of unpaired setae on anterior side of manubrium, short accp3-s-setae and long p1 setae on Abd.V (Table
Distribution and ecology
Known from three neighboring localities of inner part of East Asia (Fig.
Derivatio nominis
The species is common in areas around Amur River lowlands.
Folsomia brevisensilla
Material
Far East of Russia, Magadanskaya Region, Ten’kinsky District, village Kulu, 04.ix.1995, coll. S. Bukhkalo.
Remarks
The species resembles F. inoculata sharing with the latter species a middle-sized furca, short s-setae on body, their position on tergites, and undifferentiated ventral setae on Th.III. Nevertheless, F. brevisensilla does not possess several unique characteristics of the latter species, e.g., large and tubular accp3-s on Abd.V and the absence of subapical seta on posterior side of dens.
Distribution
It is the most northern species of the ‘inoculata’ group since known so far only in the basin of Kolyma River (NE Asiatic part of Russia).
Folsomia imparis , sp. n.
Type material
Holotype, female, Japan, Hokkaido Island, Shiretoko Peninsula, trail to Mont. Rausu, deciduous forest (Acer, Quercus, Sorbus, Magnolia, Daphniphillum macropodium), litter, 19.viii.2016, 354 m alt., 44.1083°N, 145.0893°E, coll. M. Potapov and N. Kuznetsova. Paratypes, 12 specimens from the same location; three specimens from Russia, Far East, S Primorye, Ussuriysky Reserve, Komarovskoye Forest District, Turova Nipple, Khripunovsky Pass, mixed forest with Pinus koraiensis on slope, rotten wood, 22.vii.2016, coll. M. Potapov and N. Kuznetsova; four specimens from same region, but Shkotovsky District, trail to Mont. Khualaza, deciduous forest, rotten wood and litter, 21.vii.2016, coll. M. Potapov and N. Kuznetsova. The material from Japan and Russia is deposited in
Other material
Far East of Russia, Primorsky Krai, Bikin River near confluence to Amba River, mixed forest litter, 29.ix.2009, coll. O. Smirnova; Ussuriyski District, Ussuriyski Reserve, decaying wood, 5.x.2004, coll. M. Potapov, L. Deharveng, R. Pomorski, and A. Bedos; Shkotovsky District, trail to Khualaza Mount, rotten wood, 21.vii.2016, coll. M. Potapov and N. Kuznetsova; Partizansky District, vicinities of Ekaterinovka, Chondalaz (= Lazovy) Range, oak litter, 26.ix.2004, coll. M. Potapov, L. Deharveng, R. Pomorski, and A. Bedos; Khasansky District, vicinities of Barabash, oak litter, 200–400 m alt., 27.ix.2004, coll. M. Potapov; Sakhalin, Kholmsky District, South Kamysh Ridge, Spamberg Mt., moss in mixed and coniferous forests, 14–15.vi.2017, coll. A. Kuprin; Korsakovsky District, vicinities of Korsakov, forest litter, 16.vi.2017, coll. A. Kuprin; Khabarovsky Krai, suburbs of Khabarovsk, Voronez highlands, leaf litter near river bank. 26.iv.2010, coll. M.Potapov; Verkhnebureinsky District, western part of Badjal Range, upper flow of Irungda River (tributory of Amgun’ River), 1900 m alt., subalpine litter and moss, 23.vi.2014, coll. A. Brinev.
North Korea. Hamgyong-namdo Province (= South Hamgyong), SW from Tanchon, 30.v.1987, coll. A. Szeptycki; Yanggang-do Province, Rimjong-su Waterfall, litter, 7.vii.1985, coll. A. Szeptycki.
Material of F. sp. aff. imparis. Khabarovsky Krai (western part), Bureyskoye Reservoir, Nizny Mel’gin Bay, 50.5539°N, 131.3970°E, 12.ix.2009, coll. M. Babykina; Amurskaya Region, ~3 km N Zeya, oak litter, 22.viii.2014, coll. M. Potapov and N. Kuznetsova.
Diagnosis
Blind. Dorsal macrosetae (Md) present on both Th.II and Th.III. Sensillary formula complete (43/22235; 10/100). Medial s-setae on body tergites long, set in p-row. Ventral setae on Th.III present. Manubrium on anterior side with 3–5+3–5 paired and 2–3 unpaired axial setae, dens with 23–27 anterior setae. Mucro bidentate.
Description
Body size from 0.9 to 1.4 mm. Body without pigmentation, its shape as in F. amurica sp. n. Cuticle with fine hexagonal primary granulation (“smooth”). Ocelli absent. PAO slender, constricted, 1.4–1.7 as long as width of Ant.I and 1.7–1.9 as long as inner unguis length. Labium with five usual papillae (A–E), guard setae e7 absent, three proximal and four basomedian setae. Ventral side of a head with 4+4 postlabial setae. Ant.I with 15–17 common setae, two ventral s-setae (s) and three bms, one long (inseparable from common setae) and two short, Ant.II with three bms and one latero-distal s, Ant.III with one bms and with five distal s (including one lateral), without additional s-setae (Fig.
Common setae long and sparse. Sensillary formula as 43/22235 (s), 10/100 (ms). Tergal s-setae thin and long. Medial s-setae on Th.II–Abd.III situated in posterior position, on Abd.I–III between Md and Mdl. Abd.V with five s-setae arranged as four ones (as, accp1, accp2, accp3), long and slender, and one latero-ventral, short (‘4+1’ pattern) (Figs
Unguis of normal shape, without lateral and inner teeth. Empodial appendage as long as 0.4–0.5 unguis. Tibiotarsi with 26–28 setae on legs I–II and 33–37 on leg III. Upper and lower subcoxae of legs I–III with 0,1/3,~7/5–6,7–8 setae, respectively. Coxae of leg I with three front setae. Tibiotarsal tenent setae pointed, some setae on distal half of tibiotarsi thickened. Ventral tube with 4+4 latero-distal and seven posterior setae (four in distal transversal row and three in more proximal position), anteriorly without setae. Tenaculum with 4+4 teeth and a seta. Anterior furcal subcoxae with 10–12, posterior one with five setae. Anterior side of manubrium with 3–5+3–5 pair setae, and 2–3 (rarely one) axial unpaired setae (Figs

F. imparis sp. n. (40–46) and F. macrochaetosa (47–49) 46 End of abdomen, lateral view 41 Furca, lateral view (unpaired setae marked with grey) 42 Anterior side of dens 43–45 Chaetotaxy of anterior side of furca, variations 46Ant.III 47 End of abdomen 48 Furca, anterior view 49 Dens, posterior view. Notation of setae of posterior and lateral sides of manubrium: ap, M1, ml1, L1, l1, l2; LB–latero-basal setae, C–setae of central area.
Remarks
Folsomia imparis sp. n. is well defined by the presence of unpaired setae on anterior side of manubrium shared only with F. hidakana (belongs to another subgroup) and F. setifrontalis (has anterior setae on ventral tube). Main differences from the other species of the subgroup ‘macrochaetosa’ are shown in Table
A typical form, called preliminary as f. 1 (Fig.
Additionally, a closely related form was also recorded in two more eastern localities (see the Material part, F. sp. aff. imparis). It differs from all species listed in Table
Distribution and ecology
The species is widely distributed in Far East of Russia, Korea and in the most northern part of Japan (Fig.
Derivatio nominis
The species has unpaired setae on anterior side of manubrium.
Folsomia macrochaetosa
Type Material
Three paratypes labelled as “Magadanskaya Region, vicinities of Magadan, Snow Valley, dwarf-wood belt. 18.ix.1974. coll. Berman.” Kept in Zoological Institute (S.-Petersburg).
Other material
Far East of Russia, Khabarovsky Krai, Vicinities of Nikolaevsk-na-Amure, lower flow of Amgun’ River, larch and birch forests, 1.vii.1990, coll. N. Ryabinin; Primorsky Krai, Ussuriysky Reserve, Komarovskoye Forest District, mixed forest, 22.vii.2016, coll. M. Potapov and N. Kuznetsova; Terneysky District, Sikhote-Alinski Reserve, Kabany station, hardwood with Rhododendron, litter, 08.viii.2017, coll. N. Kuznetsova, A. Geras’kina, A. Kuprin; South Kuril Islands, Kunashir Isl., Krugly Cape and 5 km E Yu-Kuril’sk, viii–ix.1997, coll. Y. Marusik.
South Korea, Gangwon-do, Palsan Mt., mixed forest in foothills, litter, 09.ix.2017, coll. A. Kuprin.
Description
Body size from 0.9 to 1.5 mm. Body without pigmentation, cuticle with fine primary granulation. Ocelli absent. PAO slender, constricted, 1.4–1.6 as long as width of Ant.I and 1.4–2.2 as long as inner unguis length. Labium with five usual papillae (A–E), guard setae e7 absent, three proximal and four basomedian setae. Ventral side of a head with 4+4 postlabial setae. Ant.I with 15–16 common setae, s-setae of antennae as in F. imparis sp. n. Organite stick-like, small.
Common setae long. Sensillary formula as 43/22235 (s), 10/100 (ms). Tergal s-setae long, medial ones on Th.II–Abd.III situated in posterior position. Abd.V with five s-setae arranged as four ones (as, accp1, accp2, accp3), long and slender, and one latero-ventral, short (‘4+1’ pattern), accp3 s-setae somewhat shorter than accp2 (accp2 : accp3 = 1.1–1.5). p1-setae on Abd.V long (Fig.
Unguis of normal shape. Empodial appendage as long as 0.50–0.55 of unguis. Chaetotaxy of tibiotarsi and subcoxae similar to F. amurica sp. n. and F. imparis sp. n. Number of front setae on coxae of leg I varies (2–3), variant with two setae common for specimens from Magadan and Kunashir. Ventral tube with 4+4 latero-distal and 6–8 posterior setae, anteriorly without setae. Tenaculum with 4+4 teeth and a seta. Anterior furcal subcoxae with 11–12, posterior one with five setae. Anterior side of manubrium with 2–3+2–3 pairs of setae (2+2 usually in males), without axial unpaired setae (Figs
Remarks
F. amurica sp. n., F. imparis sp. n., F. macrochaetosa, and F. setifrontalis combine a group of species with long macrosetae and furca, sparse setae covering, and posterior position of median s-setae on all tergites. The differences between the species of this subgroup are shown in Table
Differentiated characters of three species of the ‘macrochaetosa’ subgroup.
| Species | Abd.V : accp3-s | Abd.V : a1:p1 | Paired setae on manubrium | Unpaired setae on manubrium |
|---|---|---|---|---|
| F. amurica sp. n. | short | 0.9–1.1 | 4–6+4–6 | absent |
| F. imparis sp. n. | long | f.1: 1.4–2.3 f.2: 1.0–1.2 | 3–5+3–5 | 2–3 (1) |
| F. macrochaetosa | long | 1.1–1.3 | 2(3)+2(3) | absent |
| F. setifrontalis | long | 1.6–2.3 | 3–5+3–5 | 0–2 |
Distribution
Scattered records all over the coastal areas from Magadan (Russian Far East) to South Korea (Fig.
Folsomia setifrontalis
Material
Far East of Russia, Primorsky Krai, Partizansky District, vicinities of Ekaterinovka, Chondalaz (=Lazovy) Range, oak litter, 26.ix.2004, coll. M. Potapov, L. Deharveng, R. Pomorski, and A. Bedos; Shkotovsky District, vicinities of Anisimovka and near trail to Khualaza Mount, forest litter, 10-12.ix.2001, coll. M. Potapov, Y. Bu and H. Cheng-Wang; Terneysky District, Sikhote-Alimski Reserve, Blagodatny station, oak wood on slope, rotten wood, 07.viii.2017, coll. N. Kuznetsova, A. Geras’kina, A. Kuprin.
Remarks
Folsomia setifrontalis is sharply defined by the presence of anterior setae on ventral tube that is a unique character for the genus. Populations from Primorsky Krai differ from the type specimens (South Kuril Islands) by the presence of unpaired setae on manubrium and longer accp3-s on Abd.V. Considering this variability, a wider diagnosis is proposed for the species. So far, the chaetotaxy of ventral tube remain a key characteristic of this species.
Distribution and ecology
Less common than the sympatric F. imparis sp. n. and F. macrochaetosa. Rare records in forest litter of southern part of Far East of Russia.
Subgroup ‘laconica’
Folsomia laconica , sp. n.
Type material
Holotype, female, Far East of Russia, Amurskaya Region, Khingansky Reserve, near (~6 km W) Kundur, valley of Karapcha River, northern steep slope, mixed forest with Abies, litter, 19.viii.2014, coll. M. Potapov and N. Kuznetsova. 15 paratypes from the same location and ten paratypes from Khingansky Reserve, ~10 km E Uril, coniferous forest (Pinus koraiensis, Abies, Picea), 7.x.2009, coll. M. Babykina. Deposited in
Other material
Amurskaya Region, Khingansky Reserve, ~15–20 km SE Uril, oak wood litter, 8.x.2009, coll. M. Babykina. North Korea, Yanggang-do Province, ~1.5 km SE Mupho, litter under Rhododendron and Alnus, 5.vii.1985, coll. A. Szeptycki.
Diagnosis
Blind. Dorsal macrosetae (Md) present on both Th.II and Th.III. Sensillary formula incomplete (33/22224; 10/000). Medial s-setae on body tergites long, set in p-row. Ventral setae on Th.III present. Anterior side of manubrium with 3+3 setae, no unpaired axial setae present. Dens with 17–20 anterior setae, its posterior side with three setae in basal part. Mucro bidentate.
Description
Body size approximately 1.3 mm (Fig.
Common setae sparse, macrosetae long (Fig.
Unguis of normal shape, without lateral and inner teeth. Empodial appendage as long as ~0.5 of unguis. All tibiotarsi with additional setae: 28–29 on legs I–II and >35 on leg III. Upper and lower subcoxae of legs I–III with 0,1/3,7/6,8 setae, respectively. Coxae of leg I with three front setae. Tibiotarsal tenent setae pointed, few setae on distal half of tibiotarsi III slightly thickened. Ventral tube with 4+4 latero-distal and 6–7 posterior setae (4 in distal transversal row), anteriorly without setae. Tenaculum with 4+4 teeth and a seta. Anterior furcal subcoxae with 12–14, posterior one with five setae. Anterior side of manubrium with 3+3 setae (Fig.
Remarks
Folsomia laconica sp. n. most resembles F. trisensilla sp. n. but differs having three (vs four) basal setae on posterior side of dens) (Figs


Distribution and ecology
Known from three locations in Russian Far East and North Korea (Fig.
Derivatio nominis
The species shows the most laconic chaetotaxy of p-row on Abd.V.
Folsomia trisensilla , sp. n.
Type material
Holotype, female, Far East of Russia, Khabarovsky Krai, Vaninsky District, 10 km N Vysokogorny, valley of Mulinka River, closed Picea and Abies forest on NE slope, litter, ~ 750 m alt., 29.ix.2011, coll. M. Potapov. Ten paratypes from the same location and six paratypes from the nearly same location, in litter of coniferous forests at different altitudes (600 and 900 m alt.), 29.ix.2011, coll. M. Potapov. Deposited in
Other material
Far East of Russia, Khabarovsky Krai. Different sites of type locality, litter and rotten wood, 29.ix.2011, coll. M. Potapov; Kamchatka. Nearby Anavgai and Esso settlements, different larch forests, litter and rotten wood, ~ 500 m alt., 4.vii.2012, coll. M. Potapov; Primorsky Krai. Terneysky District, Sikhote-Alimski Reserve, Kabany station, forest litter, 08.viii.2017, coll. N. Kuznetsova, A. Geras’kina, A. Kuprin; East Siberia: Chitinskaya Region, near foothills of Daursky Range, valley of Ilya River, ~ three km station Ara-Ilya, 50.9253°N, 113.1783°E, 887 m alt., mixed forest with Betula and Larix, 11.vii.2014, coll. A. Gulgenova.
Diagnosis
Blind. Dorsal macrosetae (Md) present on both Th.II and Th.III. Sensillary formula incomplete (33/22224; 10/000). Medial s-setae on body tergites long, set in p-row. Ventral setae on Th.III present. Anterior side of manubrium with 3+3 paired setae, no unpaired axial setae present. Dens with 12–16 anterior setae, its posterior side with four setae in basal part. Mucro bidentate.
Description
Body size from 1.0 to 1.3 mm, rather tubular (Fig.
Common setae long. Sensillary formula as 33/22224 (s), three s-setae lost (as described for the ‘laconica’ subgroup (Figs
Empodial appendage as long as 0.5–0.6 of unguis. All tibiotarsi with additional setae: 24–26 on legs I–II and 30–35 on leg III. Upper and lower subcoxae of legs I–III with 0,1/3,7–8/6–7,7–8 setae, respectively. Coxae of leg I with three (rarely two on one side) front setae. Tibiotarsal tenent setae pointed, some setae of distal whorl of tibiotarsi thickened. Ventral tube with 4+4 latero-distal and 7–8 posterior setae (four in distal transversal row and 3–4 in more proximal position), anteriorly without setae. Tenaculum with 4+4 teeth and a seta. Anterior furcal subcoxae with 8–10, posterior one with four setae. Anterior side of manubrium with 3+3 setae (rarely 2+3) (Fig.

Remarks
Two setae at the middle of posterior side of dens are always absent in the type populations (Fig.
An odd population was found near Uril (Amurskaya Region) differing from the typical ones by having 4+4 (vs 3+3) anterior setae on manubrium (left out of diagnosis of the new species so far).
For difference between F. trisensilla sp. n. and F. laconica sp. n. see Remarks to the latter.
Distribution and ecology
Known from forest litter in three localities in Eastern Asia (Fig.
Derivatio nominis
The new species name reflects the presence of three (vs four, as common for the genus) s-setae on dorsal side of Abd.V that is characteristic of the ‘laconica’ subgroup.
Folsomia tertia , sp. n.
Type material
Holotype, female, Far East of Russia, Khabarovsky Krai, vicinities of Khabarovsk, Bol’shoy Khekhtsyr Range, ~10 km N Korfovsky, mixed forest litter, 28.vi.2007, coll. E. Sokolova. Ten paratypes from the same location. Deposited in
Diagnosis
Blind. Dorsal macrosetae (Md) present on both Th.II and Th.III. Sensillary formula incomplete (33/22224; 10/000). Medial s-setae on body tergites long, set in p-row. Ventral setae on Th.III present. Anterior side of manubrium with 2+2 setae, no unpaired axial setae present. Dens with 13–16 anterior setae, its posterior side with four setae in basal part. Mucro bidentate.
Description
Body size from 0.9 to 1.2 mm, shape of corpus rather tubular, slender. Without pigmentation. Cuticle with fine hexagonal primary granulation (“smooth”). PAO slender, insignificantly constricted, 1.6–1.8 as long as width of Ant.I and 2.1–2.2 as long as inner unguis length. Labium complete, guard setae e7 present, three proximal and four basomedian setae. Ventral side of head with 4+4 postlabial setae. Ant.I with 14–15 common setae, two ventral s-setae (with one thick) and three basal micro s-setae (bms): two dorsal (short and long) and one ventral. Ant.II with three bms and one latero-distal s, Ant.III with one bms and with five distal s (including one lateral), without additional s-setae. Organite normal, small.
Sensillary formula as 33/22224 (s), three s-setae lost (as in other species of the subgroup, Fig.
Empodial appendage as long as 0.55–0.60 of unguis. Tibiotarsi with few additional setae: 22–24 on legs I–II and ~29 on leg III. Upper and lower subcoxae of legs I–III with 0,1/3,7/3–5,~8 setae, respectively. Coxae of leg I with two front setae. Tibiotarsal tenent setae pointed, few setae of whorl of tibiotarsi insignificantly thickened. Ventral tube with 4+4 latero-distal and 5–7 posterior setae (four in distal transversal row and 1–3 in more proximal position), anteriorly without setae. Tenaculum with 4+4 teeth and a seta. Anterior furcal subcoxae with 7–9, posterior one with 5–6 setae. Anterior side of manubrium with 2+2 setae (Fig.

F. tertia sp. n. 65 Position of macrosetae, setae of p-row, and s-setae on corpus 66 End of abdomen 67 Posterior side of manubrium and genital area in adult female 68 Furca, lateral view. Notation of setae of posterior and lateral sides of manubrium: ap, M1, ml1, M2, l1; LB–latero-basal setae, C–setae of central area.
Remarks
The new species belongs to the ‘laconica’ subgroup due to incomplete s-set on body. It differs from two other members, F. laconica sp. n. and F. trisensilla sp. n., by reduced chaetotaxy on anterior (2+2 instead of 3–4+3–4 setae) and posterior sides (1+1 vs 2+2 lateral setae) of manubrium. F. tertia sp. n. has the shortest macrosetae among species of the subgroup and somewhat resembles members of the ‘tatarica’ subgroup.
Distribution
Known only from type locality.
Derivatio nominis
The new species is the third (tertius in Latin) species of the ‘laconica’ subgroup.
Subgroup ‘tatarica’
Folsomia calcarea , sp. n.
Type material
Holotype, female, Russia, European part, Samarskaya District, Samarskaya Luka National Park, calcareous stone-pit, soil, 02.v.2010, coll. Y. Shveenkova. Six paratypes from the same location and maple forest nearby. Deposited in
Diagnosis
Blind. Body slender. Dorsal macrosetae (Md) present on both Th.II and Th.III. Sensillary formula incomplete (43/22235; 10/000). Medial s-setae on body tergites in anterior position. Th.III with 1+1 ventral setae. Anterior side of manubrium with 1+1 setae, dens with 6–7 anterior setae. Mucro bidentate.
Description
Body size near 1.1 mm (for the largest subadult female). Body slender, tubular (Fig.
Common setae short. Sensillary formula as 43/22235 (s), 10/000 (ms) (Fig.
Unguis of normal shape, without lateral and inner teeth. Empodial appendage as long as 0.4–0.5 of unguis. Tibiotarsi with few setae: 21–22 on legs I–II and 24–27 on leg III. Upper and lower subcoxae of legs I–III with 0,1/3,5–7/4–5,6–6 setae, respectively. Coxae of leg I with two front setae. Ventral tube with 4+4 latero-distal and 4–5 posterior setae (four in distal transversal row), anteriorly without setae. Tenaculum with 4+4 teeth and a seta. Anterior furcal subcoxae with 3–4, posterior one with three setae. Anterior side of manubrium with 1+1 setae (Fig.

Remarks
Two key characters, short furca and anterior position of medial s-setae on body tergites, are shared with only F. torpeda. F. calcarea sp. n. differs by ms-setae on Abd.I missing (vs present in F. torpeda) and fewer number of setae on posterior side of manubrium. The absence of lateral s on Ant.III is shared only with F. breviseta sp. n. but value of this characteristic requires further study.
Distribution and ecology
Known only from the type locality. The species possibly belongs to calciphilous fauna.
Derivatio nominis
The species was recorded in calcareous soil.
Folsomia tubulata , sp. n.
Type material
Holotype, adult male, Russia, East Siberia, Yakutia (Sakha Republic), Lensky District, Chayandinsky allotment, July of 2011, coll. V. Boeskorov. Two paratypes from the same location. Deposited in
Diagnosis
Blind. Body slender. Dorsal macrosetae (Md) present on both Th.II and Th.III. Sensillary formula complete (43/22235; 10/100). Medial s-setae on body tergites set in p-row. Th.III with 2+2 ventral setae. Anterior side of manubrium with 1+1 setae, dens with 5–7 anterior setae. Mucro bidentate.
Description
Body size 0.8–0.9 mm. Body slender, tubular (Fig.
Common setae short. Sensillary formula as 43/22235 (s). Medial s-setae on Th.II–Abd.III situated in p-row, on Abd.I–III between Md and Mdl (Figs
Empodial appendage as long as 0.4–0.5 of unguis. Tibiotarsi with 22 setae on legs I–II and 25–27 on leg III. Upper and lower subcoxae of legs I–III with 0,1/1,6–7/3–4,5–6 setae, respectively. Coxae of leg I with two front setae. Ventral tube with 4+4 latero-distal and three (seen in only one specimen) posterior setae, anteriorly without setae. Tenaculum with 4+4 teeth and a seta. Anterior furcal subcoxae with 6–7, posterior one with three setae. Anterior side of manubrium with 1+1 setae (Fig.

Remarks
Within ‘tatarica’ group, only F. baida (Ural Mts.) has medial s-setae in posterior position on all tergites. F. tubulata sp. n. shows more reduced furca: one pair of anterior setae on manubrium (vs two pairs in F. baida), latero-central setae on manubrium absent (vs present), 5–7 setae on anterior side of dens (vs 9–10). Similar structure of furca is known for F. tatarica (steppe zone of Eastern Europe) which has another s-pattern on tergites (Table
Distribution
Known from the type locality.
Derivatio nominis
The species is named after tubular shape of body.
Folsomia inoculata
Syn.: Folsomia ezoensis Yosii, 1965
Type material of J. Stach
Two adult females from the collection of J. Stach labelled as “Polonia, Czarhohora, 28.VI.1922, leg. Smraczynski. F. inoculata”. Kept in the Institute of Systematics and Evolution of Animals, Polish Academy of Sciences in Krakow, Poland.
Additional material
East Asia. Japan, Honshu Island. Nagano Prefecture, E Chino city, Kitayama, surroundings of Mugikusa Hutte, 2255 m alt., 36.0404°N, 138.3679°E, coniferous green moss forest with Tsuga, north slope, litter, 10.viii.2016; ibidem, 36.0653°N, 138.3410°E, stony meadow, grass turf, 11.viii.2016, coll. M. Potapov and N. Kuznetsova; Hokkaido Island. Japan, Hokkaido Island, Shiretoko Peninsula, trail to Mont. Rausu, different forests, litter and rotten wood, 19.viii.2016, from 350 to 1100 m alt.; Shiretoko Peninsula, nearby Shiretoko Pass, 593 m alt., 44.0617°N, 145.0913°E, stony mixed forest with Betula ermanii, 17.viii.2016; Shiretoko Peninsula, surroundings of Utoro, 500-year mixed forest, rotten wood, 20.viii.2016, 97 m alt., 44.1006°N, 145.0584°E, coll. M. Potapov and N. Kuznetsova.
China: Jilin Province, 5.viii.2009, Nearby Tian Lake, Changbai Mts., 1718 m alt., coniferous forest, soil under tree, coll. D. Wu.
Far East of Russia, Primorsky Krai, Shkotovsky District, Pidan Mount, ~800 m alt., rotten wood, 20.ix.2004, coll. M.Potapov, L.Deharveng, R.Pomorski, and A. Bedos; Shkotovsky District, trail to Mont. Khualaza, deciduous forest, rotten wood. 21.vii.2016, coll. M. Potapov and N. Kuznetsova; Sakhalin, Kholmsky District, South Kamysh Ridge of the Western Sakhalin Mountains, Spamberg Mt., mixed forest on slope, litter, 15.vi.2017, coll. A. Kuprin; Yuzhno-Sakhalinsk, Susunaysky Range, Chekhov peak, litter on top, 16.vi.2017, coll. A. Kuprin; Khabarovsky Krai, Sikhote-Alin Range, Nanaisky District, ~ 15 km N road Khabarovsk-Sovetskaya Gavan, Golaya mount. massif, Studeny Pass, coniferous forest, rotten wood, 28.vi–07.vii.2017, coll. A. Brinev; Sikhote-Alin Range, Vaninsky District, nearby Vysokogorny, valley of Mulinka River, rotten wood, ~ 600 m alt., 29.ix.2011, coll. M. Potapov; Vaninsky District, nearby Datta, coastal larch-wood, 28.ix.2011, coll. M. Potapov; Kamchatka, Elizovsky District, vicinities of Malki, 53.3219°N, 157.5502°E, 260 m alt., Betula ermanii forest, litter and rotten wood, 26.vi.2012, coll. M. Potapov and N. Kuznetsova.
Additionally, specimens from 32 localities, i.e. Ukraine (Skolevskiye Beskids), Bosnia (Perucica), Germany (Helgoland Isl., Zittau Mts, and Bavarian Alps), France (Mont Blanc), Russia (Komi, Middle Ural Mts.), Caucasus (Teberda, Guzeripl, Tsey, Khosta, Krasnaya Polyana, Lagonaki, and several other locations in Western part of North Caucasus), Armenia (Dilizhan), Georgia (Batumi, Kutaisi), Turkey (one unprecise locality, coll. L. Deharveng), Kazakstan (West Altai), Russia, West Siberia (Altai Mts.) and East Siberia (Podkamennaya Tunguska, Shira, W Sayan Mts), were examined.
Description
Body stout, very characteristic, head massive, with swollen front (Fig.
Common setae short. Sensillary formula as 43/22235 (s). Micro s-setae as 10/100 (ms). Tergal s-setae short and distinct. Medial s-setae on Th.II–III in front of p-row, on Abd.I–III in posterior position, between Md and Mdl. Abd.V with five s-setae arranged as three short (as, accp1, accp2), one lateral long and tubular, and one latero-ventral, short (‘3+1+1’ pattern) (Fig.
Unguis of normal shape, without lateral and inner teeth. Empodial appendage usually longer than half of unguis (0.5–0.7). All tibiotarsi with additional setae: 23–27 setae on legs I–II and >30 setae on leg III, as a whole. Upper and lower subcoxae of legs I–III with 0,1/5–7,8–12/7–9,8–10 setae, respectively. Coxae of leg I with two front setae. Ventral tube with 4–5+4–5 latero-distal and 6–7 posterior setae (with four in distal transversal row), anteriorly without setae. Tenaculum with 4+4 teeth and one or two setae. Anterior furcal subcoxae with 11–16, posterior one with four setae. Anterior side of manubrium with 2+2 setae (rarely 2+3 or 1+2). Posterior side of manubrium with 4–5+4–5 latero-basal, two apical setae (ap), 2+2 setae in distal transversal row, pair of lateral setae present or absent (see the Discussion part), and 4–5(3–6)+4–5(3–6) in central part. Dens normally with 10–14 anterior setae (the whole range is 8–16). Posterior side of dens crenulated and with four setae: three (very rarely two) basal and one at the middle, no subapical setae. Mucro bidentate. Ratio of manubrium : dens : mucro = 3.1–4.9 : 2.4–4.3 : 1.
Remarks
Folsomia inoculata is a rather peculiar species due to several characteristics. On Abd.V the differentiation of s-setae is unique: accp3-s is well-marked, tubular, and longer than three shortened and thin s-setae of “dorsal triplet” (shown in detail on fig.14 in
Available vast material on this species shows a wide variation in several characters (chaetotaxy of manubrium and dens, shape of PAO, body length) which, however, are individual or population-dependent and does not indicate several species.
According to the original description, PAO is not constricted in F. inoculata, which was also shown in associated figures by
The modern detailed description of the species is given in
Size of the body ranges between 0.9 and 1.7. Specimens from eastern populations appears to be smaller than in western ones, but the whole variation is strongly overlapping (0.9–1.5 vs 1.1–1.7 mm, respectively).
The performed multivariate analysis of metric morphology did not reveal any irregularities, and noticeable differences between eastern and western populations were not detected (Fig.
The species is facultatively parthenogenetic and its populations mostly consist of females. Males were seen by us only in four “central” localities: in Middle Ural mountains (upper flow of Pechora River), East Siberia (Podkamennaya Tunguska), Caucasus (Aibga Range), Turkey, and Kazakstan (West Altai).

F. inoculata 81 Appearance 82 Position of macrosetae, setae of p-row, and s-setae on corpus 83PAO, variations (83, 84 Japan 85 Caucasus) 86–88 Manubrium, lateral view (Germany: Helgoland, coll. J. Schulz (86), Caucasus: Aibga (87), Japan: central Honshu (88) 89 Scatterplot of 84 individuals from five large regions of Palearctic basing on nine length ratios (for explanations see the Methods part). Abbreviations: pr, l2–groups of setae on posterior side of manubrium (notation as in Fjellberg, 2007), C–Caucasus, E–Europe, FE–Far East of Russia, J–Japan, S–Siberia, U–Ural.
Distribution and ecology
The species is widely distributed in the region (Fig.
Distributional range of F. inoculata appears to be restricted to the Holarctic. In the Palearctic we still have not seen specimens from the eastern areas of East Siberia, such as Buryat Republic and Amurskaya District, despite intensive collections in appropriate sites. Thus, all populations can probably be divided to ‘western’ and ‘eastern’ which are inseparable by morphology for the present. In Scandinavia and the westernmost part of Europe F. inoculata is very rare and appears to be an alien species (absent, for example, in the Iberian Peninsula). In the Nearctic, the species is also infrequent although it occurs at least on the Pacific coast of USA (Oregon, Cascade Range, coll. A. Smolis, our identification, new record). Folsomia inoculata does not occur in the Arctic; the northernmost record is north-east corner of Komi Republic (67.50°N, NE European part of Russia) (
The species often occurs in forest litter while apparently preferring rotten wood where it can be very abundant. Folsomia inoculata is the most dendrophilous species of the ‘inoculata’ group and, very likely, in the genus, having associated shape of body and crushing mouth parts. The species is also sporadically recorded in specific sites enriched by organic matter.

A key to the species of Folsomia of the ‘inoculata’ group
| 1 | Laterally positioned accp-3 s-seta much longer than three dorsal s-setae on Abd.V (Figs |
inoculata Stach, 1947 (Holarctic) |
| – | Accp-3 s-seta as long as or shorter than three dorsal s-setae on Abd.V (Figs |
2 |
| 2 | More than eleven setae on anterior side of dens | 7 |
| – | Fewer than eleven setae on anterior side of dens | 3 |
| 3 | Medial s-setae in front of p-row on Th.II–III and Abd.I–II (Fig. |
4 |
| – | Medial s-setae in p-row on Th.II–III and Abd.I–II (Fig. |
5 |
| 4 |
Abd.I without ms-setae (Fig. |
calcarea sp. n. (south-east of European part of Russia) |
| – |
Abd.I with ms-setae (as in Fig. |
torpeda Potapov, 2006 (north and east of European part of Russia, north-west of Siberia) |
| 5 | Medial s-setae in front of p-row on Abd.III (as in Fig. |
tatarica Martynova, 1964 (European part of Russia) |
| – | Medial s-setae in p-row on Abd.III (Fig. |
6 |
| 6 | Dens with 5–7 anterior setae. Manubrium with one pair of setae on anterior side (Fig. |
tubulata sp. n. (Yakutia) |
| – | Dens with 9–10 anterior setae. Manubrium with two pairs of setae on anterior side | baida Potapov, 2006 (north and north-east of European part of Russia) |
| 7 | Medial setae on ventral side of Th.III present (Fig. |
8 |
| – | Medial setae on ventral side of Th.III absent (Fig. |
16 |
| 8 | Medial s-setae on Th.III–Abd.II absent (s-formula: 42/11235) | bashkira Potapov, 2006 (Ural) |
| – | Medial s-setae on Th.III–Abd.II present (s-formula: 43/22235 or 33/22224) | 9 |
| 9 | Medial s-setae in front of p-row on Th.II–III (Fig. |
brevisensilla Potapov & Babenko, 2000 (North-East of Russia) |
| – | Medial s-setae within p-row on Th.II–III (Fig. |
10 |
| 10 | One and three dorsal s-setae on Abd.IV and V, respectively (Fig. |
11 |
| – | Two and four dorsal s-setae on Abd.IV and V, respectively (Fig. |
13 |
| 11 | Manubrium with 2+2 setae on anterior side and no more than 3+3 setae in central part of posterior side (Figs |
tertia sp. n. (Far East of Russia) |
| – | Manubrium with 3+3 or more setae on anterior side and more than 3+3 setae in central part of posterior side (Fig. |
12 |
| 12 |
Abd.V with p1-setae shorter than accp1 s-setae (Fig. |
laconica sp. n. (Amurskaya District (Far East of Russia), North Korea) |
| – |
Abd.V with p1-setae as long as accp1 s-setae (Fig. |
trisensilla sp. n. (East Siberia, Far East of Russia) |
| 13 | Setae on anterior side of ventral tube absent | 14 |
| – | Setae on anterior side of ventral tube present | setifrontalis Potapov & Marusik, 2000 (south of Far East of Russia) |
| 14 | Anterior side of manubrium with 2–3+2–3 setae (Fig. |
macrochaetosa Martynova, 1977 (Far East of Russia) |
| – | At whole, seven or more setae on anterior side of manubrium (Figs |
15 |
| 15 | Unpaired setae on anterior side of manubrium absent (Figs |
amurica sp. n. (Amurskaya District (Far East of Russia), Inner Mongolia Province (North China)) |
| – | Unpaired setae on anterior side of manubrium present (Figs |
imparis sp. n. (Japan, south of Far East of Russia) |
| 16 | Medial s-setae in front of p-row on Abd.I–III (Figs |
breviseta sp. n. (North-East Asia) |
| – | Medial s-setae within p-row on Abd.I–III (Figs |
hidakana Uchida & Tamura, 1968 (Japan, Korea, south of Far East of Russia) |
Acknowledgements
We would like to express our sincere thanks to Drs Vasily Alpatov, Marina Babykina, Anne Bedos, Vasily Boeskorov, Alexey Brinev, Yun Bu, Sergey Bukhkalo, Huang Chen-Wang, Louis Deharveng, Tatyana Dobrolyubova, Wolfram Dunger, Arne Fjellberg, Saori Fujii, Anna Geraskina, Natalia Goloschapova, Ayuna Gulgenova, Ryo Kitagawa, Anastasia Korotkevich, Nikolay Kozlov, Alexander Kremenitsa, Arkady Lelej, Andrey Matalin, Igor Melnik, Akira Mori, Taizo Nakamori, Keiko Niijima, Kei-ichi Okada, Andrey Ptashinsky, Seikoh Saitoh, Juergen Schulz, Maxim Shashkov, Dmitry Shitikov, Yulia Shveenkova, Olga Sleptsova, Olga Smirnova, Yelena Sokolova, Sofia Stebaeva, Yasuhiko Suma, Anastasia Taskaeva, Lilia Vanyavina, Ivan Vtorov, Wanda Weiner, and Dong-Hui Wu, for kindly providing material on Collembola or for field assistance.
The authors are grateful to the management and staff of the Shiretoko National Park (Japan, Hokkaido), Ussurisky Nature Reserve (Russia, Primorsky Krai), “Land of Leopard” National Park (Primorsky Krai), Sikhote-Alin Nature Reserve (Primorsky Krai), State Reserve of Laso (Primorsky Krai), Alkhanai National Park (Russia, Zabaikal’sky Krai), Khingansky State Nature Reserve (Russia, Amurskaya Region), Zeysky State Nature Reserve (Amurskaya Region), Caucasian Biosphere Reserve (Russia, North Caucasus), who provided the collecting permit and the favorable conditions to our field work. Our special thanks are to Taizo Nakamori (Yokohama National University) whose hospitality was very helpful during our stay in Japan. We also indebted to Javier Arbea and an anonymous reviewer for their critical comments.
The study was partly supported by Japan–Russia Research Cooperative Program of RFBR (project No. 16-54-50068) for M.Potapov, N.Kuznetsova, A.Babenko, and A. Kuprin.
References
- Babenko AB, Potapov MB, Taskaeva AA (2017) The collembolan fauna of the East European tundra. Russian Entomological Journal 26(1): 1–30.
- Fjellberg A (2007) The Collembola of Fennoscandia and Denmark. Part II: Entomobryomorpha and Symphypleona. Fauna Entomologica Scandinavica 42: 1–264. https://doi.org/10.1163/ej.9789004157705.i-265
- Fujii S, Takeda H (2012) Succession of collembolan communities during decomposition of leaf and root litter: Effects of litter type and position. Soil Biology & Biochemistry 54: 77–85. https://doi.org/10.1016/j.soilbio.2012.04.021
- Furuno K, Hasegawa M, Hisamatsu M, Ichisawa K, Itoh R, Niijima K, Suma Y, Tamura H, Tanaka S (2000) List of collembolan species recorded from Japan and their Japanese names. Edaphologia 66: 75–88. [In Japanese with English summary]
- Hasegawa M, Fukuyama K, Makino S, Okochi I, Tanaka H, Okabe K, Goto H, Mizoguchi T, Sakata T (2009) Collembolan community in broad-leaved forests and in conifer stands of Cryptomeria japonica in Central Japan. Pesquisa Agropecuária Brasileira 44: 881–890. https://doi.org/10.1590/S0100-204X2009000800012
- Hishi T, Tashiro N, Maeda Y, Inouei S, Cho K, Yamauchi K, Ogata T, Mabuchi T (2012) Soil depth distribution and the patterns of alpha- and beta-diversity of families of soil Collembola in cool-temperate deciduous natural forests and larch plantations of northern Japan. Edaphologia 91: 9–20.
- Iwanami M, Tsuchiya D, Niijima K (1980) Density and species composition of Acarina and Collembola in Tama, Tokyo, in relation to forest types and soil properties. Japanese Journal of Ecology 30: 145–154. [In Japanese with English summary]
- Kurcheva GF (1977) Soil invertebrates of Soviet Far East. Nauka, Moscow, 131 pp. [In Russian]
- Kutyreva LT (1979) Fauna and population of collembolans in soils of spruce-broadleaf forests of South Primorye. In: Chernova NM (Ed.) Fauna i Ekologiya bespozvonochnykh.MGPI, Moscow, 53–60. [In Russian]
- Kutyreva LT (1984) Springtails in secondary oak forests of Primorsky Krai. In: Gilyarov MS, Chernova NM (Eds) Fauna i Ekologiya nogokhvostok.Nauka, Moscow, 118–123. [In Russian]
- Kutyreva LT (1988) Complexes of springtails in anthropogenic forests. In: Izmeneniye rastitelnogo i zhivotnogo mira pod vliyaniyem anthropogennoi deyatelnosti v usloviyakh Dal’nego Vostoka. KGPI, Khabarovsk, 142–148. [In Russian]
- Lee BH (1973) Étude de la faune Coréenne des Collemboles. I. Liste des Collemboles de Corée et description de trois espèces. Revue d’Ecologie et de Biologie du Sol 10: 435–449.
- Martynova EF (1973) To diagnostics of springtails of the genus Folsomia Willem (Collembola, Isotomidae) occurring on the territory of the USSR. Fauna of Siberia. 16. Nauka, Novosibirsk, 7–23. [In Russian]
- Natuhara Y, Imai C, Takeda H (1994) Classification and ordination of communities of soil arthropods in an urban park of Osaka City. Ecological Research 9: 131–141. https://doi.org/10.1007/BF02347488
- Niijima K, Hasegawa M (2011) Classification of the Family Isotomidae Börner, 1913 (Apterygota: Collembola) from Japan 1. Anurophorinae Borner, 1906 and Proisotominae Stach, 1947. Edaphologia 89: 29–69. [In Japanese]
- Potapov M (2001) Isotomidae. In: Dunger W (Ed.) Synopses on Palearctic Collembola.Vol. 3, Abhandlungen und Berichte des Naturkundemuseums Gorlitz 73, 1–603.
- Potapov MB, Greenslade P (2010) Redescription of Folsomia loftyensis Womersley with notes on the sensillary arrangement of the genital segment in the genus (Collembola: Isotomidae). Zoologischer Anzeiger 249: 13–20. https://doi.org/10.1016/j.jcz.2010.01.004
- Potapov M, Marusik Yu (2000) New and little known Folsomia Willem, 1902 (Collembola: Isotomidae) from South Kuriles. Russian Entomological Journal 9: 99–102.
- Schulz HJ (1999) Bemerkenswerte sächsische Collembolenfunde, insbesondere aus Heidemooren der Oberlausitz, und eine Aktualisierung des Verzeichnisses der Springschwänze für Sachsen (Collembola). Entomologische Nachrichten und Berichte 43: 233–236.
- Solntseva EL, Molodova LP (1979) Springtails of South Sakhalin. In: Chernova NM (Ed.) Fauna i Ekologiya bespozvonochnykh.MGPI, Moscow, 34–37. [In Russian]
- Stach J (1947) The Apterygotan fauna of Poland in relation to the world-fauna of this group of insects. Family Isotomidae. Polska Akademia Umiejętności, Acta Monographica Musei Historiae Naturalis, Krakow, 488 pp.
- Suma Y (1997) Collembolan fauna of ‘heat islands’, Akan National Park. Sylvicola 15: 19–28. [In Japanese]
- Suma Y (2008) Collembolan fauna of Mt. Shari Prefectural natural Park, Eastern Hokkaido. Jezoensis 34: 79–86. [In Japanese]
- Takeda H (1995) Changes in the collembolan community during the decomposition of needle litter in a coniferous forest. Pedobiologia 39: 304–317.
- Tamura H, Chiba S (1977) Collembola. In: Kitazawa Y (Ed.) JIBP Synthesis 15.Ecosystem analysis of the subalpine coniferous forest of the Shigayama IBP Area, Central Japan, University of Tokyo Press, Tokyo, 88–97.
- Tanaka M (1970) Ecological studies on communities of soil Collembola in Mt. Sobo, southwest Japan. Japanese Journal of Ecology 20(3): 102–110.
- Uchida H, Tamura H (1968) Descriptions and records of Collembola from Hokkaido. II. Kontyu 36: 1–13.
- Yamauchi S, Suma Y (1999) Collembola fauna of the Towada Mountain Range and Mts. Hakkoda-san, Aomori Pref., Japan. Journal of Natural History of Aomori 4: 51–57. [In Japanese]
- Yamauchi S, Suma Y (2009) Collembolen Fauna in Mt. Hashikami-dake, Hashikami-machi Town, Aomori Prefecture, northern Japan. Bulletin of the Aomori Prefectural Museum 38: 9–16. [In Japanese]
- Yosii R (1939) Isotomid Collembola of Japan. Tenthredo 2: 348–391.
- Yosii R (1965) On some Collembola of Japan and adjacent countries. Contributions from the Biological Laboratory, Kyoto University 19: 1–71.
- Yosii R (1977) Critical check list of the Japanese species of Collembola. Contributions from the Biological Laboratory, Kyoto University 25: 141–170.