Research Article |
|
Corresponding author: Jiang-Li Tan ( tanjl@nwu.edu.cn ) Academic editor: Michael S. Engel
© 2021 Jiang-Li Tan, Cornelis van Achterberg, Jia-Xuan Wu, Hang Wang, Qi-Jing Zhang.
This is an open access article distributed under the terms of the Creative Commons Attribution License (CC BY 4.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Citation:
Tan JL, Achterberg C van, Wu JX, Wang H, Zhang QJ (2021) An illustrated key to the species of Gasteruption Latreille (Hymenoptera, Gasteruptiidae) from Palaearctic China, with description of four new species. ZooKeys 1038: 1-103. https://doi.org/10.3897/zookeys.1038.64978
|
Abstract
Four new species of the genus Gasteruption Latreille, 1797 (Hymenoptera: Evanioidea: Gasteruptiidae: Gasteruptiinae) are described from China. Three are from Shaanxi (NW China; G. granulatum Tan & van Achterberg, sp. nov., G. pedion Tan & van Achterberg, sp. nov., and G. reductum Tan & van Achterberg, sp. nov.) and one from S China and Ningxia (G. kexinae Tan & van Achterberg, sp. nov.). Eleven species are newly recorded for Shaanxi (G. abeillei Kieffer, 1912, G. amoyense Pasteels, 1958, G. bimaculatum Pasteels, 1958, G. corniculigerum Enderlein, 1913, G. latitibia Zhao, van Achterberg & Xu, 2012, G. minutum (Tournier, 1877), G. nigritarse (Thomson, 1883), G. parvicollarium Enderlein, 1913, G. sinarum Kieffer, 1911, G. subtile (Thomson, 1883) and G. brevicuspis Kieffer, 1911). The newly-recorded species and the new species are keyed and illustrated. Two new synonyms are proposed: G. rufescenticorne Enderlein, 1913, with G. japonicum Cameron, 1888, syn. nov. and G. oriplanum Kieffer, 1911, with G. minutum (Tournier, 1877), syn. nov.
Keywords
Inner Mongolia, new record, Ningxia, Shaanxi, Xinjiang
Introduction
Gasteruptiidae are very slender apocritan hymenopterans with elongated “neck” (propleuron), swollen hind tibiae and compressed petiolate metasoma attached very high on the propodeum. Up to now, 511 species are recognised as valid in two subfamilies, Gasteruptiinae (with four genera) (
The adults frequently feed on flowers with easily accessible nectar (especially families Apiaceae, Asteraceae and Euphorbiaceae), but likely at least some Gasteruption species feed on both nectar and pollen (
All known gasteruptiids from the Palaearctic Region belong to the subfamily Gasteruptiinae and to the genus Gasteruption Latreille, 1797. Up to 2018, 33 species were known from China, of which seven were found in the NW Chinese Province Shaanxi, which is 21% of the total for China. However,
Material and methods
The specimens were collected by sweep nets or in Malaise traps. The material was directly killed and stored in 70% ethanol and subsequently prepared according to the AXA method (
Observations and descriptions were made with an Opto-Edu A230903 stereomicroscope and a fluorescent lamp. Photographic images were made with the Keyence VHX-5000 digital microscope and processed with Adobe Photoshop CS5, mostly to adjust the size and background. For the identification,
The antesternal carina (van Achterberg in
The following abbreviations are used for the depositories:
ZIL Zoological Institute, Lund;
Taxonomy
Gasteruption
Remarks
Gasteruption Latreille, 1797: 113;
Key to species of the genus Gasteruption Latreille from Palaearctic China
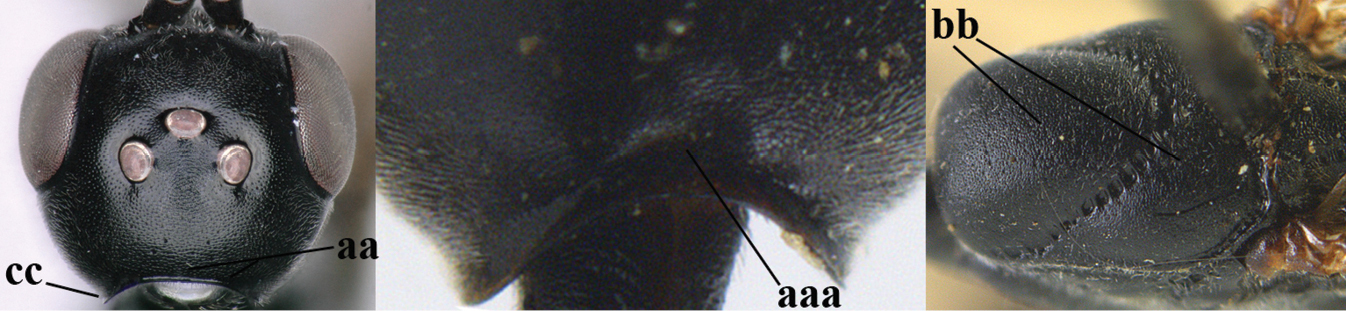
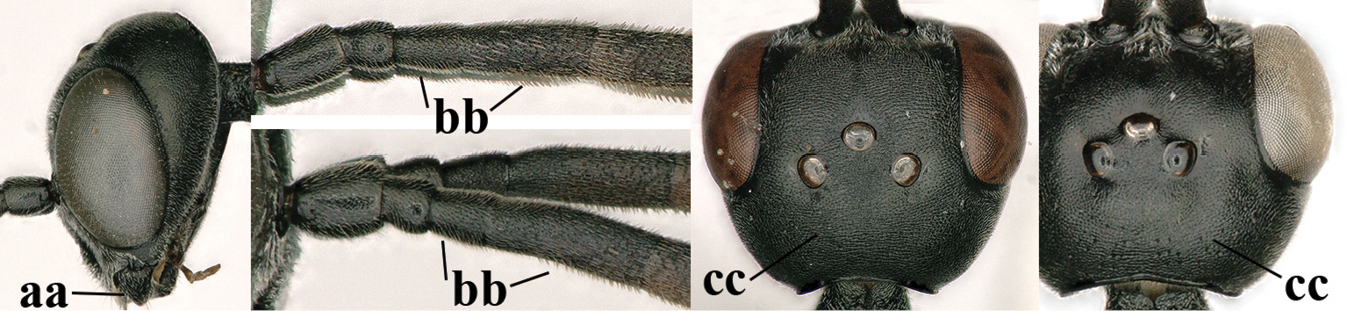
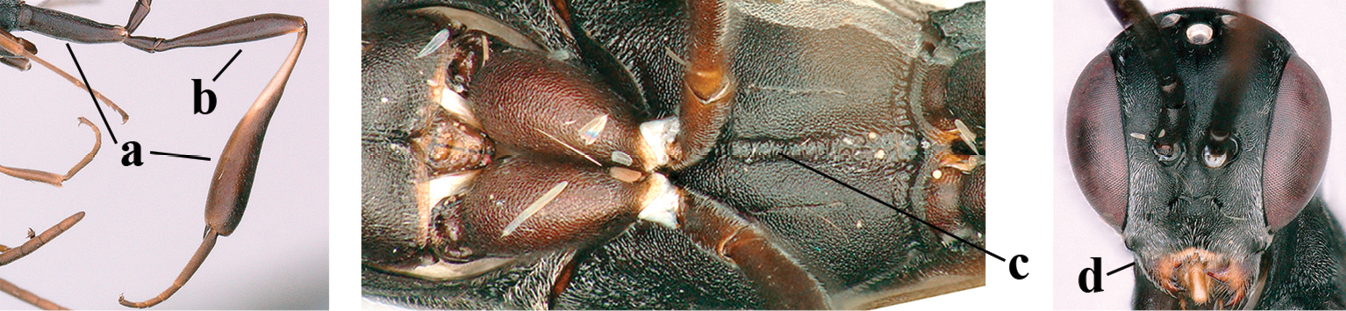
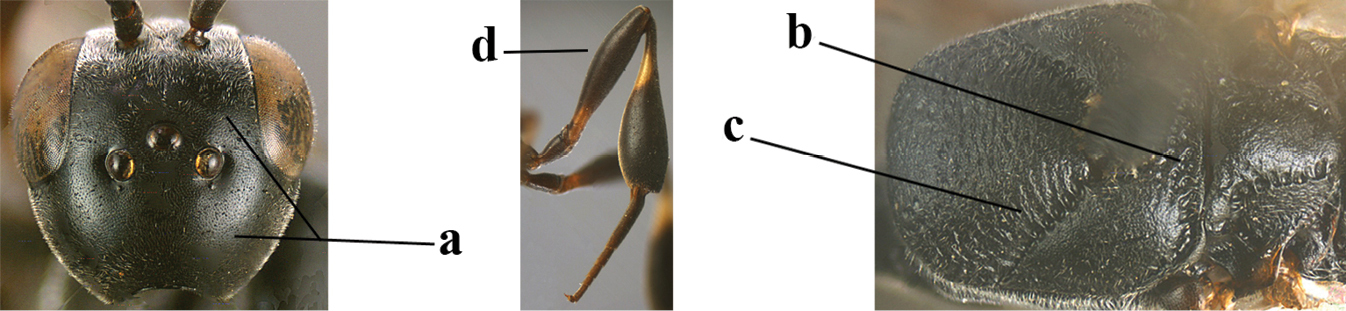
| 1 | Ovipositor present (a); antenna with 14 segments (b) (females) | 2 |
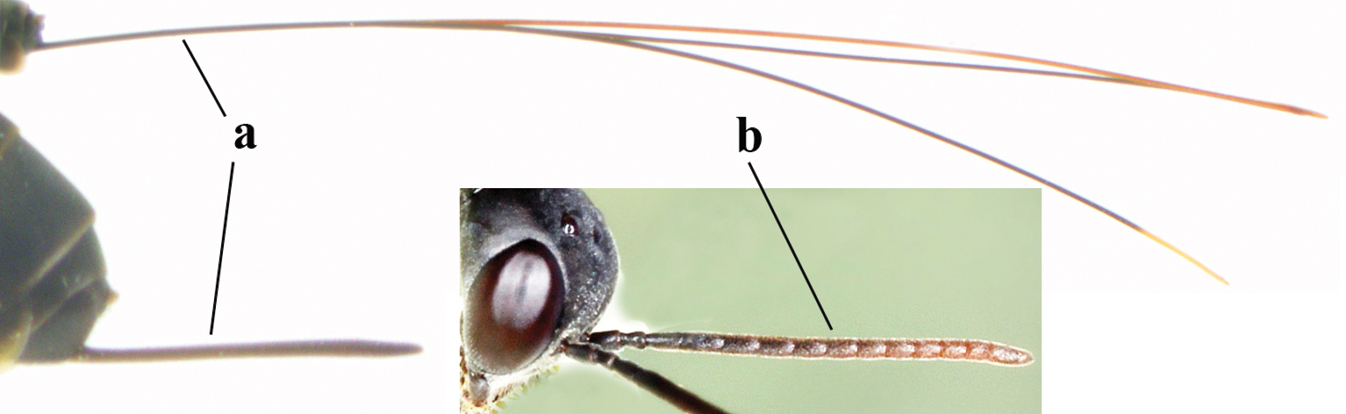
|
||
| – | Ovipositor absent (aa); antenna with 13 segments (bb) (males); [if males are unknown, the species is provisionally inserted] | 38 |

|
||
| 2 | Apex of ovipositor sheath blackish or dark brown; if narrowly pale apically, then white, ivory or brownish-yellow part at most 0.3× as long as hind basitarsus (a); (intermediate species are included in both alternatives) | 3 |

|
||
| – | Apex of ovipositor sheath distinctly white or ivory (but rarely pale brown) and pale part 0.3–8.0× as long as hind basitarsus (aa) | 23 |

|
||
| 3 | Ovipositor sheath 0.6–2.0× as long as hind tibia and 0.3–1.2× as long as hind tibia and tarsus combined (a); incision of hypopygium shallow V-shaped and up to apical 0.2 (b) or absent; occipital carina obsolescent to narrowly lamelliform medio-dorsally (c) | 4 |

|
||
| – | Ovipositor sheath 3.0–7.0× as long as hind tibia and 1.9–4.0× as long as hind tibia and tarsus combined (aa); incision of hypopygium often deep and slit-like up to apical 0.3–0.5 (bb); occipital carina obsolescent (c) or distinctly lamelliform (cc) medio-dorsally | 16 |

|
||
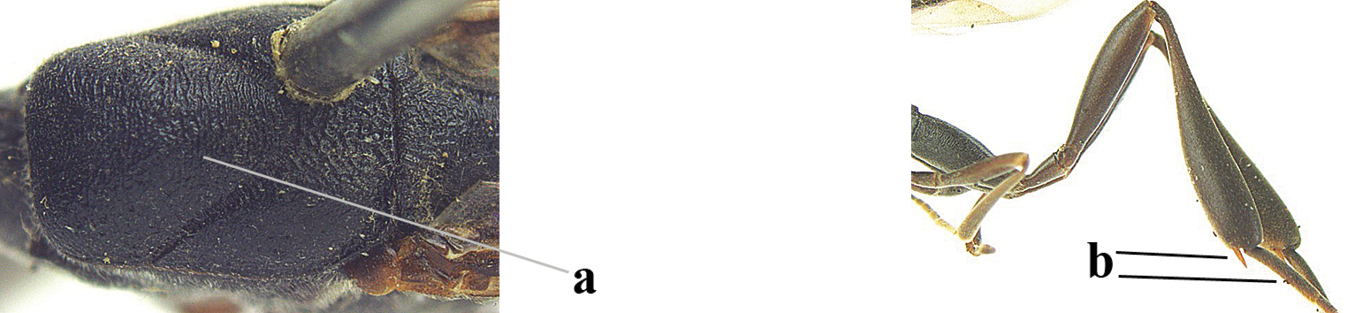
| 4 | Head in anterior view protruding below lower level of eyes, 0.5–0.6× length of second antennal segment and 0.4–0.6× basal width of mandible and mandibular condylus distinctly below lower level of eyes (a); in lateral view condylar incision of malar space remains far removed from eye (b); ovipositor sheath 0.4–0.9× as long as hind tibia (c) | G. minutum (Tournier, 1877) |

|
||
| – | Head in anterior view slightly protruding below lower level of eyes by less than half basal width of mandible and mandibular condylus near lower level of eyes (aa); in lateral view condylar incision of malar space close to eye (bb), rarely slightly wider; ovipositor sheath 0.7–2.7× as long as hind tibia (cc) | 5 |

|
||
| 5 | Clypeus with rather large shallow depression (a); mesoscutum densely reticulate-rugulose or -rugose (b); hind basitarsus stout (c); apical antennal segment 1.4–1.6× third antennal segment (d); [ovipositor sheath 0.6–1.5× as long as hind tibia] | G. hastator (Fabricius, 1804) |

|
||
| – | Clypeus with small depression or depression obsolescent (aa); mesoscutum mainly densely coriaceous or rugulose (bb); hind basitarsus more slender (cc), rarely similarly stout; apical antennal segment at most 1.2× as long as third antennal segment (dd) | 6 |

|
||
| 6 | Mesoscutum coarsely (often “crater”-like) punctate (a); head distinctly emarginate medio-posteriorly (b); head less protruding in lateral view (c) and narrower in anterior view (d) | 7 |

|
||
| – | Mesoscutum predominantly densely coriaceous, at most with some shallow punctures (aa); head truncate medio-posteriorly or nearly so (bb); head more protruding in lateral view (cc) and wider in anterior view (dd) | 8 |

|
||
| 7 | Hind tibia about as long as hind femur and trochanter combined or slightly longer (a); head somewhat longer in dorsal (b) and lateral (c) view; head directly narrowed behind eyes in dorsal view (d) | G. formosanum Enderlein, 1913 |

|
||
| – | Hind tibia 1.1–1.2× as long as hind femur and trochanter combined (aa); head somewhat shorter in dorsal (bb) and lateral (cc) view; head roundly narrowed behind eyes in dorsal view (dd) | G. sinicola (Kieffer, 1924) |

|
||
| 8 | Vertex strongly bulging above upper level of eyes (a); head comparatively long in dorsal view (b); ovipositor sheath 1.2–1.7× as long as hind tibia and apex of ovipositor wide and with distinct dorsal teeth (c); propleuron slightly less robust in lateral view (d); [hind tibia moderately slender; propleuron antero-dorsally and pronotum ventrally coriaceous; pronotum convex antero-ventrally] | G. parvicollarium Enderlein, 1913 |

|
||
| – | Vertex at most moderately bulging above upper level of eyes (aa); head shorter in dorsal view (bb); ovipositor sheath usually shorter; if 1.2–1.9× longer than hind tibia, then apex of ovipositor narrow and with minute dorsal teeth (cc); propleuron robust in lateral view (dd) | 9 |

|
||
| 9 | Hind tibia slender (a); pronotal sides antero-dorsally granulate (b); side of pronotum slender and with narrow and weakly crenulated grooves (c); ovipositor sheath 1.3–1.9× as long as hind tibia; [hind basitarsus elongate; hind tibia dark brown to yellowish-brown ventro-basally] | G. granulatum sp. nov. |

|
||
| – | Hind tibia distinctly inflated (aa); pronotal sides antero-dorsally coriaceous or rugulose (bb); side of pronotum robust and with wider and distinctly crenulated grooves (cc); ovipositor sheath 0.7–1.4× as long as hind tibia | 10 |

|
||
| 10 | Mandible black, dark brown or reddish-brown basally (a), rarely narrowly so and largely brownish-yellow; basal depression of mandible rather large and deep (b); tegula dark brown or brown (c); fifth (= pre-apical) sternite dark brown, blackish or narrowly pale medio-apically (d) | 11 |

|
||
| – | Mandible pale yellow basally (aa); basal depression of mandible often smaller and shallower (bb); tegula yellow (cc), brownish-yellow or brown (ccc); fifth sternite more or less yellowish medio-apically (dd) | 14 |

|
||
| 11 | Head in dorsal view directly narrowed posteriorly and longer (a); vertex moderately protruding above eye in lateral view (b); mesopleuron more elongated (c) | G. latitibia Zhao, van Achterberg & Xu, 2012 |

|
||
| – | Head in dorsal view rounded narrowed posteriorly and shorter (aa); vertex less protruding above eye in lateral view (bb); mesopleuron less elongated (cc) | 12 |

|
||
| 12 | Ovipositor sheath with curved bristles (“velcro”-type; a); lateral buccal area shallowly depressed (b); hypostomal bridge often longer (behind buccal area at underside of head; c); [pronotum antero-laterally evenly finely sculptured; POL 1.7–2.3× width of anterior ocellus] | G. nigritarse (Thomson, 1883) |

|
||
| – | Ovipositor sheath with normal straight setae, either largely bristly and erect (aa) or mostly adpressed and finely setose (aaa); lateral buccal area distinctly depressed (bb); hypostomal bridge usually shorter (bb); [sculpture of pronotum antero-laterally variable] | 13 |

|
||
| 13 | Ovipositor sheath medially only with short adpressed pale setae at 50× (lacking upward directed dark brown and short bristly setae) and subapically slightly bristly setose (a); ovipositor sheath not or slightly widened subapically (b), 0.9–1.4× as long as hind tibia (only to measure if fully exserted); second and third antennal segments usually more robust (c); occipital carina wider latero-dorsally (d); [medially sculpture of mesoscutum at 60× variable, often dissimilar to very fine sculpture of vertex and more or less rugulose] | G. assectator (Linnaeus, 1758) |

|
||
| – | Ovipositor sheath medially with erect setae or bristles at 50× and subapically distinctly bristly setose (aa); ovipositor sheath widened subapically (bb), 0.6–1.1× as long as hind tibia; second and third antennal segments usually more slender (cc); occipital carina narrower latero-dorsally (dd); [medially sculpture of mesoscutum at 60× variable, frequently similar to fine sculpture of vertex or somewhat coarser | G. abeillei Kieffer, 1912 |

|
||
| 14 | Hind basitarsus rather stout and at least partly ivory dorsally (a); mesoscutum somewhat coarser sculptured (b); fifth metasomal sternite widely pale yellowish posteriorly (c); [hind tibia dark ventrally and hind femur black] | G. flavimarginatum van Achterberg, 2014 |

|
||
| – | Hind basitarsus slender and entirely dark brown or brown dorsally (aa); mesoscutum finely sculptured (bb); fifth sternite narrowly pale yellowish posteriorly or mainly dark brown (cc) | 15 |
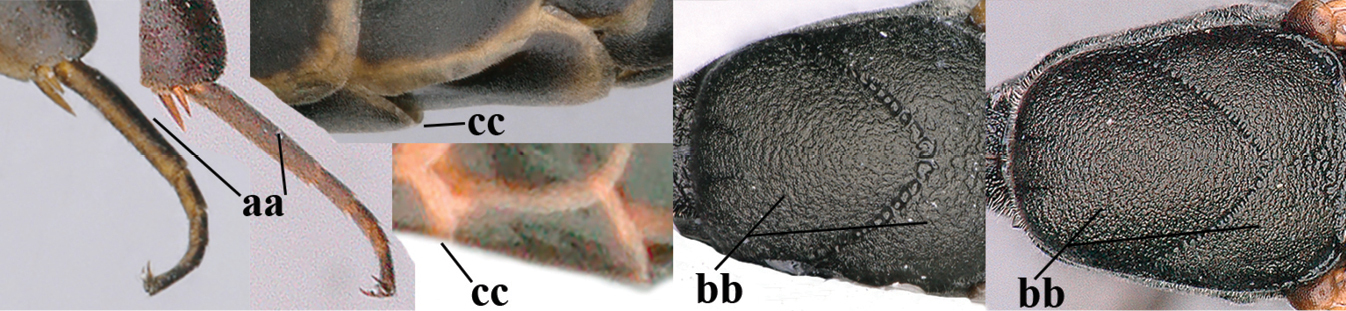
|
||
| 15 | Posterior ocellus situated near upper level of vertex (a); head in dorsal view directly narrowed posteriorly (b); ovipositor sheath 0.5–0.7× as long as hind tibia (c); [hind femur often partly dark reddish-brown; malar space 0.2–0.3× basal width of mandible] | G. bicoloratum Tan & van Achterberg, 2016 |

|
||
| – | Posterior ocellus situated distinctly below upper level of vertex (aa); head in dorsal view gradually narrowed posteriorly (bb); ovipositor sheath 0.8–1.1× as long as hind tibia (cc) | G. brevicuspis Kieffer, 1911 |

|
||
| 16 | Vertex of ♀ with reversed V-shaped emargination medio-posteriorly (a; emargination hardly developed in ♂), flat (b) and smooth, shiny and long dorsally (c); mesoscutum of ♀ finely transversely rugose to nearly smooth (d); [dorsal apical teeth of ovipositor distinct] | G. bimaculatum Pasteels, 1958 |

|
||
| – | Vertex of ♀ truncate medio-posteriorly (aa) or reversed U-shaped emarginate (aaa), shorter and moderately convex (bb); if vertex more or less emarginate and/or flat, then vertex finely sculptured, with satin sheen and shorter (cc); mesoscutum of ♀ punctate, punctate-rugose or transversely wrinkled (dd) | 17 |

|
||
| 17 | Head rather elongate and below eyes slightly enlarged, minimum length of malar space 0.3–0.4× second antennal segment (a); head distinctly reversed U-shaped emarginate medio-posteriorly (b); mandible and malar space brown (c); hind tarsus brownish apically, paler than basally (d); [apex of ovipositor sheath ivory; first metasomal tergite orange or yellowish-brown] | G. dimidiatum Semenov, 1892 |

|
||
| – | Head less elongate and below eyes not enlarged, minimum length of malar space 0.1–0.2× second antennal segment (aa); head shallowly emarginate medio-posteriorly (bb); if intermediate (bbb), then mandible brownish-yellow and contrasting with colour of malar space (cc); apically hind tarsus as dark brown as basally (dd); [apex of ovipositor sheath dark brown or black] | 18 |

|
||
| 18 | Mesosoma sparsely setose laterally (a); pale apical part of ovipositor sheath 0.3–1.0× as long as hind basitarsus, apex ivory, brownish-yellow or brown (b); hind femur dark brown or black (c) | 19 |

|
||
| – | Mesosoma densely setose laterally (aa); apex of ovipositor sheath blackish or mainly dark brown (bb); if apex pale, then at most 0.3× as long as hind basitarsus (bbb); hind femur orange or reddish-brown (cc), but black in G. shengi | 20 |

|
||
| 19 | Middle lobe of mesoscutum coarsely punctate (a); vertex strongly convex (b); mandible largely blackish or dark brown (c); mesosoma less elongated (d) | G. sinarum Kieffer, 1911 |

|
||
| – | Middle lobe of mesoscutum without punctures and finely transversely wrinkled (aa); vertex moderately convex (bb); mandible yellowish (cc); mesosoma distinctly elongated (dd) | G. pannuceum Tan & van Achterberg, 2016 |

|
||
| 20 | Hind tibia more slender (a); first metasomal tergite dark brown (b); middle lobe of mesoscutum remotely punctate (c); hind femur black or blackish-brown (d); mesoscutum and head black (e) | G. shengi Tan & van Achterberg, 2016 |

|
||
| – | Hind tibia more inflated (aa), but intermediate in G. argentifrons and G. coloratum (21bb); first tergite reddish or orange (bb); middle lobe of mesoscutum densely punctate or punctate-rugose (cc); hind femur orange brown to dark brown (dd), but partly or entirely black in G. coloratum; mesoscutum often paler than head (ee) or both reddish | 20 |

|
||
| 21 | Ovipositor sheath 3.1–4.4× as long as hind tibia (a); hind tibia more inflated (b); vertex longer setose (c); mesosoma largely or entirely black laterally (d); [hind femur and tibia (except basally) similarly coloured, orange brown or dark brown] | G. dilutum Semenov, 1892 |

|
||
| – | Ovipositor sheath approx. 7.0× as long as hind tibia (aa); hind tibia less inflated (bb); vertex shorter setose (cc); mesosoma often dark reddish or orange brown laterally (dd) | 22 |

|
||
| 22 | Basal half of hind coxa mainly transversely rugose or punctate dorsally (a); apex of ovipositor sheath largely dark brown or brown (b); pronotum longer setose (c); [hind coxa orange brown or blackish] | G. coloratum Zhao, van Achterberg & Xu, 2012 |

|
||
| – | Basal half of hind coxa superficially coriaceous dorsally (aa); apex of ovipositor sheath ivory or brownish-yellow (bb); pronotum shorter setose (cc) | G. argentifrons Semenov-T.-S. & Kostylev, 1928 |

|
||
| 23 | Head with medial depression in front of occipital carina and with pair of lateral depressions (a); if shallow, then head in dorsal view nearly parallel-sided behind eyes (b); occipital carina wide lamelliform medio-dorsally (c) | 24 |

|
||
| – | Head flat or evenly convex in front of occipital carina (aa); if with a shallow depression in front of occipital carina (aaa), then head directly narrowed behind eyes (bb) and occipital carina at most moderately lamelliform medio-dorsally (cc) | 26 |

|
||
| 24 | Propleuron moderately robust and 0.8–1.0× as long as mesoscutum in front of tegula (a); vertex medially weakly convex in lateral view (b); head shorter in dorsal view (c); mandible often pale in anterior view medially (d); [white or ivory part of ovipositor sheath 1.7–3.3× longer than hind basitarsus] | G. oshimense Watanabe, 1934 |

|
||
| – | Propleuron slender and 1.0–1.2× as long as mesoscutum in front of tegula (aa); vertex medially nearly flat in lateral view (bb); head comparatively long in dorsal view (cc); mandible often darker in anterior view (dd) | 25 |

|
||
| 25 | Head in dorsal view distinctly narrowed behind eyes (a); middle lobe of mesoscutum with fine transverse elements anteriorly (b; sometimes coarsely sculptured); propleuron in ventral view less slender anteriorly (c); white or ivory apical part of ovipositor sheath 2.4–3.8× longer than hind basitarsus (d) | G. corniculigerum Enderlein, 1913 |

|
||
| – | Head in dorsal view nearly parallel-sided behind eyes (aa); middle lobe of mesoscutum only coriaceous between punctures (bb); propleuron in ventral view more slender anteriorly (cc); white or ivory apical part of ovipositor sheath 1.0–2.1× longer than hind basitarsus (dd) | G. kexinae sp. nov. |

|
||
| 26 | Ovipositor sheath comparatively wide and about 0.9× as long as hind tibia, 0.3× as long as metasoma and 0.2× as long as body (a); middle lobe of mesoscutum rather protuberant in lateral view (b); pronotal tooth slender and acute | G. assectoides Zhao, van Achterberg & Xu, 2012 |
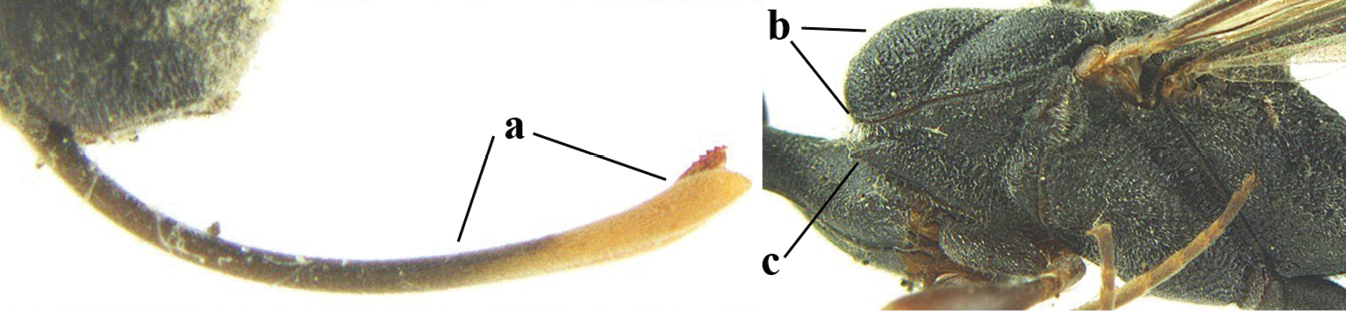
|
||
| – | Ovipositor sheath comparatively narrow and 1.1–9.0× as long as hind tibia, 0.6–2.8× as long as metasoma and 0.4–1.4× as long as body (aa); middle lobe of mesoscutum less protuberant in lateral view (bb); if convex (bbb), then pronotal tooth wider and rather blunt (cc) | 27 |

|
||
| 27 | Ovipositor about 0.4× as long as body and 0.6× as long as metasoma (a); hind coxa very slender (b); ovipositor widened apico-ventrally and more or less angularly up-curved apically in dead specimens (c) | G. angulatum Zhao, van Achterberg & Xu, 2012 |
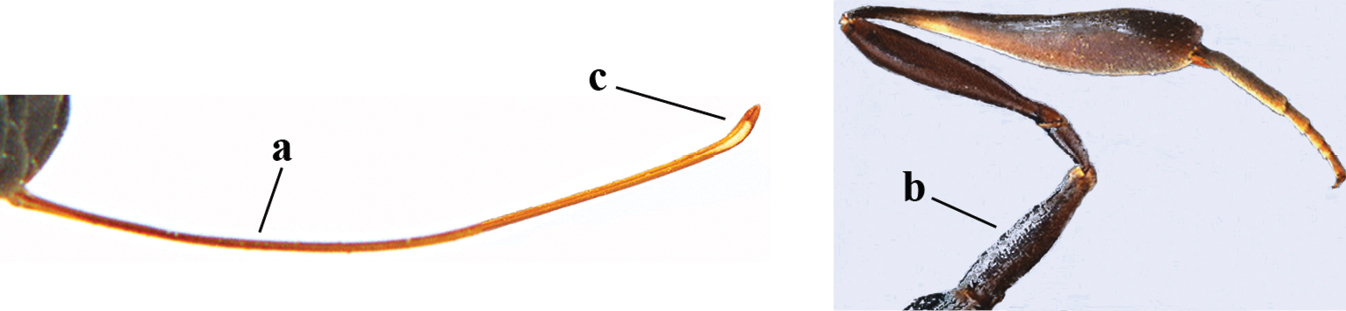
|
||
| – | Ovipositor 0.8–1.4× as long as body and 1.2–1.9× as long as metasoma (aa); hind coxa slightly less slender (bb); ovipositor narrow apico-ventrally and nearly straight or gradually up-curved apically (cc) | 28 |

|
||
| 28 | Hind femur and tibia widened (a); hind basitarsus robust (b); head slightly narrowed in dorsal view (c); head slender in anterior view (d) and face narrower than clypeus (e); hind basitarsus entirely dark brown (f); [ovipositor sheath about 1.4× as long as body and 8.5× as long as hind tibia] | G. huangshii Tan & van Achterberg, 2016 |

|
||
| – | Hind femur and tibia slender (aa); hind basitarsus more slender (bb); head in dorsal view distinctly narrowed (cc); head in anterior view subglobular (dd); if slender (ddd), then face as wide as clypeus (ee); hind basitarsus often partly ivory (ff) | 29 |

|
||
| 29 | Pale apical part of ovipositor sheath 0.2–1.0× as long as hind basitarsus (a); occipital carina non-lamelliform medio-dorsally (b) and mesoscutum distinctly punctate or rugose (c); if narrow lamelliform (bb), then hypopygium yellowish-brown or yellow; [ovipositor sheath 1.0–1.3× as long as body] | 30 |

|
||
| – | Pale apical part of ovipositor sheath 1.1–3.5× as long as hind basitarsus (aa); occipital carina narrow lamelliform medio-dorsally (bb) and hypopygium dark brown or black; if non-lamelliform (bbb), then mesoscutum very finely coriaceous and, at most, punctulate (cc) or transversely rugulose | 32 |

|
||
| 30 | Hypopygium dark brown or black (a); mesoscutum between coarse punctures with satin sheen and densely punctulate (b); head longer in dorsal view (c); face with short setosity laterally (d); [mandible dark brown or brown in anterior view] | G. sinarum Kieffer, 1911 |

|
||
| – | Hypopygium yellowish-brown or yellow (aa); mesoscutum between punctures rather shiny and sparsely punctulate (bb); head shorter in dorsal view (cc); face with long setosity laterally (dd) | 31 |

|
||
| 31 | Head slightly elongated ventrally (a); mandible largely dark brown in anterior view (b); mesosoma dorsally black (c), hind coxa (d), hind femur (e) and hind basitarsus (f) dark brown | G. dimidiatum Semenov, 1892 |

|
||
| – | Head short ventrally (aa); mandible yellow in anterior view (bb); mesosoma dorsally (cc), hind coxa (dd) and femur (ee) reddish or orange brown; hind basitarsus mainly ivory (ff) | G. argentifrons Semenov T.-S. & Kostylev, 1928 |

|
||
| 32 | Notauli with transverse rugae posteriorly and shallow (a) and anteriorly narrow, finely crenulate or nearly smooth (b); middle lobe of mesoscutum very finely transversely rugulose (c) and vertex (d) dull and finely coriaceous (as temple dorsally (e)) | G. reductum sp. nov. |

|
||
| – | Notauli only crenulate posteriorly and distinctly impressed (aa) and anteriorly wider and moderately crenulate (bb); middle lobe of mesoscutum very finely coriaceous (cc); if finely transversely rugulose, then vertex (dd) and temple dorsally (ee) shiny and largely smooth | 33 |

|
||
| 33 | Occipital carina non-lamelliform medio-dorsally and narrow laterally (a); head gradually narrowed in dorsal view (b) and short (c); vertex in lateral view more (d) or less (dd) above level of ocelli; head dorsally with satin sheen (e) | 34 |

|
||
| – | Occipital carina narrowly lamelliform medio-dorsally and moderately wide laterally (aa); head usually directly narrowed posteriorly in dorsal view (bb); if gradually narrowed (bbb), then head longer (cc); vertex in lateral view near level of ocelli (dd); head dorsally more or less shiny (ee) | 35 |

|
||
| 34 | Mesoscutum densely punctate (a) and punctulate-coriaceous between punctures (b); pronotal side more or less rugose or rugulose ventrally (c); white or ivory apical part of ovipositor sheath 1.1–1.9× as long as hind basitarsus (d) | G. subtile (Thomson, 1883) |

|
||
| – | Mesoscutum at most sparsely punctulate (aa) and very finely coriaceous between minute punctures (bb); pronotal side entirely finely coriaceous ventrally (cc); white or ivory apical part of ovipositor sheath 2.1–2.6× as long as hind basitarsus (dd) | G. pedion sp. nov. |

|
||
| 35 | Head elongate-elliptical in dorsal view (a); propleuron 1.1–1.2× as mesoscutum in front of tegula (b); mesosoma more slender in lateral view (c); mesoscutum less convex dorsally (d); hind tibia slender (e) | G. amoyense Pasteels, 1958 |

|
||
| – | Head trapezoid in dorsal view (aa); propleuron 0.9–1.0× mesoscutum in front of tegula (bb); mesosoma less slender in lateral view (cc); mesoscutal lobes distinctly convex dorsally (dd); hind tibia more robust (ee) or slender (e) | 36 |

|
||
| 36 | Pale apical part of ovipositor sheath 3.0–3.5× as long as hind basitarsus (a); fourth antennal segment 1.7–2.3× as long as third antennal segment (b); vertex shiny and largely smooth or finely punctulate (c), without shallow depression medio-posteriorly (d) and in lateral view distinctly convex (e); [hind basitarsus black] | G. tonkinense Pasteels, 1958 |

|
||
| – | Pale apical part of ovipositor sheath 1.1–2.4× as long as hind basitarsus (aa); fourth antennal segment 1.2–1.9× as long as third antennal segment (bb); head dorsally with satin sheen and finely sculptured (cc), rather often with shallow depression medio-posteriorly (dd; G. japonicum) and vertex less convex (ee) | 37 |

|
||
| 37 | Lobes of mesoscutum distinctly convex in lateral view (a); middle mesoscutal lobe more protuberant in dorsal (b) and lateral (c) view; fourth antennal segment 1.7–1.9× as long as third segment (d); hind tibia more slender (e); [occipital carina fine and non-lamelliform dorsally; vertex in front of occipital carina without depression] | G. sinepunctatum Zhao, van Achterberg & Xu, 2012 |

|
||
| – | Lobes of mesoscutum comparatively flat in lateral view (aa); middle mesoscutal lobe less protuberant in dorsal (bb) and lateral (cc) view; fourth antennal segment 1.2–1.5× as long as third segment (dd); hind tibia less slender (ee); [hind basitarsus often partly ivory] | G. japonicum Cameron, 1888 |

|
||
Males
| 38 | Vertex with distinct medio-posterior depression or a groove with two minute tubercles in front of distinctly lamelliform occipital carina (a); mesoscutum medially transversely or obliquely rugulose, in large specimens transversely rugose (b); occipital carina wide lamelliform (c) | 39 |

|
||
| – | Vertex flat or evenly convex in front of occipital carina (aa); if slightly depressed (aaa), then mesoscutum mainly punctate medially (bb) or occipital carina narrow lamelliform (cc) | 41 |
|
||
| 39 | Propleuron moderately robust and 0.8–1.0× as long as mesoscutum in front of tegula (a); fifth antennal segment 2.2–2.9× as long as third segment (b); head shorter in dorsal view (c) | G. oshimense Watanabe, 1934 |

|
||
| – | Propleuron slender and 1.0–1.2× as long as mesoscutum in front of tegula (aa); fifth antennal segment 1.8–2.1× as long as third segment (bb); head comparatively long in dorsal view (cc) | 40 |

|
||
| 40 | Middle lobe of mesoscutum with transverse elements (a); mesoscutum distinctly rugose medio-posteriorly (b); hind femur slightly more slender (c) | G. corniculigerum Enderlein, 1913 |

|
||
| – | Middle lobe of mesoscutum without transverse elements, only coriaceous between punctures (aa); mesoscutum only punctate medio-posteriorly (bb); hind femur slightly widened (cc) | G. kexinae sp. nov. |

|
||
| 41 | Clypeus with rather large shallow depression (a); hind basitarsus rather stout (b); mesoscutum reticulate or rugose (c); [head and scapus more or less orange or reddish-brown, but sometimes entirely black] | G. hastator (Fabricius, 1804) |

|
||
| – | Clypeus with small depression or depression obsolescent (aa); hind basitarsus often more slender (bb) or mesoscutum coriaceous or rugulose (cc) | 42 |

|
||
| 42 | Head in anterior view distinctly protruding below lower level of eyes (a), in lateral view condylar incision of malar space remains distinctly removed from eye, malar area behind indentation square and at least 0.8× as long as second antennal segment (= pedicellus) and 0.6–0.9× basal width of mandible (b); [mesoscutum densely coriaceous and matt, similar to vertex] | G. minutum (Tournier, 1877) |

|
||
| – | Head in anterior view hardly protruding below lower level of eyes (aa), in lateral view condylar incision of malar space close to eye and malar area behind indentation transverse and 0.3–0.5× as long as second antennal segment and 0.2–0.3× basal width of mandible (bb), rarely slightly longer (aaa, bbb) | 43 |

|
||
| 43 | Mesoscutum only coriaceous or finely rugulose medially (a), at most with a few shallow punctures; [♂ unknown of G. assectoides, G. granulatum and G. pannuceum and are provisionally included] | 44 |

|
||
| – | Mesoscutum with several distinctly impressed punctures medially (aa; but often shallow in G. japonicum) or reticulate-rugose (aaa) | 60 |

|
||
| 44 | Hind tibia slender, hardly to moderately inflated (a); mesoscutum often very finely and regularly sculptured (b) | 45 |

|
||
| – | Hind tibia distinctly inflated (aa); mesoscutum mainly densely coriaceous or irregularly rugulose (aa), especially near notauli (bb) | 54 |

|
||
| 45 | Mesoscutum distinctly regularly transversely rugulose (a); hind tibial spurs yellowish and distinctly contrasting with dark hind basitarsus (b) | G. assectoides Zhao, van Achterberg & Xu, 2012 |
|
||
| – | Mesoscutum mainly finely coriaceous or superficially irregularly rugulose (aa); hind tibial spurs more or less brown and less contrasting with hind basitarsus (bb) | 46 |

|
||
| 46 | Fourth antennal segment 1.8–3.5× as long as third segment (a); face rather narrow (b) | 47 |

|
||
| – | Fourth antennal segment 1.4–1.7× as long as third segment (aa; unknown of G. pannuceum and G. granulatum); face wide (bb) | 50 |

|
||
| 47 | Hind femur rather robust (a); second antennal segment (= pedicellus) more slender (b); middle lobe of mesoscutum less protuberant in lateral view (c); [fourth antennal segment 2.5–2.9 × as long as third segment] | G. huangshii Tan & van Achterberg, 2016 |

|
||
| – | Hind femur slender (aa); pedicellus robust (bb); middle lobe of mesoscutum more protruding in lateral view (cc) | 48 |

|
||
| 48 | Mandible medially yellow (a); fourth antennal segment 2.5–3.5× as long as third segment (b); vertex largely smooth or densely punctulate (c) | G. sinepunctatum Zhao, van Achterberg & Xu, 2012 |

|
||
| – | Mandible medially black or dark brown (aa); fourth antennal segment 1.8–2.3× as long as third segment (bb); vertex coriaceous (cc) | 49 |
|
||
| 49 | Mesoscutum with fine transverse rugulae (a); propleuron less elongate (b); middle lobe of mesoscutum less protruding (c) | G. reductum sp. nov. |

|
||
| – | Mesoscutum with transverse rugulae absent or with very fine rugulae (aa); propleuron more elongate (bb); middle lobe of mesoscutum more protruding (cc) | G. pedion sp. nov. |

|
||
| 50 | Head strongly convex in lateral view (a); head longer in dorsal view (b); hind coxa and tibia rather robust (c); [mandible dark brown] | G. parvicollarium Enderlein, 1913 s. str. |

|
||
| – | Head moderately convex in lateral view (aa); head rather short in dorsal view (bb); hind coxa and tibia slender (cc) | 51 |

|
||
| 51 | Hind coxa distinctly narrower than hind tibia (a); hind femur narrower medially (b); mesosternal sulcus narrow and largely smooth (c); mandible dark brown basally (d) | G. angulatum Zhao, van Achterberg & Xu, 2012 |
|
||
| – | Hind coxa about as wide as hind tibia (aa); hind femur wider medially (bb); mesosternal sulcus wide and coarsely crenulate (cc); mandible brownish-yellow basally (dd) | 52 |

|
||
| 52 | Anterior half of propleuron parallel-sided and long in ventral view (a); propleuron 1.1–1.2× as long as mesoscutum up to tegula in lateral view (b); head comparatively long in dorsal view (c); [eye elongate in lateral view; ♂ unknown] | G. amoyense Pasteels, 1958 |

|
||
| – | Anterior half of propleuron narrowed anteriorly and rather short in ventral view (aa); propleuron 0.8–0.9× as long as mesoscutum up to tegula in lateral view (bb); head short in dorsal view (cc) | 53 |

|
||
| 53 | Mesoscutum densely and finely granulate (a); scutellum granulate and rather flat (b); frons granulate and matt (c); [hind tibia slender and yellowish-brown ventro-basally] | G. granulatum sp. nov. |

|
||
| – | Mesoscutum rugulose or punctate (aa); scutellum rugulose and slightly more convex (bb); frons largely smooth, and more or less shiny (cc); [♂ unknown] | G. pannuceum Tan & Achterberg, 2016 |

|
||
| 54 | Frons and vertex shiny and smooth (a); mesoscutum rugose medio-posteriorly (b) and near notauli (c); hind femur slightly wider (c); [mesoscutum with satin sheen] | G. latitibia Zhao, van Achterberg & Xu, 2012 |
|
||
| – | Frons and vertex matt or with satin sheen, micro-sculptured (aa); mesoscutum medio-posteriorly rugulose (bb) or mainly punctate and near notauli coriaceous or rugulose (cc); hind femur usually more slender (dd) | 55 |

|
||
| 55 | Apex of paramere yellowish-brown (a); pre-apical metasomal sternite distinctly yellowish posteriorly (b); mandible yellowish basally (c) | G. flavimarginatum van Achterberg, 2014 |

|
||
| – | Apex of paramere dark brown (aa); pre-apical metasomal sternite at most narrowly yellowish posteriorly (bb); colour of mandible variable, often darkened basally (cc) | 56 |

|
||
| 56 | Mandible largely pale yellowish (a) | 57 |

|
||
| – | Mandible largely brown, dark brown or black (aa); [additional females needed for reliable identification] | 58 |

|
||
| 57 | Middle lobe of mesoscutum distinctly protruding (a); head longer and rather trapezoid in dorsal view (b); hind tibia dark brown or blackish ventrally (c); scutellum wider (d) | G. brevicuspis Kieffer, 1911 |

|
||
| – | Middle lobe of mesoscutum hardly protruding (aa); head shorter and transverse in dorsal view (bb); hind tibia more or less yellowish-brown ventrally (cc); scutellum narrower (dd) | G. bicoloratum Tan & van Achterberg, 2016 |

|
||
| 58 | Hind tibia with ill-defined pale subbasal patch (a); hypostomal bridge usually medium-sized (b) | G. nigritarse (Thomson, 1883) |

|
||
| – | Hind tibia usually with well-defined subbasal ivory patch (aa); hypostomal bridge narrow (bb) | 59 |

|
||
| 59 | Mesoscutum often coarser sculptured than vertex (a); head directly narrowed behind eyes (b); temple rather convex (c) | G. assectator (Linnaeus, 1758) |

|
||
| – | Mesoscutum slightly coarser sculptured than vertex (aa); head less directly narrowed behind eyes (bb); temple less convex (cc) | G. abeillei Kieffer, 1912 |

|
||
| 60 | Posteriorly vertex flat in lateral view and long (a); head smooth and shiny dorsally (b); head moderately narrowed posteriorly (c); mesosoma laterally often paler than dorsally (d) | G. bimaculatum Pasteels, 1958 |

|
||
| – | Posteriorly vertex convex in lateral view and usually shorter (aa); sculpture of head dorsally variable (bb); if smooth (bbb), then more narrowed posteriorly (cc); mesosoma usually unicoloured (dd) or dorsally paler than laterally | 61 |

|
||
| 61 | Hind coxa orange brown or dark brown (a); mesoscutum conspicuously setose (b); metasoma largely reddish-brown (c); [mandible orange yellow or yellowish-brown; mesoscutum finely or coarsely punctate and often with narrow interspaces] | 62 |

|
||
| – | Hind coxa black (aa); mesoscutum less conspicuously setose (bb); metasoma often largely dark brown or black (cc) | 64 |

|
||
| 62 | Mesoscutum finely punctate, with rather wide interspaces (a); hind tibia yellowish-brown or indistinctly infuscate basally (b); apical half of hind basitarsus partly dark brown and only apically ivory (c) | G. dilutum Semenov, 1892 |

|
||
| – | Mesoscutum coarsely punctate and interspaces narrower (aa); hind tibia distinctly dark brown basally (bb); apical half of hind basitarsus mainly ivory (cc) | 63 |

|
||
| 63 | Basal half of hind coxa rugose dorsally (a); outer side of hind tibia (except basally) dark brown or blackish (b); head subtruncate posteriorly in dorsal view (c) | G. coloratum Zhao, van Achterberg & Xu, 2012 |

|
||
| – | Basal half of hind coxa superficially coriaceous dorsally (aa); ventral half of outer side of hind tibia orange-brown (bb); head emarginate posteriorly in dorsal view (cc) | G. argentifrons Semenov-T.-S. & Kostylev, 1928 |

|
||
| 64 | Third antennal segment slightly longer than wide and similar to pedicellus (a); ventral half of pronotal side largely superficially coriaceous to nearly smooth, only grooves crenulate (b); pronotum partly densely setose (c) | 65 |

|
||
| – | Third antennal segment less robust compared to pedicellus and distinctly longer than wide (aa); ventral half of pronotal side largely moderately reticulate-rugose, at most ventrally coriaceous (bb); pronotum sparsely setose (cc) | 66 |

|
||
| 65 | Malar space narrow (a); head less emarginate medio-posteriorly (b); mesoscutum usually with less coarse sculpture (c); scutellum mainly micro-sculptured (d), at most with few large punctures | G. shengi Tan & van Achterberg, 2016 |

|
||
| – | Malar space slightly wider (aa); head distinctly emarginate medio-posteriorly (bb); mesoscutum with coarser sculpture (cc); scutellum coarsely punctate (dd) | G. dimidiatum Semenov, 1892 |

|
||
| 66 | Hind tibia distinctly inflated and bulging ventrally (a); head directly narrowed posteriorly in dorsal view (b); third antennal segment slender, 1.6–1.9× as long as second segment (c); [mesoscutum distinctly “crater-like” punctate; head concave medio-posteriorly] | 67 |

|
||
| – | Hind tibia slender and hardly bulging ventrally (aa); head usually gradually narrowed posteriorly in dorsal view (bb); third antennal segment robust, 1.2–1.7× as long as second segment (cc) | 68 |

|
||
| 67 | Hind tibia strongly inflated (a); third antennal segment slightly more robust (b) | G. sinicola (Kieffer, 1924) |

|
||
| – | Hind tibia less inflated (aa); third antennal segment slender (bb) | G. formosanum Enderlein, 1913 |

|
||
| 68 | Occipital carina moderately wide (a); vertex distinctly convex (b); [middle lobe of mesoscutum coarsely punctate laterally] | G. tonkinense Pasteels, 1958 |

|
||
| – | Occipital carina narrow lamelliform or non-lamelliform (aa); vertex comparatively flat (bb) | 69 |

|
||
| 69 | Occipital carina narrow lamelliform (a) and vertex slightly depressed medio-dorsally (b); mesoscutum less coarsely punctate (c) | G. japonicum Cameron, 1888 |

|
||
| – | Occipital carina less or non-lamelliform (aa) and vertex flat medio-dorsally (bb); mesoscutum more or less coarsely punctate (cc) | 70 |

|
||
| 70 | Lateral lobe of mesoscutum rather matt and at most finely punctate (a); head slightly enlarged below eyes in anterior view (b) | G. subtile (Thomson, 1883) |

|
||
| – | Lateral lobe of mesoscutum with satin sheen and with coarse punctures (aa); head not enlarged below eyes in anterior view (bb) | G. sinarum Kieffer, 1911 |

|
||
Systematics
Gasteruption abeillei
Gasteruption abeillei
Kieffer, 1912: 228, 231, 251;
Trichofoenus breviterebrae
Watanabe, 1934: 285;
Additional material
3 ♀ + 3 ♂ (

Distribution
China (Shaanxi), Russia (Far East), Europe. New for Shaanxi.


Gasteruption amoyense
Gasteruption amoyense
Pasteels, 1958: 178–179;
Gasteruption curiosum
Pasteels, 1958: 177–178;
Additional material
1 ♀ (

Notes
Similar to the West Palaearctic G. syriacum Szépligeti, 1903, because of the slender pronotum (Fig.

Distribution
China (Fujian, Hong Kong, Hunan, Shaanxi, Zhejiang). New for Shaanxi and Fujian; Shaanxi is the first record from the Palaearctic Region.
Gasteruption angulatum
Gasteruption angulatum
Additional material
1 ♂ (
Distribution

Notes
The male from Baolongyu (Qinling Mts, Shaanxi) illustrated by

Gasteruption assectator
Ichneumon assectator Linnaeus, 1758: 566.
Gasteruption assectator;
Gasteruption margotae
Madl, 1987: 225–227. Synonymised with G. assectator (Linnaeus) by
Distribution
China (Heilongjiang; Hubei (1800 m alt.), Jilin (740 m alt.), Shanxi (1148–1700 m alt.), Qinghai (3300–4288 m alt.), Tibet (2800 m alt.), Xinjiang (1148–2425 m alt.), Russia (Far East), Europe.
Notes
The interpretation of the very similar Gasteruption boreale (Thomson, 1883) is problematic because the lectotype is a male and the most reliable differences are found in the setosity of the ovipositor sheath of the female. The somewhat elongated malar space indicates that it belongs to a species of the G. assectator aggregate with entirely adpressed setosity of the ovipositor sheath and the ovipositor sheath is usually somewhat longer than the hind tibia. So far, no females of typical G. boreale are known from China. Gasteruption assectator is easily confused with G. latitibia Zhao, van Achterberg & Xu, 2012; if the head is distinctly convex dorsally and minute pronotal teeth are present, the specimen most likely belongs to the latter species.
Gasteruption bicoloratum
Gasteruption bicoloratum Tan & van Achterberg, 2016: 86–90; van Achterberg et al. 2019: 4.
Additional material
1 ♀ (
Distribution
China (Gansu, Shaanxi), Russia (Far East). New for Gansu.
Gasteruption bimaculatum
Gasteruption bimaculatum
Pasteels, 1958: 191–192 (only holotype ♂);
Gasteruption obscuripenne Pasteels, 1958: 189–190 (p.p.).
Additional material
1 ♀ + 1 ♂ (

Distribution
China (Fujian, Guangxi, Hainan, Henan, Shaanxi, Tibet, Yunnan); Burma. New for Shaanxi and its most northern record.

Notes
The specimens from Shaanxi have the sculpture of the mesoscutum reduced, especially in the female (Fig.

Gasteruption brevicuspis
Gasteruption brevicuspis Kieffer, 1911: 196, 1912: 293–294; van Achterberg et al. 2019: 4.
Gasteruption terebrelligerum
Enderlein, 1913: 324;
Additional material
3 ♀ + 5 ♂ (

Notes
This species is similar to G. assectator (Linnaeus), but differs by having a slightly longer head in dorsal view, the vertex more convex in lateral view, the hind tibia more slender, the pronotal teeth more or less developed and the mandibles brownish-yellow or yellow (dark brown in G. assectator). Closely related to G. bicoloratum Tan and van Achterberg and differs mainly by the shape of the head as illustrated in the key.

Distribution
China (Fujian, Gansu, Hubei, Hunan, Shaanxi, Shanxi, Taiwan, Yunnan), India, Myanmar, Russia (Far East). New for Gansu and Shaanxi.

Gasteruption coloratum
Gasteruption coloratum Zhao, van Achterberg & Xu, 2012: 38–40.
Additional material

Distribution
China (Xinjiang).
Notes
Both specimens reported here are darker than the holotype (and, so far, only known specimen) of G. coloratum, especially the hind leg and the scapus (

Gasteruption corniculigerum
Gasteruption corniculigerum
Enderlein, 1913: 322–323;
Additional material
1 ♀ (

Distribution
China (Fujian, Guangdong, Guangxi, Guizhou, Hainan, Hunan, Shaanxi, Taiwan, Zhejiang). New for Shaanxi and for the Palaearctic Region.

Gasteruption granulatum , sp. nov.
Gasteruption parvicollarium;
Type material
Holotype, ♀ (

Diagnosis
Head of ♀ trapezoid in dorsal view, medio-posteriorly truncate or nearly so, in anterior view hardly protruding below lower level of eyes and mandibular condylus near lower level of eyes; occipital carina narrow and non-lamelliform medio-dorsally; vertex, at most, moderately bulging above upper level of eyes; clypeus with obsolescent depression; propleuron antero-dorsally granulate, rather robust in ventral and lateral; side of pronotum slender and with narrow and weakly crenulated grooves; mesoscutum matt and densely finely granulate; hind tibia slender and basitarsus elongate; hind tibia dark brown to yellowish-brown ventro-basally; apex of ovipositor narrow and with minute dorsal teeth; ovipositor sheath 1.2–1.8× as long as hind tibia and its apex mainly dark brown; apical sternite of ♂ and paramere entirely dark brown.

Easily confused with G. parvicollarium Enderlein, but G. granulatum has the vertex, at most, moderately bulging above upper level of eyes (strongly bulging in G. parvicollarium), head normal in dorsal view (distinctly elongated); apex of ovipositor narrow and with minute dorsal teeth (wider and with coarse dorsal teeth); propleuron less elongate in ventral view (more elongate) and robust in lateral view (somewhat more slender).
Description
Holotype, female, length of body 14.0 mm, of fore wing 6.9 mm.
Head. Vertex and frons with satin sheen, very finely granulate, moderately convex (Fig.
Mesosoma. Length of mesosoma 2.1× its height; propleuron robust and 0.8× as long as mesoscutum in front of tegula (Fig.
Metasoma. Ovipositor sheath 5.3 mm, 0.4× as long as body, 0.5× as long as metasoma and 1.6× as long as hind tibia; ovipositor sheath with dense cover of fine brownish and adpressed setae, its apical half slender; ovipositor apex narrow and its dorsal teeth minute; hypopygium shallowly V-shaped emarginate medio-posteriorly (Fig.
Colour. Black; apical fifth of antenna largely brown; tegulum and mandible brownish-yellow (except narrow dark borders); clypeus latero-ventrally and humeral plate dark brown; third–sixth metasomal tergites very narrowly apically pale greyish, fourth–sixth sternites apically pale greyish; fore and middle legs (except coxae) largely, hind trochantellus, hind femur apico-ventrally, hind tibial spurs, hind basitarsus apically and more or less second-fourth hind tarsal segments, yellowish-brown; remainder of hind leg (including basal half of tibia and excluding coxa), veins and pterostigma dark brown; wing membrane slightly infuscate, except basally; apex of ovipositor sheath mainly dark brown (Fig.
Male. Similar to female (including fine granulate sculpture of mesoscutum and scutellum, Fig.

Variations
According to
Distribution
China (Shaanxi). According
Etymology
The name is derived from granulum (Latin for small grain or seed) because of the granulate mesoscutum and scutellum.
Gasteruption japonicum
Gasteruption japonicum
Cameron, 1888: 134;
Gasteryption (!) sibiricum Semenov, 1892: 24; van Achterberg et al. 2019: 5 (synonymised with G. japonicum).
Gasteruption rufescenticorne
Enderlein, 1913: 324–325 (only female lectotype; not
Gasteruption sinense var. minus
Kieffer, 1924: 78;
Additional material
1 ♀ + 2 ♂ (
Notes
The female lectotype of Gasteruption rufescenticorne Enderlein proved to be a synonym of G. japonicum (syn. nov.).
Distribution
. China (Fujian, Gansu, Heilongjiang, Hubei, Hunan, Inner Mongolia, Jilin, Ningxia, Shaanxi, Shanghai, Sichuan, Taiwan, Xinjiang, Yunnan, Zhejiang); Japan (Honshu, Hokkaido).
Gasteruption kexinae , sp. nov.
Gasteruption rufescenticorne; Zhao, van Achterberg & Xu, 2012: 75–80 (paralectotype; not lectotype).
Type material
Holotype, ♀ (

Diagnosis
Head of ♀ comparatively long and nearly parallel-sided behind eyes in dorsal view, with deep medial depression in front of occipital carina and with pair of shallow lateral depressions (Fig.
Easily confused with G. corniculigerum Enderlein, 1913; differs mainly by the shape of the head in dorsal view (nearly parallel-sided behind eyes in female and distinctly contracted in G. corniculigerum), the sculpture of the middle lobe of the mesoscutum (without fine transverse elements anteriorly; present in G. corniculigerum), the anteriorly more slender propleuron and the shorter pale part of the ovipositor sheath (1.0–2.1× versus 2.8–3.5× as long as hind basitarsus in G. corniculigerum).
Description
Holotype, female, length of body 16.1 mm, of fore wing 6.5 mm.
Head. Vertex and frons with satin sheen and densely punctulate; vertex weakly convex medio-posteriorly in front of deep medio-posterior depression and a pair of shallow elliptical lateral depressions (Figs

Mesosoma. Length of mesosoma 2.2× its height; propleuron rather slender and 1.2× as long as mesoscutum in front of tegula, in ventral view anteriorly distinctly narrowed; pronotal side rugose ventrally, setosity not obscuring sculpture and crenulated grooves wide, antero-ventral tooth small and triangular (Fig.
Metasoma. Ovipositor sheath 14.6 mm, 0.9× as long as body, 1.4× as long as metasoma and 5.6× as long as hind tibia; ovipositor sheath with dense cover of very fine adpressed setae, its apical white part 1.5× as long as hind basitarsus; emargination of hypopygium 0.4× length of hypopygium (Fig.
Colour. Black; antenna (except blackish four basal segments and apical segment) largely dark brown; mandible largely brown (Fig.
Male. Very similar to female (including fine coriaceous sculpture of mesoscutum, Fig.

Variations
Body length of ♀ 14.6–16.1 mm, of ♂ 12.5–16.5 mm; propleuron 1.0–1.2× as long as mesoscutum in front of tegula; ovipositor sheath 0.9–1.0× as long as body; white or ivory apical part of ovipositor sheath 1.0–2.1× longer than hind basitarsus; antenna (except basally) dark brown or brown.
Distribution
China (Fujian, Guangxi, Hainan, Hubei, Hunan, Jiangsu, Ningxia, Shanghai, Taiwan, Zhejiang).
Etymology
Named after the first author of the revision of the Gasteruptiidae from China, Ms Ke-xin Zhao, for her excellent cooperation and taxonomical insight.
Notes
The female lectotype of G. rufescenticorne Enderlein became available after the revision by

Gasteruption latitibia
Gasteruption latitibia Zhao, van Achterberg & Xu, 2012: 62–65.
Additional material
18 ♀ + 5 ♂ (

Distribution
China (Fujian, Guizhou, Hubei, Hunan, Shaanxi (410–1508 m alt.), Zhejiang). New for Shaanxi and Zhejiang.

Gasteruption minutum
Foenus minutus
Tournier, 1877: ix;
Faenus minutus;
Gasteruption minutum;
Foenus longigena
Thomson, 1883: 849;
Gasteruption longigena;
Gasteruption oriplanum
Kieffer, 1911: 210;
Type material
Lectotype
of G. minutum here designated, ♀ (

Additional material
1 ♂ (

Distribution

Notes
Unfortunately, the holotype of G. oriplanum is a male and has the head severely damaged (figs 166–173 in

Gasteruption nigritarse
Foenus nigritarsis
Thomson, 1883: 849;
Gasteruption nigritarse; Schletterer 1885: 310;
Type material
The female lectotype from Lund (Scania, S Sweden) was selected by

Additional material
3 ♀ (

Gasteruption nigritarse (Thomson), female, Shaanxi 176 head lateral 177 details of hypostomal bridge 178 mesosoma lateral 179 mesosoma dorsal 180 fore wing 181 hind leg 182 head anterior 183 head dorsal 184 apex of metasoma ventral 185 details of apex of ovipositor sheath 186 base of antenna 187 apex of antenna.
Distribution
Europe; China (Gansu, Shaanxi). Collecting a couple of this species in the small NWU garden (at the Taibai campus in the very centre of Xi’an) on bush-killer, Cayratia japonica (Thunb.) Gagnep. was a real surprise. New for China, Gansu, Shaanxi and even for the East Palaearctic Region.

Notes
The Chinese specimens have the temple slightly longer than in European specimens (the eye/temple ratio is the same in the examined specimens) and the hind tibia is slightly more inflated. Both are likely part of clinal variation and, therefore, the Chinese specimens are considered to be conspecific.

Gasteruption oshimense
Gasteruption oshimensis Watanabe, 1934: 283–284.
Gasteruption oshimense;
Additional material
2 ♂ (

Distribution
China (Gansu, Guizhou, Henan, Hunan, Jilin, Shaanxi), Japan. New for Gansu.
Notes
One of the most common species in China; large specimens tend to have the head less narrowed, less shiny and more sculptured in dorsal view (Figs

Gasteruption oshimense Watanabe, large female, Shaanxi 198 head lateral 199 mesosoma lateral 200 mesosoma dorsal 201 hind leg 202 apex of ovipositor sheath lateral 203 head anterior 204 head dorsal 205 wings 206 apex of metasoma ventral 207 medio-posterior depression of head latero-dorsal.

Gasteruption parvicollarium
Gasteruption parvicollarium
Enderlein, 1913: 323–324;
Additonal material
2 ♀ (

Notes
The proper recognition of a species depends a lot on the size and quality of the type series and Gasteruption species are no exception. It becomes very complicated if the holotype (and only available type specimen) is a deformed male from Taiwan as in the case of G. parvicollarium Enderlein. The illustrated associated female by


Distribution

Gasteruption pedion , sp. nov.
Type material
Holotype, ♀ (

Diagnosis
Head gradually narrowed in dorsal view and with satin sheen (Fig.

Gasteruption pedion Tan & van Achterberg, sp. nov., female, holotype 235 head lateral 236 mesosoma lateral 237 mesosoma dorsal 238 fore wing 239 hind leg 240 head anterior 241 head dorsal 242 apex of metasoma ventral 243 apex of ovipositor sheath 244 base of antenna 245 apex of antenna.
Easily confused with G. sinepunctatum Zhao, van Achterberg & Xu, 2012, but the new species has the mesoscutal lobes flattened and very finely coriaceous without transverse elements (mesoscutal lobes bumpy and sculpture with very fine transverse elements in G. sinepunctatum), the vertex rather matt and densely finely sculptured with very fine transverse rugulae (rather sparsely to densely punctulate and rather shiny), the mandible black (largely brownish-yellow), the metasoma of ♀ black ventrally (largely yellowish-brown) and the fore coxa black (dark brown).
Description
Holotype, female, length of body 15.0 mm, of fore wing 7.8 mm.
Head. Vertex and frons with rather matt, very finely and densely coriaceous, on vertex mixed with very fine transverse elements; vertex moderately convex in lateral view (Fig.
Mesosoma. Length of mesosoma twice its height; propleuron rather robust and 0.9× as long as mesoscutum in front of tegula, in ventral view rather robust and less narrowed than in G. sinepunctatum; pronotal side entirely granulate-coriaceous, except for wide crenulated grooves and sparsely setose, with obtuse and rather large lobe-shaped tooth antero-ventrally (Figs
Metasoma. Ovipositor sheath 13.6 mm, 0.9× as long as body, 1.3× as long as metasoma and 4.3× as long as hind tibia; ovipositor sheath with dense cover of very fine adpressed setae, its white apical part (becoming ivory more basally) 2.6× as long as hind basitarsus; apical half of hypopygium emarginate medio-posteriorly.
Colour. Black (including mandible); subapically antenna somewhat brownish ventrally; tegula, legs (but coxae black, hind tibia with large ivory ventro-basal patch, ivory basal patch of fore and middle tibia and middle basitarsus (except apex)), veins and pterostigma dark brown; wing membrane slightly brownish; apex of ovipositor white (Fig.
Male. Very similar to female (including fine sculpture of mesoscutum, but usually somewhat coarser (Fig.

Variations
Body length of ♀ 12.3–15.7 mm, of ♂ 12.2–14.7 mm; fourth antennal segment of ♀ 1.5–1.6× as long as third segment; ovipositor sheath 0.9–1.1× as long as body; apical white part of ovipositor sheath 2.1–2.6× as long as hind basitarsus; propleuron 0.8–0.9× as long as mesoscutum in front of tegula.
Distribution
China (Jiangsu, Shaanxi, Yunnan).
Etymology
From “pedion” (Greek for “flat, plain”), because of the flat and evenly coriaceous mesoscutum.
Gasteruption reductum , sp. nov.
Type material
Holotype, ♀ (

Diagnosis
Head in dorsal view distinctly narrowed, moderately convex and medio-posteriorly flat, without a depression in front of occipital carina; mandibular condylus near lower level of eyes; temple dorsally dull and finely coriaceous; fourth antennal segment of ♀ 1.2–1.3× as long as third segment; third antennal segment of ♂ 1.4× as long as second segment; occipital carina non-lamelliform medio-dorsally propleuron; 0.8–0.9× as long as mesoscutum in front of tegula; notauli narrow, finely crenulate and posteriorly reduced and with transverse rugae; mesoscutum rather flat, with satin sheen and without rugae or punctures in lateral view; mesoscutum mainly very finely coriaceous mixed with very fine transverse rugulae, but medio-posteriorly with distinct transverse rugae; hind femur and tibia slender; ovipositor sheath 1.0–1.1× as long as body; apical white part of ovipositor sheath 1.2–2.5× as long as hind basitarsus; apical sternite of ♂ entirely dark brown and paramere densely whitish setose, with its apex dark brown.

Gasteruption reductum shares with G. graciloides van Achterberg, 2019, from Far East Russia, the peculiar sculpture of the mesoscutum and the slender body, but the new species has the notauli narrow, finely crenulate and posteriorly reduced and with transverse rugae (notauli medium-sized, moderately crenulate and posteriorly distinctly impressed and only crenulate in G. graciloides) and the vertex distinctly protruding above level of ocelli (hardly protruding above level ocelli in G. graciloides). The new species differs from G. pedion sp. nov. mainly by the reduced notauli (distinctly impressed in G. pedion), the propleuron distinctly narrowed anteriorly in ventral view (hardly narrowed), the very finely transversely rugulose mesoscutum (only very finely granulate-coriaceous) and the small ocelli (larger).
Description
Holotype, female, length of body 9.1 mm, of fore wing 4.3 mm.
Head. Frons very finely coriaceous and with satin sheen; vertex very finely coriaceous with very fine transverse rugulae, moderately convex and medio-posteriorly flat, without a depression; head rather gradually contracted behind eyes in dorsal view and temples slightly rounded (Fig.
Mesosoma. Length of mesosoma 1.9× its height; propleuron moderately robust, shiny and 0.9× as long as mesoscutum in front of tegula; pronotal side granulate-coriaceous, except for wide crenulate-rugose grooves and sparsely setose, with distinct acute tooth antero-ventrally (Figs
Metasoma. Ovipositor sheath 9.7 mm, 1.1× as long as body, 1.5× as long as metasoma and 5.5× as long as hind tibia; ovipositor sheath with dense cover of fine adpressed setae, its white apical part 1.7× longer than hind basitarsus; apical half of hypopygium emarginate medially.
Colour. Black; apex of apical antennal segment brown; basal half of mandible dark brown and apical half dark reddish-brown; tegulum, humeral plate, veins and pterostigma largely brown; fore and middle tibiae (but basally and apically somewhat paler) and tarsi, trochantelli and hind tarsus dark brown; hind tibia baso-ventrally with elongate ivory patch; hind tibial spurs brown; wing membrane slightly brownish; ivory or whitish apex of ovipositor sheath basally brownish and remainder of sheath dark brown (Fig.
Male. Similar to female (including very fine transverse sculpture of mesoscutum: Fig.

Variations
Body length of ♀ 9.1–11.5 mm, of ♂ 9.5 mm; fourth antennal segment of ♀ 1.2–1.3× as long as third segment; ovipositor sheath 1.0–1.1× as long as body; apical white part of ovipositor sheath 1.2–2.5× as long as hind basitarsus; propleuron 0.8–0.9× as long as mesoscutum in front of tegula.
Distribution
China (Shaanxi, Hebei).
Etymology
Name derived from “reductus” (Latin for “withdrawn”) because of the narrow and posteriorly reduced notauli.
Gasteruption sinarum
Gasteruption sinarum
Kieffer, 1911: 205–206, 1912: 229, 264;
Gasteruption sinense
Kieffer, 1924: 77–78;
Additional material
1 ♀ (

Distribution
China (Anhui, Beijing, Guangdong, Guangxi, Guizhou Henan, Hubei, Hunan, Inner Mongolia, Jiangsu, Liaoning, Ningxia, Shaanxi, Shandong, Shanghai, Tianjin, Zhejiang). New for Shaanxi.


Gasteruption sinepunctatum
Gasteruption sinepunctatum
Zhao, van Achterberg & Xu, 2012: 85;
Additional material
1 ♀ (

Notes
Gasteruption sinepunctatum Zhao, van Achterberg & Xu, 2012, is a large (about 15 mm body length or more) species described from C. China (Zhejiang) with paratypes from Taiwan, Jilin and Tibet. A large (13 mm body length) male from Taiwan has been associated with this species (
Distribution
China (Jilin, Shaanxi, Taiwan, Tibet, Zhejiang).
Gasteruption subtile
Foenus subtilis Thomson, 1883: 847.
Gasteruption subtile;
Gasteruption kriechbaumeri
Schletterer, 1889: 384, 389, 395, 396, 426;
Gasteruption sabulosum
Schletterer, 1889: 390, 396, 423;
Gasteruption poecilothecus Kieffer, 1911: 205.
Gasteruption poecilothecum;
Gasteruption rossicum
Semenov Tian-Shanskij & Kostylev, 1928: 89;
Type material
Lectotype of G. subtile ♀ (ZIL) from Sweden, “Norl” [= Norrland], “subtilis”, “Lectotypus Foenus subtilis Thoms., ♀, K.-J. Hedqvist, det. 1972”. Holotype of G. poecilothecum, ♀ (BMNH), “Type”, B.M. Type 3.a.164”, “Gasteruption poecilothecus Kieff.”, “[Far East Russia or North China], Amoor [= Amur River= Heilongjiang] / 71 25”, “Determined by Dr. Kieffer”.
Additional material
1 ♀ (
Distribution
China (Hebei, Heilongjiang, Inner Mongolia, Jilin, Xinjiang); Europe (alpine-boreal); Mongolia; Far East Russia. New for Inner Mongolia and Shaanxi.
Notes
The specimens identified as G. poecilothecum (including the holotype) fall within the variation range of G. subtile, resulting in its synonymy with the latter species.


Results and discussion
In this paper, we reviewed the Palaearctic Chinese species of Gasteruption and provide a new and extensively-illustrated identification key. The association of the sexes in Gasteruption is problematic in several cases and, for recent revisions, the association is based on morphological similarity of the sexes and similar collection data, keeping in mind that males are, in general, more coarsely sculptured than females and have a shorter head in dorsal view. However, smaller males are less sculptured than larger ones and, often, several species occur together at the same locality. In future, presence of DNA data and of reared specimens may solve the problem, but, for the moment, we have to rely on extensive collections per site and precise comparisons of similar specimens. In a few cases, no males are available with distinctive features and probably non-sexual characters of the female are tentatively used for the inclusion in the key as far as possible.
We reported 14 additional species (of which three are new to science) from Shaanxi, bringing the total for Shaanxi to 18 species, which is 46% of the total 39 species known from China. Although coming closer to the 60% hypothesis proposed by
Acknowledgements
Thanks are due to Sergey Belokobylskij (St. Petersburg), Christer Hansson (Lund), Bernhard Merz (Geneva), David Notton and Gavin Broad (London), Masahiro Ohara (Sapporo), Hege Vårdal (Stockholm), Zoltán Vas and Sándor Csösz (Budapest), Claire Villemant and Agnièle Touret-Alby (Paris) and Dominique Zimmermann (Vienna) for the loan of the types. The research was supported jointly by the Foundation the National Natural Science Foundation of China (NSFC, No. 31872263, 31572300, 31201732), the Agricultural Sci-Tech Innovation Programme of Xi’an Science and Technology Bureau (No.201806116YF04NC12-1) and the Opening Foundation of Shaanxi Key Laboratory for Animal Conservation (Northwest University).
References
- Abeille de Perrin EEA (1879) Essai de classification des espèces françaises du genre Faenus Fabricius. Bulletin de la Societé d’ Histoire Naturelle de Toulouse 13: 260–279.
- Alexander KNA (1983) Gasteruption minutum (Tournier) (Hym., Gasteruptiidae) on Anglesey. Entomologist’s Monthly Magazine 119: 150.
- Bogusch P, van Achterberg C, Šilhán K, Astapenková A, Heneberg P (2018) Description of mature larvae and ecological notes on Gasteruption Latreille (Hymenoptera, Evanioidea, Gasteruptiidae) parasitizing hymenopterans nesting in reed galls. Journal of Hymenoptera Research 65: 1–21. https://doi.org/10.3897/jhr.65.26645
- Cameron P (1888) Descriptions of one new genus and some new species of parasitic Hymenoptera. Proceedings of the Manchester Literary and Philosophical Society 26: 117–137.
- Capron E (1880) Notes on Hymenoptera. Entomologist 13: 89. https://doi.org/10.5962/bhl.part.27902
- Crosskey RW (1951) The morphology, taxonomy and biology of the British Evanioidea (Hymenoptera). Transactions of the Royal Entomological Society of London 102: 247–301. https://doi.org/10.1111/j.1365-2311.1951.tb00749.x
- Dalla Torre CG de [= KW von] (1902) 2. Subfam. Gasteruptionidae: 1063–1075. In: Catalogus Hymenopterorum hucusque descriptorum systematicus et synonymicus 3: 1–1141.
- Dolfuss H (1982) Die Gasteruptioniden des Bezirkes Scheibbs (Niederösterreich) (Insecta: Hymenoptera, Evanioidea, Gasteruptionidae). Entomofauna 3(2): 21–25.
- Ferrière C (1946) Les Gasteruption de la Suisse (Hym. Evaniidae). Mitteillungen der Schweizerischen Entomologischen Gesellschaft Bern 20: 232–248.
- Hanson PE, Gauld ID (1995) The evaniomorph parasitoid families. Chapter 8. In: Hanson PE, Gauld ID (Eds) The Hymenoptera of Costa Rica. Oxford University Press, Oxford.
- Hanson PE, Gauld ID (2006) Hymenoptera de la région neotropical. Memoirs of the American Entomological Institute 77: 1–994. [Enlarged translation of: Hanson, PE, ID Gauld (Eds), 1995. The Hymenoptera of Costa Rica: 1–893.]
- He J-H (2004) Hymenopteran Insect Fauna of Zhejiang. Science Press, Beijing.
- Hedicke H (1939) Hymenopterorum Catalogus 11, Gasteruptiidae. s’Gravenhage, 54 pp.
- Hedqvist K-J (1973) Notes on the superfamily Evanioidea in Sweden with keys to families, genera and species (Hym., Apocrita). Entomologisk Tidskrift 94(3–4): 177–187.
- Hellén W (1950) Die Evaniiden Finnlands (Hym.). Notulae Entomolgicae 30: 1–5.
- Houston TF (1987) A second contribution to the biology of Stenocolletes bees (Hymenoptera: Apoidea: Stenotritidae). Records of the Western Australian Museum 13: 189–201.
- Jennings JT, Austin AD (1997a) Revision of the Australian endemic genus Hyptiogaster Kieffer (Hymenoptera: Gasteruptiidae), with descriptions of seven new species. Journal of Natural History 31: 1533–1562. https://doi.org/10.1080/00222939700770821
- Jennings JT, Austin AD (1997b) Revision of Aulacofoenus Kieffer (Hymenoptera: Gasteruptiidae), hyptiogastrine wasps with a restricted Gondwanic distribution. Invertebrate Taxonomy 11: 943–976. https://doi.org/10.1071/IT97003
- Jennings JT, Austin AD (2002) Systematics and distribution of world hyptiogastrine wasps (Hymenoptera: Gasteruptiidae). Invertebrate Systematics 16: 735–811. https://doi.org/10.1071/IT01048
- Jennings JT, Austin AD (2004) Biology and host relationships of aulacid and gasteruptiid wasps (Hymenoptera: Evanioidea): a review. In: Rajmohana K, Sudheer K, Girish Kumar P, Santhosh S (Eds) Perspectives on Biosystematics and Biodiversity. University of Calicut, Kerala, 187–215.
- Jennings JT, Parslow B (2014) Gasteruption youngi sp. nov. (Hymenoptera: Evanioidea: Gasteruptiidae) from South Australia; an unusual species with trichoid sensilla on the ovipositor sheaths. Zootaxa 3872(1): 95–100. https://doi.org/10.11646/zootaxa.3872.1.9
- Johansson N, van Achterberg C (2016) Revision of the Palaearctic Gasteruption assectator aggregate (Hymenoptera, Gasteruptiidae), with special reference to Sweden. ZooKeys 615: 73–94. https://doi.org/10.3897/zookeys.615.8857
- Kieffer JJ (1911) Étude sur les évaniides exotiques (Hym.) du British Museum de Londres. Annales de la Société Entomologique de France 80: 151–231. https://doi.org/10.5962/bhl.part.2974
- Kieffer JJ (1912) Evaniidae. Das Tierreich 30: 1–431.
- Kieffer JJ (1924) Evaniides de Chine. Annales de la Societe scientifique de Bruxelles 23: 74–80.
- Kofler A, Madl M (1990) Über Evanioidea von Osttirol (Hymenoptera, Evaniidae, Gasteruptiidae, Aulacidae). Linzer Biologische Beiträge 22(2): 319–324.
- Kozlov MA (1988) Fam. Gasteruptiidae: 244–247. In: Skarlato OA (Ed.) Keys to the fauna of the USSR 158, 3. Hymenoptera 3: 1–268. [Russian] [Translation, 1994: 404–410.]
- Latreille PA (1797) Précis des caractères génériques des insectes, disposes dans un ordre naturel. Paris and Bordeaux, [i–xiv +] 201 pp. [+ 1–7] [Often cited as 1796 but according to Evenhuis (1997), published in 1797] https://doi.org/10.5962/bhl.title.58411
- Lotfalizadeh H, Masudi-Rad S, Mehrvar A (2017) Review of the superfamily Evanioidea (Hymenoptera) in Iran with four new records. Journal of Insect Biodiversity and Systematics 3(2): 141–151.
- Macedo ACC (2009) Generic classification for the Gasteruptiinae (Hymenoptera: Gasteruptiidae) based on a cladistic analysis, with the description of two new Neotropical genera and the revalidation of Plutofoenus Kieffer. Zootaxa 2075: 1–32. https://doi.org/10.11646/zootaxa.2075.1.1
- Macedo ACC (2011) A revision of Gasteruption Latreille (Hymenoptera: Gasteruptiidae) in the Neotropical region. Zootaxa 3030: 1–62. https://doi.org/10.11646/zootaxa.3030.1.1
- Madl M (1987a) Über Gasteruptiidae aus Oberösterreich (Hymenoptera, Evanioidea). Linzer Biologische Beiträge 19(2): 401–405.
- Madl M (1987b) Über Gasteruptiidae aus Niederösterreich (Hymenoptera, Evanioidea). Faunistische Abhandlungen Staatliches Museum für Tierkunde Dresden 15(4): 21–25.
- Madl M (1988) Die Gasteruptiidae des Bundeslandes Salzburg (Hymenoptera, Evanioidea). Verhandlungen der Zoologisch-Botanischen Gesellschaft in Österreich 125: 37–40.
- Madl M (1989a) Über Gasteruptiidae aus Tirol und Vorarlberg (Hymenoptera, Evanioidea). Berichte des naturwissenschaftlich-medizinischen Vereins in Innsbruck 76: 159–163.
- Madl M (1989b) Über Gasteruptiidae aus Jugoslawien (Hymenoptera, Evanioidea). Nachrichtenblatt der Bayerischen Entomologen 38: 40–45.
- Madl M (1990a) Beitrag zur Kenntnis des Gasteruptiidae Griechenlands (Insecta, Hymenoptera, Evanioidea). Faunistische Abhandlungen Staatliches Museum für Tierkunde Dresden 17(14): 127–130.
- Madl M (1990b) Über Gasteruptiidae aus Karnten und Steiermark (Hymenoptera, Evanioidea). Carinthia II 180(100): 479–484.
- Malyshev SI (1965) Lebensweise und Instinkte der primitiven Schlupfwespen Gasteruptiidae (Hymenoptera). Zoologische Jahrbücher. Abteilung für Systematik, Ökologie und Geographie der Tiere 92: 239–288.
- Mao M, Gibson T, Dowton M (2015) Higher-level phylogeny of the Hymenoptera inferred from mitochondrial genomes. Molecular Phylogenetics and Evolution 84: 34–43. https://doi.org/10.1016/j.ympev.2014.12.009
- Mikó I, Rahman SR, Anzaldo SS, van de Kamp T, Parslow BA, Tatarnic NJ, Wetherington MT, Anderson J, Schilder RJ, Ulmer JM, Deans AR, Hines HM (2019) Fat in the leg: Function of the expanded hind leg in Gasteruptiid wasps (Hymenoptera: Gasteruptiidae). Insect Systematics and Diversity 3(1): 1–16. https://doi.org/10.1093/isd/ixy020
- Neumayer J, Schwarz M, Bregant E (1999) Vorläufiges Verzeichnis ausgewählter Hautflügler Kärntens (Hymenoptera ohne Formicidae und Apidae). In: Holzinger WE, Mildner P, Rottenburg T, Wieser C (Eds) Rote Listen gefährdeter Tiere Kärntens Naturschutz in Kärnten 15: 213–231. [Klagenfurt]
- Orlovskytė S, Budrys E, Budrienė A (2018) Check-list of gasteruptiid wasps (Hymenoptera: Gasteruptiidae) of Lithuania, with new data on trophic interactions. Lietuvos Entomologų Draugijos Darbai 2(30): 119–126.
- Pagliano G, Scaramozzino PL (2000) Gasteruptiidae italiani (Hymenoptera: Evanioidea). Bollettino del Museo di Zoologia dell’Università di Torino 17: 5–38. [figs 1–37]
- Parslow BA, Schwarz MP, Stevens MI (2020) Review of the biology and host associations of the wasp genus Gasteruption (Evanioidea: Gasteruptiidae). Zoological Journal of the Linnean Society 189: 1105–1122. https://doi.org/10.1093/zoolinnean/zlaa005
- Pasteels JJ (1958) Révision du genre Gasteruption (Hymenoptera, Evanioidea, Gasteruptionidae). V. Espèces indomalaises. Bulletin et Annales de la Société Royale Entomologique de Belgique 94: 169–213.
- Saure C (2001) Trigonalyoidea, Evanioidea, Stephanoidea: 29–30. In: Dathe HH, Taeger A, Blank SM (Eds) Verzeichnis der Hautflügler Deutschlands (Entomofauna Germanica 4). Entomologische Nachrichten und Berichte, Dresden. Beiheft 7: 1–178.
- Saure C, Schmid-Egger C, van Achterberg C (2017) Order Hymenoptera, family Gasteruptiidae. Arthropod Fauna of the UAE 6: 190–224.
- Scaramozzino PL (1995) Hymenoptera Trigonalyoidea, Evanioidea, Stephanoidea. In: Minelli A, Ruffo S, Posta S (Eds) Checklist delli Specie della Fauna Italiana 94: 1–4.
- Schletterer A (1889) Die Hymenopteren-Gruppe der Evaniiden. Annalen des Kaiserlich-Königliches Naturhistorischen Hofmuseums 4: 373–546.
- Schmidt K (1969) Beiträge zur Kenntnis der Hymenopterenfauna des Mittelrheingebietes, insbesondere des Mainzer Sandes. Mainzer Naturwissenschaftliches Archiv 8: 292–302.
- Schmiedeknecht O (1930) Die Hymenopteren Nord- und Mitteleuropas mit Einschluss von England, Südschweiz, Südtirol und Ungarn nach ihren Gattungen und zum grossen Teil auch nach ihren Arten analytisch bearbeitet, 2nd edn. Jena, 1062 pp.
- Šedivý J (1958) Die tschechoslowakische Arten der Gasteruptioniden (Hym.). Acta Societas Entomologicae Cechosloveniae, Praha (= Časopis Československé Společnosti Entomologické) 55: 34–43.
- Tan J-L, van Achterberg C, Tan Q-Q, Chen X-X (2016) Four new species of Gasteruption Latreille from NW China, with an illustrated key to the species from Palaearctic China (Hymenoptera, Gasteruptiidae). ZooKeys 612: 51–112. https://doi.org/10.3897/zookeys.612.9751
- Thomson CG (1883) XXX. Öfversigt av de I Sverige funna arter av Hymenoptera-slägtet Foenus. Opuscula Entomologica 9: 845–850.
- Tournier H (1877) Tableau synoptique des especes europeennes de genre Foenus Fabr. (Hymenopteres). Annales de Societé Entomologique de Belgique 20: vi–x.
- van Achterberg C (1988) Revision of the subfamily Blacinae Foerster (Hymenoptera, Braconidae). Zoologische Verhandelingen Leiden 249: 1–324.
- van Achterberg C (1993) Illustrated key to the subfamilies of the Braconidae (Hymenoptera: Ichneumonoidea). Zoologische Verhandelingen Leiden 283: 1–189.
- van Achterberg C (2009) Can Townes type Malaise traps be improved? Some recent developments. Entomologische Berichten Amsterdam 69: 129–135.
- van Achterberg C, Grootaert P, Shaw MR (2010) Chapter 17 – Flight interception traps for arthropods. In: Eymann J, Degreef J, Häuser C, Monje JC, Samyn Y, VandenSpiegel D (Eds) Manual on field recording techniques and protocols for All Taxa Biodiversity Inventories and Monitoring. Abc Taxa 1/2, 423–462.
- van Achterberg C (2013) De Nederlandse Gasteruptiidae of Hongerwespen (Hymenoptera: Evanioidea). Nederlandse Faunistische Mededelingen 39: 55–87.
- van Achterberg C, Talebi AA (2014) Review of Gasteruption Latreille (Hymenoptera, Gasteruptiidae) from Iran and Turkey, with the description of 15 new species. ZooKeys 458: 1–188. https://doi.org/10.3897/zookeys.458.8531
- van Breugel P (2014) Gasten van bijenhotels. EIS Kenniscentrum Insecten en andere ongewervelden and Naturalis Biodiversity Center, Leiden, 486 pp. [free download:] www.bijenhotels.nl/pdf/GastenVanBijenhotels.pdf
- van der Smissen J (2010) Teil IV. Beitrag zur Stechimmen-fauna Südfrankreichs (Ardèche, Drôme, Gard, Vaucluse) Hymenoptera Aculeata: Apidae, Chrysididae, Scoliidae, Vespidae, Pompilidae, Sphecidae). Verhandlungen des Vereins für Naturwissenschaftliche Heimatforschung zu Hamburg e.V. 43: 355–415.
- Wall I (1994) Seltene Hymenopteren aus Mittel-, West- und Südeuropa (Hymenoptera Apocrita: Stephanoidea, Evanoidea, Trygonalyoidea). Entomofauna 15(14): 137–184.
- Watanabe C (1934) On Evaniidae and Gasteruptionidae from Japan (Hymenoptera). Transactions of the Sapporo Natural History Society 13(3): 280–286.
- Wisniowski B (2004) Nowe stanowiska gatunkow z rodzaju Gasteruption Latreille, 1796 (Hymenoptera: Gasteruptiidae) w Polsce. Wiadomosci Entomologiczne 23(2): 117–118.
- Zhao K-X, van Achterberg C, Xu Z-F (2012) A revision of the Chinese Gasteruptiidae (Hymenoptera, Evanioidea). ZooKeys 237: 1–123. https://doi.org/10.3897/zookeys.237.3956
- Žikić V, van Achterberg C, Stanković SS, Dubaić JB, Četković A (2014) Review of the Gasteruptiidae (Hymenoptera: Evanioidea) from the territory of the former Yugoslavia, with three newly reported species. Zootaxa 3793(5): 573–586. https://doi.org/10.11646/zootaxa.3793.5.5