Research Article |
|
Corresponding author: Michael J. Sharkey ( msharkey@uky.edu ) Academic editor: Jose Fernandez-Triana
© 2022 Michael J. Sharkey, Austin Baker, Ramya Manjunath, Paul D. N. Hebert.
This is an open access article distributed under the terms of the Creative Commons Attribution License (CC BY 4.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Citation:
Sharkey MJ, Baker A, Manjunath R, Hebert PDN (2022) Description of Chilearinus Sharkey gen. nov. and status of Nearctic Earinus Wesmael, 1837 (Braconidae, Agathidinae) with the description of new species. ZooKeys 1099: 57-86. https://doi.org/10.3897/zookeys.1099.81473
|
Abstract
The Neotropical members formerly included in Earinus Wesmael, 1837 are transferred to a new genus, Chilearinus Sharkey gen. nov. Presently three Nearctic species of Earinus are recognized, i.e., Earinus erythropoda Cameron, 1887, Earinus limitaris Say,1835, and Earinus zeirapherae Walley, 1935, and these are retained in Earinus. Earinus chubuquensis Berta, 2000 and Earinus scitus Enderlein, 1920 are transferred to Chilearinus, i.e., C. chubuquensis, and C. scitus, comb. nov. One other species is transferred to Chilearinus, i.e., Microgaster rubricollis Spinola, 1851, Chilearinus rubricollis, comb. nov. Two other Neotropical species, Earinus hubrechtae Braet, 2002 and Earinus bourguignoni Braet, 2002 were described under the genus Earinus but are here transferred to Lytopylus, L. hubrechtae, and L. bourguignoni comb. nov. Two new species of Chilearinus are described, C. covidchronos and C. janbert spp. nov. The status of Agathis laevithorax Spinola,1851, Agathis rubricata Spinola,1851, and Agathis areolata Spinola, 1851 is discussed. A neotype is designated for Earinus limitaris (Say, 1835) and diagnosed with a COI barcode. Earinus austinbakeri and Earinus walleyi spp. nov. are described. The status of both Earinus and Chilearinus in the Americas is discussed. A revised key to the genera of Agathidinae of the Americas is presented.
Keywords
Accelerated taxonomy, BIN code, COI barcode Hymenoptera, COI DNA barcode, conservation, Ichneumonoidea
Introduction
Neotropical species formerly included in Earinus Wesmael, 1837 are transferred to a new genus, Chilearinus Sharkey gen. nov. Presently three Nearctic species of Earinus are recognized, i.e., Earinus erythropoda Cameron, 1887, Earinus limitaris Say,1835, and Earinus zeirapherae Walley, 1935, and these are retained in Earinus. Earinus chubuquensis Berta, 2000 and Earinus scitus Enderlein, 1920 are transferred to Chilearinus, i.e., C. chubuquensis and C. scitus, comb. nov. One other species is transferred to Chilearinus, i.e., Microgaster rubricollis Spinola, 1851, Chilearinus rubricollis, comb. nov. Two other Neotropical species, Earinus hubrechtae Braet, 2002, and Earinus bourguignoni Braet, 2002 were described under the genus Earinus but are here transferred to Lytopylus, L. hubrechtae, and L. bourguignoni comb. nov. Two new species of Chilearinus are described, C. covidchronos and C. janbert spp. nov. The status of Agathis laevithorax Spinola,1851, Agathis rubricata Spinola,1851, and Agathis areolata Spinola, 1851 is discussed. A neotype is designated for Earinus limitaris (Say, 1835) and diagnosed with a COI barcode. Earinus austinbakeri and Earinus walleyi spp. nov. are described. The status of both Earinus and Chilearinus in the Americas is discussed. A revised key to the genera of Agathidinae of the Americas is presented.
Methods
DNA extraction and sequencing
Molecular work was carried out at the CBG using standard protocols. A leg from each frozen-then-oven-dried specimen was destructively sampled for DNA extraction using a glass fiber protocol (
The BOLD database can be used to identify specimens using the following steps: (1) navigate to the identification tab of the BOLD Systems database (http://www.boldsystems.org/index.php/IDS_OpenIdEngine); (2) paste the COI sequence of the query organism (in forward orientation) into the query box and search against the appropriate library (e.g., All Barcode Records on BOLD, Species Level Barcode Records, etc.); (3) the search results page shows the top hits based on percentage similarity starting with the closest matches (This page also provides additional information to help verify the identity of a match, such as links to the BIN where specimen data, including images, can be found, a distribution map, and a tree-based identification tool); (4) use the Tree-Based Identification button to generate a neighbor-joining tree and find the query taxon (name in red). This allows you to visualize how distant the query sequence is from the closest matches.
Taxonomic account
Chilearinus , gen. nov.
Type species
Chilearinus janbert Sharkey, sp. nov.
Etymology
A conjunction of Chile, where 90% of the species are likely to be found, and Earinus, a reference to the probable sister group of the species, based on preliminary analyses. The genus is masculine.
Diagnosis
Notauli absent; hind coxal cavities open; tarsal claws with basal lobes; second submarginal cell quadrate, never petiolate; foretibia lacking sclerotized spines/pegs; hind wing Cub strong and emanating from an angle on the basal cell. Most similar morphologically to Earinus and Lytopylus. Earinus and Chilearinus do not have overlapping distributions. The former is restricted to the Nearctic and the latter to the Neotropics; therefore, there is little chance of confusing the two. Nonetheless, the lack of pegs on the foretibia of members of Chilearinus and the morphological characters given in the key (below) can also be employed to differentiate them. Members of Lytopylus differ most significantly in that they lack vein Cub in the hind wing. See couplet 25 in the key below.
Description
Head. Lateral carina on frons (as found in members of Alabagrus) absent; interantennal space slightly raised above antennal sockets; gena not extended ventroposteriorly into sharp prominence; mandible dorsoventrally flattened (twisted); labial palpus with 4 segments, third segment slightly more than ½ length of apical segment. Mesosoma. Propleuron lacking a sharp bump; notauli absent; mesoscutum smooth with a median pit (presumably a remnant of notauli), postscutellar depression absent; propodeum mostly smooth, sometimes with weak smooth sculpture medially; sclerite between hind coxal cavities and metasomal foramen absent. Precoxal groove absent or smooth and weakly impressed. Legs. Foretibia lacking dull pegs (unlike Earinus); mid- and hind tibia with blunt apical or preapical pegs; all tarsal claws with a rounded basal lobe. Wings. Forewing RS+Ma vein mostly present but not usually completely tubular; second submarginal cell large, quadrate and usually (perhaps always) higher than long; RS of forewing complete to wing margin; hind wing r and r-m cross veins absent; hind wing vein Cub strong and emanating from an angle on the basal cell. Metasoma. First median tergite smooth, longer than apical width, lateral longitudinal carina absent or weak and short; remaining terga smooth; ovipositor ranging from as long as the body to twice the length of the body, but this is based on small sample of a few dozen species.
Biology
Unknown.
Diversity and distribution
This is a species-rich genus with hundreds of species, based on specimens identified by MS. It is widespread in Chile and southern Argentina. A few species are found at high altitudes as far north as Ecuador and Colombia.
Notes
It is almost pointless to present a morphological key to the five recognized species of Chilearinus as they represent just five species out of hundreds. Many undescribed species will undoubtedly key to these named species. The only way to handle species-rich undocumented genera such as Chilearinus is to include COI barcode data in the diagnoses. We know this diagnostic is sufficient to differentiate all but a few species of Agathidinae (
Key to the few described species of Chilearinus
| 1 | Forewing with two yellow bands | C. scitus |
| – | Forewing evenly colored, weakly infuscate | 2 |
| 2 | Mesonotum orange | C. rubricollis |
| – | Mesonotum black | 3 |
| 3 | Hind femur entirely yellow except extreme apex dorsally | C. janbert |
| – | Hind femur mostly or entirely black | 4 |
| 4 | Hind femur black except extreme apex yellow | C. chubuquensis |
| – | Hind femur entirely black | C. covidchronos |
Chilearinus covidchronos , sp. nov.
Holotype
♀, Chile, Región IX, PN Nahualbuta, 37.809°S, 73.016°W, 3680' [1122 m], 9–12.i.2000, Malaise trap, Webb and Yeates (Canadian National Collection).
Diagnosis
COI barcode. BOLD sample ID H1145. BOLD BIN code BOLD:AAV0870. GenBank Accession Code OL702761.
AATTTTATATTTTATATTTGGAATTTGATCGGGAATTTTAGGTTTATCAATAAGTTTAATTATTCGAATAGAATTAAGAGTAGGGG
GTAATTTTATTGGTAATGATCAAATTTATAATAGAATTGTNGCTGCTCATGCTTTTATTATAATTTTTTTTATAGTTATACCAATTA
TAATTGGAGGATTTGGAAATTGATTAATTCCATTAATATTGGGGGGGCCAGATATAGCTTTCCCTCGAATAAATAATATAAGATT
TTGATTATTAATTCCTTCATTATTATTATTAATTTTAAGGTCTTTAATTAATGTTGGGGTAGGTACTGGATGAACTGTTTATCCTCC
TTTATCATTAAATATAAGTCATAGTGGTATATCTGTAGATTTAGCTATTTTTTCTTTACATATTGCTGGAATTTCTTCAATTATAGGTG
CTATAAATTTTATTACAACTATTTTAAATATGTGAATAATTAATATTAAAATTGATAAAATACCTTTATTAGTTTGATCAATTTTAA
TTACGGCAATTTTATTATTATTATCTTTGCCAGTTTTAGCTGGAGCTATTACTATATTATTAACAGATCGTAATTTAAATACTAGAT
TTTTTGATCCTTCTGGAGGAGGAGATCCAATTTTATATCAACATTTATTT
Morphological diagnosis
See key.
Paratypes
None.

Chilearinus janbert , sp. nov.
Holotype
♀, Chile, Región IX, PN Nahualbuta, 37.493°S, 72.582°W, 1168 m, 8.ii.2005, Heraty, (Canadian National Collection).
Diagnosis
COI barcode. BOLD sample ID H12114. BOLD BIN: BOLD:AEM7846. GenBank Accession Code OL702760.
TTTTAGGATTATCAATAAGTTTAATTATTCGAATAGAATTAAGAGTAGGTGGTAATTTTATTGGTAATGATCAAATTTATAATAG
GATTGTNACTGCTCATGCTTTTATTATAATTTTTTTTATAGTTATACCAATTATAATTGGAGGATTTGGAAATTGATTAATTCCAT
TAATATTAGGGGGTCCAGATATAGCCTTCCCTCGAATAAATAATATAAGATTTTGATTATTAATTCCTTCATTATTATTATTAATT
TTAAGATCTTTAATTAATGTTGGAGTAGGTACTGGATGAACTGTTTATCCTCCTTTATCATTAAATATAAGTCATAGTGGTATATC
TGTAGATTTGGCTATTTTTTCTTTACATATTGCTGGAATTTCTTCAATTATAGGGGCTATAAATTTTATTACAACTATTTTAAATA
TATGAATAATTAATATTAAAATTGATAAAATACCTTTATTAGTTTGATCAATTTTGATTACAGCAATTTTATTATTATTATCTTTA
CCAGTTTTAGCTGGGGCTATTACTATATTATTAACAGATCGTAATTTAAATACTAGATTTTTTGATCCTTCTGGAGGGGGAGATCC
AATTTTATATCAACATTTATTTTGATTTTT
Morphological diagnosis
See key.
Paratypes
None.
Etymology
A conjunction of Paul Hebert and Dan Janzen in recognition of their enormous contributions towards the conservation of nature.
Earinus
Note
In the Americas, there are three previously recognized species of Earinus, i.e., E. erythropoda Cameron, 1887, E. limitaris (Say, 1835), and E. zeirapherae Walley, 1935, and here we describe two more, Earinus austinbakeri sp. nov. and Earinus walleyi sp. nov. In the Nearctic, Earinus is common and widespread with the southernmost record being the sole recognized specimen of E. erythropoda from northern Sonora state, Mexico. Earinus differs from Chilearinus in the possession of pegs/spines in the foretibia and the characters given in the key.

Based on the collection in the Hymenoptera Institute (MS’s personal collection, which will eventually be deposited in the CNC) and borrowed specimens, there are probably between eight and 12 species in the Nearctic region. They are extremely similar in color, but there are obvious differences among specimens in body dimensions, degree of punctation, color of the hind coxae, ocellar configuration, ovipositor length, length and density of setae on the ovipositor sheath, and dimensions of the first metasomal tergum. Unfortunately, these are not sufficient to allow confident delineation of species limits. For example, the differences in the key between E. limitaris and E. erythropoda are trivial. There are numerous specimens scattered over the Nearctic region that will key to E. erythropoda, but they might all be E. limitaris, or the two nominal species may be conspecific, or there may be multiple cryptic species. Likewise, there are probably a number of undescribed Nearctic species that will key to either E. zeirapherae or E. austinbakeri. In other words, the key is sufficient to discriminate among the barcoded species and E. zeirapherae but not among these and the undescribed species. The key is presented in part to satisfy the code of Zoological Nomenclature to act as a diagnosis for E. austinbakeri and E. walleyi. Only dense sampling of COI barcodes and perhaps other genes will supply the information necessary to delimit Nearctic Earinus species.
Key to the species of Earinus of North America
| 1 | A Mid- and hind coxae slightly (A) to distinctly (AA) melanic, darker than their respective femora | E. zeirapherae |
| – | B Mid- and hind coxae mostly or entirely pale (yellow to orange) concolorous with their respective femora | 2 |
 |
||
| 2(1) | A Face distinctly punctate. Body length > 6 mm (average = 6.8 mm.) | 3 |
| – | B Face mostly smooth with shallow punctation. Body length < 6 mm (average = 5.3 mm) | 5 |
 |
||
| 3(2) | A Facial punctures deeper and wider | E. limitaris variation, or perhaps E. sp. nov. |
| – | B Facial punctures shallower and narrower | 4 |
 |
||
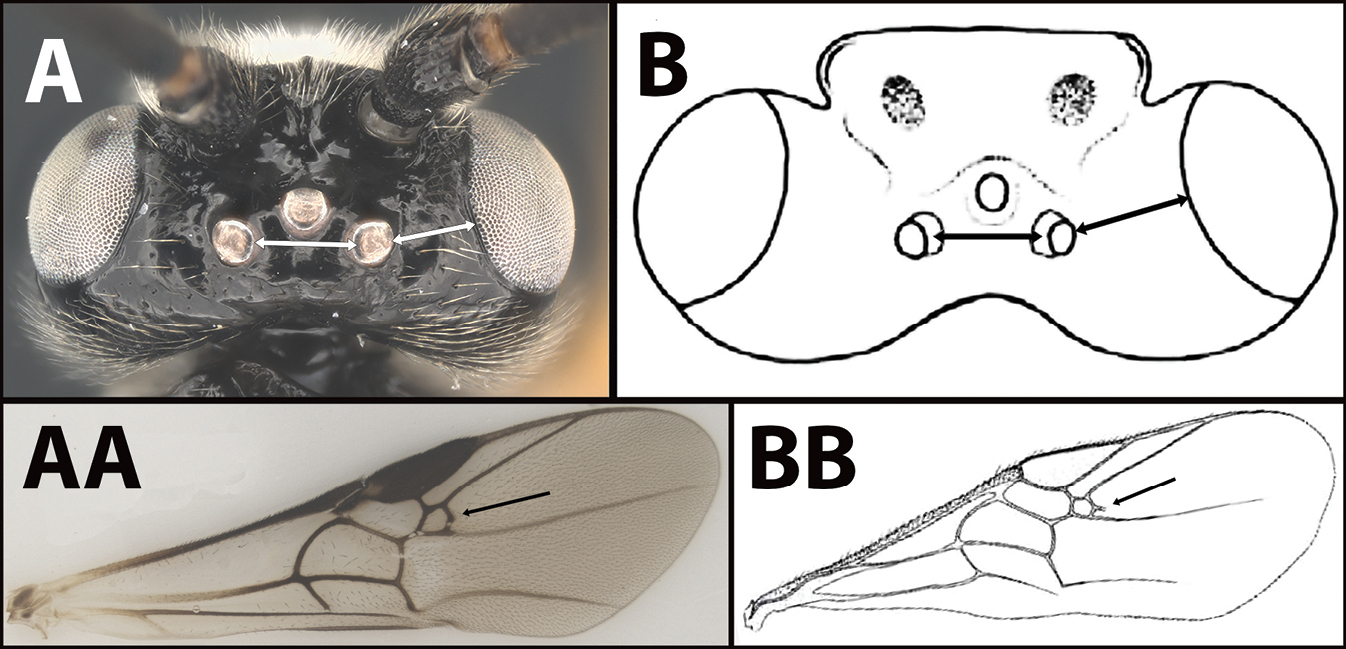
| 4(3) | A Distance between lateral ocelli longer than distance between lateral ocellus and eye. AA Second submarginal cell lacking distinct 2RS2 vein | E. limitaris |
| – | B Distance between lateral ocelli equidistant or shorter than distance between lateral ocellus and eye. BB Second submarginal cell with distinct 2RS2 vein |
E. erythropoda
.[Line drawings modified from |
 |
||
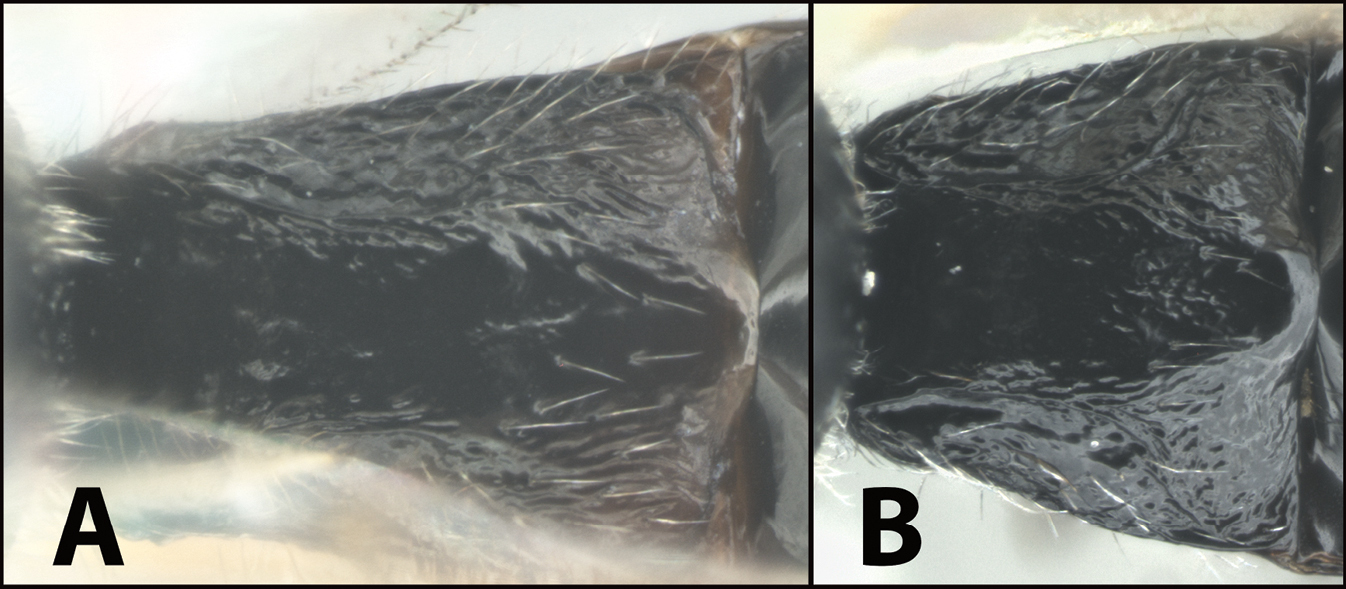
| 5(2) | A First metasomal tergum relatively longer and slimmer, distinctly longer than wide | E. austinbakeri |
| – | B First metasomal tergum relatively shorter and broader, about as long as wide | E. walleyi |
 |

Earinus austinbakeri , sp. nov.
Holotype
♀, Canada, Ontario, Ferris Provincial Park, 44.2829°N, 77.7963°W, 131 m, 05–20.Jun.2014 (Canadian National Collection). BOLD sample ID BIOUG33065-A05, BOLD BIN code BOLD:ADL5164. GenBank Accession Code OM158425.
Diagnosis
Consensus barcode based on four specimens.
ATTTTATATTTTATATTTGGGATTTGATCYGGAATTGTGGGKTTATCAATAAGTTTAATTATTCGTATGGARTTAAGAGTAGGGG
GBAATTTAATTGGKAATGATCAAATTTATAATAGTATTGTTACTGCTCATGCATTTATTATAATTTTTTTTATAGTTATRCCAATT
ATAATTGGTGGGTTTGGTAATTGGTTAATTCCTTTAATATTAGGRGGTCCCGATATRGCTTTCCCTCGAATGAAYAATATAAGRTT
TTGATTATTAATTCCTTCTTTATTATTATTAATTTTAAGATCTTTAATTAATATTGGGGTTGGAACTGGTTGAACGGTYTATCCTC
CTTTATCATTRAATATAAGTCATAGTGGTATATCTGTTGATTTGGCTATTTTYTCTTTACATATTGCGGGRATTTCTTCTATTATA
GGGGCAATAAATTTTATTACTACTATTTTAAATATATGAATAATAAATATTAAAGTTGATAAAATGTCTTTATTRATTTGATCAAT
TTTAATTACTGCTATTTTATTATTATTATCTTTACCTGTTTTAGCRGGRGCAATTACTATATTATTAACAGATCGTAATTTAAATA
CAAGATTTTTTGATCCTTCTGGAGGTGGGGATCCAATTTTATATCAACATTTATTT
Morphological diagnosis
Very similar to E. austinbakeri but differing by the characters given in the key as well as having the ovipositor sheath more setose. The COI barcodes of the two species differ by 6.29% (p-distance), reinforcing the conclusion that they are different species.

Paratypes
BIOUG01028-C01, BIOUG01028-F12, BIOUG32793-A05. These are sample IDs; the data for these specimens can be found by searching for these codes on BOLD (http://www.boldsystems.org).

Distribution
The holotype and paratypes were found at two localities just north and northeast of Lake Ontario. This species may be widespread throughout the eastern USA as far south as the Carolinas.
Etymology
Named in honor Austin Baker, hymenopterist extraordinaire.
Earinus erythropoda
Holotype
♀, “N. Sonora, Mexico, Morrison” (British Museum Natural BM3c893, viewed).
Notes
The sole identified specimen is the holotype. It differs little from many specimens that are widespread in the United States. It could be that they all belong to E. limitaris, or several more species may have similar morphologies. COI barcode data are needed. Several line drawings, modified from
Earinus limitaris
Bassus limitaris Say, 1835.
Neotype
♂, USA, West Virginia, Hardy County, 3 mi. NE Mathias, 38°55'N, 78°49'W, 30.viii–19.ix.2005 (Canadian National Collection). BOLD sample ID H1141. BOLD BIN code BOLD:AAU8493. GenBank Accession Code OM237775.
Diagnosis
Consensus COI barcode based on 9 specimens.
AATTTTATATTTTATATTTGGAATTTGATCAGGAATTTTAGGTTTATCAATAAGATTAATTATTCGAATAGAATTAAGDATAGGTG
GTAATTTRATTGGTAATGATCAAATTTATAATAGTGTTGTTYCTGCTCATGCTTTTATTATAATTTTTTTTATAGTTATACCAATTAT
GATTGGRGGRTTTGGRAATTGATTAGTTCCTTTAATATTGGGRGGTCCTGATATAGCTTTYCCTCGAATAAATAATATAAGATTTT
GATTATTAATTCCTTCTTTATTATTATTAATTTTGAGTTCTTTAATTAATATTGGGGTRGGGACTGGKTGAACAGTTTATCCTCCRT
TATCTTTAAATATAAGRCATAGTGGAATATCAGTTGATTTAGCTATTTTTTCATTACATATYGCAGGAATTTCTTCAATTATAGGGG
CAATAAATTTTATTACTACTATYATAAATATATGAATAATAAATATTAAAATTGATAAAATACCTTTATTAGTTTGATCAATTTTAA
TTACTGCTATTTTATTATTATTATCATTRCCAGTTTTAGCTGGRGCAATTACTATATTATTAACAGATCGAAATTTRAATACAAGAT
TTTTTGATCCTTCTGGAGGGGGGGATCCAATTTTATATCAACATTTATTT
Morphological diagnosis
See key.
Other specimens with barcode data
ASGLE-0444, ASGLE-0446, ASGLE-0449, ASGLE-0451, ASGLE-0452, ASGLE-0445, BIOUG01022-D11, BIOUG32892-B07. These are sample IDs; data on them can be found by searching for these codes on BOLD (http://www.boldsystems.org).
Biology
The following are listed as hosts of E. limitaris by
Notes
There are 15 specimens from one locality in Quebec that are in a different BIN (BOLD:ADF5580) which differs by only 2.54% (p-distance) from E. limitaris (Earinus sp. in Fig.
Like many of Say’s types, the type of B. limitaris is lost (
The following is from Say’s original description.
“B[assus] limitaris. Black; feet honey-yellow.
Inhabits Missouri and Indiana.
Body black: palpi white: thorax longitudinally indented behind the middle: wings nearly hyaline, at base yellowish; nervures fuscous; stigma large; first cubital cell complete; second rather large, quadrangular: radial cellule also rather large: feet honey-yellow; posterior pair of tibiae whitish, their tips and annulus near the base black; posterior pair of tarsi black.

Length seven twentieths of an inch.
Var. a. Maxillary palpi, first joint black.
♀ Oviduct hairy, decurved, somewhat robust.”
Except for the body length, this description is consistent with all of the estimated 8–12 Nearctic species of Earinus. We have a number of specimens of what we believe to be E. limitaris. The neotype was selected because it is geographically closest to the two specimens included in

Distribution
Unknown, except for barcoded specimens (West Virginia, southern Ontario), as well as either Missouri or Indiana, or both. It is unknown if
Earinus walleyi , sp. nov.
Holotype
♀, Canada, Manitoba, Churchill pump house, 15 km S Churchill, Goose Creek Road, 58.3734°N, 94.1342°W, 3–7.vii.2007, Malaise trap (Canadian National Collection). BOLD sample ID. 07PROBE-20853, BOLD BIN code BOLD:AAF9894. GenBank Accession Code FJ413805.
Diagnosis
Consensus barcode based on four specimens.
TATTTTATATTTTATATTTGGAATTTGATCAGGTATTGTAGGTTTATCAATAAGATTAATTATTCGAATGGAATTAAGAGTGGGR
GGTAATTTAATTGGRAATGATCAAATTTATAATAGTATTGTTACTGCTCATGCTTTTATTATAATTTTTTTTATAGTTATACCTAT
TATAATTGGGGGRTTTGGTAATTGATTARTCCCATTAATATTGGGAGGTCCTGATATAGCTTTCCCTCGTATAAATAATATGAGAT
TTTGATTATTAATCCCYTCTTTATTAATATTAATTTTAAGATCTTTAATTAATATTGGAGTAGGGACTGGTTGGACAGTTTATCCT
CCKTTATCATTAAATATAAGTCATAGTGGAATATCTGTTGATTTGGCTATTTTTTCTTTACATATTGCGGGRGTTTCTTCTATTAT
AGGGGCAATAAATTTTATTACTACTATTTTAAATATRTGAATAATAAATATTAAAATTGATAAAATGTCTTTATTAATTTGATCAA
TTTTAATTACTGCTATTTTATTATTATTRTCTTTACCAGTTTTAGCAGGAGCTATTACTATATTATTAACAGATCGTAATTTAAAT
ACAAGATTTTTTGATCCTTCYGGAGGGGGTGACCCAATTTTATATCAACATTTATTT
Morphological diagnosis

Paratypes
All are from the same locality as the holotype, 07PROBE-23096, 07PROBE-23097, 09PROBE-A0304. These are specimen IDs; more data on the specimens can be found by searching for these codes on BOLD (http://www.boldsystems.org).

Distribution
Unknown but likely widespread in Alaska and northern and mid-latitudinal areas of Canada. Some or all records in
Etymology
Named in honor Stuart Walley (RIP), former research scientist at the Canadian National Collection and author of E. zeirapherae.
Earinus zeirapherae
Holotype
♀, Grand River, Nova Scotia, 11.May.1932 (M. L. Prebble) No. 3847 (Canadian National Collection, viewed).
Biology
The following are all reported as hosts by
Notes
The holotype (Fig.


Key to the New World genera of Agathidinae
(Modified from
| 1 | A Forewing venation greatly reduced; RS absent and crossvein r present only as a short stub; Neotropical, rare | Mesocoelus |
| – | B Forewing venation moderately reduced; apical abscissa of RS absent, or mostly so, but crossvein r complete to junction of RS; Neotropical and rare | 2 |
| – | C Forewing venation not significantly reduced; apical abscissa of RS complete or almost complete to wing margin; widespread, common (99+ % of specimens) | 4 |
 |
||
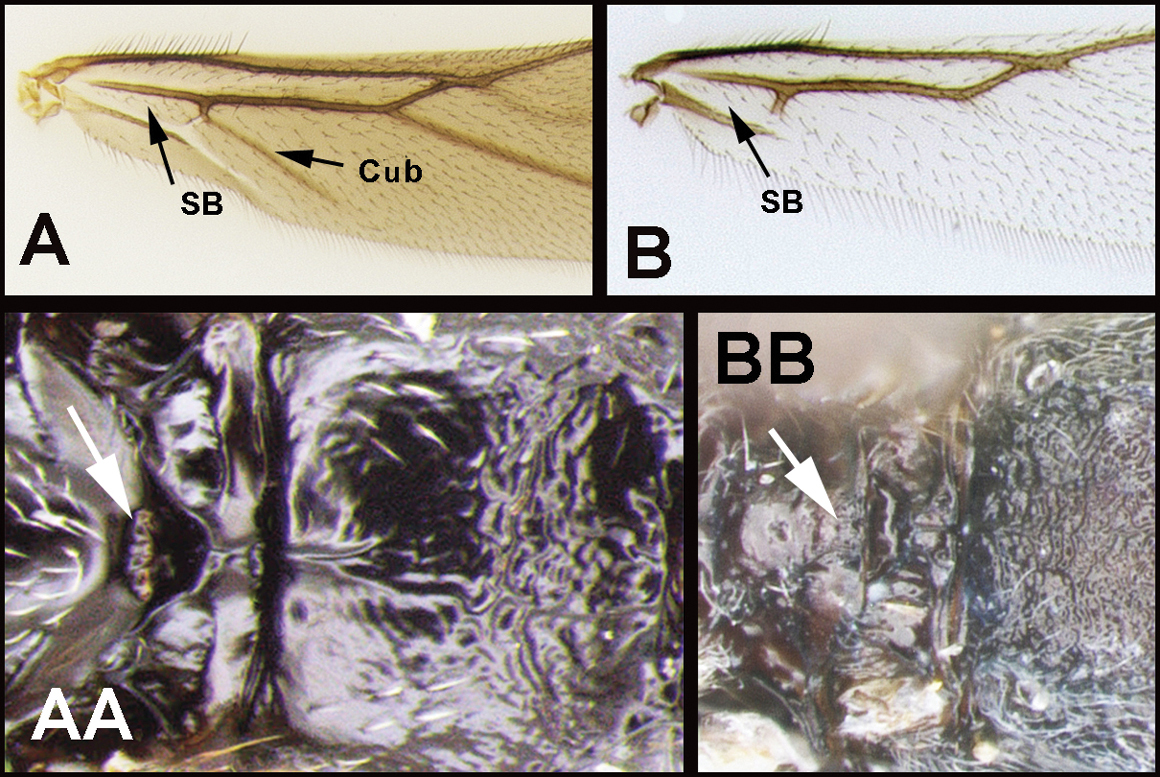
| 2(1) | A Hind wing subbasal (SB) cell 4-sided with vein Cub emanating from an angle in the cell AND/OR AA Posterior surface of scutellum with a semi-circular or arc-shaped depression (post-scutellar depression) | Therophilus (in part) |
| – | B Hind wing subbasal (SB) cell 3-sided. If Cub vein is present, it emanates from a straight vein. BB Post scutellar depression absent, but rugose sculpture usually present | 3 |
 |
||
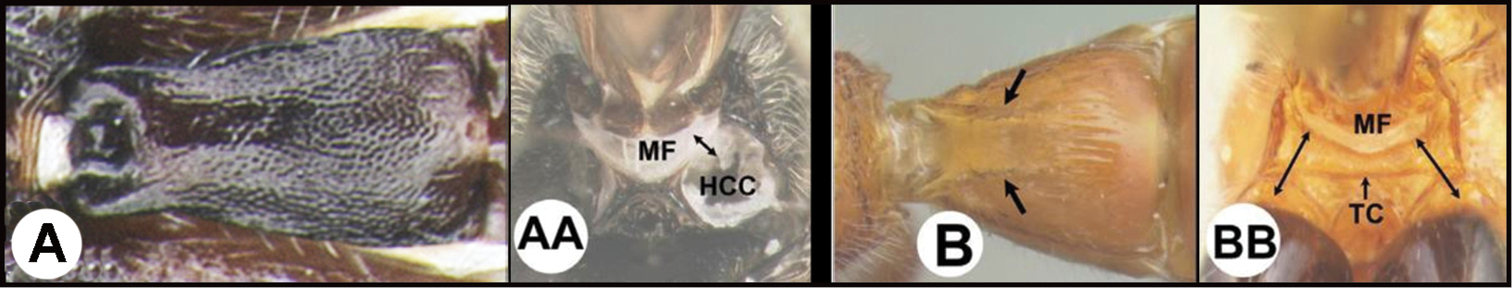
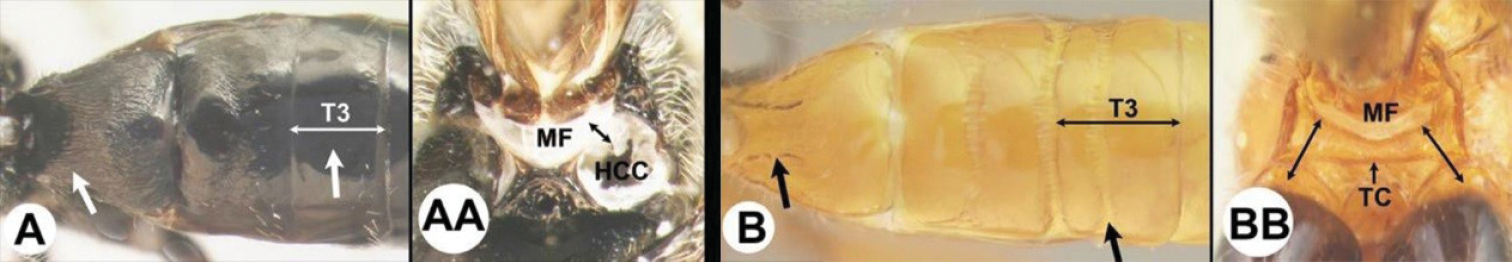
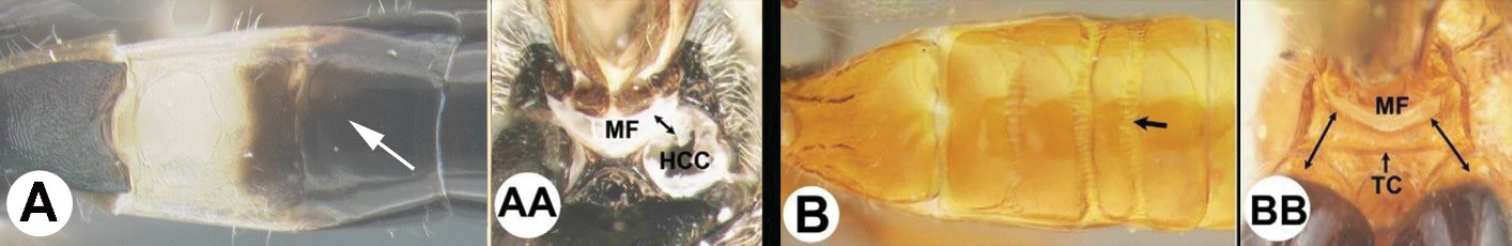
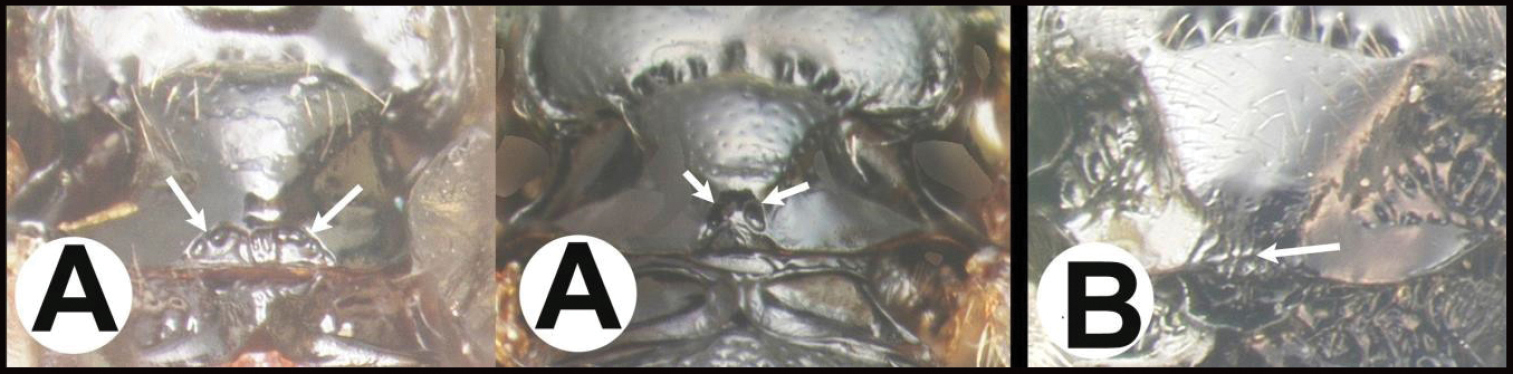
| 3(2) | A Median area of first tergum not raised above lateral portions and granulate or striogranulate. AA Hind coxal cavities (HCC) open to metasomal foramen or narrowly closed and positioned partly above ventral margin of metasomal foramen (MF) | Plesiocoelus |
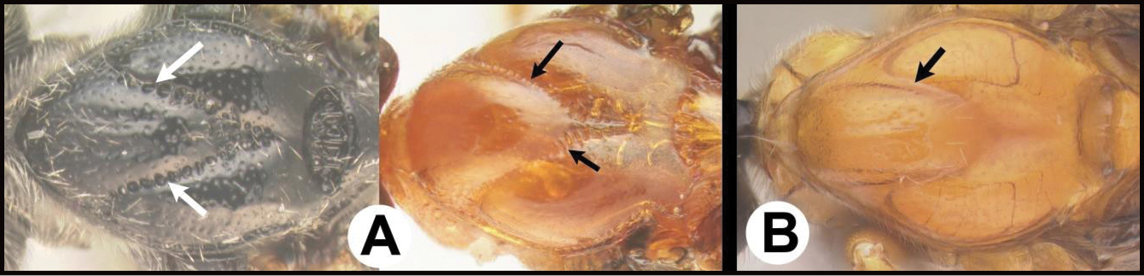
| – | B Median area of first tergum raised above lateral portions, sculpture variable but often smooth or smoothly striate. BB Hind coxal cavities closed and positioned completely below the metasomal foramen; ventral margin of metasomal foramen with a strong, relatively straight transverse carina (TC) | Aerophilus (in part) |
 |
||
| 4(1) | A Fore tarsal claws bifid | 5 |
| – | B Fore tarsal claws simple, with distinct basal lobe | 9 |
| – | C Fore tarsal claws simple, lacking a distinct basal lobe | 31 |
 |
||
| 5(4) | A Forewing areolet quadrate, not or only slightly narrower anteriorly. AA Ovipositor as long as or longer than half the length of metasoma | 7 |
| – | B Forewing areolet triangular or if quadrate much narrower anteriorly. BB Ovipositor shorter than half the length of metasoma | 6 |
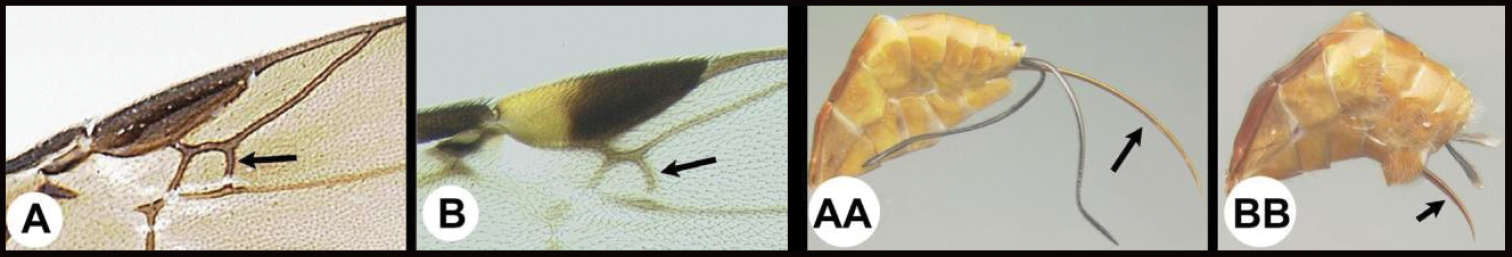 |
||
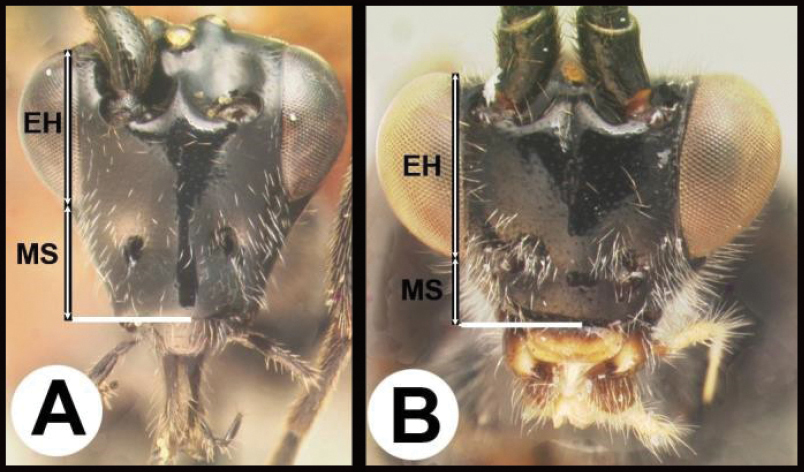
| 6(5) | A Gena expanded into a flange posteriorly; malar space (MS) >½ length of eye height (EH); Neotropical, rare | Hemichoma |
| – | B Gena not modified into a flange posteriorly; malar space (MS) <½ length of eye height (EH); widespread, common | Zelomorpha |
 |
||
| 7(5) | A Body predominantly orange/yellow. AA Frons bordered by a carina posteriorly; widespread, common | 8 |
| – | B Body predominantly black. BB Frons not bordered by a carina posteriorly; southern USA through the tropical Neotropics | Zacremnops |
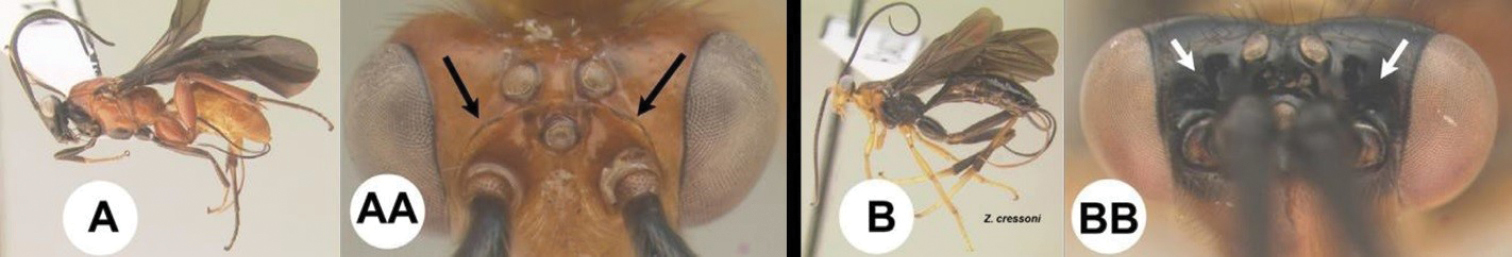 |
||
| 8(7) | A Propodeum and hind coxa with granulate sculpture; first metasomal tergum almost 3× wider at apex than at base; rare; Neotropical, rare | Labagathis |
| – | B Propodeum and hind coxa lacking granulate sculpture; first metasomal tergum not nearly 3× wider at apex than at base; common; widespread, relatively common | Cremnops |
 |
||
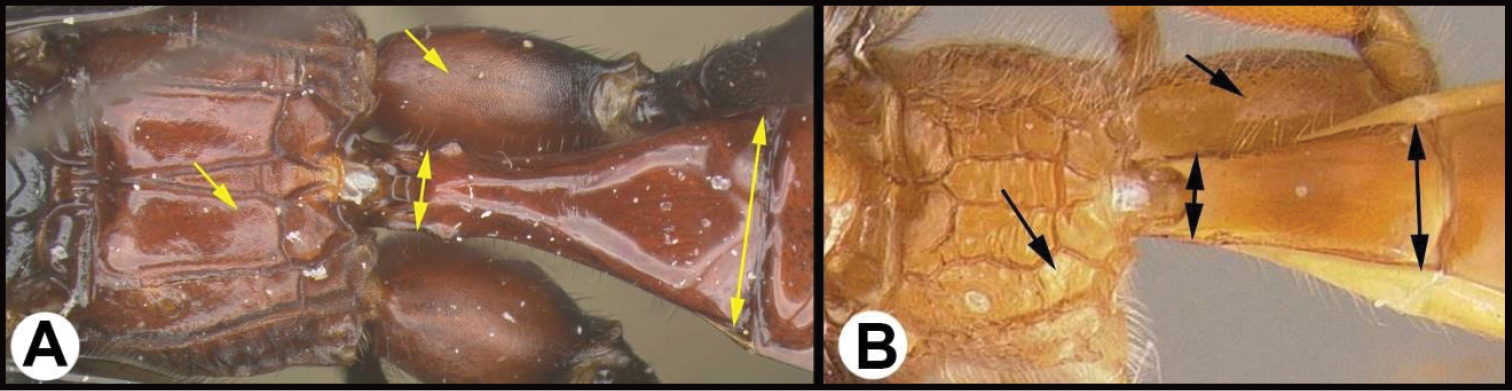
| 9(4) | A Notauli present, though sometimes weak | 10 |
| – | B Notauli completely absent | 25 |
 |
||
| 10(9) | A Ventral margin of clypeus projecting; width of temple longer than width of eye in lateral view; Nearctic, rare | Gelastagathis |
| – | B Ventral margin of clypeus not projecting; width of temple shorter than width of eye in lateral view; widespread, common | 11 |
 |
||
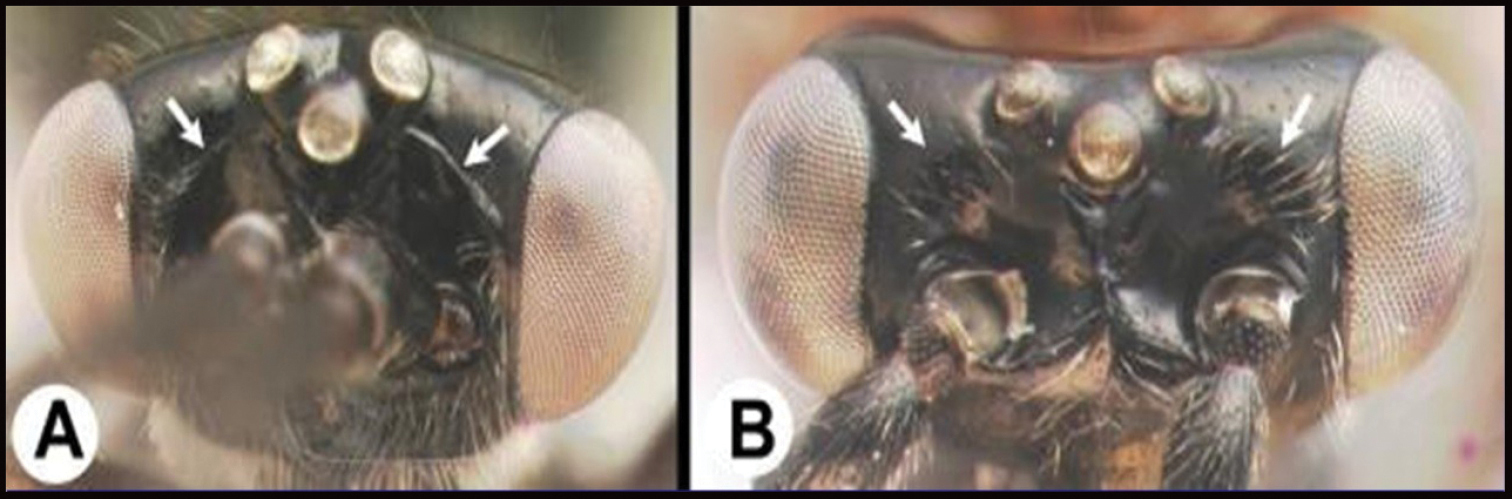
| 11(10) | A Frons bordered by carinae or grooves posteriorly | 12 |
| – | B Frons not bordered by carinae or grooves posteriorly | 14 |
 |
||
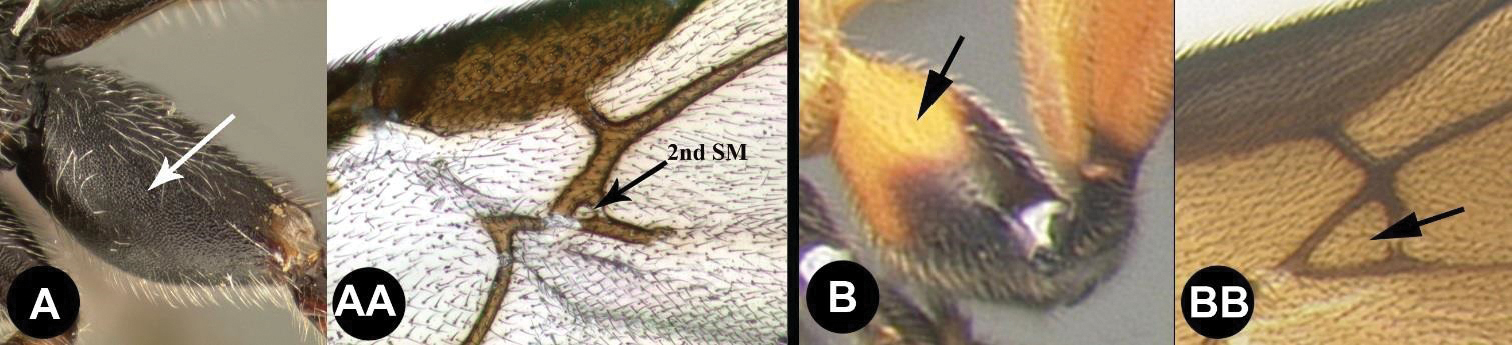
| 12(11) | A Hind coxa with granulate sculpture. AA Second submarginal cell minute or absent; Neotropical, rare | Trachagathis |
| – | B Hind coxa smooth, lacking granulate sculpture. BB Second submarginal cell of normal dimensions; widespread, relatively common | 13 |
 |
||
| 13(12) | A First metasomal tergum smooth with two widely spaced converging carinae forming a tear-shaped basal area; Neotropical, rare | Pharpa |
| – | B First metasomal tergum usually smooth and convex, or BB with a median longitudinal carina, or, BBB rarely with 2 carinae in which case the tergum has more extensive sculpture; widespread, common | Alabagrus |
 |
||
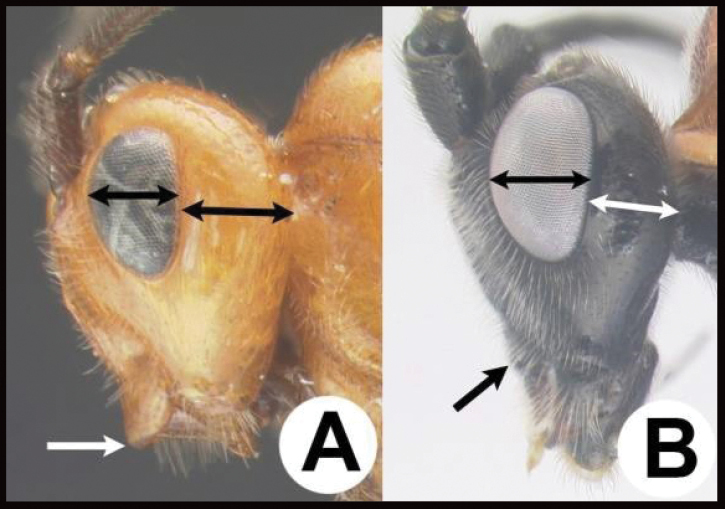
| 14(11) | A Malar space (MS) distinctly >½ eye height (EH). Head shape in frontal view elongate, at least as high (measure from ventral margin of clypeus) as wide | 15 |
| – | B Malar distance usually (95%) ≤½ eye height. Head shape in frontal view wide, wider than high (measure from ventral margin of clypeus) | 16 |
 |
||
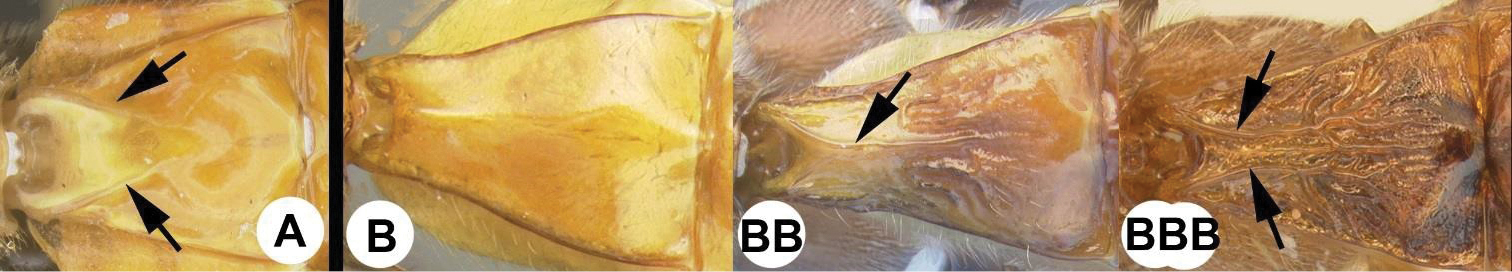
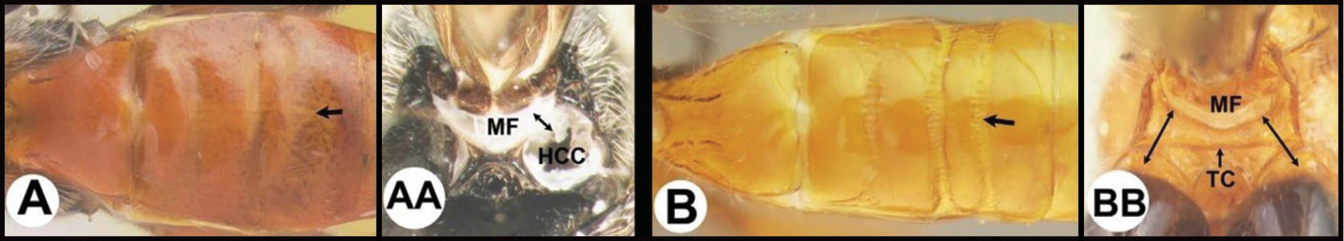
| 15(14) | A Third tergum completely smooth; pair of carinae on first tergum not prominent. AA Hind coxal cavities (HCC) open to metasomal foramen or narrowly closed and positioned partly above ventral margin of metasomal foramen (MF); common in the Nearctic, very rare in the Neotropics | Agathis (in part) |
| – | B Third tergum usually (95%) partly or completely sculptured, often sculpture confined to narrow line along transverse depression; pair of carinae on first tergum prominent. BB Hind coxal cavities closed and positioned completely below the metasomal foramen; ventral margin of metasomal foramen with a strong, relatively straight transverse carina (TC); widespread, common | Aerophilus (in part) |
 |
||
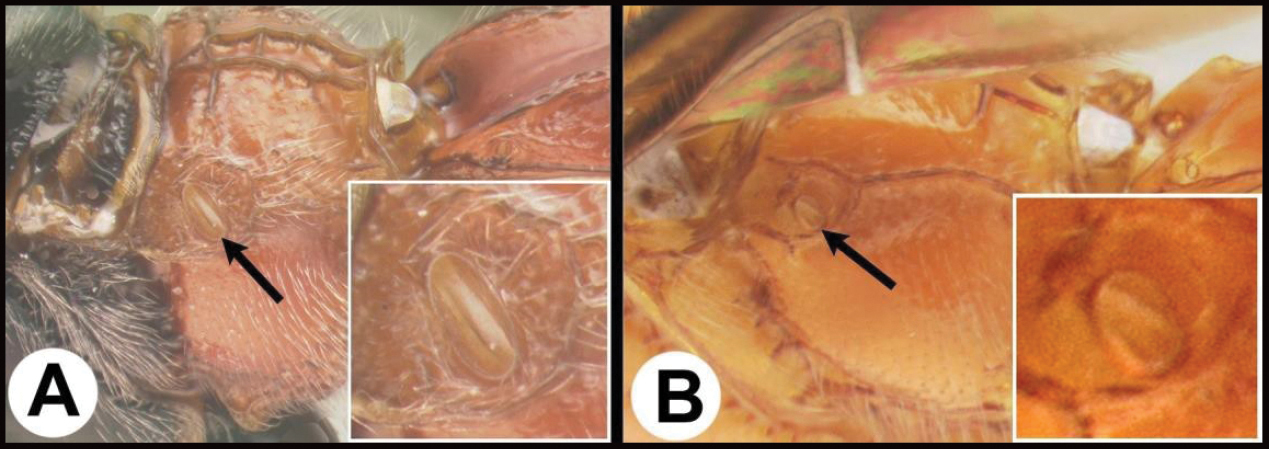
| 16(14) | A Propodeal spiracle elongate, > 2× longer than wide; widespread, common | Pneumagathis |
| – | B Propodeal spiracle circular or oval, < 2× longer than wide | 17 |
 |
||
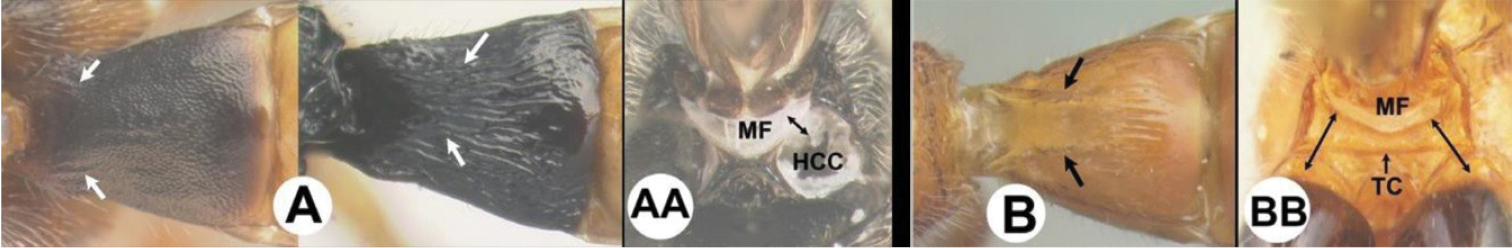
| 17(16) | A Pair of carinae on first tergum NOT prominent. AA Hind coxal cavities (HCC) open to metasomal foramen or narrowly closed and positioned partly above ventral margin of metasomal foramen (MF) | 18 |
| – | B Pair of carinae on first tergum prominent; BB Hind coxal cavities closed and positioned below the metasomal foramen; ventral margin of metasomal foramen with a strong, relatively straight transverse carina (TC); widespread, common | Aerophilus (in part) |
 |
||
| 18(15) | A First tergum completely smooth, or rarely with some punctures posterolaterally | 20 |
| – | B First tergum with sculpture | 19 |
 |
||
| 19(18) | A Cub vein of hind wing long and partly tubular, apical margin of subbasal (SB) cell angled; widespread, common | Therophilus (in part) |
| – | B Cub vein of hind wing weak or absent and never tubular; apical margin of subbasal cell (SB) straight. Nearctic and northern Neotropics, i.e., Mexico and Central America, rare | Agathirsia (in part) |
 |
||
| 20(18) | A Notauli pitted or crenulate | 21 |
| – | B Notauli smooth | 24 |
 |
||
| 21(17) | A Third tergum usually entirely smooth or weakly and partly coriarious (leather-like), if with different sculpture (especially in transverse depressions) then pair of longitudinal carinae on first metasomal tergum weaker than in B or absent. AA Hind coxal cavities (HCC) open to metasomal foramen or narrowly closed and positioned partly above ventral margin of metasomal foramen (MF) | 22 |
| – | B Third tergum usually partly or completely sculptured, often sculpture confined to narrow line along transverse depression. B Pair of longitudinal carinae on first metasomal tergum present and extending past spiracles. BB Hind coxal cavities closed and positioned entirely below the metasomal foramen (MF); ventral margin of metasomal foramen with a strong, relatively straight transverse carina (TC); widespread, common | Aerophilus (in part) |
 |
||
| 22(21) | A First tergum partly or completely granulate; widespread, common | Neothlipsis |
| – | B First tergum otherwise sculptured, usually striate or rugosostriate | 23 |
 |
||
| 23(22) | A Posterior apex of scutellum with a distinct depression in the form of a semicircle or two distinct pits; widespread, common | Therophilus (in part) |
| – | B Posterior apex of scutellum lacking depression, smooth to rugose; common in the Nearctic, very rare in the Neotropics | Agathis (in part) |
 |
||
| 24(19) | A Propleuron with a distinct protuberance; gena expanded into an acute angle posteroventrally; Neotropical, rare | Zamicrodus |
| – | B Propleuron flat or weakly convex, lacking a distinct protuberance; genae not expanded and rounded posteroventrally; Neotropical, rare | Aphelagathis |
 |
||
| 25(9) | A Second submarginal cell of forewing quadrate. AA Cub vein of hind wing present and often tubular; subbasal cell angled distally at junction of Cub | 26 |
| – | B Not combining the above character states. B. Second submarginal cell usually triangular. BB Cub vein of hind wing usually absent or not tubular and subbasal cell not angled distally; widespread, common | 27 |
 |
||
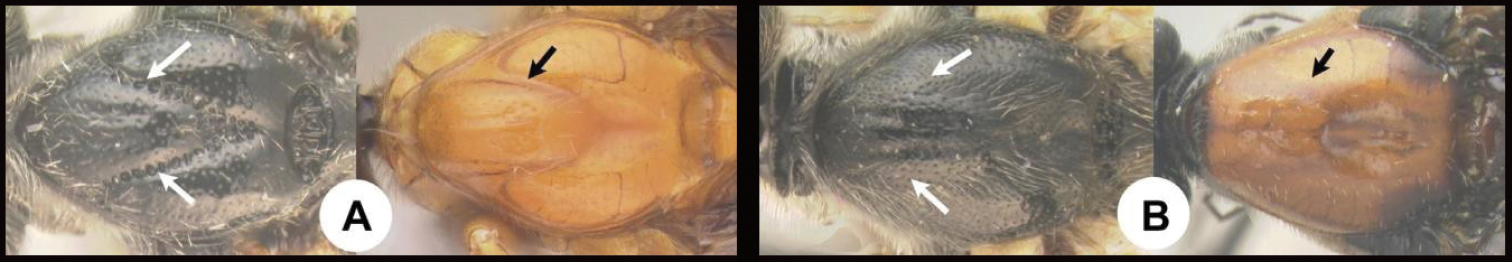
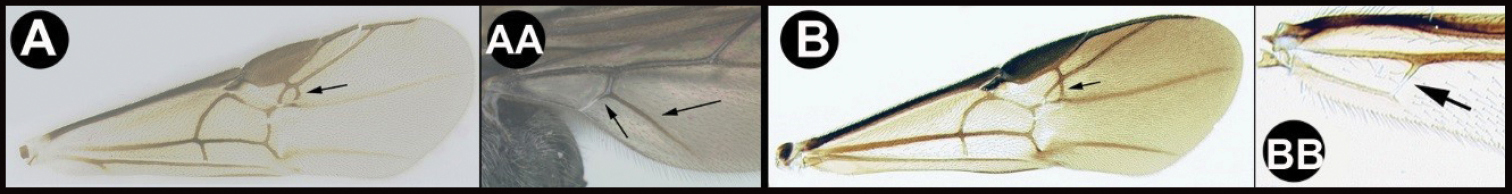
| 26(25) | A Face distinctly punctate AA. Body and antenna black, all femora pale (yellow to orange), hind tibia pale (yellow to orange with a melanic apex and a melanic ring or lateral spot subbasally. AAA Holarctic and Oriental (in the Americas from northern Canada and rarely as far south as northern Mexico) | Earinus |
| – | B Face smooth with very tiny punctation BB Often brightly colored and otherwise not as above. BBB Neotropical: Chile, and southern Argentina, and rarely in high altitudes of the Andes far north as Colombia and Ecuador | Chilearinus |
 |
||
| 27(25) | A Third tergum completely smooth. AA Hind coxal cavities (HCC) open to metasomal foramen (MF), or narrowly closed such that the ventral part of the metasomal foramen is below the dorsal margin of the hind coxal cavities | 28 |
| – | B Third tergum usually (95% of specimens encountered) partly or completely sculptured, often sculpture confined to narrow lines along transverse depressions. BB Hind coxal cavities closed and positioned completely below the metasomal foramen; ventral margin of metasomal foramen with a strong, relatively straight transverse carina; widespread, common | Aerophilus (in part) |
 |
||
| 28(27) | A Spurious vein, RS2b, well developed. AA Ovipositor barely exerted, much shorter than metasoma; Neotropical, very rare | Marjoriella |
| – | B Spurious vein, RS2b, lacking. BB Ovipositor at least as long as metasoma | 29 |
 |
||
| 29(28) | A Second submarginal cell smaller than its dorsal stem; apical abscissa of RS curving towards fore margin of wing; Neotropical, very rare | Smithagathis |
| – | B Second submarginal cell larger than its dorsal stem; apical abscissa of RS straight | 30 |
 |
||
| 30(29) | A Posterolateral corner of gena sharp; propleuron with a protuberance; Neotropical, rare | Amputoearinus |
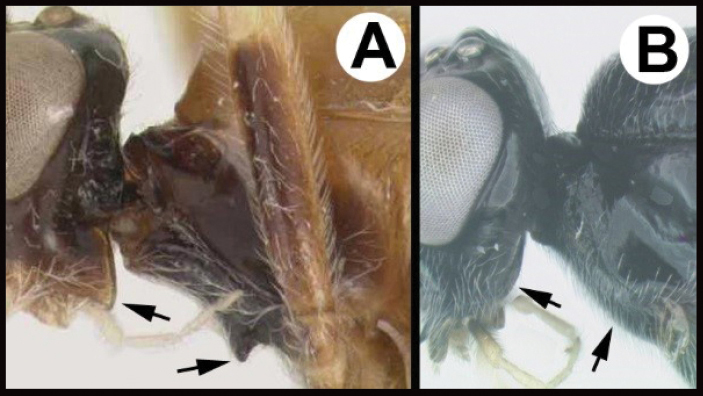
| – | B Posterolateral corner of gena rounded; propleuron evenly convex, lacking a protuberance; widespread, common | Lytopylus |
 |
||
| 31(4) | A Notauli absent, mesoscutum completely smooth; Neotropical, rare | Sesioctonus |
| – | B Notauli present though sometimes only indicated anteriorly or posteriorly; widespread and common in the Nearctic, rare in the Neotropics | 32 |
 |
||
| 32(31) | A First tergum smooth, lacking microsculpture and pair of longitudinal carinae, at most with punctures laterally | 33 |
| – | B First tergum with microsculpture, usually in the form of longitudinal striae or rugae; widespread and common in the Nearctic, extremely rare in the Neotropics | Agathis (in part) |
 |
||
| 33(32) | A Ovipositor barely exerted, shorter than half the length of metasoma; Nearctic and Central America, rare | Crassomicrodus |
| – | B Ovipositor at least as long as half the metasoma, often much longer; Nearctic and Central America, rare | Agathirsia (in part) |
 |
References
- Berta DC (2000) Contribución sobre las especies neotropicales del género Earinus Wesmael (Himenóptera: Braconidae, Agathidinae). Boletin de la Asociacion Espanola de Entomologia 24: 229–241. https://notablesdelaciencia.conicet.gov.ar/handle/11336/78424
- Braet Y (2002) Contribution to the knowledge of Agathidinae (Hymenoptera Braconidae) from French Guiana with description of two new species of Earinus Wesmael, 1837. Belgian Journal of Entomology 4: 41–51.
- Ivanova NV, Grainger CM (2007) CCDB protocols, COI amplification. http://ccdb.ca/site/wp-content/uploads/2016/09/CCDB_Amplification.pdf [accessed 1 July 2019]
- Ivanova NV, Dewaard JR, Hebert PDN (2006) An inexpensive, automation‐friendly protocol for recovering high‐quality DNA. Molecular Ecology Resources 6(4): 998–1002. https://doi.org/10.1111/j.1471-8286.2006.01428.x
- Muesebeck CFW (1927) A revision of the parasitic wasps of the subfamily Braconinae occurring in America north of Mexico. Proceedings of the United States National Museum 69(2642): 1–73. https://doi.org/10.5479/si.00963801.69-2642.1
- Say T (1835) Descriptions of new North American Hymenoptera, and observations on some already described. Boston Journal of Natural History 1(3): 210–305. https://www.biodiversitylibrary.org/page/32413933
- Sharkey MJ (1997) Subfamily Agathidinae. In: Wharton RA, Marsh PM, Sharkey MJ (Eds) Manual of the New World genera of Braconidae (Hymenoptera). Special Publication of the International Society of Hymenopterists, Vol. 1., 69–84.
- Sharkey MJ, Meierotto S, Chapman EG, Janzen DJ, Hallwachs W, Dapkey T, Solis MA (2018) Alabagrus Enderlein (Hymenoptera, Braconidae, Agathidinae) species of Costa Rica, with an emphasis on specimens reared from caterpillars in Area de Conservación Guanacaste. Contributions in Science 526: 31–180. https://doi.org/10.5962/p.320146
- Sharkey MJ, Janzen DH, Hallwachs W, Chapman EG, Smith MA, Dapkey T, Brown A, Ratnasingham S, Naik S, Manjunath R, Perez K, Milton M, Hebert PDN, Shaw SR, Kittel RN, Solis MA, Metz MA, Goldstein PZ, Brown JW, Quicke DLJ, van Achterberg C, Brown BV, Burns JM (2021) Minimalist revision and description of 403 new species in 11 subfamilies of Costa Rican braconid parasitoid wasps, including host records for 219 species. ZooKeys 1013: 1–665. https://doi.org/10.3897/zookeys.1013.55600
- Spinola M (1851) Icneumónidos. In: Gay C (Ed.) Historia física y politica de Chile. Zoologia 6: 471–550.
- Walley GS (1935) Five new species of Braconidae with host records of additional species. Canadian Entomologist 67(3): 55–61. https://doi.org/10.4039/Ent6755-3
- Yu DSK, van Achterberg C, Horstmann K (2016) Taxapad, Ichneumonoidea. Vancouver. http://www.taxapad.com
- Zamani A, Vahtera V, Sääksjärvi IE, Scherz MD (2020) The omission of critical data in the pursuit of “revolutionary” methods to accelerate the description of species. Systematic Entomology 46(1): 1–4. https://doi.org/10.1111/syen.12444