Research Article |
|
Corresponding author: Ilgoo Kang ( ikang1@lsu.edu ) Academic editor: Jose Fernandez-Triana
© 2020 Ilgoo Kang, Scott R. Shaw, Nathan P. Lord.
This is an open access article distributed under the terms of the Creative Commons Attribution License (CC BY 4.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Citation:
Kang I, Shaw SR, Lord NP (2020) Two new species and distribution records for the genus Bohayella Belokobylskij, 1987 from Costa Rica (Hymenoptera, Braconidae, Cardiochilinae). ZooKeys 996: 93-105. https://doi.org/10.3897/zookeys.996.59075
|
Abstract
Two new species of Bohayella Belokobylskij, 1987 from Costa Rica are described: Bohayella geraldinae Kang, sp. nov. and Bohayella hansoni Kang, sp. nov. These are new distribution records for the genus in the Neotropical region. In addition, a key to species of the genus Bohayella of Costa Rica is presented. The current work elevates the number of species included in Bohayella from nine to eleven.
Keywords
Morphology, New World, parasitoid wasp, taxonomy
Introduction
Costa Rica is one of the biodiversity hotspots, and a total estimated hymenopteran fauna in the country is ~ 20,000 species, including ~ 2,000 estimated species of braconid wasps (
Cardiochiles nigricans Mao, 1949 (
The first author (IK) had the opportunity to examine Costa Rican cardiochiline specimens housed in University of Wyoming Insect Museum (
Materials and methods
Specimens for this project were provided by
Results
Bohayella
Type species
Bohayella tobiasi Belokobylskij, 1987.
Diagnosis
(based on
Members of the genus can be identified by setose compound eyes (length and density variable); ventro-posteriorly moderately extended gena (Fig.
Key to species of the genus Bohayella of Costa Rica
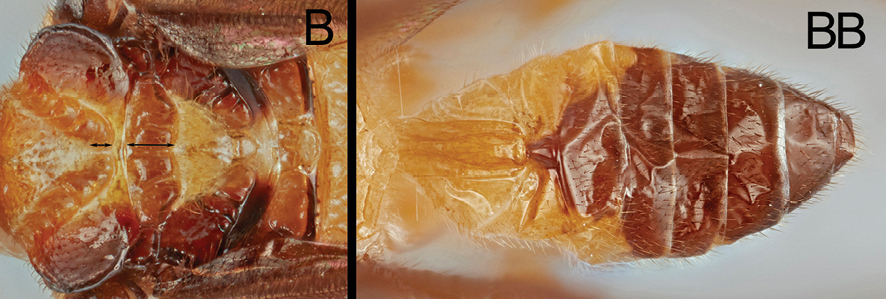
| 1 | Median crenula of notauli as long as median crenula of scutellar sulcus (A); scutellar sulcus with one median crenula (A); T3–T8 mostly pale (AA) | B. geraldinae sp. nov. |

|
||
| – | Median crenula of notauli shorter than median crenula of scutellar sulcus (B); scutellar sulcus with three crenulae (B); T3–T8 mostly melanistic (BB) | B. hansoni sp. nov. |
|
||
Bohayella geraldinae , sp. nov.
Material examined
Holotype Costa Rica • ♀; female, Heredia, 3 km S. Puerto Viejo OTS, La Selva; 100 m; x.1992; P. Hanson; huertos Malaise trap set by G. Wright. Paratypes Costa Rica • 1 ♀; same data as for holotype; xi.1992 • 1 ♂; male; same collecting data as for preceding; 10°26'N, 84°01'W; 4. iv. 1987; H. A. Hespenheide.
Diagnosis
Bohayella geraldinae sp. nov. can be recognized by the following combination of characters: apical maxillary palpomere as long as fifth maxillary palpomere; median crenula of notauli as long as median crenula of scutellar sulcus; scutellar sulcus with one median crenula; hind basitarsus antero-posteriorly slightly expanded; dorsal metasoma mostly pale.
Description
Female. Body 4.6–4.8 mm. Forewing length: ~ 4.2 mm Antenna length: ~ 4.8 mm. Head. Antenna 34-segmented. Interantennal space with well-developed median carina. POL ~ 1.38× longer than diameter of anterior ocellus (11:8) (Fig.
Male. Body ~ 5.0 mm. Same as female except for the following characters: antenna 32-segmented, melanistic color does not reach the dorsal margin of foramen magnum.
Color. Body mostly pale; the following areas are melanistic: antenna, vertex, frons, dorsal occiput, maxillary palpus, labial palpus, lateral mesonotal lobe (pale basally), lateral scutellum, margin of metanotum, apical fore femur, fore tibia, apical fore tarsus, apical mid femur, mid tibia, apical mid tarsus, apical hind femur, basal and apical hind tibia, apical hind tarsus, posterior T5 and T6 (weakly), entire T7 and T8, ovipositor sheath. Wings entirely infuscate, stigma entirely melanistic.
Host
Unknown.
Distribution
Bohayella geraldinae sp. nov. is known only from the La Selva Biological Station owned and managed by Organization for Tropical Studies (OTS) in Heredia, Costa Rica at an elevation of 100 m (Figs



Etymology
This species is named in honor of Dr Geraldine Wright, a former student of the second author (SRS), Rhodes Scholar, professor in the Department of Zoology in the University of Oxford (United Kingdom), and the person who set the trap that collected the specimens.
Bohayella hansoni , sp. nov.
Material examined
Holotype Costa Rica • ♀; female, Puntarenas, San Vito, Estac. Biol., Las Alturas; 1,500 m; vi.1992; Paul Hanson; traps #1 + #2, Malaise. Paratypes Costa Rica • 2 ♀; same data as for holotype • 2 ♀; same collecting data as for preceding • 1 ♀; female; same collecting data as for preceding; 1,700 m; 11.iv.1993.
Diagnosis
Bohayella hansoni sp. nov. can be distinguished from B. geraldinae sp. nov. by the following characters: apical maxillary palpomere slightly longer than fifth maxillary palpomere; median crenula of notauli ~ 0.38× longer than median crenula of scutellar sulcus; scutellar sulcus with three crenulae; hind basitarsus cylindrical; dorsal metasoma mostly melanistic.
Description
Female. Body 3.9–4.1 mm. Forewing length: 3.9–4.1 mm Antenna length: 4.1–4.5 mm. Head. Antenna 32–34-segmented. Interantennal space with well-developed median carina. POL 1.22× longer than diameter of anterior ocellus (11:9) (Fig.

Color. Body mostly pale; the following areas melanistic: antenna, vertex, frons, dorsal occiput, maxillary palpus, labial palpus, lateral mesonotal lobe (basally pale), lateral scutellum, margin of metanotum, apical fore femur, fore tibia, apical fore tarsus, apical mid femur, mid tibia, apical mid tarsus, apical hind femur, basal and apical hind tibia, apical hind tarsus, T2–T8, ovipositor sheath. Wings entirely infuscate, stigma entirely melanistic.
Male. Unknown.
Host
Unknown.
Distribution
Bohayella hansoni sp. nov. is known only from the Las Alturas Biological research station owned and operated by Stanford University in Las Alturas, San Vito, Costa Rica at the elevations of 1,500 m and 1,700 m (Figs

Etymology
This species is named in honor of Dr Paul Hanson, collaborator and professor at the Escuela de Biología, Universidad de Costa Rica. He worked tirelessly for many years collecting and sorting Costa Rican braconids from Malaise samples. SRS is very grateful for his dedication to Hymenoptera studies.
Discussion
Most genus-level diagnostic characters are shared by both Old World and New World members (B. geraldinae sp. nov. and B. hansoni sp. nov.). None of the New World members have a mostly black body, 5-segmented maxillary palpi, or apically infuscate forewings. The following characters are only shared by New World members: angled Rs vein of forewing (Figs
Specimens of B. hansoni sp. nov. collected at altitudes above 1,500 m have more melanistic metasoma than specimens of B. geraldinae sp. nov. collected at a low altitude of 100 m (Figs
The elevation of Costa Rica ranges from sea level to 3,819 m (
Acknowledgements
The first author is grateful to all members of Louisiana State Arthropod Museum and the Department of Entomology as well as LSU Agricultural Center for financial support. We thank Drs Michael Sharkey and James Whitfield for their invaluable help and advice. We also thank Dr Paul Hanson in the Universidad de Costa Rica for the loan of specimens. This study was partially supported by NSF DEB #1841704 to NPL. Research support for SRS was partly provided by National Science Foundation grant DEB 14-42110 (Dimensions of Biodiversity Program). This work was also supported by Wyoming Agricultural Experiment Station funding to SRS provided through the USDA National Institute of Food and Agriculture, McIntire-Stennis project 1021111. Any opinions, findings, and conclusions expressed are those of the authors and do not necessarily reflect the views of the National Science Foundation.
References
- Abe Y, Nishimura T, Maeto K (2013) Causes of polymorphic melanism and its thermoregulatory function in a parasitoid wasp Meteorus pulchricornis (Hymenoptera: Braconidae). European Journal of Entomology 110(4): 627–632. https://doi.org/10.14411/eje.2013.085
- Aguirre H, Shaw SR, Rodríguez‐Jiménez A (2018) Contrasting patterns of altitudinal distribution between parasitoid wasps of the subfamilies Braconinae and Doryctinae (Hymenoptera: Braconidae). Insect Conservation and Diversity 11(3): 219–229. https://doi.org/10.1111/icad.12265
- Beeson CF, Chatterjee SN (1935) On the biology of the Braconidae (Hymenoptera). Indian Forest Records 1: 105–138.
- Belokobylskij SA (1987) A new genus of the subfamily Cardiochilinae (Hymenoptera, Braconidae) from the USSR Far East. Zoologicheskiy Zhurnal 66(2): 302–304.
- Cameron P (1910) On some Asiatic species of the subfamilies Spathiinae, Doryctinae, Rhogadinae, Cardiochilinae and Macrocentrinae in the Royal Berlin Zoological Museum. Wiener Entomologische Zeitschrift 29: 93–100. https://doi.org/10.5962/bhl.part.23337
- Dangerfield PC (1995) The systematics of the genera of Cardiochilinae (Hymenoptera: Braconidae) with a revision of Australasian species. PhD Thesis. Adelaide, Australia: University of Adelaide, 343 pp. http://hdl.handle.net/2440/18664
- Dangerfield PC, Austin AD, Whitfield JB (1999) Systematics of the world genera of Cardiochilinae (Hymenoptera: Braconidae). Invertebrate Systematics 13(6): 917–976. https://doi.org/10.1071/IT98020
- de Souza AR, Mayorquin AZ, Sarmiento CE (2020) Paper wasps are darker at high elevation. Journal of Thermal Biology 89: 102535. https://doi.org/10.1016/j.jtherbio.2020.102535
- Fernandez-Triana JL, Whitfield JB, Smith MA, Hallwachs W, Janzen DH (2014) Revision of the neotropical genus Sendaphne Nixon (Hymenoptera, Braconidae, Microgastrinae). Journal of Hymenoptera Research 41: 1–29. https://doi.org/10.3897/JHR.41.8586
- Gaston K, Gauld I, Hanson P (1996) The size and composition of the hymenopteran fauna of Costa Rica. Journal of Biogeography 23(1): 105–113. https://doi.org/10.1046/j.1365-2699.1996.00978.x
- Hanson PE, Gauld ID (1995) The Hymenoptera of Costa Rica. Oxford University Press, Oxford, 893 pp.
- Hartshorn GS, Peralta R (1987) Preliminary description of primary forests along the La Selva-Volcan Barva altitudinal transect, Costa Rica. In: Almeda F, Pringle CM (Eds) Tropical Rainforests: Diversity and Conservation. California Academy of Science, San Francisco, 281–295.
- Harris RA (1979) Glossary of surface sculpturing. Occasional Papers in Entomology 28: 1–31.
- Holdridge LR (1967) Life zone ecology. Tropical science center. San Jose, Costa Rica, 266 pp.
- Huddleston T, Walker AK (1988) Cardiochiles (Hymenoptera: Braconidae), a parasitoid of lepidopterous larvae, in the Sahel of Africa, with a review of the biology and host relationships of the genus. Bulletin of entomological research 78(3): 435–461. https://doi.org/10.1017/S0007485300013201
- Kang I, Long KD, Sharkey MJ, Whitfield JB, Lord NP (2020) Orientocardiochiles, a new genus of Cardiochilinae (Hymenoptera, Braconidae), with descriptions of two new species from Malaysia and Vietnam. ZooKeys 971: 1–15. https://doi.org/10.3897/zookeys.971.56571
- Mao YT (1949) The species of ichneumon-flies of the genus Cardiochiles occurring in America north of Mexico. Proceedings of the United States National Museum 99: 229–266. https://doi.org/10.5479/si.00963801.99-3237.229
- McDade LA, Hartshorn GS (1994) La Selva Biological Station. In: McDade LA, Bawa KS, Hespenheide HA, Hartshorn GS (Eds) La Selva: ecology and natural history of a neotropical rain forest. University of Chicago Press, Chicago, 6–14.
- Mercado I, Wharton RA (2003) Mexican cardiochiline genera (Hymenoptera: Braconidae), including a preliminary assessment of species-groups in Toxoneuron Say and Retusigaster Dangerfield, Austin and Whitfield. Journal of Natural History 37(7): 845–902. https://doi.org/10.1080/00222930110097167
- Mora R, Hanson PE (2019) Widespread Occurrence of Black-Orange-Black Color Pattern in Hymenoptera. Journal of Insect Science 19(2): 1–12. https://doi.org/10.1093/jisesa/iez021
- Nixon G (1965) A reclassification of the tribe Microgasterini (Hymenoptera: Braconidae). Bulletin of the British Museum (Natural History) Entomology series 2: 1–284.
- Sanford Jr RL, Paaby P, Luvall JC, Phillips E (1994) Climate, Geomorphology, and Aquatic systems. In: McDade LA, Bawa KS, Hespenheide HA, Hartshorn GS (Eds) La Selva: ecology and natural history of a neotropical rain forest. University of Chicago Press, Chicago, 19–33.
- QGIS Development Team (2019) QGIS Geographic Information System. Open Source Geospatial Foundation Project. http://qgis.osgeo.org
- Wesmael C (1835) Monographie des Braconides de Belgique. Nouveaux Mémoires de l’Academie Royale des Sciences et Belles-lettres Bruxelles 9: 1–252.
- Wilkinson DS (1930) New species and host records of Braconidae. Bulletin of Entomological Research 21(4): 481–487. https://doi.org/10.1017/S0007485300024822
- Yu DS, Achterberg C van, Horstmann K (2012) Taxapad 2012, Ichneumonoidea 2011. Database on USB Flash drive. Ottawa, Ontario. http://www.taxapad.com