Research Article |
|
Corresponding author: Simon van Noort ( svannoort@iziko.org.za ) Academic editor: Norman Johnson
© 2022 Simon van Noort, Scott Richard Shaw, Robert S. Copeland.
This is an open access article distributed under the terms of the Creative Commons Attribution License (CC BY 4.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Citation:
van Noort S, Shaw SR, Copeland RS (2022) Revision of the endemic African genus Dinapsis (Dinapsini, Megalyridae, Hymenoptera) with description of seven new species. ZooKeys 1112: 27-122. https://doi.org/10.3897/zookeys.1112.82307
|
Abstract
The endemic Afrotropical genus Dinapsis is revised, and seven new species are described and illustrated: D. bicolor van Noort & Shaw, sp. nov., D. gamka van Noort & Shaw, sp. nov., D. igneus van Noort & Shaw, sp. nov., D. spinitibia van Noort & Shaw, sp. nov., D. taita van Noort & Shaw, sp. nov., D. tricolor Shaw & van Noort, sp. nov., D. zulu Shaw & van Noort, sp. nov. The distribution of the Central African Republican species D. centralis Shaw & van Noort, 2009 is extended to include Cameroon, Kenya and Uganda. Dinapsis turneri Waterston, 1922, previously only known from the poorly preserved holotype female, is redescribed based on newly collected specimens. The distribution of this Western Cape species is extended to include the Eastern and Northern Cape provinces of South Africa. Four distinct species groups within the genus are proposed and diagnosed. An illustrated identification key to all described species of Dinapsis is provided. Online interactive Lucid keys to Afrotropical megalyrid genera and Dinapsis species are available at http://www.waspweb.org.
Keywords
Africa, barcode, holotype, identification key, new species, parasitoid, taxonomy, wasp
Introduction
The Megalyridae comprise eight extant genera and five extinct genera (
Dinapsis was originally placed in the family Dinapsidae, erected by
The hosts of most megalyrid genera and species are unknown, but available evidence suggests that they are idiobiont endoparasitoids of concealed insect larvae. The only detailed biological observations are for a few species of Megalyra (
We here revise the mainland African and Mauritian species of Dinapsis and provide identification keys to all the described Dinapsis species, including those named so far from Madagascar. There are numerous additional undescribed species from Madagascar that will be treated in a future revision. Images and online interactive Lucid identification keys to the Afrotropical megalyrid genera and Dinapsis species are available at WaspWeb (http://www.waspweb.org) (
Materials and methods
As far as possible morphological terminology follows the Hymenoptera Anatomy Ontology project (HAO Portal hymao.org); where terms are not defined on HAO,
The following abbreviations are used in this paper:
F1-F12 sequential antennal flagellomere segments, basal to distal;
LOL lateral ocellar line, shortest distance between inner margins of median and lateral ocelli;
OOL ocular ocellar line, shortest distance from inner eye orbit and outer margin of posterior ocellus;
POL posterior ocellar line, shortest distance between inner margins of posterior ocelli;
T1-T5 sequential tarsomere segments, basal to distal.
A leg from each of ten specimens of Dinapsis, representing seven species, was submitted to BOLD for DNA barcoding. DNA extracts were obtained from the legs at BOLD using a glass fibre protocol (
Images were acquired at SAMC with a Leica LAS 4.9 imaging system, comprising a Leica Z16 microscope with a Leica DFC450 Camera and 0.63× video objective attached. The imaging process, using an automated Z-stepper, was managed using the Leica Application Suite v. 4.9 software installed on a desktop computer. Diffused lighting was achieved using a Leica LED 5000 Dome. Additional images were acquired at the University of Wyoming using Leica Application Suite (Leica Microsystems) and image stacking software, Combine ZM and Zeren Stacker v. 1.04. SEM images were acquired at UWIM using an Environmental Scanning Electron Microscope at the UW Microscopy Core Facility. Dry uncoated specimens were mounted on metal stubs for SEM study at low operating voltages and photography, then the glue was dissolved, and the specimens were remounted, undamaged, on pins and points with their original data. An SEM label was added to indicate which specimens were studied. All images included in this paper, as well as additional images, and online interactive keys to Dinapsis species are available on WaspWeb (https://www.waspweb.org) (
Lucid pathway and Lucid matrix keys were developed using Lucid Builder v. 4.0.23. Character matrices were generated and edited using Microsoft Excel; matrices were then used as input into Lucid matrix key production (
Distribution maps were generated using SimpleMappr, an online data resource in the public domain (
Depositories
CAS California Academy of Sciences, San Francisco, USA (Brian Fisher)
ICIPE International Centre of Insect Physiology and Ecology, Nairobi, Kenya (Robert Copeland)
NHMD Natural History Museum of Denmark, University of Copenhagen, Denmark (Lars Vilhelmsen)
NMSA KwaZulu-Natal Museum, Pietermaritzburg, South Africa (Tricia Pillay)
UWIM UW Insect Museum, Laramie, USA (Scott Shaw)
Systematics
Identification keys
Standard dichotomous keys to genera of African Megalyridae and to the described species of Dinapsis are presented below. Online interactive Lucid pathway and Lucid matrix keys are available on WaspWeb (
Identification key to genera of African Megalyridae
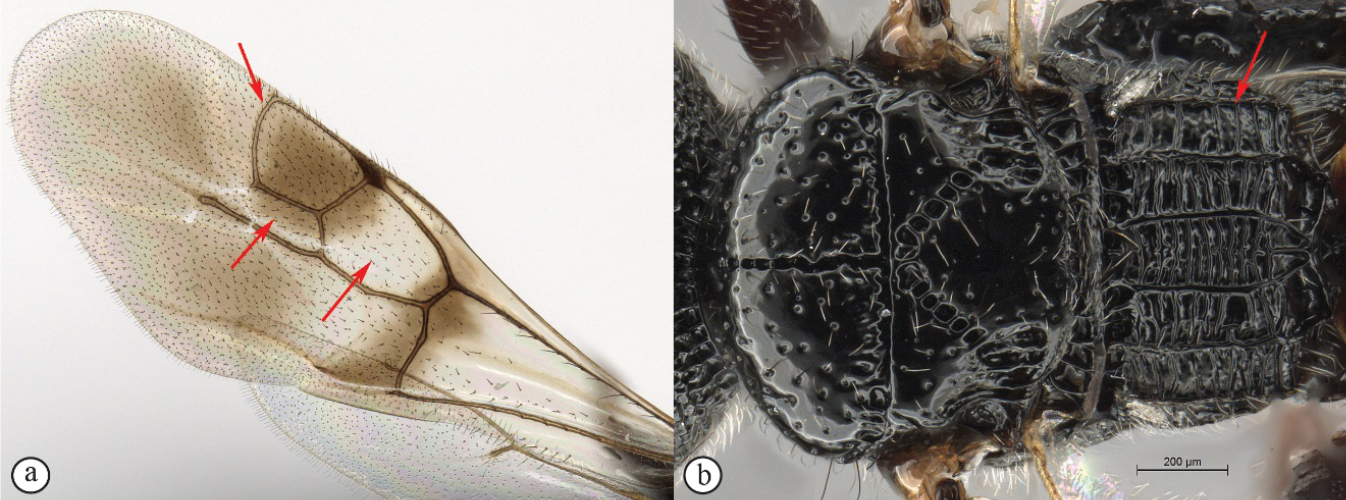
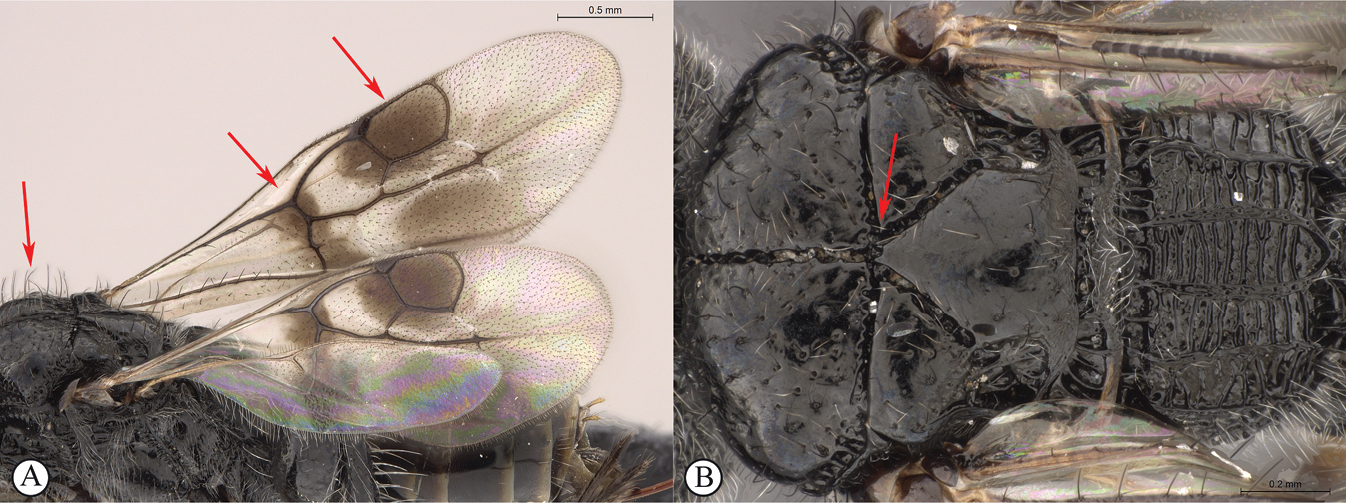
| 1 | Mesoscutum with a weak median sulcus (A); propodeum with a shallow, longitudinal median sulcus (A); eyes not surrounded by an orbital carina (B); fore wing with veins 1mcu and Cu1 present (C) | Megalyra (one introduced species in South Africa: Megalyra fasciipennis) |

|
||
| – | Mesoscutum with a strong longitudinal median sulcus (a); propodeum without a longitudinal median sulcus (a); eyes surrounded by an orbital carina (b); fore wing veins 1mcu and Cu1 spectral (c, d) | 2 |

|
||
| 2 | Fore wing with one submarginal cell (A); radial vein reduced, radial cell open (A); propodeum rugulose without parallel longitudinal carinae (B) | Megalyridia (monotypical genus: Megalyridia capensis is restricted to Eastern, Northern, and Western Cape provinces of South Africa) |

|
||
| – | Fore wing with two submarginal cells (a); radial cell closed (a); propodeum with distinctive pattern of six longitudinal, nearly parallel carinae, each tract with lateral transverse carinae that may be reduced in number (b) | Dinapsis |
|
||
Dinapsis
Type species

Diagnosis
Dinapsis species have a distinctive forewing venation pattern, with vein Rs curving abruptly to the anterior wing margin to form a short, abruptly truncate marginal cell (cell 2R1) (Fig.

Recognition
The genus Dinapsis can be keyed out from the two other megalyrid genera present in Africa using the above generic key that is also available as an online interactive Lucid key on WaspWeb (https://www.waspweb.org) (
Biology
Unknown. Specimens are most commonly collected in Malaise traps, and less frequently in yellow pan traps, via tree canopy fogging, or through leaf litter sifting. To our knowledge only a single living specimen of this genus has ever been photographed (Fig.
Species richness
The genus Dinapsis was previously known from ten Madagascan and two continental African species (
D. albicauda Mita & Shaw, 2020 (Madagascar)
D. albicoxa Hedqvist, 1967 (Madagascar)
D. bicolor van Noort & Shaw, sp. nov. (South Africa)
D. centralis Shaw & van Noort, 2009 (Central African Republic, Cameroon, Kenya, Uganda)
D. cresta Mita & Shaw, 2020 (Madagascar)
D. gamka van Noort & Shaw, sp. nov. (South Africa)
D. hirtipes Hedqvist, 1967 (Madagascar)
D. igneus van Noort & Shaw, sp. nov. (Mauritius)
D. luteus Mita & Shaw, 2020 (Madagascar)
D. nubilis Hedqvist, 1967 (Madagascar)
D. oculohirta Hedqvist, 1967 (Madagascar)
D. planifrons Mita & Shaw, 2020 (Madagascar)
D. scriptus Mita & Shaw, 2020 (Madagascar)
D. seyrigi Hedqvist, 1967 (Madagascar)
D. spinitibia van Noort & Shaw, sp. nov. (Tanzania)
D. taita van Noort & Shaw, sp. nov. (Burundi, Kenya)
D. tricolor Shaw & van Noort, sp. nov. (Kenya, South Africa)
D. turneri Waterston, 1922 (South Africa)
D. zulu Shaw & van Noort, sp. nov. (South Africa)
Distribution
Burundi, Cameroon, Central African Republic, Kenya, Madagascar, Mauritius, South Africa, Tanzania, Uganda (Figs
Barcoding
Of the ten Dinapsis samples submitted to BOLD (representing seven species: D. bicolor, D. gamka, D. igneus, D. taita, D. tricolor, D. turneri, D. zulu) DNA was successfully extracted from seven specimens represented by five species (Table
Details of barcode data for the seven Dinapsis specimens from which DNA was successfully extracted.
| Dinapsis species | Type status | Country | Sample ID | BOLD sequence code | BIN URI |
|---|---|---|---|---|---|
| D. bicolor | Holotype ♀ | South Africa | 38754_A03_SAM-HYM-P088338 | FSA189521 | BOLD:AEH7061 |
| D. igneus | Holotype ♀ | Mauritius | 8754_A01_Din_ign_fem | FSA189321 | None |
| D. igneus | Paratype ♂ | Mauritius | 38754_A02_Din_ign_mal | FSA189421 | None |
| D. taita | Paratype ♀ | Burundi | 08672-MEGSPBURH9 | KINS160911 | BOLD:AAZ9109. |
| D. tricolor | Paratype ♀ | Kenya | 38754_A07_ |
FSA189921 | None |
| D. tricolor | Paratype ♀ | South Africa | 38754_A08_NMSA-HYM-000546 | FSA190021 | None |
| D. zulu | Paratype ♀ | South Africa | 38754_A09_NMSA-HYM-000539 | FSA190121 | None |
Dinapsis species groups
Dinapsis hirtipes species group. This species-group was proposed by
Dinapsis oculohirta species group. In this group of species, the eyes are “hairy,” being covered with numerous small setae (Fig.
In this paper we define the oculohirta species group as comprising D. oculohirta from Madagascar, D. centralis from central Africa, as well as seven new species from the African mainland and Mauritius described below (D. bicolor, D. gamka, D. igneus, D. spinitibia, D. taita, D. tricolor, and D. zulu). Dinapsis oculohirta, described from Madagascar, has not been found on the African mainland yet. Otherwise, the named species in this group are from the African mainland. However, additional new species in this group from Madagascar are planned to be covered in a separate paper.
Despite these species being morphologically similar, this group is probably a paraphyletic grade, and may be just a convenient way to group many basal species in the genus. Hairy eyes also occur in the sister-genus Ettchellsia from south-east Asia, so out-group comparison suggests that the setose eyes of this species group are a plesiomorphic condition, and perhaps the lack of ocular setae (bare eyes) is a synapomorphy uniting the three other Dinapsis species groups. The species in the oculohirta group, if not comprising a monophyletic lineage, are at least inferred to be the most basal lineages in the genus.
The South African species Dinapsis turneri Waterston has for many years been difficult to assign into a group with other species, especially since until now it was known only from the holotype specimen. Morphologically the holotype of D. turneri looks similar to species in the oculohirta species group but the eyes do not appear to have setae. This point is debatable because the eyes of the holotype specimen are oddly shrunken and it is hard to observe whether setae are present or not (see Figs
Dinapsis nubilus species group. In this species group the mesoscutum has distinctive lobe-like protuberances postero-laterally (see key couplet 4, images A, B). These lobes are parts of a projecting carina running obliquely on the lateral mesoscutal surface that appear lobe-like in dorsal view. A few species assigned to other groups (such as D. seyrigi and D. tricolor) to a lesser extent have the same carina projecting somewhat, but in these it is not so lobe-like in dorsal view. This feature is shared with Dinapsis albicoxa Hedqvist, the only other species now assigned to this small, but distinctive group. This group is known only from Madagascar.
Dinapsis seyrigi species group. In this species group the gena behind the orbital carina is smooth (see key couplet 3, image A). This group consists of the typical Dinapsis seyrigi and several other undescribed species from Madagascar. None has yet been found on the African mainland. The species are not frequently collected, but the smooth lateral areas of the head are quite distinctive. Some of the undescribed species have distinctive patches of silver setae on the sides of the mesopleuron. This is probably a monophyletic group (other groups all have very coarse sculpture on the sides of the head).
Key to species of Dinapsis
| 1 | Vertex produced dorsally forming a raised carina (A, B); subquadrate in anterior view (A) (Dinapsis hirtipes Hedqvist species group) | 14 |

|
||
| – | Vertex normal, evenly rounded (a, b) | 2 |

|
||
| 2 | Two large obvious sharply-pointed spines in close apposition on hind tibial apex (A); mesoscutum with sharply-projecting wave-shaped lateral teeth and strong transverse ridges (B) | Dinapsis spinitibia van Noort & Shaw, sp. nov. |

|
||
| – | Hind tibia without apical spines (a); mesoscutum laterally rounded, mesoscutal transverse ridges absent (b) | 3 |
|
||
| 3 | Gena behind the orbital carina smooth (A); vertex polished, with only a few scattered minute punctures and setae (B); propodeum medially polished, mostly lacking transverse carinae between the submedian longitudinal carinae (B) | Dinapsis seyrigi Hedqvist |

|
||
| – | Gena behind orbital carina sculptured, or densely punctate (a); vertex often sculptured (a), if smoother then with denser punctation and setae (b); propodeum medially usually with well-developed transverse carinae between the submedian longitudinal carinae (b) | 4 |

|
||
| 4 | Mesoscutum with a distinctive and strongly projecting carina (lateral mesoscutal margin projected as a flange) situated posterolaterally that appears as a lobe-like lateral protuberance in dorsal view (A, B); mesoscutum always smooth (A) | 5 |

|
||
| – | Mesoscutum without a strongly projecting carina situated postero-laterally (a), or if carina slightly projecting (b) then adjacent mesoscutal surface with large fovea (b) | 6 |

|
||
| 5 | Wings infuscate (brownish) without obvious bands (A); head weakly foveate-reticulate, face and vertex with polished areas between fovea, setae sparse and short (B) | Dinapsis nubilis Hedqvist |

|
||
| – | Wings with transverse dark bands (a); head coarsely reticulate, with long setae (b) | Dinapsis albicoxa Hedqvist |

|
||
| 6 | Vertex with a medial row of punctures, which may be fused to form a shallow groove, between ocelli (A, B) | Dinapsis oculohirta Hedqvist |

|
||
| – | Vertex without a medial row of punctures between ocelli (a, b) | 7 |

|
||
| 7 | Vertex and mesoscutal lobes coarsely rugulose-reticulate (A); scutoscutellar sulcus demarcated by broad, longitudinally striate furrow (A); flagellum uniformly dark, or occasionally lighter in basal 1/3 (B) | Dinapsis centralis Shaw & van Noort |

|
||
| – | Vertex finely or coarsely punctate, mesoscutal lobes polished with sparse punctures or scattered fovea (a); scutoscutellar sulcus demarcated by foveate furrow (a); flagellum often with medial white band (b) | 8 |

|
||
| 8 | Mesoscutum projecting anteriodorsally as a bilobed crest (A), distinct in lateral view (B); head and mesosoma densely rugulose-punctate, with metallic greenish bronze sheen (requires good lighting to discern) (A, B) | Dinapsis igneus van Noort & Shaw, sp. nov. |

|
||
| – | Mesoscutum not projecting dorsally (a, b); mesosoma polished or punctate, sometimes with scattered foveae, non-metallic (a, b) | 9 |

|
||
| 9 | Metasoma pale orange-brown, contrasting with black head and mesosoma (A, B); ovipositor short, subequal in length to metasoma (B) | Dinapsis bicolor van Noort & Shaw, sp. nov. |

|
||
| – | Metasoma black or dark brown, body unicolourous (a, b), mesoscutum may have small orange patches; ovipositor long, as long as mesosoma and metasoma combined (b) | 10 |

|
||
| 10 | Vertex coarsely punctate (A, B); dorsal mesoscutal areas laterally with fovea, contrasting with polished medial mesoscutal area (A, B) | 11 |

|
||
| – | Vertex finely punctate (a, b); dorsal mesoscutal areas laterally without fovea, polished with fine punctation (a, b) | 12 |

|
||
| 11 | Fovea on dorsal mesoscutal areas indistinct, small (A); scutoscutellar sulcus demarcated by narrow furrow with fovea indistinct (A); mesoscutum with orange patches (A, B) | Dinapsis taita van Noort & Shaw, sp. nov. |

|
||
| – | Fovea on dorsal mesoscutal areas large and distinct (a); scutoscutellar sulcus demarcated by strongly foveate furrow (a); mesoscutum uniformly black (a) | Dinapsis tricolor Shaw & van Noort, sp. nov. |

|
||
| 12 | Forewing with narrow dark bands, basal band absent from costal cell (A); apical infuscation diffuse (A); short, dark setae on dorsal surface of mesoscutum (A); scutoscutellar sulci meet the transscutal articulation independently (B) | Dinapsis turneri Waterston |
|
||
| – | Forewing with broad dark bands, basal band extending to anterior wing margin through costal cell (a); apical infuscation darker, obviously demarcated subparallel to apical wing margin (a); longer, dark setae on dorsal surface of mesoscutum; scutoscutellar sulci meet before reaching the transscutal articulation (b) | 13 |

|
||
| 13 | Head small, height in lateral view equivalent to mesosomal height (A); postocular furrow broad, 1/2 genal width behind postocular carina, quarter of eye width (B) | Dinapsis zulu Shaw & van Noort, sp. nov. |

|
||
| – | Head large, height in lateral view 1.5× greater than mesosomal height (a); postocular furrow narrow, 1/5 of genal width, 1/7 of eye width (b) | Dinapsis gamka van Noort & Shaw, sp. nov. |

|
||
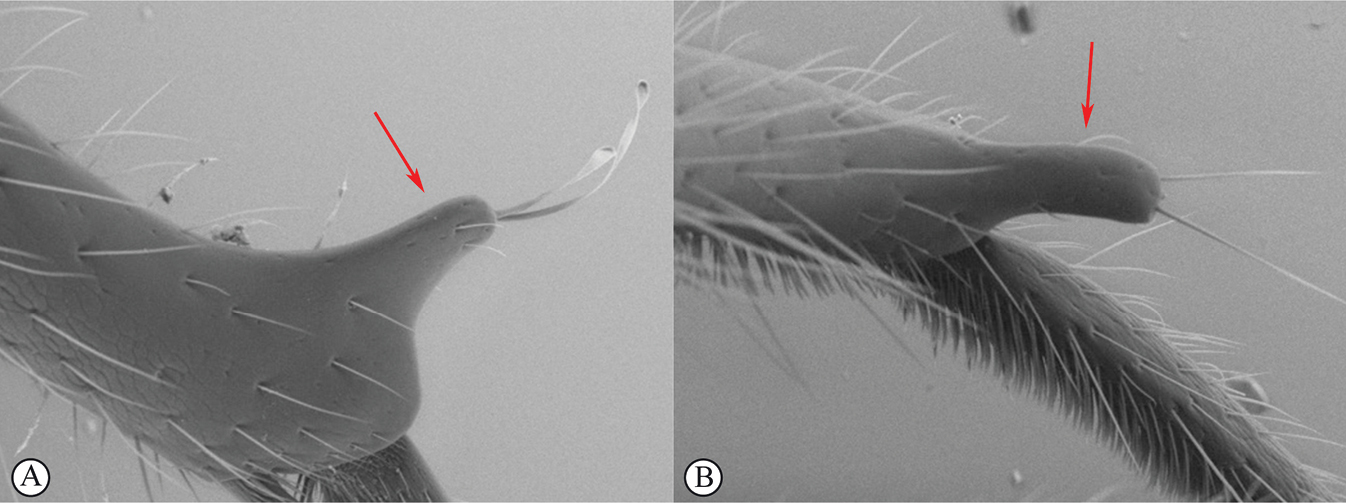
| 14 | Distal prong present on hind tibia (A, B) | 15 |
|
||
| – | Distal prong absent on hind tibia (a, b) | 16 |

|
||
| 15 | Setae on distal prong spatulate (A); female body mostly brownish (B) | Dinapsis planifrons Mita & Shaw |

|
||
| – | Setae on distal prong simple (a, b); female body mostly blackish (b) | Dinapsis scriptus Mita & Shaw |

|
||
| 16 | Transverse carina on vertex strongly developed (A, B), width of carina wider than minimum distance between inner eye margins in anterior view (A) | 17 |

|
||
| – | Transverse raised carina on vertex less developed (a, b), width of carina narrower than minimum distance between inner eye margins in anterior view (b) | 18 |

|
||
| 17 | Forewing with dark bands; posterior margin of raised vertex (A) and anterior margin of mesoscutum excavated medially (B) | Dinapsis hirtipes Hedqvist |

|
||
| – | Forewing without dark band; posterior margin of raised vertex (a) and anterior margin of mesoscutum flat (b), at most slightly excavated medially | Dinapsis luteus Mita & Shaw |

|
||
| 18 | Head round, as long as wide in lateral view (A); dorsal surface of head rounded in lateral view (A); ovipositor whitish excluding brown apex (B) | Dinapsis albicauda Mita & Shaw |

|
||
| – | Head oval, longer than wide in lateral view (a); dorsal surface of head less rounded in lateral view (a); ovipositor entirely brown (b) | Dinapsis cresta Mita & Shaw |

|
||
Dinapsis albicauda
Material examined
Holotype . Madagascar • ♀; Fianarantsoa Prov., Forêt d’Atsirakambiaty, 7.6 km 285° WNW Itremo; 20°35'36"S, 046°33'48"E; 1550 m a.s.l.; 22–26 Jan. 2003; Fisher, Griswold et al. leg.; California Acad. of Sciences; yellow pan trap; in montane rainforest; code: BLF7154 (CAS).
Distribution
(Fig.
Comments
Dinapsis albicauda is known only from the type-locality. Dinapsis albicauda is similar to D. cresta Mita & Shaw but may be distinguished by its ovipositor being white-coloured basally, and by its head being rounded in lateral view. For a full species description and more information on its distribution and distinguishing characteristics, see
Dinapsis albicoxa
Material examined
Holotype
. Madagascar • ♀; Mandraka; Feb. 1944; A. Seyrig;

Dinapsis albicoxa holotype female (
Additional non-type specimens examined
Madagascar • 1 ♀; Fianarantsoa Province, 7 km W Ranomafana,-21.263064°, 47.383935°; 1100 m a.s.ls; 22–31 October 1988; W.E. Steiner leg.; Malaise trap in small clearing; montane rain forest;
Distribution
(Fig.
Comments
Dinapsis albicoxa is currently only known from a few localities in southeast Madagascar. The holotype locality (Mandraka) is situated east of Manjakandriana (Antananarivo Province) There is a tiny settlement (-18.905361, 47.920253) 10 km east of Manjakandriana near the Mandraka Catholic church. The Mandraka River runs through this region and there is a Mandraka waterfall in the area. Seyrig may have been referring to the river/waterfall area rather than the precise settlement. Ivondro is located in the Atsimo-Atsinanana region.
Dinapsis albicoxa is a distinctive species that is named for its white hind coxa (Fig.
Dinapsis bicolor , sp. nov.
Material examined
Holotype
. South Africa • ♀; Northern Cape, Tankwa National Park, Renoster River; 490 m a.s.l.; 32°14.704'S, 20°05.824'E; 17 Aug.–9 Sep. 2014; S. van Noort leg.; Malaise trap; Acacia karoo thicket; Tanqua Wash Riviere; Succulent Karoo; TKW14-ACA1-M07; IMAGED WaspWeb LAS 4.9
Diagnosis
This species is easily recognised by the distinctly bi-coloured body (Fig.

Dinapsis bicolor van Noort & Shaw, sp. nov. holotype female SAM-HYM-P088338 (
Distribution
(Fig.
Comments
Dinapsis bicolor is probably also present in the Western Cape, as the Tankwa type locality is situated directly on the border between these two provinces. This is an arid-adapted species, collected from low rainfall regions in the Succulent Karoo and Bokkeveld Sandstone Fynbos, reflected in the orange colour of the metasoma a common colour present on the body of wasp species from arid regions.

Dinapsis bicolor van Noort & Shaw, sp. nov. holotype female SAM-HYM-P088338 (
Etymology
This species is named for the contrasting colour between the metasoma and the rest of the body. Noun in apposition.
Barcode sequence for holotype specimen
38754_A03_SAM-HYM-P088338 (sequence code in BOLD:FSA1895-21) BIN URI: BOLD:AEH7061.
Nucleotide sequence
TTTATTGGGTCGTCTATAAGTATAATTATTCGGATAGAATTAAGGGTTCCTGGTTCATTTATTGGTAATGATCAAATTTATAATTCAATTGTTACAGCTCATGCTTTTATTATAATTTTTTTTATAGTTATACCTTTTATAATAGGAGGTTTTGGTAATTGATTGTTACCATTAATATTAGGAGCTCCTGATATATCTTATCCACGTTTAAATAATTTAAGATTTTGATTATTAATTCCTTCTTTATTATTTTTATTAATAAGATTTTATGTAGGTGGTGGTACAGGTACAGGGTGAACTGTGTATCCCCCATTGTCTTCAAATATATTTCATTCTGGAATAAGGGTAGATCTATCAATTTTTAGATTACATTTAGCTGGAATTTCATCAATTTTAGGCTCAGTTAATTTTATTTCAACAATTTTAAATTTGCGTAATATTAAATTATCAGTAAGTAATTTAAGGTTATTTATTTGATCAGTGTTTTTAACAGCTATTTTATTATTATTATCTTTACCTGTATTAGCAGGGGCTATTACTATATTGTTAACGGATCGTAATTTAAATACAACATTTTTTGACCCTTCAGGGGGCGGTGATCCAATTTTATATCAACATTTATTT.
Description
Holotype female. Body length 3.5 mm excluding ovipositor.
Colour. Head and mesosoma black with minute white setae, interspersed with darker longer setae; metasoma orange-brown, dorsally darker on last three tergites. Scape, pedicel, F1, F2, fore coxa, trochanters, tibiae, tarsi, ovipositor dark orange-brown; mandibles and hind leg reddish brown. Remaining antennal segments, fore and mid coxae and femora, wing venation, and ovipositor sheath dark brown. Eyes and ocelli silvery. Wing membrane clear except two dark brown pigmented bands across forewing with the apical band extending in a lighter infuscation almost to apical margin, and all the way to posterior margin in apical 1/2 of wing.
Head oval, 1.44 × wider than height; vertex, frons, and face evenly sparsely punctate, interstices polished and 1–3 × greater than puncture width; ocelli small, OOL 3.0 × ocellar diameter; ocellar triangle equilateral; eye large and slightly protuberant, nearly parallel in anterior view, but diverging slightly ventrally; eye densely and evenly covered with minute white ocular setae; eye margined posteriorly by foveate groove; postocular orbital carina absent; antenna with 11 flagellomeres having flagellar length/width ratios as follows: F1 = 3.5, F2 = 3.0, F3-F11 = 2.5, F12 (apical flagellomere) = 3.0; apical flagellomere same width as basal flagellomeres; temple adjacent to ocular orbital carina punctate and polished, becoming areolate-reticulate towards occipital carina, temple width 0.88 × eye width in lateral view; malar space length 1.0 × mandible width basally; occiput areolate-reticulate, occipital carina wide and crenulate.
Mesosoma. Pronotum polished, laterally excavated with a row of large oblong foveae situated dorsally and posteriorly; the mesoscutal anterior plate polished, with a medial row of punctures, and a lateral carina bounding a foveate groove; mesoscutum as long as wide, mesoscutal lobes sparsely punctate, polished, antero-lateral corners smoothly rounded without tubercles; medial mesoscutal furrow jagged with foveae; transscutal articulation a smooth furrow, anterior edge crenulated, posterior edge smooth; scutoscutellar sulci comprising a line of adjacent large foveae, anteriorly meeting before reaching transscutal articulation; scutellar disc medially polished, with scattered punctures laterally; scutellar disc medially devoid of setae, peripherally rimmed with erect setae; mesopleuron antero-laterally strongly foveate and sparsely setose, medially polished with sparse punctures and setae, and with large median mid-pit; propodeum medially with strongly developed transverse carinae between submedian longitudinal carinae, progressively less transverse carinae present from central to lateral longitudinal tracks.
Legs. Apex of fore tibia with comb of 13 or 14 stout spines; hind coxa sparsely punctate, weakly covered with long, silky, white setae not obscuring surface; hind femur stout, polished, 2.6 × longer than wide, outer surface of hind femur sparsely, but evenly covered with long, erect, white setae, inner surface of hind femur polished sparsely punctate with very short setae; surface of hind tibia polished, sparsely punctate with long erect white setae dorsally and ventrally, shorter adpressed setae laterally; dorsal setae lacking spatulate tips; inner median margin of hind tibia with a dense longitudinal patch of shorter white setae; hind basitarsus long, subequal in length to remaining four tarsomeres combined; basitarsus ventrally with sparse preening brush consisting of numerous short, white setae, inclined posteriorly; basitarsus dorsally with normal hair-like setae, lacking spatulate tips; T2, T3, and T4 each short, T2 twice as long as wide, T3 and T4 ca. as long as wide; all tarsomeres with normal hair-like setae; tarsal claw simple, strongly curved.
Wings. Forewing length 2.8 mm, 2.95 × longer than wide; wing basally with cells R and 1A largely devoid of setae; 1R1 and 1M with larger, more sparse setae compared to wing apical of these cells, which is evenly covered with smaller, more dense setae; wing clear, with two dark pigmented vertical bands. Basal wing band narrowest dorsally, starting at basal corner of cell 1M and anterior end of cell R, extending ventrally to cover entire cell 2CU and 3A; apical wing band wider, starting at base of pterostigma, and anterior end of 1R1, extending apically to almost cover entire marginal cell 2R1, ventrally to almost cover entire cell 1+2RS, ventrally wider and more diffuse, with infuscate pigmentation extending across cells 2+3M and 3CU, to lower wing margin and extending towards apical margin not quite reaching wing edge; forewing venation with vein Rs apically curving abruptly towards anterior wing margin to form short, truncate marginal cell 2R1; apical segment of vein M long, extending beyond apex of marginal cell, vein M with small white bulla situated almost 3/5 of vein length. Hind wing with apical stub of vein Rs 2/3 of shortest width between the propodeal submedian longitudinal carinae.
Metasoma in dorsal view 1.47 × longer than wide, with seven dorsally visible terga, all smooth and shining except for terga 6 and 7, which are finely shagreened; exposed portion of ovipositor, in lateral view, sub-equal in length to metasomal length; ovipositor sheaths setose, strongly curled (an artefact of preservation).
Variation present in paratype females
Hind leg may be lighter reddish brown in colour than holotype. Body length 2.25–3.8 mm. Forewing length 1.9–2.8 mm.
Dinapsis centralis
Material examined
Holotype
. Central African Republic • 1 ♀; Prefecture Sangha-Mbaéré, Parc National de Dzanga-Ndoki, 38.6 km. 173˚ S. Lidjombo; 2°21.60'N, 16°03.20'E; 350 m a.s.l.; 22 May 2001; lowland rain forest; S. van Noort leg.; sweeping; CAR01-S230; SAM-HYM-P0024654;
Other material examined
Cameroon • 1 ♀; S.W. Province, Korup [5.2014°N, 8.8934°E], Big Rock Camp; 27 Dec. 1980–10 Jan. 1981; D. Jackson leg.; BMNH(E) 2007-19;
Diagnosis
In the key to African Dinapsis species by

Distribution
(Fig.
Comments
This is a rainforest and coastal forest associated species and is expected to be widespread across the central African region. The Cameroon record represents the northernmost confirmed distribution of Dinapsis in the Afrotropical region. The Uganda specimens are darker than other specimens, with the legs and metasoma being mostly black rather than dark brown. The Kenyan specimen was collected in Mrima Hill forest at an elevation of 212 m and co-occurs geographically with D. tricolor.

Dinapsis cresta
Material examined
Holotype. Madagascar • ♀; Province Diego-Suarez, Parc National Montagne d’Ambre; 12°31'13"S, 49°10'45"E; 1125 m a.s.l.; 4–19 Mar. 2001; R. Harin’Hala leg.; Malaise trap MA-01-01D-05; CASENT2009739; CAS. Paratype data listed in Mita and Shaw, 2020.
Distribution
(Fig.
Comments
Dinapsis cresta is currently known only from the type specimens from three localities in northern and eastern Madagascar. Dinapsis cresta is most similar to D. albicauda, from which it can be distinguished by its flatter face and entirely brown ovipositor. For a full species description and more information on its distribution see
Dinapsis gamka , sp. nov.
Material examined
Holotype. South Africa • ♀; Western Cape, Gamkaberg Nature Reserve; 33°43.663'S, 21°57.600'E; 940 m a.s.l.; 4 Oct. 2010–25 Jan. 2011; S. van Noort leg.; Malaise trap; Rooiberg Sandstone Fynbos; GB09-FYN1-M52; SAM-HYM-P048061;

Dinapsis gamka van Noort & Shaw, sp. nov. holotype female, SAM-HYM-P048061 (
Diagnosis
Morphologically similar to D. zulu, both species possessing long mesoscutal setae and the same wing pattern. However, the height of the head in D. gamka is distinctly taller than in D. zulu, where the head and mesosoma are of nearly equal height (the head of D. gamka is 1.5× taller than the mesosoma height). Additionally, the postocular furrow of D. gamka is noticeably more narrow than in D. zulu. Similar also to D. taita, both species having a polished mesoscutum with distinctly raised anterior knobs, but only the very peaks of the knobs may be faintly orange-brown (or may be black as in specimen from Grootbos, which also has faint, very small fovea on part of the mesoscutum) in D. gamka, whereas the mesoscutal plate and larger areas of the dorsal surface encompassing the raised knobs are distinctly reddish orange in D. taita. Head finely punctate, with moderately long setae, whereas it is distinctly rugulose in D. taita. Scutoscutellar sulcus demarcated by broad, foveate furrow, which is very narrow in D. taita. Legs and hind coxae black. Hind tibia with long setae. Forewing shiny, sparsely setose, with a characteristic broad, black distal band, centrally extending as infuscation towards the distal wing margins, but not reaching the margin on any side. Hind tibia with long black setae dispersed between smaller white setae.

Dinapsis gamka van Noort & Shaw, sp. nov. holotype female, SAM-HYM-P048061 (
Distribution
(Fig.
Etymology
This species is named after the Khoisan word (gamka) for lion. Noun in apposition.

Dinapsis gamka van Noort & Shaw, sp. nov. paratype female, Grootbos PNR, SAM-HYM-P088412 (
Description
Holotype female. Body length 4.1 mm excluding ovipositor.
Colour. Head and mesosoma black. Head with a dense covering of white setae on occiput, brown, more widely spaced setae on face and frons; mesoscutal plate black, peaks of anterio-lateral mesoscutal knobs may be faintly orange-brown; metasoma dark brown. Scape, pedicel, F1 to F4 and F8 to F12, fore coxae, trochanters, tibiae dark brown. F5, tarsi, ovipositor orange-brown; mandibles and hind leg reddish brown. F6 and F7 white forming a median anellus on antennae. Eyes and ocelli silvery. Wing membrane clear except for two broad dark brown pigmented bands across forewing with the apical band extending as an infuscation towards, but not reaching apical margin.

Dinapsis gamka van Noort & Shaw, sp. nov. paratype female, Grootvadersbosch NR, SAM-HYM-P043550 (
Head oval, 1.25 × wider than high; vertex, frons, and face evenly sparsely punctate, interstices polished and 1–3 × greater than puncture width; ocelli small, OOL 3.0 × ocellar diameter; all ocelli bounded by a semi-circular depression on the side facing outer edge of the triangle; ocellar triangle equilateral; eye large and slightly protuberant, nearly parallel in anterior view, but diverging slightly ventrally; eye densely and evenly covered with minute white ocular setae; eye margined posteriorly by foveate groove; postocular orbital carina absent; antenna with 12 flagellomeres having flagellar length/width ratios as follows: F1 = 3.5, F2-F4 = 3.0, F5-F8 and F12 = 2.5, F9-F11 = 2.0; apical flagellomere same width as basal flagellomeres; temple adjacent to ocular orbital carina sparsely punctate and polished, becoming more densely punctate towards occipital carina, temple width 0.5 × eye width in lateral view; malar length 1.3 × mandible width basally; occiput punctate; occipital carina wide and crenulate.

Mesosoma. Pronotum polished, laterally excavated with a row of large oblong foveae situated dorsally and posteriorly on the margin with the mesopleuron; medially with angled, central tri-radiating rows of foveae. Mesoscutal anterior plate polished, with a medial row of punctures, and a lateral carina bounded by a foveate groove; mesoscutum 0.88 × wider than long, mesoscutal lobes polished, with scattered long setae, antero-lateral corners tinged orange, smoothly rounded without tubercles; medial mesoscutal furrow deep and sinusoidally jagged with fovea; transscutal articulation a smooth furrow, anterior edge crenulated, posterior edge smooth; scutoscutellar sulci comprising a line of adjacent large fovea, anteriorly meeting before reaching transscutal articulation; scutellar disc medially polished, with scattered erect, long, brown setae laterally; mesopleuron antero-laterally deeply foveate with short white setae, medially polished with dense patch of long, erect white setae, with large median mid-pit. Metanotum with raised, setose medial area flanked anteriorly by narrow foveate depression laterally grading into wider, larger foveae. Propodeum medially with strongly developed transverse carinae between submedian longitudinal carinae, progressively less transverse carinae present from central to lateral longitudinal tracks. Medial track anteriorly with two deep fovea, lateral tracks each anteriorly with three or four deep foveae.
Legs. Apex of fore tibia with comb of stout spines; hind coxa sparsely punctate, densely covered with long, silky, white setae obscuring surface on ventral 1/2; hind femur stout, polished, 2.14× longer than wide, outer surface of hind femur sparsely, but evenly covered with long, erect, white setae, inner surface of hind femur polished sparsely punctate with very short setae; surface of hind tibia polished, with long erect white setae dorsally, shorter setae laterally and ventrally; dorsal setae lacking spatulate tips; inner ventral margin of hind tibia with a dense longitudinal patch of shorter whitish yellow setae; hind basitarsus long, subequal in length to remaining four tarsomeres combined; basitarsus ventrally with dense preening brush consisting of numerous short, whitish yellow setae, inclined anteriorly; basitarsus dorsally with normal long, white setae, lacking spatulate tips; T2 twice as long as wide, T3 1.8× as long as wide, T4 ca. as long as wide, T5 3 × as long as wide; all tarsomeres with normal hair-like setae; tarsal claw simple, strongly curved.
Wings. Forewing length 2.85 mm, 3.16× longer than wide; wing basally with cells R and 1A largely devoid of setae; 1R1 and 1M with very small, sparse setae compared to wing apical of these cells, which is evenly covered with small, scattered setae; wing clear and overall appearing polished, with two dark pigmented vertical bands. Basal wing band narrowest posteriorly, covering basal 1/3 of cell 1M, anterior 1/3 of cell R, and anterior 2/5 of cell 1Cu, extending posteriorly to wing margin, covering entire cell 2CU and 3A; apical wing band wider, starting at base of pterostigma, and anterior end of 1R1, extending apically to cover entire marginal cell 2R1, posteriorly to cover entire cell 1+2RS, posteriorly wider and more diffuse, with infuscate pigmentation extending across cells 2+3M and 3CU, but not reaching lower wing margin, and extending towards apical margin but not reaching wing edge resulting in a clear semi-circular band parallel to the apical wing margin; forewing venation with vein Rs apically curving abruptly towards anterior wing margin to form short, truncate marginal cell 2R1; apical segment of vein M long, extending beyond apex of marginal cell, vein M with small white bulla situated at 2/3 of vein length. Hind wing with apical stub of vein Rs 2/3 of shortest width between the propodeal submedian longitudinal carinae.
Metasoma in dorsal view 1.67 × longer than wide, with seven dorsally visible terga, all polished; exposed portion of ovipositor, in lateral view 2.0 × longer than metasomal length; ovipositor sheaths setose, strongly curled (an artefact of preservation).
Variation
There is variation in the colouring of the mesoscutum across localities, with the paratype female from Grootbos Nature Reserve having the distinctly raised anterior knobs on the mesoscutum black, and additionally this specimen has faint, very small foveae on part of the mesoscutum, which is usually polished. The Grootvadersbosch specimen has only very faint indications of brownish orange colouration on the knobs. For now, we regard this variation as being intra-specific.
Dinapsis hirtipes
Material examined
Holotype. Madagascar • ♀; Ivondro; [18.238933°S, 49.366374°E]; Museum Paris; Apr. 1941; A. Seyrig [blue labels]; Coll. Mus. Tervuren [white label]; HOLOTYPUS Dinapsis hirtipes sp. n. ♀ K-J Hedqvist det. 1967 [red label];
Distribution
(Fig.
Comments
Dinapsis hirtipes is known only from the type- locality at Ivondro, Madagascar. Ivondro is located in the Atsimo-Atsinanana region. Dinapsis hirtipes is a distinctive species with a raised (crested) vertex (Fig.

Dinapsis hirtipes holotype female (
Dinapsis igneus , sp. nov.
Material examined
Holotype. Mauritius • ♀; Black River Gorges Natl. Pk., Pétrin; 20°24'31"S, 57°28'11"E; 674 m a.s.l.; 7 Dec. 2016; S.A. Marshall leg.; debu00396997; BARCODE BOLD:FSA1893-21; IMAGED WaspWeb LAS 4.9

Diagnosis
Dinapsis igneus can be assigned to the oculohirta species group by the presence of dense ocular setae on the eyes. It can be distinguished from D. oculohirta, and others in that group, by being the only Dinapsis species with a slight metallic sheen to the integument (as seen under a microscope with illumination). The mesoscutum has a characteristic strongly raised bilobed crest situated anteriorly (Fig.

Dinapsis igneus van Noort & Shaw, sp. nov. holotype female (

Distribution
(Fig.
Comments
Known only from Black River Gorges National Park in both the montane rainforest and upland heath forest habitat. This is the first megalyrid species to be described from an island that is volcanic in origin, and not a more ancient fragment of continental rock. Previous authors have commented on the association of Megalyridae with ancient continental landmasses (

Dinapsis igneus van Noort & Shaw, sp. nov. paratype male (
Etymology
This species is named after the Latin for fiery (igneus) in reference to the provenance of the species from Mauritius, which is a volcanic island formed ca. 8 million years ago. The species epithet is to be treated as an adjective.
Barcode sequence for holotype female
38754_A01_Din_ign_fem (sequence code in BOLD: FSA1893-21) BIN URI: None (sequence too short).
Nucleotide sequence for holotype female
CTATAAGAATAATTATTCGTATGGAACTTAGAGTTCCGGGTTCATTTATTGGAAATGATCAGATTTATAATTCTATTGTGACTGCACATGCTTTTATTATAATTTTTTTTATAGTAATACCTTTTATAATGGGAGGTTTTGGTAATTGGTTATTGCCCTTAATGTTAGGGGCTCCTGATATGTCTTACCCTCGTCTTAATAATTTAAGGTTTTGATTGTTGGTTCCTTCTTTGTTATTTTTATTAATA.
Barcode sequence for paratype male
38754_A02_Din_ign_mal (sequence code in BOLD: FSA1894-21) BIN URI: None (sequence too short).
Nucleotide sequence for paratype male
TTGAGCAGGGCTTATTGGATCATCTATAAGAATAATTATTCGTATGGAACTTAGAGTTCCGGGGTCATTTATTGGGAATGATCAGATTTATAATTCTATTGTAACTGCACATGCTTTTATTATAATTTTTTTTATAGTAATACCTTTTATAATGGGAGGGTTTGGTAATTGATTATTGCCTTTGATGTTAGGGGCCCCTGATATGTCTTATCCTCGTCTTAATAATTTAAGGTTTTGATTATTGGTTCCTTCTTTGTTATTTTTACTAATAAGGTTTTATGTGGGCAGAGGTACAGGAA.
Description
Holotype female. Body length 4.0 mm excluding ovipositor.
Colour. Head and mesosoma with greenish bronze sheen. Head with a dense covering of white setae on occiput, darker, more widely spaced setae on face and frons; mesoscutal plate with a greenish bronze sheen. Mesoscutal plate with anteriodorsal bilateral peaks. Anteriolateral mesoscutal knobs absent; metasoma brown, lighter yellowish brown along posterior tergal margins. Scape, pedicel, F1, fore coxae and mid coxae, tibiae and tarsi yellowish brown. F2 and F3 light brown; F4 to F12 dark brown with light offset rows of multiporous plate sensilla; trochanters whitish yellow. Hind femur dark brown, except for light yellowish brown apex. Ovipositor orange-brown. Eyes and ocelli silvery. Wing membrane clear without dark bands.
Head round, only slightly (1.13 ×) wider than high; vertex, frons, and face evenly strongly punctate, interstices polished, 1–2 × greater than puncture width; ocelli small, OOL 2.0 × ocellar diameter; all ocelli bounded by a semi-circular depression on the side facing outer edge of the triangle; ocellar triangle isosceles (POL:LOL - 3:4); eye large and hardly protuberant, not parallel in anterior view, strongly diverging dorsally and ventrally; eye densely and evenly covered with minute white ocular setae; eye margined posteriorly by foveate groove; postocular orbital carina weakly present; antenna with 12 flagellomeres having flagellar length/width ratios as follows: F1 = 5, F2-F4 = 4.0, F5-F9 = 2.5, F10-F11 = 2.2 F12 = 2.75; apical flagellomeres distinctly wider than basal flagellomeres; temple adjacent to ocular orbital carina coarsely rugulose, temple width 0.67× eye width in lateral view; malar length equivalent to mandible width basally; occiput coarsely rugulose; occipital carina wide laterally, narrower dorsally and crenulate.
Mesosoma. Pronotum polished, laterally excavated with a row of large quadrate to oblong foveae situated posteriorly on the margin with the mesopleuron; foveae along dorsolateral margin faint and shallow; medially with angled central row of six foveae, dorsal fovea largest, 3 × length of others. Mesoscutal anterior plate polished, with a medial suture grading ventrally into a row of approximately five foveae, and lateral carinae bounded by weak foveae; glabrous except for dorsal fifth which is setose as in rest of mesoscutum; mesoscutum 1.1 × wider than long, shoulders rounded, mesoscutal lobes absent; medially mesoscutum punctate; medial mesoscutal furrow deep, narrow with weakly jagged edge; transscutal articulation a smooth, narrow furrow, anterior edge weakly jagged, posterior edge straight; scutoscutellar sulci medially comprising a continuous shallow groove with defined foveae; anteriorly meeting before reaching transscutellar articulation; scutellar disc punctate; mesonotum dorsally covered with dense white setae; mesopleuron antero-laterally shallowly foveate, dorsal fovea elongate, extending entire dorsal length of mesopleuron, with short white setae except for medially polished area surrounding large median mid-pit. Metanotum with raised, setose (long white setae as on mesoscutum) medial area flanked laterally by depression with 3–5 foveae. Propodeum medially polished with strong transverse carinae between the submedian longitudinal carinae defining the three central tracks; lateral longitudinal tracks with defined transverse carinae. All five tracks anteriorly with two or three deep foveae.
Legs. All legs with long, white setae, each seated in a darker basal socket, contrasting with surrounding pale integument creating weakly spotted appearance. Apex of fore tibia with comb of stout spines; hind coxa polished, with sparse, shorter, white setae; hind femur stout, polished, 2.3 × longer than wide, outer surface of hind femur sparsely covered with short, white setae; inner surface of hind femur polished with very short setae; surface of hind tibia polished, with long, erect white setae dorsally and ventrally, shorter setae laterally; dorsal setae lacking spatulate tips; inner ventral margin of hind tibia with a dense longitudinal patch of shorter white setae; hind basitarsus long, 1.5 × length of remaining four tarsomeres combined; basitarsus ventrally with dense preening brush consisting of numerous short, brown setae, inclined posteriorly; basitarsus dorsally with normal long, white setae, lacking spatulate tips; T2 twice as long as wide, T3 1.3 × longer than wide, T4 ca. as long as wide, T5 ca. 4 × as long as wide; all tarsomeres with normal hair-like setae, but also with scattered elongate, stronger setae projecting from dorsal surface; tarsal claw simple, strongly curved.
Wings. Forewing length 3.1 mm, 3 × longer than wide; wing surface evenly covered with small, scattered setae, including basal cells R and 1A; wing clear, without dark vertical bands. Forewing venation with vein Rs apically curving abruptly towards anterior wing margin to form short, truncate marginal cell 2R1; apical segment of vein M abbreviated, not extending beyond apex of marginal cell, vein M with small white bulla situated at a quarter of vein length. Hind wing with apical stub of vein Rs 2/3 of shortest width between the propodeal submedian longitudinal carinae.
Metasoma in lateral view 1.75 × as long as wide, with seven dorsally visible terga, all polished; exposed portion of ovipositor relatively short, in lateral view 0.76 × metasomal length; dorsal valve with 14 serrations, ventral valve smooth; ovipositor sheaths setose, strongly curled (an artefact of preservation).
Variation
Paratype male has body length 3.75 mm, and forewing length 3.0 mm.
Dinapsis luteus
Material examined
Holotype . Madagascar • ♀; Toliara Prov., Parc National de Zombitse; 19.8 km 84°E Sakaraha; 770 m a.sl.; 5–9 Feb. 2003; 22°50'36"S, 044°42'36"E; Fisher, Griswold et al.; California Acad. of Sciences; yellow pan trap, tropical dry forest; coll. code: BLF7505; CASENT2053467; CAS. Paratype data listed in Mita and Shaw, 2020.
Distribution
(Fig.
Comments
Dinapsis luteus is known from localities in southern and eastern Madagascar (Antananarivo, Fianarantsoa, Toamasina, and Toliara provinces). Dinapsis luteus is similar to D. hirtipes but can be distinguished by the absence of dark wing bands on the forewings, and instead having the wings entirely light yellow-coloured. For a full species description and more information on its distribution and distinguishing characteristics, see
Dinapsis nubilis
Material examined
Holotype
. Madagascar • ♀; Ranomafana [21.259457°S, 47.454986°E]; Museum Paris; Aug. 1940; A. Seyrig [blue labels]; Coll. Mus. Tervuren [white label]; HOLOTYPUS Dinapsis nubilis sp. n. ♀ K-J Hedqvist det. 1967 [red label];

Distribution
(Fig.
Comments
Dinapsis nubilis is only known from the type locality of Ranomafana, Madagascar. This small town lies adjacent to the Ranomafana National Park, which is in the south-eastern region of Madagascar in Haute Matsiatra and Vatovavy. Dinapsis nubilis is most closely related to D. albicoxa, which both share the unusual characteristic of having lobe-like extensions off the sides of the mesoscutum (see discussion in species-group section). Dinapsis nubilis can be distinguished from D. albicoxa (which has banded wings) by the wings being entirely dark-coloured (Fig.
Dinapsis oculohirta
Material examined
Holotype. Madagascar • ♀; Esira [24.325870°S, 46.708602°E]; Museum Paris; Sep. 1940; A. Seyrig leg. [blue labels]; Coll. Mus. Tervuren [white label]; HOLOTYPUS Dinapsis oculohirta sp. n. ♀ K-J Hedqvist det. 1967 [red label];
Additional specimens examined
Madagascar • 1 ♀; Toliara Province, Forêt de Kirindy, 15.5 km 64˚ENE Marofandilia, 26 November–3 December 2008; 20°04'07"S, 044°39'34"E, California Acad. of Sciences, B.L. Fisher leg., Malaise trap; tropical dry forest, 30 m a.s.l.; BLF-18284-56; CASENT2237108; CAS • 1 ♀; Tulear Province, Beroboka village, 45 km NE Morandava; 430 ft a.s.l.; 22–30 May 2008; 19°58.65'S, 44°39.92'E; California Acad. of Sciences; M. Irwin, R. Harin’Hala leg.;, Antsarongaza dry forest; Malaise; MG-45A-24; CASENT2187500 CAS • 1 ♂; same data except Antsarongaza gallery forest; MG-45B-26; CASENT2187503 CAS • 1 ♀; Tulear Province, Androimpano Forest, 5 km E of Itampolo; 11–18 Dec. 2009; 24°39.02'S, 43°57.79'E; M. Irwin, R. Harin’Hala leg.; Malaise; path in dry forest; 130 m a.s.l.; MG-54C-53; CASENT2187501 CAS • 1♂; Tulear Province, Beza Mahafaly Reserve, Parcelle 1 near research station; 10–29 Apr. 2002; 23°41.19'S, 44°35.46'E; M.E. Irwin, F.D. Parker, R. Harin’Hala leg.; Malaise trap in dry deciduous forest; 165 m a.s.l.; MA-02-14A-23; CASENT2187502 CAS • 1 ♀; same data except 9–20 Sep. 2002; MA-02-14A-35; CASENT2118387 CAS • 1 ♂; same data except 1–8 Feb. 2002; MA-02-14B-14; CASENT2118380 CAS • 1 ♀; Tulear Province, Berenty Special Reserve, 85 m a.s.l.; 8 km NW Amboasary; 25°00.40'S, 46°18.20'E; 26 Jan.–5 Feb. 2003; M.E. Irwin, F.D. Parker, R. Harin’Hala leg.; Malaise trap; gallery forest; MA-02-22-14; CASLOT 016980; SEM CAS • 1 ♀; same data except 24 Mar.–3 Apr. 2003; MA-02-22-20; CASENT2118383 CAS • 1 ♂; same data except 27 Dec. 2002–7 Jan. 2003; MA-02-22-11; CASENT2118382 CAS • 1 ♀; same data except 22–30 Nov. 2003; spiny forest; MA-02-22A-04; CASENT2118384 CAS.
Distribution
(Fig.
Comments
Dinapsis oculohirta is known only from southern Madagascar (Anosy and Toliara [= Tulear] provinces). Dinapsis oculohirta was named for the conspicuous dense setae that cover the compound eyes (Fig.

Dinapsis planifrons
Material examined
Holotype. Madagascar • ♀; Antsiranana; Ambato forest, 26.6 km 33°, NE Ambanja; 150 m; 8 December 2004; 13°27'52"S, 048°33'06"E; California Acad. of Sciences; B.L. Fisher; rainforest; yellow pan trap; BLF11517; CASLOT 014881; SEM CAS. Paratype data listed in Mita and Shaw, 2020.
Additional specimens examined
Madagascar • 3 ♀♀, 1 ♂, Antisirana Province, Makirovana Forest; 225 m a.s.l.; 28 Apr.–8 May 2011; 14°10'14"S, 049°57'15"E; California Acad. of Sciences; B.L. Fisher; rainforest; yellow pan trap; BLF26522; BLF27041; CASENT2237138, CASENT2237139, CASENT2237145, CASENT2237146; CAS • 1 ♂, Antisirana Province, SAVA Region, Vohemar District, Antsahabelela rain forest, 9 km SW Daraina Binara; 16–23 Feb. 2011; 13°15.03'S, 049°37.00'E; California Acad. of Sciences; M. Irwin, R. Harin’Hala leg.; Malaise; in humid forest; MG-58-20; CASENT2237021; CAS.
Distribution
(Fig.
Comments
Dinapsis planifrons is known only from forests in western and northern Madagascar (Antisirana, Fianarantsoa, Mahajanga, and Toliara provinces). Dinapsis planifrons is a very distinctive species, not likely to be confused with any other. It is the most common species of the Dinapsis hirtipes species group, known as “crested wasps” because of the smooth flat-faced head with the vertex conspicuously produced dorsally as a crested ridge. D. planifrons can be distinguished from all other species by having the hind tibia apex produced into an extended prong with long apical seta with spatulate tips. For a full species description and more information on its distribution and distinguishing characteristics, see
Dinapsis scriptus
Material examined
Holotype. Madagascar • ♀; Province Fianarantsoa, near Isalo National Park, at stream, east of Interpretive Center; 17–24 Mar. 2004; 22°37.60'S, 45°21.49'E; 750 m a.s.l.; R. Harin’Hala leg.; Malaise trap in open area; MA-02-11A-16; CASLOT 016972; CAS. Paratype data listed in Mita and Shaw, 2020.
Distribution
(Fig.
Comments
Dinapsis scriptus is known from localities in northern, central, and southwestern Madagascar (Antsiranana, Fianarantsoa, and Toliara provinces). Dinapsis scriptus is distinguished from the other Dinapsis species by the hind tibia having a well-developed distal prong with only simple setae (see couplet 14 of the key to species). A similar prong is also present on the hind tibia of D. planifrons but that species has spatulate setae on the distal prong. For a full species description and more information on its distribution and distinguishing characteristics, see
Dinapsis seyrigi
Material examined
Holotype. Madagascar • ♀; Trafonomby [24.549938°S, 46.725393°E]; Museum Paris; Aug. 1940; A. Seyrig leg. [blue labels]; Coll. Mus. Tervuren [white label]; HOLOTYPUS Dinapsis seyrigi sp. n. ♀ K-J Hedqvist det. 1967 [red label];
Additional specimens examined
Madagascar • 1 ♀; Fianarantsoa Province, 7 km W Ranomafana; [21.263064°S, 47.383935°E]; 900 m a.s.l.; 20–31 Jan. 1990; W.E. Steiner leg.; Malaise trap in small clearing; montane rain forest;
Distribution
(Fig.
Comments
The type-locality of Trafanomby is now located inside the Androhahela National Park. Dinapsis seyrigi can be easily distinguished from all other species treated in this paper by the smooth (unsculptured) condition of the gena (Fig.

Dinapsis spinitibia , sp. nov.
Material examined
Holotype. Tanzania • ♂; Tanga, Muheza Dist. Kwamgumi For. Res.; 4°57S 38°44E; 170–220 m a.s.l.; 9 Nov. 1995; Fog 18-CD, NHMD, Denmark; U. Copenhagen UDSM Canopy Fogging Project; S.H. McKamey et al. leg.; Dinapsis sp. ♂, Megalyridae; IMAGED WaspWeb LAS 4.9
Diagnosis
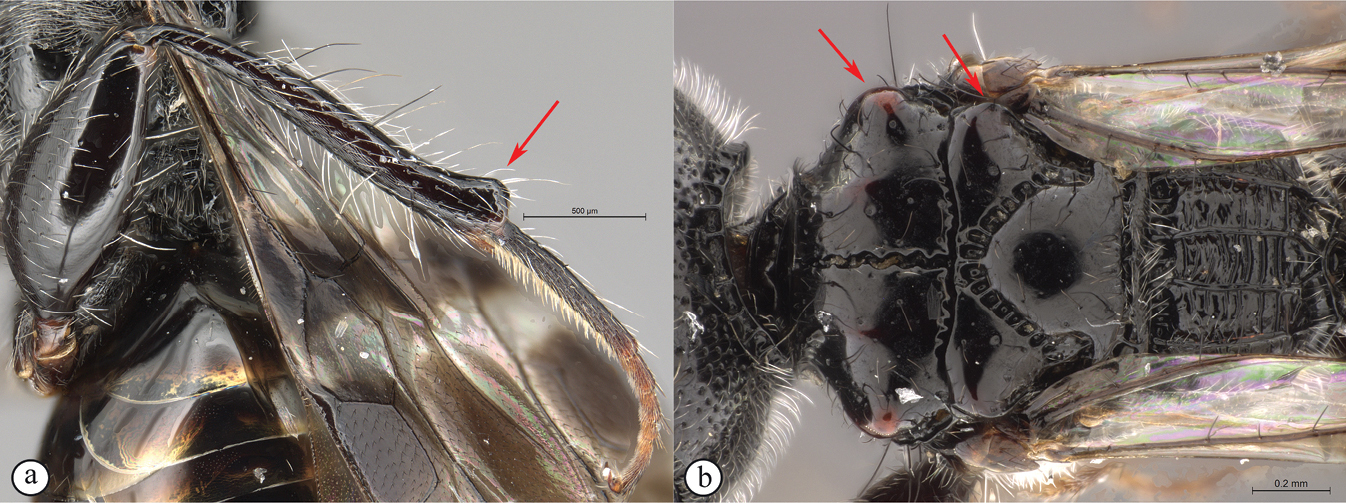
Extremely stout bodied, uniquely with two sharply elongate, parallel, stout spines positioned in close apposition on hind tibial apex (Fig.

Distribution
(Fig.

Comments
Dinapsis spinitibia is known only from the Kwamgumi Forest Reserve.

Dinapsis spinitibia van Noort & Shaw, sp. nov. holotype female (NHMD) A hind leg, antiaxial view B tibial spines, lateral view C tibial spines, basitarsus, antiaxial view D tibial spines, basitarsus, axial view E habitus, lateral view (specimen in ethanol prior to being mounted) F hind leg antiaxial view (specimen in ethanol prior to being mounted). Scale bars: 200 µm (A, B, D); 100 µm (C).
Etymology
This species is named after the sharply pointed, stout spines present on the apex of the hind tibia (Fig.
Description
Holotype female. Body length 3.6 mm.
Colour. Head orange-brown; mandibles grading to dark brown in apical 1/2. Mesosoma with black ground plan, dorsally diffused with small orange-brown areas on mesoscutum. Pronotum, mesopleuron and propodeum black. Metasoma light brown. Head with a covering of sparse white setae which are longer on face and frons. Scape, pedicel, F1 and basal F2 yellowish brown, F3–F12 orange-brown. Coxae and trochanters orange-brown, femora and tibiae dark orange-brown, tarsi yellowish brown. Eyes and ocelli silvery. Wing membrane clear except for two light brown pigmented patches extending partly across forewing as semi-bands.
Head oval, 1.25 × wider than high; vertex, frons, and face coarsely foveate; ocelli small, OOL 1.6 × ocellar diameter; all ocelli bounded by a partial semi-circular depression on the side facing outer edge of the triangle; ocellar triangle equilateral; eye large and slightly protuberant, medially parallel in anterior view, but diverging dorsally and ventrally; eye devoid of ocular setae; eye margined posteriorly by wide foveate groove bounded by strong carina; these carinae extend through genae to meet anteriorly above toruli; foveae absent in section of groove present on gena; genae massive; postocular orbital carina weakly present; antenna with 12 thin, elongate flagellomeres having flagellar length/width ratios as follows: F1 = 8.5 (very narrow basally), F2-F4 = 10.0, F5 = 6.7, F6-F7 = 6.4, F8 = 6.0, F9 = 7.0, F10 = 5.7, F11 = 5.2, F12 = 7.3; temple adjacent to ocular orbital carina coarsely foveate, temple large, width 1.4 × eye width in lateral view; malar length equivalent to mandible width basally; occiput largely polished between postocular carina and occipital carina; occipital carina evenly wide, crenulate.
Mesosoma. Pronotum polished, laterally excavated with a row of large oblong foveae situated posteriorly on the margin with the mesopleuron, no foveae along dorsolateral margin. Mesoscutal anterior plate polished, with a medial suture grading into a row of punctures, anterior plate with strong lateral carinae extending dorsally into two teeth-like projections; mesoscutum 0.9 × wider than long, with strong transverse carina extending into two lateral, dorsally projecting teeth; sparse, short setae; anterodorsally with two diffuse parallel orange-brown elongate patches; axillae and posterolaterally on the scutellum with diffuse orange-brown patches; medial mesoscutal furrow with jagged edges and foveae; transscutal articulation a smooth, narrow furrow, anterior edge largely straight except medially, posterior edge straight; scutoscutellar sulcus poorly defined, absent medially, laterally comprising three elongate foveae; scutellar disc strongly rugulose, with scattered erect, short white setae; mesopleuron coarsely foveate with short white setae, in posterior medial 1/2 polished area grading into striations anteriorly and abutting vertical row of foveae posteriorly, with large median mid-pit. Metanotum with raised, setose medial area flanked laterally by depression with 3–5 foveae. Propodeum medially polished with strong transverse carinae between the submedian longitudinal carinae defining the three central tracks; lateral longitudinal tracks with defined transverse carinae. All five tracks anteriorly with deep foveae.
Legs. All legs with white setae. Apex of fore and mid tibiae with comb of four or five stout spines; hind coxa shagreened, with sparse, small, white setae; hind femur stout, shagreened, 2.4 × longer than wide, with strong, long, erect white setae projecting from dorsal and ventral margin, outer surface of hind femur sparsely, covered with short, erect, white setae; inner surface of hind femur polished with very short setae; surface of hind tibia rugulose, with two sharp stout, but elongate black (grading to white apically) spines, in close apposition, projecting from dorsoapical margin (almost as long as basitarsus 0.83 ×); two hind tibial spurs strong, curved, longitudinally striate with minute setae; hind tibia with long, erect, white setae dorsally and ventrally, shorter setae laterally; dorsal setae lacking spatulate tips; inner ventral margin of hind tibia with a dense longitudinal patch of shorter white setae; hind basitarsus long, 1.4 × length of remaining four tarsomeres combined; basitarsus ventrally with dense preening brush consisting of numerous short, white setae, inclined posteriorly; basitarsus dorsally with two or three strong, long, erect white setae, lacking spatulate tips; T2 and T3 slightly longer than wide, T4 ca. as long as wide, T5 3.4 × as long as wide; all tarsomeres with normal small hair-like setae; tarsal claw simple, weakly curved.
Wings. Forewing length 3.15 mm, 3.3 × longer than wide; wing basally with cells R and 1A largely devoid of setae, but with widely spaced strong erect setae projecting from surrounding veins; remaining wing surface evenly, but sparsely covered with small, scattered setae; basally and medially with scattered stronger, short setae; wing clear, with two light brown pigmented patches, positioned in two vertical semi-bands. Basal wing band narrowest dorsally, covering basal 1/10 of cell 1M, posterior 1/5 of cell R, and anterior 1/8 of cell 1Cu, extending ventrally to wing margin, diffusely covering basal 1/2 of cell 2CU and all of 3A, both of which are hardly demarcated by ephemeral veins; apical wing band reduced to two patches; dorsal patch starting at base of pterostigma, and anterior end of 1R1, extending apically to cover basal posterior quarter of marginal cell 2R1, ventrally to cover almost entire cell 1+2RS, ventrally wider and more diffuse; ventral patch starting below vein M and extending to but not reaching posterior margin of wing; forewing venation with vein Rs apically curving abruptly towards anterior wing margin to form short, truncate marginal cell 2R1; apical segment of vein M long, extending beyond apex of marginal cell, vein M with small white bulla 1/3 of vein length. Hind wing with apical stub of vein Rs 2/3 of shortest width between the propodeal submedian longitudinal carinae.
Metasoma in dorsal view 1.3 × as long as wide, with seven dorsally visible terga, all polished, with row of long erect, white setae, terminally with white setae in dense patch; pygostyles long (as long as hind tarsus 2), elongate, projecting.
Dinapsis taita , sp. nov.
Material examined
Holotype. Kenya • ♀; Taita Hills, Ngangao Forest; 3.36100°S, 38.34186°E; 1848 m a.s.l.; Malaise trap; indigenous forest; 3–17 Oct. 2012; R. Copeland leg.; ICIPE 49131;
Diagnosis
Morphologically similar to D. tricolor, however, D. taita may be distinguished by its mesoscutum being more weakly foveate and having light orange-brown colour patches on the mesoscutal lobes (Fig.

Distribution
(Fig.
Comments
Dinapsis taita is known from single sites in Kenya and Burundi, the latter representing a new country record for the genus. In Kenya, the species was collected in the large, wet Ngangao forest in the Taita Hills at an elevation of 1848 m. To our knowledge this is the highest elevation of any Dinapsis species.

Dinapsis taita van Noort & Shaw, sp. nov. holotype female ICIPE 49131 (
Etymology
This species is named after the Taita Hills encompassing the holotype locality. Noun in apposition.

Barcode sequence for paratype specimen from Burundi
(sequence code in BOLD:KINS1609-11) BIN URI: BOLD:AAZ9109.
Nucleotide sequence
TTCTTTGTATTTTATTTTTGCTATTTGATCTGGTTTAATTGGATCTTCATTAAGTATAATTATTCGAATAGAATTAAGAGTTCCGGGTTCTTTTATTGGTAATGATCAAATTTATAATTCTATTGTTACAGCTCATGCATTTATTATAATTTTTTTTATAGTTATACCTTTTATAATAGGTGGATTTGGAAATTGACTTCTTCCTTTAATATTAGGGGCTCCTGATATATCTTATCCTCGTTTGAATAATTTAAGATTTTGATTATTAATCCCTTCTTTAATATTTTTGTTAATAAGATTTTATGTTGGCAGAGGAACAGGAACTGGATGAACAGTTTATCCTCCTTTATCGTCTAATACATTTCATTCTAGAATAAGAGTAGATTTATCTATTTTTAGTCTTCATATTGCTGGTATTTCATCAATTTTAGGATCTGTAAATTTTATTTCTACAATATTAAATTTACAGCATGTTTATTTAAAATTAAATAGACTTAGTTTATTTATTTGATCTGTTTTTTTGACAGCTATTTTACTTTTATTATCTTTACCTGTATTAGCAGGTGCAATTACAATATTATTGACTGATCGTAATTTAAATACAACTTTTTTTGATCCCTCGGGGGGGGGGGANCCAATTTTATATCAACNTCTTTTT.
Description
Holotype female. Body length 4.6 mm excluding ovipositor.
Colour. Head mostly black except for genae, face, clypeus and mandibles which are orange-brown. Mesosoma with black ground plan diffused with orange areas on mesoscutal knobs and mesoscutal plate, and ventral mesopleuron. Mesopleuron laterally, and pronotum diffuse orange-brown. Propodeum black. Metasoma dark brown. Head with a covering of white setae on occiput, smaller brown setae on face and frons. Scape, pedicel and F1 yellowish brown, F2–F4 dark brown, F5 basally light brown grading to whitish yellow, F6 and F7 whitish yellow and F8–F12 light brown. Coxae ventrally and trochanters white; coxae laterally and dorsally grading to orange-brown, Rest of legs yellowish brown. Ovipositor orange-brown. Eyes and ocelli silvery. Wing membrane clear except for two broad light brown pigmented bands across forewing with the apical band not extending as infuscation towards apical margin.
Head oval, 1.38 × wider than high; vertex, frons, and face coarsely punctate, interstices absent; ocelli small, OOL twice ocellar diameter; only lateral ocelli bounded by a partial semi-circular depression on the side facing outer edge of the triangle; ocellar triangle an isosceles (POL:LOL - 6:7); eye large and hardly protuberant, not parallel in anterior view, strongly diverging dorsally and ventrally; eye densely and evenly covered with minute white ocular setae; eye margined posteriorly by foveate groove; postocular orbital carina weakly present; antenna with 12 flagellomeres having flagellar length/width ratios as follows: F1 = 4.6, F2-F3 = 4.0, F4 = 3.0 F5-F11 = 2.5 F12 = 1.5; apical flagellomeres slightly wider than basal flagellomeres; temple adjacent to ocular orbital carina coarsely punctate, temple width 0.7 × eye width in lateral view; malar length equivalent to mandible width basally; occiput coarsely punctate; occipital carina wide laterally, narrower dorsally and crenulate.
Mesosoma. Pronotum polished, laterally excavated with a row of large oblong foveae situated posteriorly on the margin with the mesopleuron, no foveae along dorsolateral margin, dorsally with transverse excavation. Mesoscutal anterior plate polished, with a medial suture grading medially into a row of approximately five punctures, and a lateral carina bounded laterally by weak foveae; mesoscutum as wide as long, mesoscutal lobes polished, with scattered shallow punctures and sparse, short setae, antero-lateral areas orange, smoothly rounded except for shallow punctures; sparse shallow foveae scattered posterolaterally; medial mesoscutal furrow deep, narrow and straight edged; transscutal articulation a smooth, narrow furrow, anterior edge straight, posterior edge straight; scutoscutellar sulci medially comprising a continuous shallow groove with very weakly defined septa, laterally grading into three large separate foveae; anteriorly meeting before reaching transscutal articulation; scutellar disc medially polished, with scattered erect, white setae laterally and posteriorly; mesopleuron anteriorly shallowly foveate with short white setae, in anterior 1/2 medially polished, with large median mid-pit. Metanotum with raised, setose medial area flanked laterally by depression with 3–5 foveae. Propodeum medially polished with strongly reduced transverse carinae between the submedian longitudinal carinae defining the three central tracks; lateral longitudinal tracks with defined transverse carinae. All five tracks anteriorly with two or three deep foveae.
Legs. All legs with white setae, each seated in a dark basal socket, contrasting with surrounding pale integument creating spotted appearance. Apex of fore tibia with comb of stout spines; hind coxa polished, with sparse, small, white setae; hind femur stout, polished, 2.4 × longer than wide, outer surface of hind femur sparsely covered with short, erect, white setae; inner surface of hind femur polished with very short setae; surface of hind tibia polished, with long erect white setae dorsally and ventrally, shorter setae laterally; dorsal setae lacking spatulate tips; inner ventral margin of hind tibia with a dense longitudinal patch of shorter white setae; hind basitarsus long, 1.3 × length of remaining four tarsomeres combined; basitarsus ventrally with dense preening brush consisting of numerous short, white setae, inclined posteriorly; basitarsus dorsally with normal long, white setae, lacking spatulate tips; T2 and T3 twice as long as wide, T4 ca. as long as wide, T5 4.5 × as long as wide; all tarsomeres with normal hair-like setae, but also with scattered elongate, stronger setae projecting from dorsal surface; tarsal claw simple, strongly curved.
Wings. Forewing length 3.3 mm, 3 × longer than wide; wing basally with cells C, R and 1A with longer, more sparsely spaced setae; remaining wing surface evenly covered with small, scattered setae; wing clear, with two light brown pigmented vertical bands. Basal wing band narrowest medially, covering ventral medial portion of cell C, basal 1/2 of cell 1M, anterior ends of cells R and 1Cu, extending ventrally to wing margin, covering all of cell 2CU and 3A, both of which are only weakly demarcated by ephemeral veins; apical wing band wider, starting at base of pterostigma, and anterior end of 1R1, extending apically to cover entire marginal cell 2R1, posteriorly to cover entire cell 1+2RS, ventrally wider and more diffuse, with infuscate pigmentation extending across cells 2+3M and 3CU, and reaching lower wing margin; light infuscation covering medial section of apical wing area; forewing venation with vein Rs apically curving abruptly towards anterior wing margin to form short, truncate marginal cell 2R1; apical segment of vein M long, extending beyond apex of marginal cell, vein M with small white bulla situated at mid length of vein. Hind wing with apical stub of vein Rs 2/3 of shortest width between the propodeal submedian longitudinal carinae.
Metasoma in dorsal view 2 × as long as wide, with seven dorsally visible terga, all polished; exposed portion of ovipositor, in lateral view 2.1 × longer than metasomal length; dorsal valve with approximately 14 serrations, ventral valve smooth; ovipositor sheaths setose, strongly curled (an artefact of preservation).
Dinapsis tricolor , sp. nov.
Material examined
Holotype. South Africa • ♀; KwaZulu-Natal, Louwsberg, Sanyati Farm; 1090 m a.s.l.; 27°34'S, 31°17.9'E; 30 Oct.–18 Dec. 2005; Malaise trap (MTR); M. Mostovski leg.; NMSA-HYM 002030; NMSA. Paratypes. South Africa • 2 ♀♀; same data as holotype NMSA-HYM 000547; NMSA-HYM 000548; NMSA • 2 ♀♀; La Mercy, site 1;85 m a.s.l.; 29°37'41.0"S, 31°06'45.4"E; yellow pan trap; M. Mostovski leg.; NMSA-HYM 000545; NMSA-HYM 000546; NMSA • 1 ♂; Eshowe, nr. Ntumeni N.R.; 680 m a.s.l.; 28°52'08"S, 31°22'41"E; 20 Apr.–26 Oct. 2007; Malaise trap; Kolyada and Mostovski leg.; NMSA-HYM 000544; NMSA • 2 ♀♀; KwaZulu-Natal, Ndumo Game Reserve; [26.881013°S, 32.252109°E]; 7–8 Nov. 2002; C. Desjardins leg.; yellow pan trap;


Diagnosis
Morphologically similar to D. taita, however, D. tricolor may be distinguished by the mesoscutum being coarsely foveate and usually entirely black (Figs

Distribution
(Fig.
Comments
Dinapsis tricolor is only known from forested localities in Kenya and South Africa but is likely to be present all along the East African coastal regions from Kenya to South Africa, a sub-biogeographical unit comprising similar faunal and floral assemblages throughout the region. All 14 Kenyan specimens of D. tricolor were collected in wet, canopy-forest habitats. Four of the sites are small relict coastal forests (Mrima Hill, Muhaka, Buda, and Longomwagandi forest in the Shimba Hills National Park). The latter is under the jurisdiction of the Kenya Wildlife Service while the others are the responsibility of the Kenya Forest Service. Mrima, Muhaka and Buda are also kayas, sacred forests where cultural and religious activities are still held. As such they also receive a measure of protection from encroachment through the overview of local elders. Kasigau mountain forms, along with the nearby Taita Hills, the northernmost extension of the eastern arc mountains, a chain of ancient non-volcanic crystalline mountains centred mostly in Tanzania. Dinapsis tricolor was also collected in riverine forest at the base of the Taita Hills. This species was also found in the Kenyan highlands in Njuki-ini forest, once continuous with the great forests of Mount Kenya but now cut off because of population pressures. The Gwasi hills site is on the edge of Lake Victoria in western Kenya. The forests on the upper side and the top of the hill have been largely destroyed as a result of agricultural expansion. Finally, Marsabit mountain in northern Kenya is essentially a desert mountain, but as the only geological feature of any great elevation in the xeric north, any moisture that accompanies easterly winds tends to precipitate out over the mountain. Additional water comes from the frequent mists that cover the entire mountain and descend to the forest floor at certain times of the year. Within Kenya, then, D. tricolor has by far the widest geographic distribution of Dinapsis species, found from Taita Hills in the south to Marsabit Mountain in the north, and from the Kenyan coast in the east to the shores of Lake Victoria in the west. Its altitudinal distribution is also great, with specimens collected from near sea level to 1471 m (Njuki-ini Forest) and ca. 1500 m in the western Gwasi hills.

Dinapsis tricolor Shaw & van Noort, sp. nov. paratype female, Kenya ICIPE 49121 (ICIPE) A habitus, dorsal view B habitus, lateral view C head, mesosoma, dorsal view D head, mesosoma, lateral view. E head, anterior view F head, dorsal view. Scale bars: 2000 µm (A, B); 500 µm (C, D); 200 µm (E, F).
Etymology
Dinapsis tricolor is named for its distinctive tri-coloured antennae (Fig.

Dinapsis tricolor Shaw & van Noort, sp. nov. paratype female, Kenya ICIPE 49121 (ICIPE) A mesosoma, dorsal view B mesosoma, lateral view C metasoma, dorsal view D metasoma, lateral view E hind and mid legs, antiaxial view F wings, dorsal view (inset: data labels). Scale bars: 200 µm (A, B); 500 µm (C–F).
Barcode sequence for paratype specimens ICIPE 49121
specimen code: 38754_A07_

Dinapsis tricolor Shaw & van Noort, sp. nov. paratype male, Kenya ICIPE 49129 (
Nucleotide sequence for ICIPE 49121
AATAGAATTAAGAGTCCCAGGTTCTTTTATTGGTAATGATCAAATTTATAATTCTATTGTGACTGCTCNTGCTTTTATTATAATTTTTTTTATAGTTATACCATTTATAATGGGGGGATTTGGAAATTGACTTCTTCCCTTAATATTAGGAGCTCCGGATATATCTTATCCTCGTCTAAATAATCTGAGATTTTGATTATTAATTCCTTCTTTAATATTTTTATTAATAAGATTTTATATTGGTAGAGGAACAGGAACAGGATGAACTAT.
Barcode sequence for paratype specimen NMSA-HYM-000546
specimen code: 38754_A08_NMSA-HYM-000546 (sequence code in BOLD: FSA1900-21) BIN URI: None (sequence too short).
Nucleotide sequence for NMSA-HYM-000546
TCTTTAAGATTATTAATTCGAGCTGAATTAGGAAGTCCAGGATCTTTAATTGGGGATGATCAAATTTATAATACTATTGTAACAGCTCATGCTTTTATTATAATTTTTTTTATAGTTATACCTATTATAATTGGGGGGTTTGGAAATTGATTAGTACCCCTAATATTAGGGGCCCCTGATATAGCATTCCCTCGAATAAATAATATAAGATTTTGACTTTTACCACCCTCTATTACTCTCTTAATCTCCAGAAGAATCGTAGAAAATGGGGCTGGAAC.
Description
Holotype female. Body length 4.5 mm excluding ovipositor.
Colour. Body mostly black to dark brown with sparse minute white setae. Antenna distinctively tri-coloured, with scape, pedicel, and F1-F2 orange; F3-F4, F5 basally, and F8–F12 dark brown; and pale brownish white flagellar band on F5 apically, all of F6, and F7. Mandible, wing venation, metasomal sternites, hypopygium apically, ovipositor and sheath dark red-brown. Fore and mid femora and tibiae red-brown to bright orange. Trochanters, trochantellus and coxae (except hind coxae which are dark brown to black basally), white, orange or yellow. Hind femur and tibia black to dark red-brown, apices may be orange-brown. Eyes and ocelli silvery. Wing membrane clear except two dark brown pigmented bands across forewing.
Head round, 1.11 × wider than height; vertex, frons, and face evenly foveate-reticulate; ocelli small, OOL 1.80 × ocellar diameter; ocellar triangle equilateral; eye large and slightly protuberant, nearly parallel in anterior view; eye evenly covered with distinct ocular setae; eye margined posteriorly by coarsely foveate groove and two distinct postocular orbital carinae; antenna with 12 flagellomeres having flagellar length/width ratios as follows: F1 = 5.0, F2 = 4.0, F3 = 3.5, F10 = 2.5, F11 = 2.5, F12 (apical flagellomere) = 3.75; apical flagellomere not wider than basal flagellomeres; temple areolate-reticulate, temple width 1.25 × eye width in lateral view; gena areolate-reticulate medially and ventrally; malar space length 1.0 × mandible width basally; border of occipital carina coarsely foveate.
Mesosoma. Pronotum punctate and finely setose medio-laterally, except laterally with coarsely foveate median depression and posteriorly with evenly foveate margin; mesoscutum as wide as long, mesoscutal lobes coarsely foveate, with sparse punctures and scattered erect setae, anterolateral corners lacking tubercles; median mesoscutal sulcus and scutoscutellar sulci finely and evenly foveate; transscutal articulation narrow, finely foveate; scutellar disc medially smooth and shining, with scattered punctures laterally; scutellar disc medially devoid of setae, laterally rimmed with small erect setae, and posteriorly with shorter, denser, depressed setae; axillae smooth with scattered punctures and sparse small erect setae; mesopleuron mostly rugo-punctate and evenly setose, anterior border foveate, disc with large median mid-pit, and distinctly depressed ventro-medially to conform to meso-femur shape; propodeum medially with weakly-developed transverse carinae between longitudinal carinae, submedian longitudinal carinae posteriorly diverging slightly towards middle of propodeum.
Legs. Apex of fore tibia with comb of eight stout spines; hind coxa smooth to finely shagreened, weakly covered with long, silky, white setae not obscuring surface; hind femur stout, 2.2 × longer than wide, outer surface of hind femur sparsely but evenly covered with long, erect, silky white setae, inner surface of femur smooth, shining, and mostly devoid of setae; surface of hind tibia smooth, tibia dorsally, laterally, and ventrally with long, silky white setae, dorsal setae longer and black but hair-like, lacking spatulate tips; inner median margin of hind tibia with a dense longitudinal patch of shorter white setae; hind basitarsus long, distinctly longer than remaining four tarsomeres combined; basitarsus ventrally with dense preening brush consisting of numerous short, white setae, inclined posteriorly; basitarsus dorsally with normal hair-like setae, lacking spatulate tips; T2, T3, and T4 each shorter than preceding tarsomere, with T4 being the shortest; T2 ca. 1.5 × longer than T3, and ca. 2 × longer than T4; T2–T5 with short normal hair-like setae; T4 short and compact, length equal to width; tarsal claw simple, strongly curved.
Wings. Forewing length 3.0 mm; wing covered with scattered setae, less densely setose basally, more densely and evenly setose apically; wing clear with two vertical darkly pigmented bands. Basal wing band narrowest, starting at basal corner of cell 1M, extending ventrally to cover most of cell 2CU and 3A; apical wing band wider, starting at base of pterostigma, densely covering entire marginal cell 2R1, extending apically well beyond marginal cell and diffusely approaching wing apex, ventrally covering entire cell 1+2RS, with pigmentation extending across cells 2+3M and 3CU, to lower wing margin; forewing venation with vein Rs apically curving abruptly towards anterior wing margin to form very short, truncate marginal cell 2R1; apical segment of vein M long, extending slightly beyond apex of marginal cell, vein M with small white bulla situated at mid length of vein. Hind wing with apical stub of vein Rs very short, equal to ½ shortest width between the propodeal submedian longitudinal carinae basally.
Metasoma in dorsal view 2.9 × longer than wide, with seven dorsally visible terga, all smooth and shining dorsally, finely shagreened laterally; exposed portion of ovipositor, in lateral view, 1.62 × longer than metasoma length; ovipositor sheaths minutely setose, strongly curled (an artefact of preservation), appearing much shorter than ovipositor due to post-mortem curling.
Variation in paratype females
Body length 3.4–5.5 mm. Forewing length 2.5–4.6.0 mm. Colour of F2 varies from dark brown to orange. Apical ½ to ¾ of F5 white. Ovipositor length varying from 1.6–2.0 × metasoma length. The ovipositor sheaths are normally as long as the ovipositor but may appear more or less much shorter due to post-mortem curling; however, one specimen has short sheaths that are < ¼ the ovipositor length (possibly a developmental abnormality, or as a result of damage). The Gai Hill specimen has more brightly coloured legs than the rest of the type series: fore leg coxa and trochanter very bright yellow, rest of leg bright orange; mid leg coxa orange/brown, trochanter bright yellow, rest of leg bright orange; hind leg coxa bright yellow except basally black, trochanter bright yellow, femur and tibia dark brown except the very apex of each orange/brown, tarsi light orange.
Variation in paratype males
Body length 3.0–3.6 mm. Forewing length 2.4–2.6 mm. Antenna either entirely black, or with scape, pedicel, and F1-F2 dark brown, remainder of flagellomeres black without white band on F6-F7. Hind coxa mostly white, but black basally, or completely dark brown.
Dinapsis turneri
Material examined
Holotype
. South Africa • ♀; [Western Cape], Cape Province, Ceres; [33.358903°S, 19.298398°E]; Feb. 1921; R.E. Turner leg.; Brit. Mus.; 1921-115; Dinapsis turneri, Waterst. ♀; B.M. TYPE HYM 3,a,313,
Additional material
South Africa • 1 ♀; Eastern Cape, Asante Sana Game Reserve; 32°16.762'S, 24°57.309'E; 1186 m a.s.l.; 7 Apr. –28 Jul. 2010; S. van Noort leg.; Malaise trap; Southern Karoo Riviere Riverine Woodland; ASA09-WOO1-M10; SAM-HYM-P043547;
Diagnosis
Scutoscutellar sulci meet transscutal articulation independently. The propodeum is short (twice as wide as long; Fig.

Dinapsis turneri holotype female
Distribution
(Fig.
Comments
Dinapsis turneri is associated with the Fynbos biome in the Western Cape and the Savanna biome in the Eastern Cape and appears to not be a forest-associated species.
Revised description based on holotype and freshly collected specimens
Female. Body length 3–4 mm excluding ovipositor.
Colour. Head and mesosoma black. Head with a dense covering of white setae on occiput, brown, more widely spaced setae on face and frons (rubbed off on holotype); mesoscutal plate black, mesosoma black; metasoma dark brown. Scape and pedicel orange-brown, F1–F4 and F8–F12 dark brown, F5–F7 yellowish brown forming a median anellus on antennae. Trochanter and trochantellus whitish yellow, rest of legs black, except tarsi which are lighter. Ovipositor orange-brown; mandibles reddish brown. Eyes and ocelli silvery. Wing membrane clear except for two dark brown pigmented bands across forewing, apical band wide, basal band narrow ca. 1/3 width of apical band with the apical band extending slightly as a medial infuscation towards apical wing margin.

Dinapsis turneri holotype female
Head oval, 1.25 × wider than high; vertex, frons, and face evenly sparsely punctate, interstices polished and 1–3 × greater than puncture width; ocelli small, OOL twice ocellar diameter; all ocelli bounded by a semi-circular depression on the side facing outer edge of the triangle; ocellar triangle an isosceles (LOL 1.25 × LOL); eye large and slightly protuberant, inner margins nearly parallel in anterior view, but diverging slightly ventrally; eye evenly covered with minute white ocular setae; eye margined posteriorly by foveate groove; postocular orbital carina absent; antenna with 12 flagellomeres having flagellar length/width ratios as follows: F1 = 6, F2 = 5.5, F3 = 5.0, F4 = 5.0, F5-F7 = 4.0, F8 = 4.5, F9 to F11 = 4.0, F12 = 4.5; apical flagellomere wider than basal flagellomeres; temple adjacent to ocular orbital carina punctate-rugulose, temple width 0.75 × eye width in lateral view; malar length 1.5 × mandible width basally; occiput punctate-rugulose in contrast to the polished, sparsely punctate vertex; occipital carina wide and crenulate with large fovea.

Mesosoma. Pronotum medially polished, laterally excavated with a row of large oblong foveae situated dorsally, and posteriorly on the margin with the mesopleuron; medially with angled, central tri-radiating rows of foveae, dorsal two arms of fovea joining with dorsal row of fovea circumscribing a medial polished excavation. Mesoscutal anterior plate polished, with a medial row of punctures, and a lateral carina bounded by a shallow foveate groove; mesoscutum 1.18 × wider than long, mesoscutal shoulders evenly rounded, polished, with scattered long setae; mesoscutal lobes hardly evident; medial mesoscutal furrow deep and sinusoidally jagged with fovea; transscutal articulation a smooth furrow, anterior edge crenulated, posterior edge straight; scutoscutellar sulci comprising a line of adjacent large fovea, meeting transscutal articulation independently; scutellar disc medially polished, with scattered erect setae; mesopleuron in lateral view shallowly foveate on edges, except for posterior ventral margin, medially polished with dense long, white setae covering most of mesopleuron, except for glabrous posterior patch; with large median pit. Metanotum with raised, setose medial area flanked anteriorly by narrow foveate depression laterally grading into wider, larger foveae. Propodeum medially with strongly developed transverse carinae between submedian longitudinal carinae, fewer transverse carinae present in lateral longitudinal tracks. Medial track anteriorly with two deep fovea, lateral tracks each anteriorly with two or three deep foveae.

Dinapsis turneri holotype female slide-mount
Legs. Apex of fore tibia with comb of stout spines; hind coxa sparsely punctate, densely covered with long, silky, white setae obscuring surface on ventral 1/2; hind femur stout, polished, 2.8 × longer than wide, outer surface of hind femur sparsely, but evenly covered with short, erect, white setae, inner surface of hind femur polished, sparsely punctate with very short setae; surface of elongate (7.5 × longer than wide) hind tibia polished, with long erect white setae dorsally and fewer large black setae scattered between the white setae, shorter setae laterally and ventrally; dorsal setae lacking spatulate tips; inner ventral margin of hind tibia with a dense longitudinal patch of shorter whitish yellow setae; hind basitarsus long, subequal in length to remaining four tarsomeres combined; basitarsus ventrally with dense preening brush consisting of numerous short, whitish yellow setae, inclined anteriorly; basitarsus dorsally with normal scattered long, white setae, lacking spatulate tips; T2 and T3 1.3 × as long as wide, T4 as long as wide, T5 4 × as long as wide; all tarsomeres with normal hair-like setae; tarsal claw simple, strongly curved.

Wings. Forewing length 2.9 mm, 2.9 × longer than wide; wing basally with cells R and 1A largely devoid of setae; 1R1, 1M and 2CU with very small, sparse setae compared to wing apical of these cells, which is evenly covered with small, scattered setae; wing clear with two dark pigmented vertical bands. Basal wing band much narrower than apical band, narrowest dorsally widening progressively towards ventral wing margin, covering basal 1/2 of cell 1M, anterior eighth of cells R and 1Cu, extending ventrally to wing margin, covering entire cell 2CU and 3A; apical wing band wider, starting at base of pterostigma, and anterior 1/3 of 1R1, extending apically to cover entire marginal cell 2R1, ventrally to cover entire cell 1+2RS, medially more diffuse, slightly extending towards apical wing margin; extending ventrally with infuscate pigmentation across cells 2+3M and 3CU, to lower wing margin; forewing venation with vein Rs apically curving abruptly towards anterior wing margin to form short, truncate marginal cell 2R1; apical segment of vein M long, extending beyond apex of marginal cell, vein M with small white bulla situated at 0.425× vein length. Hind wing with apical stub of vein Rs 2/3 of shortest width between the propodeal submedian longitudinal carinae.

Metasoma in dorsal view 1.57 × longer than wide, with seven dorsally visible terga, all polished; pygostyles long, setose, pale yellow; exposed portion of ovipositor, in lateral view 2.0 × longer than metasomal length; ovipositor sheaths setose, strongly curled (an artefact of preservation).
Variation
Male much smaller than female, 2.0 mm body length; flagellum uniformly yellowish brown without medial lighter coloured band section.
Dinapsis zulu , sp. nov.
Material examined
Holotype. South Africa • ♀; KwaZulu-Natal, PMB, Karkloof, 1325 m a.s.l., 29°19.1'S, 30°15.5'E, 22 Jan.–6 Feb. 2007; YPT (yellow pan trap); Kolyada & Mostovski leg.; NMSA-HYM002031; NMSA. Paratypes. South Africa • 7 ♀♀ 1 ♂; same data as holotype; NMSA-HYM000536 to 543; NMSA • 1 ♂; same data as holotype, except for 22 Sep.–3 Oct. 2005; M. Mostovski leg.; NMSA-HYM000535; NMSA • 1 ♀; Cathedral Peak N.R., Rainbow Gorge; 1480 m a.s.l.; 28°57.60'S, 29°13.61'E; 22 Sep.–17 Nov. 2006; MT [Malaise trap]; M. Mostovski leg.; NMSA • 1 ♀; Karkloof [29.297268°S, 30.231318°E]; 48 km NE Pietermaritzburg; mist forest; 27 Nov.–11 Dec. 1991; A. Mitchell leg.; Malaise trap; NMSA • 2 ♂♂; Eastern Cape, Katberg [32.473670°S, 26.670881°E]; 4,000 ft a.s.l.; 1–15 Jan. 1933; R.E. Turner leg.; Brit. Mus. 1933–1979; Dinapsis oculohirta species group, det. Scott R. Shaw 2015;
Diagnosis
In the key to African Dinapsis species by

Dinapsis zulu Shaw & van Noort, sp. nov. holotype female (NMSA) A habitus, lateral view (inset: data labels) B habitus, dorsal view C head, mesosoma, lateral view D head, mesosoma, dorsal view E mesoscutellum, propodeum, dorsal view F habitus, ventral view. Scale bars: 1000 µm (A, B, F); 500 µm (C, D); 200 µm (E).
Distribution
(Fig.

Comments
Based on currently known data, Dinapsis zulu is associated with the forest biome, which is a naturally highly fragmented habitat in South Africa (

Etymology
Dinapsis zulu is named after the Zululand region encompassing the type locality and the Zulu people. Noun in apposition.
Barcode sequences for paratype specimen
38754_A09_NMSA-HYM-000539 (sequence code in BOLD: FSA1901-21) BIN URI: None (sequence too short).
Nucleotide sequence
GCTCATGCTTTTATTATAATTTTTTTTATAGTTATACCTTTTATAATGGGAGGTTTTGGAAATTGATTATTACCTTTAATATTAGGNGCNCCTGATATATCTTACCCCCGAATAAATAATTTAAGATTTTGATTATTA
Description
Holotype female. Body length 6.8 mm excluding ovipositor.
Colour. Body mostly black to dark brown with sparse minute white setae. Scape, pedicel, F1–F5, mandible, most of legs, wing venation, metasomal sternites, hypopygium apically, ovipositor and sheath dark reddish brown. F6-F7, trochanters, and hind coxa apically pale yellowish white. Eyes and ocelli silvery. Wing membrane clear except two dark brown pigmented bands across forewing.
Head round, 1.11 × wider than height; vertex, frons, and face shallowly and evenly punctate; ocelli small, OOL 1.8 × ocellar diameter; ocellar triangle equilateral; eye large and slightly protuberant, nearly parallel in anterior view; eye lightly and evenly covered with minute ocular setae; eye margined posteriorly by irregular and shallowly foveate groove and one distinct postocular orbital carina; second postocular orbital carina entirely absent; antenna with 12 flagellomeres having flagellar length/width ratios as follows: F1 = 5.6, F2 = 6.6, F3 = 6.6, F10 = 4.0, F11 = 4.0, F12 = 3.25; apical flagellomere slightly wider than basal flagellomeres; temple punctate, temple width 1.3 × eye width in lateral view; gena punctate medially, smooth ventrally; malar space length 1.5 × mandible width basally; border of occipital carina evenly foveate.

Dinapsis zulu Shaw & van Noort, sp. nov. paratype male
Mesosoma. Pronotum smooth, except laterally with foveate median depression; mesoscutum as long as wide, mesoscutal lobes smooth, with sparse punctures and scattered tall setae, antero-lateral corners lacking tubercles; median mesoscutal sulcus and axillular grooves narrowly and evenly foveate; scutellar furrow narrow and smooth; scutellar disc medially smooth and shining, with scattered punctures laterally; scutellar disc medially devoid of setae, laterally rimmed with large erect setae, and posteriorly with shorter, denser, depressed setae; axillae smooth with scattered punctures and sparse large erect setae; mesopleuron mostly smooth and lightly setose, medially, anterior border foveate, disc with large median mid-pit, and strongly depressed ventro-medially to conform to meso-femur shape; propodeum medially with well-developed transverse carinae between longitudinal carinae, submedian longitudinal carinae diverging posteriorly towards middle of propodeum.
Legs. Apex of fore tibia with comb of eight stout spines; hind coxa smooth to finely shagreened, weakly covered with long, silky, white setae not obscuring surface; hind femur stout, 2.5 × longer than wide, outer surface of hind femur sparsely but evenly covered with long, erect, silky white setae, inner surface of femur smooth, shining, and mostly devoid of setae; surface of hind tibia smooth, tibia dorsally, laterally, and ventrally with long, silky white setae, dorsal setae longer and black but hair-like, lacking spatulate tips; inner median margin of hind tibia lacking a dense longitudinal patch of shorter white setae; hind basitarsus long, distinctly longer than remaining four tarsomeres combined; basitarsus ventrally with dense preening brush consisting of numerous short, white setae, inclined posteriorly; basitarsus dorsally with normal hair-like setae, lacking spatulate tips; T2 and T3 each short and compact, each ca. twice as long as wide; T2 and T3 with normal hair-like setae; T4 extremely short, distinctly shorter than wide; tarsal claw simple, strongly curved.
Wings. Forewing length 4.4 mm; wing covered with scattered setae, less densely setose basally, more densely and evenly setose apically; wing clear with two dark pigmented vertical bands. Basal wing band narrowest, starting at basal corner of cell 1M, extending ventrally to cover most of cell 2CU and 3A; apical wing band wider, starting at base of pterostigma, densely covering entire marginal cell 2R1, extending apically well beyond marginal cell and diffusely approaching wing apex, posteriorly covering entire cell 1+2RS, with pigmentation extending across cells 2+3M and 3CU, to lower wing margin; forewing venation with vein Rs apically curving abruptly towards anterior wing margin to form very short, truncate marginal cell 2R1; apical segment of vein M long, extending well beyond apex of marginal cell, vein M with small white bulla situated at mid length of vein. Hind wing with apical stub of vein Rs very short, equal to ½ shortest width between the propodeal submedian longitudinal carinae basally.
Metasoma in dorsal view 3.1 × longer than wide, with seven dorsally visible terga, all smooth and shining dorsally, finely shagreened laterally; exposed portion of ovipositor, in lateral view, 1.65 × longer than metasoma length; ovipositor sheaths minutely setose, strongly curled (an artefact of preservation), appearing much shorter than ovipositor due to post-mortem curling.
Variation in paratype females
Body length 3.8–6.8 mm. Forewing length 3.0–4.4 mm. Pale band on F6-F7 varying from light yellowish white to light brown. Hind coxa varying from mostly dark brown or black to mostly yellowish white, but always dark brown to black basally and yellowish white apically. Ovipositor length varying from 1.6–2.0 × metasoma length. Head colour varying from black to reddish brown. Metasoma colour varying from black to light reddish or yellowish brown.
Variation in paratype male
Body length 3.0 mm. Forewing length 2.4 mm. Antenna entirely dark brown, without a yellowish white band on F6-F7. Hind coxa entirely dark brown to black, lacking pale colour basally.
Discussion
The family Megalyridae probably first evolved during the late Triassic period in tropical areas of the fragmenting supercontinent Pangaea, but no dated phylogeny has been produced yet. Subsequent evolution of megalyrid lineages has been shaped by the processes of rifting and continental drift among the southern continents (

Distribution maps of Dinapsis species A country occurrences for the genus Dinapsis B specimen point data of described Dinapsis species plotted on a topographical relief map C Dinapsis species occurring in Madagascar and Mauritius, excluding the D. hirtipes species-group (specimen point data plotted on a topographical relief map) D Madagascan records of Dinapsis species in the D. hirtipes species group (specimen point data plotted on a topographical relief map).
Modern megalyrid species live in southern temperate forests of Australia (

Most of the Dinapsis species are associated with forest, but in South Africa a couple of species (D. bicolor and D. gamka) are associated with the Fynbos and Succulent Karoo biomes, collectively forming the Greater Cape Floristic Region (
Although the Fynbos biome has elements that are ancient, particularly those in the families Bruniaceae and Proteaceae, diversity of this biome only escalated ca. 5 mya (
Field observations suggest that megalyrid wasps simply insert their ovipositor into pre-existing cavities, frass-filled holes, or cracks, rather than drilling into solid wood (
The currently recorded distribution for the genus Dinapsis is a highly biased artefact due to undersampling. The dominance of records from Kenya, Madagascar, and South Africa are a result of more recent, focused, long-term inventory survey efforts conducted over the last 30 years by SvN (
Acknowledgements
Thanks to Marlene McKay, Richard and Kitty Viljoen (Asante Sana Game Reserve); Derric and Nicholas Wilson (Dwarsriviershoek farm, Banghoek valley); John Moodie (Honeywood Farm); Ernest and Anne Pringle (Huntly Glen Farm); Cape Nature reserve managers, Tom Barry (Gamkaberg Nature Reserve), and Adrian Fortuin (Grootvadersbosch Nature Reserve); Deborah Winterton, Jannie du Plessis (Table Mountain National Park); Conrad Strauss (Tankwa Karoo National Park); Noel Oettlé and Bettina Koelle (WWF Avontuur Farm); and Michael Lutzeyer, Paula Strauss, Sean Privett and Gary Beukman (Grootbos Foundation) for providing permission to run long-term inventory surveys on their reserves and for logistical support in the field. Aisha Mayekiso, the late Nosiphiwo Goci, Mmamotswa Mosweu, Yvonne Samuels, the late Nkosinathi Babu, Victor Mutavhatsindi, Tiyisani Chavala, Susanna das Neves, Tiffany Wynford, and Aabid Abrahams are thanked for their dedication to processing bulk insect samples at Iziko South African Museum. Thanks to all the curators listed under museum repositories for loan of specimens. In addition, our thanks are extended to retired or previous curators, Roy Danielsson (Museum of Zoology at Lund University), Claire Villemant (Muséum National d’Histoire Naturelle, Paris), and Suzanne Lewis (Natural History Museum, London) for arranging the loan of holotypes and newly collected material. Additionally, we wish to express our grateful appreciation to Steve Marshall for providing a lovely photograph of a living Dinapsis albicoxa specimen. Cape Nature, the Eastern Cape Department of Environmental Affairs, and the Northern Cape Department of Nature and Environmental Conservation provided collecting permits to SvN. SANParks provided a research permit to SvN. SvN was funded by South African NRF (National Research Foundation) grants: GUN 2068865; GUN 61497; GUN 79004; GUN 79211; GUN 81139. Part of the South African field work conducted by SvN was funded by the National Science Foundation under PlatyPBI grant No. DEB–0614764 to N.F. Johnson and A.D. Austin. Research support for SRS was partly provided by McIntire-Stennis Grant Project number WYO-530-14, Studies of Parasitoid Wasps of Forest Ecosystems, partly by the Faculty-Grant-in-Aid program from the University of Wyoming, and partly by National Science Foundation grant DEB 14-42110, from the Dimensions of Biodiversity program. Any opinions, findings, and conclusions expressed are those of the authors and do not necessarily reflect the views of the National Science Foundation. RSC gratefully acknowledges the ICIPE core funding provided by UK Aid from the Government of the United Kingdom; Swedish International Development Cooperation Agency (SIDA); the Swiss Agency for Development and Cooperation (SDC); Federal Ministry for Economic Cooperation and Development (BMZ), Germany, and the Kenyan Government. Many thanks to Lars Vilhelmsen and Toshiharu Mita for their critical review of the manuscript and to the editor Norman Johnson for his valuable input.
References
- Baltazar CR (1962) Ettchellsia philippinensis sp. nov. (Dinapsinae, Megalyridae, Hymenoptera). Philippine Journal of Science 90: 219–220.
- Binoy C, Shaw SR, Kumar PG, Santhosh S, Nasser M (2020) First discovery of a long-tailed wasp from the Indian subcontinent (Hymenoptera: Megalyroidea: Megalyridae). International Journal of Tropical Insect Science 40(4): 751–758. https://doi.org/10.1007/s42690-020-00126-7
- Born J, Linder HP, Desmet P (2007) The Greater Cape Floristic Region. Journal of Biogeography 34(1): 147–162. https://doi.org/10.1111/j.1365-2699.2006.01595.x
- Boulinier T, Nichols JD, Sauer JR, Hines JE, Pollock KH (1998) Estimating species richness: The importance of heterogeneity in species detectability. Ecology 79(3): 1018–1028. https://doi.org/10.1890/0012-9658(1998)079[1018:ESRTIO]2.0.CO;2
- Brues CT (1923) A fossil genus of Dinapsidae from Baltic amber (Hymenoptera). Psyche (Cambridge, Massachusetts) 30(1): 31–35. https://doi.org/10.1155/1923/23910
- Brues CT (1933) The parasitic Hymenoptera of the Baltic amber. Part 1. Bernstein-Forchungen (Amber Studies), Heft 3. Walter de Gruyter and Co. Berlin.
- Cameron P (1909) Description of a new genus and species of parasitic Hymenoptera, representing a new tribe, from Kuching, Borneo. Deutsche Entomologische Zeitschrift 208–209. https://doi.org/10.1002/mmnd.48019090204
- Chen H, Liuhe B, Zhang X (2021) Two new species of the family Megalyridae (Hymenoptera) from China. ZooKeys 1043: 21–31. https://doi.org/10.3897/zookeys.1043.65223
- Deng J-Y, van Noort S, Compton SG, Chen Y, Greeff JM (2020) The genetic consequences of habitat specificity for fig trees in southern African fragmented forests. Acta Oecologica 102: e103506. https://doi.org/10.1016/j.actao.2019.103506
- Eeley HAC, Lawes MJ, Piper SE (1999) The influence of climate change on the distribution of indigenous forest in KwaZulu-Natal, South Africa. Journal of Biogeography 26(3): 595–617. https://doi.org/10.1046/j.1365-2699.1999.00307.x
- Fahringer J (1928) Die Megalyriden. Archiv für Naturgeschichte. Abteiling A 92: 98–123.
- Froggatt WW (1906) Notes on the hymenopterous genus Megalyra Westwood, with descriptions of new species. Proceedings of the Linnean Society of New South Wales 31: 399–407.
- Gauld ID (1988) Evolutionary patterns of host utilization by ichneumonoid parasitoids (Hymenoptera: Ichneumonidae and Braconidae). Biological Journal of the Linnean Society. Linnean Society of London 35(4): 351–377. https://doi.org/10.1111/j.1095-8312.1988.tb00476.x
- Gauld ID (1991) The Ichneumonidae of Costa Rica, 1. Introduction, keys to subfamilies, and keys to the specie of the lower pimpliform subfamilies Rhyssinae, Pimplinae, Poemeniinae, Acaenitinae, and Cyllocerinae. Memoirs of the American Entomological Institute 47: 1–589.
- Gauld ID, Hanson PE (1995) The evaniomorph families, Chapter 8. In: Hanson PE, Gauld ID (Eds) The Hymenoptera of Costa Rica, Oxford University Press, Oxford, 185–208.
- Gess FW (1964) The discovery of a parasite of the Phoracantha beetle (Coleoptera: Cerambycidae) in the Western Cape. Journal of the Entomological Society of Southern Africa 27: 152. https://journals.co.za/doi/10.10520/AJA00128789_3853
- Gibson GAP (1985) Some pro-and mesothoracic structures important for phylogenetic analysis of Hymenoptera, with review of terms used for the structures. Canadian Entomologist 117(11): 1395–1443. https://doi.org/10.4039/Ent1171395-11
- Hacker H (1913) Some notes on Queensland insects. Memoirs of the Queensland Museum 2: 96–100.
- Hacker H (1915) Notes on the genus Megachile and some rare insects collected during 1913–1914. Memoirs of the Queensland Museum 3: 137–141.
- Harper G, Steininger M, Tucker C, Juhn D, Hawkins F (2007) Fifty years of deforestation and forest fragmentation in Madagascar. Environmental Conservation 34(4): 325–333. https://doi.org/10.1017/S0376892907004262
- Harris RA (1979) A glossary of surface sculpturing. Occasional papers in Entomology 28: 1–31.
- Hedqvist KV (1959) Hymenoptera (Ichneumonoidea): Megalyridae. South African Animal Life 6: 485–490.
- Hedqvist KV (1967) Notes on Megalyridae [Hym. Ichneumonoidea] and description of new species from Madagascar. Annales de la Société Entomologique de France 3: 239–246.
- Huber JT, Sharkey MJ (1993) Chapter 3, Structure. In: Goulet H, Huber J (Eds) Hymenoptera of the World: an identification guide to families, Research Branch, Agriculture Canada, Ottawa, 13–59.
- Ivanova NV, Dewaard JR, Hebert PD (2006) An inexpensive, automation-friendly protocol for recovering high-quality DNA. Molecular Ecology Notes 6(4): 998–1002. https://doi.org/10.1111/j.1471-8286.2006.01428.x
- Jussila R, Kapyla M (1975) Observations on Townesia tenuiventris (Hlmgr.) (Hym., Ichneumonidae) and its hosts Chelostoma maxillosum (L.) (Hym., Megachilidae) and Trypoxylon figulus (L.) (Hym., Sphecidae). Annales Entomologici Fennici 41: 81–86.
- Kawada R, Barbosa DN, Azevedo CO (2014) Cryptalyra (Hymenoptera, Megalyridae) from Maranhão, Brazil: Three new species discovered after a large collecting effort. ZooKeys 442: 85–104. https://doi.org/10.3897/zookeys.442.8237
- Lawes MJ (1990) The distribution of the Samango monkey (Cercopithecus mitis erythrarchus Peters, 1852 and Cercopithecus mitis labiatus I. Geoffroy, 1843) and forest history in southern Africa. Journal of Biogeography 17(6): 669–680. https://doi.org/10.2307/2845148
- Lawes MJ (2004) Conservation of fragmented populations of Cercopithecus mitis in South Africa: the role of reintroduction, corridors and metapopulation ecology. In: Glenn ME, Cords M (Eds) The Guenons: diversity and adaptation in African monkeys. Springer, Boston, MA, 375–392. https://doi.org/10.1007/0-306-48417-X_24
- Lawes MJ, Eeley HAC, Findlay NJ, Forbes D (2007) Resilient forest faunal communities in South Africa: A legacy of palaeoclimatic change and extinction filtering? Journal of Biogeography 34(7): 1246–1264. https://doi.org/10.1111/j.1365-2699.2007.01696.x
- Linder HP (2003) The radiation of the Cape flora. Biological Reviews of the Cambridge Philosophical Society 78(4): 597–638. https://doi.org/10.1017/S1464793103006171
- Macdevette DR, Macdevette K, Gordon IG, Bartholomew R (1989) The floristics of the Natal indigenous forests. In Gordon IG (Ed.) Natal indigenous forest: a preliminary collection of reports on indigenous forests, Natal Parks Board, 1–20.
- Madl M (2010) A catalogue of the Megalyridae (Megalyroidea), Stephanidae (Stephanoidea) and Trigonalidae (Trigonaloidea) of the Malagasy Subregion (Hymenoptera). Entomofauna. Zeitschrift für Entomologie 31: 365–372.
- Maley J (1996) The African rain forest - main characteristics of changes in vegetation and climate from the Upper Cretaceous to the Quaternary. Proceedings of the Royal Society of Edinburgh 104B: 31–73. https://doi.org/10.1017/S0269727000006114
- Mason WRM (1993) Chapter 11, Superfamilies Evanioidea, Stephanoidea, Megalyroidea, and Trigonalyoidea. In: Goulet H, Huber J (Eds) Hymenoptera of the World: an identification guide to families. Research Branch, Agriculture Canada, Ottawa, 510–520.
- McConnell W, Kull C (2014) Protecting lemurs: Madagascar’s forests. Science 344(6182): e358. https://doi.org/10.1126/science.344.6182.358-a
- Mesaglio T, Shaw SR (2022) Observations of oviposition behaviour in the long-tailed wasp Megalyra fasciipennis Westwood, 1832 (Hymenoptera: Megalyridae). Austral Ecology 47(4): 889–893. https://doi.org/10.1111/aec.13163
- Mita T, Shaw SR (2012) A taxonomic study on the genus Ettchellsia Cameron, with descriptions of three new species (Hymenoptera, Megalyridae, Dinapsini). ZooKeys 254: 99–108. https://doi.org/10.3897/zookeys.254.4182
- Mita T, Shaw SR (2020) A taxonomic study of Dinapsis Waterson, 1922 from Madagascar (Hymenoptera: Megalyridae, Dinapsini): crested wasps of the hirtipes species-group. Zootaxa 4858(1): 71–84. https://doi.org/10.11646/zootaxa.4858.1.4
- Mita T, Konishi K, Terayama M, Yamane S (2007) Two new species of the genus Carminator Shaw from Japan, the northernmost record of extant Megalyridae (Hymenoptera). Entomological Science 10(2): 201–208. https://doi.org/10.1111/j.1479-8298.2007.00213.x
- Morley RJ (2000) The African rain forest. In: Morley RJ (Ed.) Origin and evolution of tropical rainforests. John Wiley and Sons Ltd., New York, 131–161.
- Mucina L, Geldenhuys CJ (2006) Afrotemperate, subtropical and azonal forests. Strelitzia 19: 584–614.
- Naumann ID (1983) The biology of mud nesting Hymenoptera (and their associates) and Isoptera in rock shelters of the Kakadu region, Northern Territory. In: Gillespie D et al. (Eds) The rock art sites of Kakadu National Park - some preliminary research findings for their conservation and management. Australian National Parks and Wildlife Service, Special Publication 10, ANPWS, Canberra, 129–189.
- Naumann ID (1987) A new megalyrid (Hymenoptera: Megalyridae) parasitic on a sphecid wasp in Australia. Australian Journal of Zoology 26: 215–222. https://doi.org/10.1111/j.1440-6055.1987.tb00289.x
- Penev L, Sharkey M, Erwin T, van Noort S, Buffington M, Seltmann K, Johnson N, Taylor M, Thompson F, Dallwitz M (2009) Data publication and dissemination of interactive keys under the open access model. ZooKeys 21: 1–17. https://doi.org/10.3897/zookeys.21.274
- Perrichot V (2009) Long-tailed wasps (Hymenoptera: Megalyridae) from Cretaceous and Paleogene European amber. Paleontological Contributions 1: 1–35. https://doi.org/10.17161/PC.1808.5468
- Poinar Jr G, Shaw SR (2007) Megalyra baltica Poinar and Shaw n. sp. (Hymenoptera: Megalyridae), a long-tailed wasp from Baltic amber. Zootaxa 1478: 65–68.
- Quicke DLJ (2015) The Braconid and Ichneumonid Parasitoid Wasps: Biology, Systematics, Evolution and Ecology. John Wiley & Sons, New York, 704 pp. https://doi.org/10.1002/9781118907085
- Rasnitsyn AP (1975) Vysshie pereponchatokrylye mezosoya. Trudy Paleontologischeskogo Instituta Akedemii Nauk SSR 147: 1–132. [Hymenoptera Apocrita of the Mesozoic]
- Rasnitsyn AP (1977) New Hymenoptera from the Jurassic and Cretaceous of Asia. Paleontologicheskii Zhurnal 1977: 98–108. [in Russian] [English translation in. Paleontological Journal 11: 349–357]
- Ratnasingham S, Hebert PDN (2007) Bold: The barcode of life data system (www.barcodinglife.org). Molecular Ecology Notes 7(3): 355–364. https://doi.org/10.1111/j.1471-8286.2007.01678.x
- Rodd NW (1951) Some observations on the biology of Stephanidae and Megalyridae (Hymenoptera). Australian Zoologist 11: 341–346.
- Sharkey MJ, Yu DS, van Noort S, Seltmann K, Penev L (2009) Revision of the Oriental genera of Agathidinae (Hymenoptera, Braconidae) with an emphasis on Thailand including interactive keys to genera published in three different formats. ZooKeys 21: 19–54. https://doi.org/10.3897/zookeys.21.271
- Shaw SR (1987) Three new megalyrids from South America (Hymenoptera: Megalyridae). Psyche 94(1–2): 189–199. https://doi.org/10.1155/1987/28207
- Shaw SR (1988) Carminator, a new genus of Megalyridae (Hymenoptera) from the Oriental and Australian regions, with a commentary on the definition of the family. Systematic Entomology 13(1): 101–113. https://doi.org/10.1111/j.1365-3113.1988.tb00233.x
- Shaw SR (1990a) A taxonomic revision of the long-tailed wasps of the genus Megalyra Westwood (Hymenoptera: Megalyridae). Invertebrate Taxonomy 3: 1000–1052. https://doi.org/10.1071/IT9891005
- Shaw SR (1990b) Phylogeny and biogeography of the parasitoid wasp family Megalyridae (Hymenoptera). Journal of Biogeography 17(6): 569–581. https://doi.org/10.2307/2845141
- Shaw SR (2003) A new Cryptalyra species from Colombia (Hymenoptera: Megalyridae). Zootaxa 248(1): 1–4. https://doi.org/10.11646/zootaxa.248.1.1
- Shaw SR (2005) Megalyroidea, Megalyridae. Tree of Life web project page. [Available online:] http://tolweb.org/tree?group=Megalyridae&contgroup=Apocrita
- Shaw SR, van Noort S (2009) A new Dinapsis species from the Central African Republic (Hymenoptera: Megalyridae: Dinapsini). Zootaxa 2118(1): 30–36. https://doi.org/10.11646/zootaxa.2118.1.2
- Shorthouse DP (2010) SimpleMappr, an online tool to produce publication-quality point maps. https://www.simplemappr.net [Accessed 2022/01/24]
- van Noort S (2004) Ichneumonid (Hymenoptera: Ichneumonoidea) diversity across an elevational gradient on Monts Doudou in south-western Gabon. California Academy of Sciences Memoir 28: 187–216.
- van Noort S (2022) Waspweb. https://www.waspweb.org [Accessed 2022/01/16]
- van Noort S, Picker M (2011) Wasps, Bees, Ants. Class Insecta, Order Hymenoptera. In: Picker M, Griffiths C (Eds) Alien and Invasive Animals. A South African perspective. Struik Nature, 140–146.
- van Noort S, Shaw SR (2009) Megalyridia capensis (Hymenoptera: Megalyridae: Megalyridiini) a relict species endemic to South Africa. African Natural History 5: 1–8.
- van Noort S, Belokobylskij SA, Touret-Alby A (2021) Rediscovery of the endemic Afrotropical genus Spathioplites (Hymenoptera, Braconidae, Doryctinae) with major range extension records for Spathioplites phreneticus. African Invertebrates 62(2): 497–520. https://doi.org/10.3897/afrinvertebr.62.74103
- Vilhelmsen L, Perrichot V, Shaw SR (2010) Past and present diversity and distribution in the parasitic wasp family Megalyridae (Hymenoptera). Systematic Entomology 35(4): 658–677. https://doi.org/10.1111/j.1365-3113.2010.00537.x
- von Maltitz G, Mucina L, Geldenhuys C, Lawes M, Eeley H, Adie H, Bailey C (2003) Classification system for South African indigenous forests: an objective classification for the Department of Water Affairs and Forestry. Environmentek Report ENV-P-C, 2003–017, 1–284.
- Waterston J (1922) A new family of Hymenoptera from South Africa. Annals & Magazine of Natural History 10(9): 418–420. https://doi.org/10.1080/00222932208632792
- Westwood JO (1832) In: Griffith E. Supplement on the Hymenoptera. In: Cuvier G (Ed.) Animal Kingdom, Class Insecta. Whittaker, Treacher and Co., London, 389–576.
- White F (1981) The history of the Afromontane archipelago and the scientific need for its conservation. African Journal of Ecology 19(1–2): 33–54. https://doi.org/10.1111/j.1365-2028.1981.tb00651.x
- White F (1983) Vegetation of Africa: a descriptive memoir to accompany the UNESCO/AETFAT/UNSO vegetation map of Africa. UNESCO, Paris.
Supplementary material
Appendix 1
Data type: (measurement/occurrence/multimedia/etc.)
Explanation note: Lucid Interchange Format version 3 (LIF3) of the WaspWeb online Lucid matrix identification key to the Afrotropical species of Megalyridae (Hymenoptera).The LIF3 file is an XML-based file that stores all the Lucid3 key data, allowing exchange of the key with other key developers.