Research Article |
|
Corresponding author: Simon van Noort ( svannoort@iziko.org.za ) Academic editor: Bernardo Santos
© 2020 Terry Reynolds Berry, Simon van Noort.
This is an open access article distributed under the terms of the Creative Commons Attribution License (CC BY 4.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Citation:
Berry TR, van Noort S (2020) Revision of the endemic Afrotropical genus Tetractenion (Hymenoptera, Ichneumonidae) with an identification key to genera of Banchinae for the region. ZooKeys 1007: 49-84. https://doi.org/10.3897/zookeys.1007.55543
|
Abstract
The Afrotropical banchine fauna comprises 12 genera: Apophua Morley, Atropha Kriechbaumer, Cryptopimpla Taschenberg, Exetastes Gravenhorst, Glyptopimpla Morley, Himertosoma Schmiedeknecht, Lissonota Gravenhorst, Sjostedtiella Szépligeti, Spilopimpla Cameron, Syzeuctus Förster, Tetractenion Seyrig, and Tossinola Viktorov. A well-illustrated revised key to the genera using high definition images is provided, and the endemic Afrotropical genus Tetractenion is revised, previously represented by two described species. Four new species are described: T. ibayaensis sp. nov., T. pascali sp. nov., T. pseudolutea sp. nov., and T. rosei sp. nov. The first species-level identification key is provided for this rare genus. Based on morphological attributes the hypothesis is presented that the species in this genus are probably nocturnal. All images and online interactive Lucid keys are available at: www.waspweb.org and the associated underlying data is made available as Suppl. materials
Keywords
Atrophini, Banchini, Glyptini, Ichneumonoidea, Lucid identification keys, taxonomy
Introduction
Banchinae is a cosmopolitan group of moderately small to large-sized parasitoid wasps (
Most Banchinae can be readily diagnosed by the following characters: 1) a submetapleural carina anteriorly generally expanded into a lobe; 2) an arched posterior transverse carina of the propodeum; and 3) a dorsal apical notch on the ovipositor (
Morphologically, Banchinae can be subdivided into three tribes namely Banchini, Glyptini, and Atrophini (
Within the Afrotropical region, the subfamily comprises 12 genera and 187 described species: Apophua Morley, Atropha Kriechbaumer, Cryptopimpla Taschenberg, Exetastes Gravenhorst, Glyptopimpla Morley, Himertosoma Schmiedeknecht, Lissonota Gravenhorst, Sjostedtiella Szépligeti, Syzeuctus Förster, Spilopimpla Cameron, Tetractenion Seyrig, and Tossinola Viktorov. A dichotomous identification key to banchine genera within the Afrotropical region was last produced by
Tetractenion, placed in the tribe Banchini, is a very rare genus restricted to the Afrotropical region (
Materials and methods
Photographs
Specimens were either pinned or point mounted on black, acid-free cards for examination (using a Leica M205C stereomicroscope with LED light source), photography, and long-term preservation. Images were taken using the Leica LAS 4.4 system which comprised a Leica Z16 microscope with a Leica DFC450 Camera with a 0.63× video objective attached. The imaging process, using an automated Z-stepper, was managed using the Leica Application Suite V 4.4 software installed on a desktop computer. Diffused lighting was achieved using a Leica Dome. Images of the types held in Musée Royal de l’Afrique Centrale, Tervuren (
Depositories
Codens follow
Nomenclature and abbreviations
The morphological terminology follows
A antenna length, from base of scape to flagellar apex (mm);
B body length, from toruli to metasomal apex (mm);
CT (clypeus transversality index): maximum width of clypeus: median height;
F fore wing length, from tegula to wing apex (mm);
Fln (length index of flagellomere n): length: width of flagellomere n;
IO (inter-ocellar index): shortest distance between posterior ocelli: ocellus diameter;
ML (malar space length index): malar space (shortest distance between mandible base and compound eye): basal mandibular width;
OO (oculo-ocellar index): shortest distance between eye and posterior ocellus: ocellus diameter;
OT (ovipositor sheath-tibia index): length of ovipositor sheath: length of hind tibia.
The first three measurements (absolute measures) were measured on all specimens in the type series, with measurements from the primary type reported separately in brackets if necessary.
Identification keys
Identification keys were produced in two formats to facilitate accessibility by a range of end-users and to meet the requirements of publishing both static and dynamic interactive keys under an open access model (
Results
Key to Banchinae genera of the Afrotropical region

|
||
| 1 | Tergites II–IV with a median pair of (usually) deep oblique grooves that converge anteriorly and diverge posteriorly (A); Cu1 longer than cu-a, such that Cu2 arises below middle of these combined veins (nervellus of Townes) (B) | 2 |

|
||
| – | Tergites II–IV without a median pair of oblique grooves (a); Cu1 often longer than cu-a, but may be shorter (b) | 4 |

|
||
| 2 | Occipital carina strongly curved before junction with hypostomal carina (A); areolet open, i.e., vein 3rs-m absent (B) | Apophua |

|
||
| – | Occipital carina without a strong curve before junction with hypostomal carina (a); areolet closed (b) | 3 |

|
||
| 3 | Malar space 0.5–0.8× as long as basal width of mandible (A); epomia long and strong (B) | Glyptopimpla |

|
||
| – | Malar space 1.4–2.5× as long as basal width of mandible (a); epomia usually absent or indistinct, only represented as a short wrinkle (b) | Sjostedtiella |

|
||
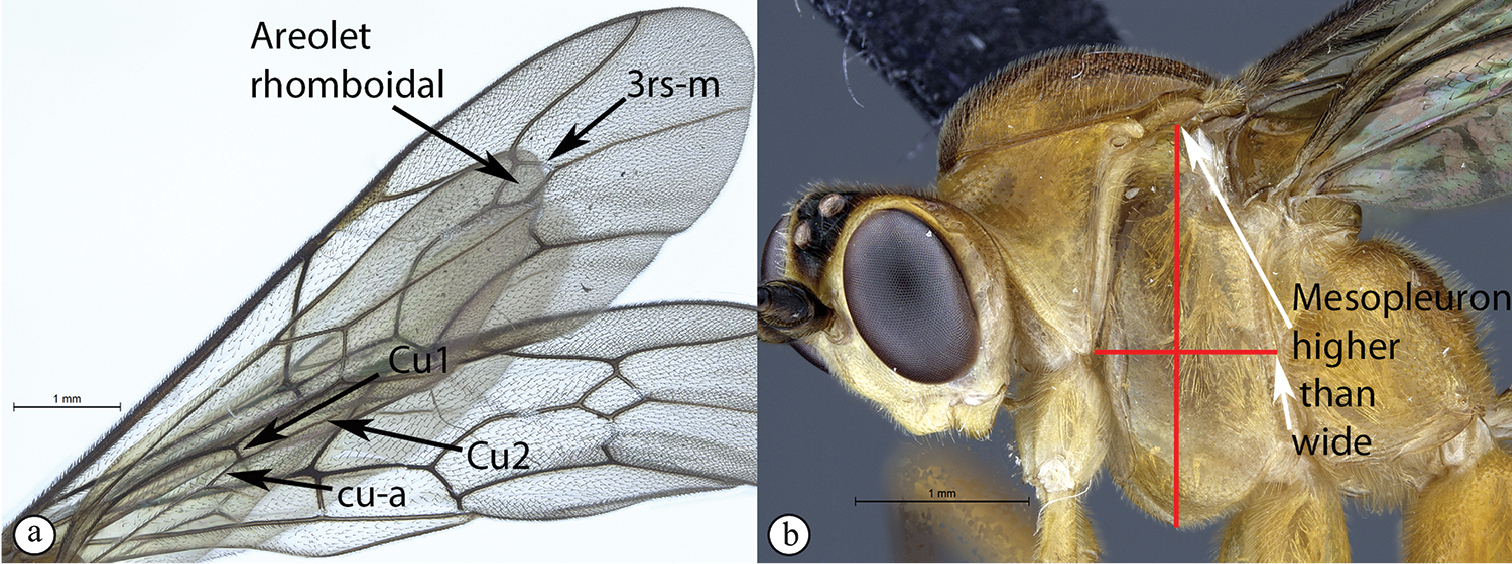
| 4 | Hind wing with Cu1 longer than cu-a such that Cu2 arises below the middle of these combined veins (nervellus of Townes), Cu2 rarely absent (A); fore wing with 3rs-m sometimes lacking, shape of areolet when closed various (A); mesopleuron usually wider than high (B) | 5 |
|
||
| – | Hind wing with Cu1 shorter than cu-a such that Cu2 arises above the middle of these combined veins (nervellus of Townes) (a); fore wing with 3rs-m always present, areolet rhomboidal (a); mesopleuron usually higher than wide (b) | 11 |

|
||
| 5 | Occipital carina joining hypostomal carina at base of mandible (A); epomia usually present (B); propodeal spiracle elliptic (C) | Syzeuctus |

|
||
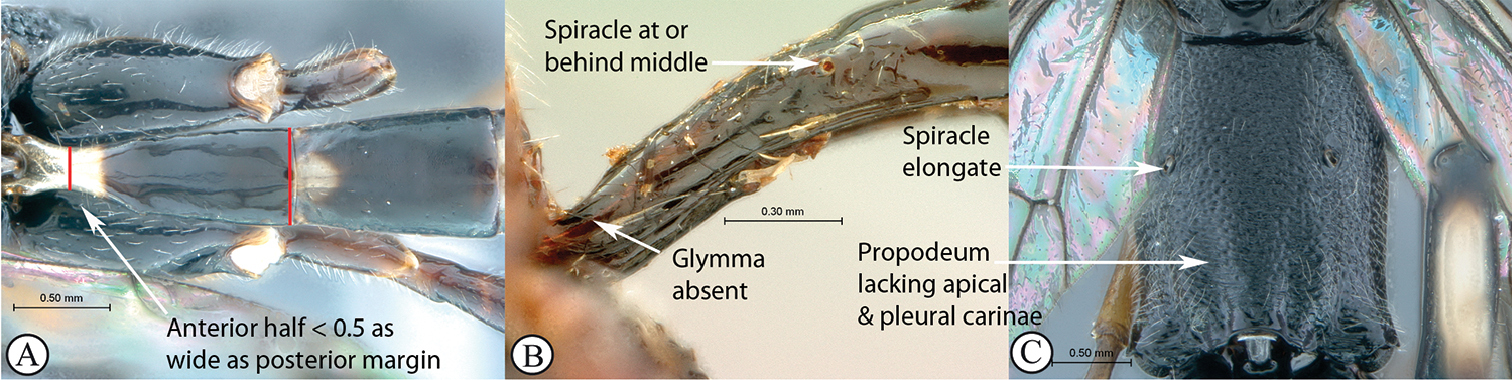
| – | Occipital carina joining hypostomal carina distant from base of mandible (a); epomia usually absent (b); propodeal spiracle circular to elongate (c) | 6 |
|
||
| 6 | Tergite I with anterior half slender, less than half as wide as posterior margin (A); glymma absent (B); with its spiracle at or behind middle (A, B); propodeum lacking carinae (C); propodeal spiracle elongate (C) | Atropha |

|
||
| – | Tergite I with anterior half about or more than half as wide as posterior margin (a); glymma present (a); with its spiracle in front of middle (a, b); propodeum usually with either an apical transverse carina or pleural carina, or both (c); propodeal spiracle usually circular to elongate (c) | 7 |

|
||
| 7 | Apex of submetapleural lobe tooth-like (A); tarsal claws simple with a single basal tooth above (B); areolet open (C); occipital carina broadly interrupted above. | Tossinola |

|
||
| – | Apex of submetapleural lobe rounded (a); tarsal claws simple or pectinate (b); areolet open or closed (c); occipital carina complete. | 8 |

|
||
| 8 | Apical 0.3–0.4 of flagellum tapered to a slender apex (A); ovipositor sheath 0.6–1.2× as long as hind tibia (B) | 9 |

|
||
| – | Flagellum not tapered at the apex (a); ovipositor sheath usually more than 1.4× as long as hind tibia (b) | 10 |

|
||
| 9 | First tergite evenly and rather strongly narrowed anteriorly , about half as wide as posterior margin (A); areolet always truncate-shaped (B); ovipositor 0.6× as long as hind tibia (C) | Cryptopimpla |

|
||
| – | First tergite stout, only moderately narrowed anteriorly, more than half as wide as posterior margin (a); areolet always open (b); ovipositor 0.8–1.2× as long as hind tibia (c). | Spilopimpla |

|
||
| 10 | First tergite nearly always with longitudinal striae (A); areolet open (B) | Himertosoma |

|
||
| – | First tergite rarely covered with longitudinal striae (a); areolet closed or sometimes lacking (b) | Lissonota |

|
||
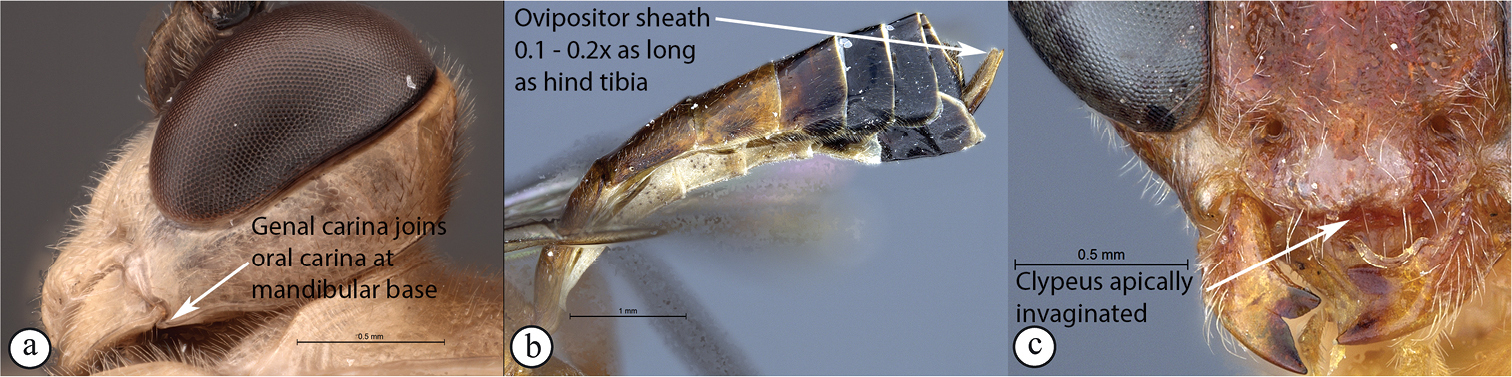
| 11 | Occipital carina joining hypostomal carina above base of mandible (A); ovipositor sheath short to long, 0.14–1.8× as long as hind tibia (B); mandibular teeth usually subequal in length (C); apical clypeal margin normal (C) | Exetastes |
|
||
| – | Occipital carina joining hypostomal carina at base of mandible (a); ovipositor sheath always short, 0.1–0.2× as long as hind tibia (b); lower tooth of mandible always longer than upper tooth (c); clypeus apically invaginated (c) | Tetractenion |
Details of morphological characters that were used to update the key to Afrotropical banchine genera
- Occipital carina with strong curve prior to meeting hypostomal carina: this character (couplet 1A), although noted in the global generic key by
Townes (1969) , was not included in the previously published generic key to Afrotropical Banchinae (Townes and Townes 1973 ). This is a strong and reliable character distinguishing Apophua from the remaining Glyptini genera, Sjostedtiella and Glyptopimpla. - Shape of the areolet when closed: the shape of the areolet has been found to be a useful character to separate the tribes/genera. When distinguishing the tribes Atrophini and Banchini, the areolet in
Exetastes and
Tetractenion is always large and rhomboidal with a very short stalk, whereas in those Atrophini that possess an areolet it is always small, but variably shaped (couplet 4A, a). An anteriorly truncate areolet (couplet 9B, veins 2rs-m and 3rs-m meeting RS separately) present in many
Cryptopimpla species has been reported (
Townes 1969 ;Sheng 2011 ;Takasuka et al. 2011 ) to be a character state that is constant for all Afrotropical Cryptopimpla species (Reynolds Berry and van Noort 2016 ). In Syzeuctus and Atropha, the areolet is triangular with a long stalk and in Lissonota the areolet, when closed, is nearly always petiolate (i.e., dorsal aspect pointed, veins 2rs-m and 3rs-m meet before RS, couplet 10c;Townes 1969 ). - Mesopleuron compressed in Exetastes Group: Tetractenion and Exetastes species have stocky bodies with the mesopleuron laterally compressed (higher than wide) and often flat whereas in Atrophini the mesopleuron is usually wider than high (dorso-ventrally compressed, couplet 4B, b).
- Distinguishing
Tossinola: the length of the ovipositor sheath relative to the hind tibia has previously been used as an additional character to separate
Tossinola from the other Afrotropical genera in the tribe Atrophini where the areolet is open (
Townes and Townes 1973 ). However, the relative lengths overlap across Lissonota, Cryptopimpla and Tossinola species, making it an unreliable character to separate these genera. While a medially, broadly interrupted occipital carina is still the most diagnostic character for the genus Tossinola, another useful character is the state of the apex of the submetapleural carinae: tooth-like in Tossinola (Townes 1969 ) but rounded in the other Afrotropical banchine genera. -
Flagellum apically tapered: as observed by
Townes (1969) , the flagella of the genera Lissonota and Himertosoma are not, or may only be weakly, apically tapered. For Cryptopimpla and Spilopimpla species, the flagella are tapered to a slender apex (couplet 8 A, a;Townes 1969 ;Reynolds Berry and van Noort 2016 ). - Distinguishing
Cryptopimpla: аll Afrotropical
Cryptopimpla possess a first tergite that is evenly and rather strongly narrowed toward the base (couplet 9A;
Reynolds Berry and van Noort 2016 ). In addition, the ovipositor sheath ca. 0.6× as long as the hind tibia is diagnostic of Cryptopimpla species (couplet 9C;Reynolds Berry and van Noort 2016 ). - Distinguishing
Himertosoma from
Lissonota: the absence of a crease separating the fifth laterotergite from the fifth metasomal tergite has been suggested as the single defining character that separates
Himertosoma from
Lissonota (couplet 10A, a;
Watanabe and Maeto 2012 ). However, this does not appear to be a defining character for Afrotropical Lissonota species as the absence/presence of the crease varies within and among species. Given that the areolet can also sometimes be open in Lissonota species, assessment of the sculpture of the first metasomal tergite is required to separate Lissonota from Himertosoma. Himertosoma species nearly always have longitudinal striae present whereas Lissonota species rarely possess either strong punctures or longitudinal striae (couplet 10B, b;Townes 1969 ). - Length of the mandibular teeth: in the global description of the genus
Exetastes by
Townes (1969) , he noted that the length of the lower mandibular tooth relative to the upper could be either equal or slightly longer/shorter. “Slightly” is a poor character description, especially concerning mandibular teeth, which wear out throughout the wasp’s life. In the description of the genus, based on Costa Rican species, byGauld et al. (2002) , all species had equal mandibular teeth. Relative length of the mandibular teeth is a more reliable character, if one of the teeth is markedly longer or shorter. For example, in Afrotropical Cryptopimpla the upper tooth is distinctly longer than the lower tooth (Reynolds Berry and van Noort 2016 ). Similarly, it has been previously noted (Townes 1969 ), and further corroborated during this revision of Tetractenion, that the upper mandibular tooth is distinctly shorter in all species, making it a diagnostic feature for the genus. While most Afrotropical Exetastes have equal mandibular teeth, Exetastes discretus (Morley 1917 ) and an undescribed species in SAMC has mandibles with the lower tooth distinctly longer than the upper. This warrants further investigation, because these two genera are closely related, as they both form part of the Exetastes group. This character may represent a transition between the two genera. - Clypeus apically invaginated: this is a diagnostic feature of
Tetractenion, while in Afrotropical
Exetastes, as has been observed in other species, the clypeal edge is convex or straight, without a median indentation (
Gauld et al. 2002 ).
Tetractenion
Tetractenion Seyrig, 1932, Mém. Acad. Malgache 11: 167. Type: Tetractenion acaule Seyrig. Monobasic.
Diagnosis
(updated from
Biology
Unknown.
Distribution
Angola, Cameroon, Democratic Republic of Congo, Kenya, Madagascar, Namibia, Nigeria, South Africa.
Species richness
T. acaule Seyrig, 1932
T. ibayaensis sp. nov.
T. luteum Seyrig, 1932
T. pascali sp. nov.
T. pseudolutea sp. nov.
T. rosei sp. nov.
Key to Afrotropical species of the genus Tetractenion

|
||
| 1 | Metasomal tergite I distinctly dorso-medially compressed, tergites IV–VIII white where weakly sclerotized (A); keels distinctly raised on mesoscutal lobes (B); notauli abbreviated, not reaching the scutellum | T. acaule |

|
||
| – | Metasomal tergite I with dorso-medial compression weak or absent, tergites IV–VIII strongly sclerotized (a); keels only slightly raised on mesoscutal lobes (b); notauli present, posteriorly meeting before reaching the scutellum | 2 |

|
||
| 2 | Metasomal tergite I stout, ca. as long as wide in dorsal view (A); pits on the mesopleuron and propodeum are large and deep (B) | T. ibayaensis sp. nov. |

|
||
| – | Metasomal tergite I elongate, ca. 2× as long as wide in dorsal view (a); pits on the mesopleuron and propodeum shallow (b) | 3 |

|
||
| 3 | Hind tarsal claw simple (A); wings with dense microtrichia, venation dark (B) | T. luteum |

|
||
| – | Hind tarsal claw pectinate (a); wings usually with sparser microtrichia, venation usually yellowish-brown (b) | 4 |

|
||
| 4 | ML 0.6 (A); head yellow, congruent with yellow body (B) | T. pseudolutea sp. nov. |

|
||
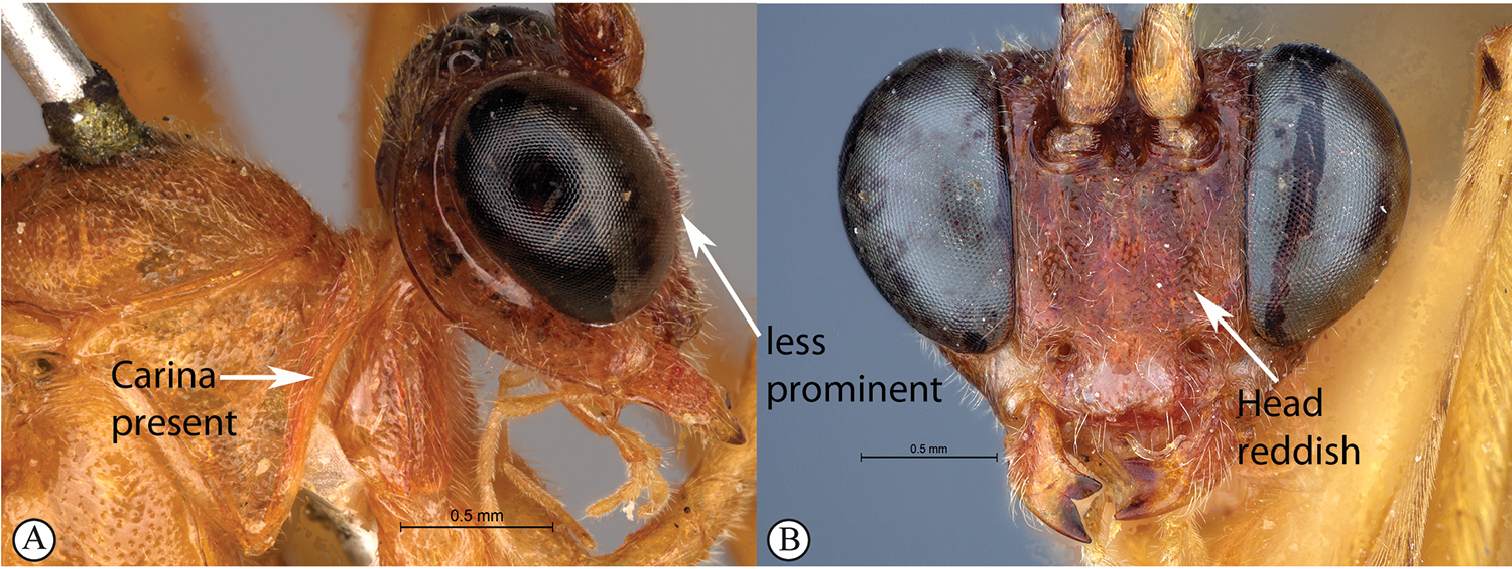
| – | ML 0.7–0.9 (a); head dark purplish-brown or reddish, contrasting with yellow body (b) | 5 |
|
||
| 5 | Pronotal collar with strong carina present (A); head reddish and less robust, face weakly three lobed (B) | T. rosei sp. nov. |

|
||
| – | Pronotal collar weakly wrinkled (a); head dark purplish-brown and more robust, face strongly three-lobed | T. pascali sp. nov. |
Species descriptions
Tetractenion acaule
Type material
Lectotype
♀: Madagascar, Rogez, Forêt Cote Est, Muséum Paris, 1.31. A. Seyrig, EY9333, [White label with TYPE written in red] [Red type label]: Lectotype ♀ Tectractenion acuale, Seyrig, 1932, designated by
Differential diagnosis
Tetractenion acaule is immediately distinguishable from all other Tetractenion species by its unique color combination of a red mesosoma and a mostly black metasoma; distinct keels are present on outer mesoscutal lobes, the notauli do not reach the scutellum; metasomal tergite I has a distinct medial compression in the dorso-ventral view, tergite II have distinct gastrocoeli, and tergites IV–VIII are dorso-posteriorly weakly sclerotized, appearing as large membranous white areas on the dorsal surface. Tetractenion acaule closely resembles T. ibayaensis as both species are similar in color, having largely fulvous bodies with a white face and the hind femur infuscate, whereas the remaining Tetractenion species are largely yellow in color with yellow hind femora. Tetractenion acaule can easily be distinguished from T. ibayaensis by having a white gena and weakly sclerotized metasomal tergites IV–VIII; the head is narrow, straight behind the eyes; a distinct carina is present on the pronotal collar; distinct keels are present on the outer mesoscutal lobes, with the notauli not reaching the scutellum; pits on the mesopleuron and propodeum are shallow; metasomal tergite I is distinctly dorso-medially compressed; gastrocoeli on tergite II are distinct; tergites IV–VIII are postero-dorsally weakly sclerotized and white; and tarsal claws on the hind leg are simple. In T. ibayaensis the gena is brown and tergites IV–VIII are strongly sclerotized; the head is rounded behind the eyes; with no more than a wrinkle present on the pronotal collar; the mesoscutal lobes are hardly present, the notauli reach the scutellum; pits on the mesopleuron and propodeum are deep; metasomal tergite I is stout and indistinctly dorso-ventrally compressed in the medial region; gastrocoeli on tergite II are indistinct; and tarsal claws on the hind legs are pectinate.
Description
(updated from
Head narrow, straight behind eyes; occiput deeply and angularly excavated, occipital carina strong, extending to lower gena at base of mandible; eyes very large; malar space almost half as long as mandibular basal width; face and clypeus finely, evenly and rather sparsely punctate on a shiny background; face with three lobes, tentorial pits deep; clypeus small, laterally convex with declivity, apically invaginated, with clypeal edge convex; mandibular teeth triangular, lower tooth longer than upper tooth; antenna long, slender and apically tapered.
Mesosoma stout; mesonotum deeply punctate, inter-punctuate spaces about as wide as punctures, rather matt, but not coriaceous; keels distinctly raised on outer mesoscutal lobes of mesoscutum, notauli abbreviated, not reaching the scutellum; apex of scutellum rounded; pronotum shining with a distinct thickened carina on collar, sparsely and very finely punctate; mesopleuron higher than wide, sparsely but more deeply punctate, speculum similarly punctate, background hardly shining, epicnemial carina ending at anterior edge of mesopleuron; shallow pits on mesopleuron and propodeum; metapleuron matt and deeply punctate; propodeum weakly convex, roughly punctate dorsally, punctate posteriorly confluently grading into transverse wrinkles, posterior transverse carina reduced, lateral longitudinal carinae present but faint, spiracle roundish-elliptic and small.
Metasoma hardly punctate at base of tergite II, indistinctly punctate beyond base; tergite I elongate, more than twice as long as wide, tapered anteriorly, glymma present, spiracle positioned slightly in front of middle and protruding, especially dorsally, with a distinct medial depression dorso-ventrally; tergite II longer than wide or subquadrate with gastrocoeli distinct; tergite III quadrate to transverse; metasomal tergites IV–VIII moderately laterally compressed; ovipositor sheath concealed or hardly protruding.
Fore wing without ramellus on Rs-M vein; Rs hardly sinuate; areolet large and quadrate with a short stalk receiving 2m-cu at center. Hind wing with Cu1 shorter than cu-a such that Cu2 arises above the middle of these combined veins. Legs very long; hind femur reaching beyond metasomal apex; length of tibia III plus tarsus III as long as body; spurs of tibia III longer than half metatarsal length; tarsal claws on hind leg simple.
Male hardly different: temples a bit less narrowed behind eyes, metasomal tergite II entirely black.
Distribution
Madagascar.

Tetractenion acaule Lectotype (
Tetractenion ibayaensis , sp. nov.
Type material
Holotype
♀: Tanzania, Mkomazi Game Reserve, Ibaya Camp, north west side, 3°57.91'S, 37°48.09'E, 22–24 April 1996, S. van Noort, Acacia/Commiphora/Combretum bushland, Yellow P. Trap, SAM-HYM-P019172 (
Differential diagnosis
Tetractenion ibayaensis is immediately distinguishable from other Tetractenion species by having a largely fulvous body and a white face, with the occiput, gena and metasomal tergites IV–VIII dark brown to black, and the hind tibia and tarsus infuscate. The clypeal and mandibular setae are long. The metasoma is hardly laterally compressed with metasomal tergite I stout, being about as long as wide. Pits on the mesopleuron and propodeum are visibly large and deep. In addition, the clypeus is hardly apically invaginated and the propodeal spiracle is distinctly circular and not circular-elliptical as in the other species.
The head is rounded behind the eyes distinguishing the species from T. acaule and T. pascali. The pronotal collar is no more than a wrinkle, separating the species from T. acaule and T. rosei. Pectinate tarsal claws on the hind legs separates the species from T. acaule and T. luteum. Metasomal tergites II and III are quadrate separating the species from all other Tetractenion species except T. acaule where tergites II and III are sometimes subquadrate and quadrate, respectively; and T. luteum where tergite III is quadrate. Sparse microtrichia on the wings distinguishes T. ibayaensis from T. luteum and T. pascali, and the pterostigma is brown separating the species from T. luteum, T. pascali, T. rosei, and T. pseudolutea.
Description
Body mostly fulvous; tibia and tarsus III brown; metasomal tergites IV–VIII brown to nearly black; head with face and area around eyes white; frons and occiput dark brown to near black; mandibles yellow with base and tips brown. Sparse microtrichia on wings, venation and pterostigma brown.
Head rounded behind eyes; occiput deeply and angularly excavated, occipital carina strong, extending to lower gena at mandibular base; malar space half as long as mandibular basal width; eyes very large; face and clypeus finely and evenly punctate on a shiny background; face with three lobes, tentorial pits deep; clypeus small, laterally convex with declivity, apically invaginated, clypeal edge convex; mandibular teeth triangular, lower tooth longer than upper tooth; clypeal and mandibular setae long; antenna long, slender and apically tapered.
Mesosoma stout and deeply punctate on a shiny background; mesopleuron higher than wide, epicnemial carina ending at anterior edge of mesopleuron; deep pits on the mesopleuron and propodeum; pronotum moderately punctate on a shiny background with no more than a wrinkle on collar; mesososcutal lobes hardly present on mesoscutum, notauli posteriorly meeting before reaching the scutellum; propodeum weakly convex, posteriorly confluently grading into weak transverse wrinkles, posterior transverse carina indistinct, lateral longitudinal carinae reduced, spiracle small and circular.
Metasoma with tergite I stout, tapered anteriorly, not distinctly dorso-ventrally compressed in medial region, glymma present, spiracle positioned in front of middle and protruding, especially dorsally, hardly punctate dorso-laterally, metasoma indistinctly punctate beyond and shining; gastrocoeli on tergite II indistinct; tergites II and III quadrate, tergites IV–VIII only slightly higher than wide.
Fore wing without ramellus on Rs-M vein; areolet large and quadrate with a short stalk receiving 2m-cu at center. Hind wing with Cu1 shorter than cu-a such that Cu2 arises above the middle of these combined veins. Legs very long; hind femur reaching beyond metasomal apex, length of tibia III plus tarsus III as long as body; spurs of tibia III longer than half metatarsal length; tarsal claws pectinate.
CT 2.1; ML 0.5; IO 1.6; OO 1.6; Fl1 4.3; OT 0.2; B 8.1 mm; A 8.1 mm; F 6.4 mm.
Etymology
Named after the type locality. Noun in apposition.

Tetractenion luteum
Type material
Holotype
♂: Nyeri, Kenya, June 1932 (
Differential diagnosis
Tetractenion luteum is immediately distinguishable from the other species in the genus as this species is the only yellow-colored Tetractenion species to possess simple hind tarsal claws, and this character is consistent in both sexes. The head is rounded behind the eyes, distinguishing the species from T. acaule and T. pascali. The malar space nearly as long as the width of the base of the mandible separates T. luteum from T. acaule, T. pseudolutea, and T. ibayaensis. The pronotal collar is weakly wrinkled, separating the species from T. acaule and T. rosei. Metasomal tergite II is longer than wide and distinguishes the species from T. ibayaensis; and a quadrate tergite III separates T. luteum from T. pseudolutea, T. pascali, and T. rosei. Furthermore, T. pascali is the only other species that possess dense microtrichia on the wings.
Description
(updated from
Head with temple short, rounded behind eyes; occiput deeply and angularly excavated, occipital carina strong, extending to lower gena at base of mandible; eyes very large, malar space a bit shorter than width of mandibular base; face and clypeus finely, evenly and rather sparsely punctate on a matt background; face with three lobes, tentorial pits deep; clypeus small, laterally convex with declivity, apically invaginated, with clypeus edge convex; mandibular teeth triangular, lower tooth longer than upper tooth; antenna about as long as body, slender and apically tapered.
Mesosoma stout, matt to sub-polished; pronotum finely punctate on a sub-polished background, no more than a wrinkle present on pronotal collar; mesoscutum moderately punctate, mesoscutal lobes hardly present, notauli posteriorly meeting before reaching the scutellum; mesonotum and mesopleuron finely punctate; mesopleuron higher than wide, epicnemial carina ending at anterior edge of mesopleuron; shallow pits on mesopleuron and propodeum; propodeum weakly convex, matt to sub-polished, moderately punctate posteriorly confluently grading into transverse wrinkles, posterior transverse carina reduced, lateral longitudinal carinae present but faint, spiracle small and circular-elliptical.
Metasoma with a sub-polished background, anterior half of tergite I and dorso-lateral region of tergite II hardly punctate, indistinctly punctate beyond base; tergite I twice as long as wide, glymma present, tapered anteriorly, weak to indistinctly dorso-ventrally depressed in the medial region, spiracle positioned in front of middle and protruding, especially dorsally; tergite II longer than wide, gastrocoeli indistinct; tergite III quadrate.
Fore wing with ramellus absent on Rs-M vein; areolet large and quadrate with a short stalk receiving 2m-cu at center. Hind wing with Cu1 shorter than cu-a such that Cu2 arises above the middle of these combined veins. Legs very long; hind femur reaching beyond metasomal apex, length of tibia III plus tarsus III as long as body, spurs of tibia III longer than half metatarsal length; fore and mid tarsal claws pectinate, hind tarsal claws simple.
Distribution
Democratic Republic of Congo, Kenya, Namibia, Nigeria, and South Africa.

Tetractenion luteum Holotype (
Tetractenion pascali , sp. nov.
Type material
Holotype
♀: Namibia, near Windhoek, between Mandume Ndemufayo Avenue and Western Bypass, 23.xii.2011, SAM-HYM-P047471 (
Differential diagnosis
Tetractenion pascali is immediately distinguishable from all other Tetractenion species by having a color combination of a largely yellow body and a dark head. The facial features are more robust compared to the other species, with the three lobes on the face prominent and the mandibles larger, and the spiracle on the second tergite of the metasoma is hardly protruding. In addition, though the posterior transverse carina may be reduced or faint in the other species, it is distinct in T. pascali. The malar space nearly as long as the width of the mandibular base separates T. pascali from T. acaule, T. pseudolutea, and T. ibayaensis. Pectinate hind tarsal claws distinguish T. pascali from T. luteum and T. acaule; and a weakly wrinkled pronotal collar separates the species from T. acaule and T. rosei. Metasomal tergites II and III are longer than wide separating the species from T. ibayaensis and T. acaule, T. luteum, and T. ibayaensis, respectively. Tetractenion luteum is the only other species besides T. pascali that possess dense microtrichia on the wings.
Description
Color : head brown, mandibles yellow from base to brown at apex. Antennae brown. Body yellow with red-brown areas on metanotum; tibia III with shades of infuscation, tarsus III infuscate. Wings with dense microtrichia, pterostigma yellow, venation brown.
Head narrowed straight behind eyes; occiput deeply and angularly excavated, occipital carina strong, extending to lower gena at mandibular base; malar space nearly as long as basal mandibular width; eyes very large; face and clypeus features robust, mandibles large; face three-lobed and punctate on a shiny background, punctures on second lobe and clypeus deeper than punctures on lobes flanking eyes, tentorial pits deep; clypeus small, laterally convex with declivity, apically invaginated, clypeal edge convex; mandibular teeth triangular, lower tooth longer than upper tooth; antenna long, slender and apically tapered.
Mesosoma stout and moderately punctate on a shiny background; pronotum with no more than a wrinkle on collar; mesoscutal lobes present on mesoscutum, notauli posteriorly meeting before reaching the scutellum; mesopleuron higher than wide, epicnemial carina present at ending at anterior edge of mesopleuron; shallow pits on mesopleuron and propodeum. Propodeum weakly convex, punctate and posteriorly confluently grading into transverse wrinkles, posterior transverse carina present and distinct, lateral longitudinal carinae present but faint, spiracle small and circular-elliptical.
Metasoma indistinctly punctate on a shiny background; tergite I elongate, twice as long as wide, tapered anteriorly, dorso-ventrally compressed in the medial region, glymma present, spiracle positioned in front of middle and hardly protruding; tergite II longer than wide, gastrocoeli indistinct; tergite III longer than wide; tergites IV–VIII higher than wide.
Fore wing without ramellus on Rs-M vein; areolet large and quadrate with a short stalk receiving 2m-cu at center. Hind wing with Cu1 shorter than cu-a such that Cu2 arises above the middle of these combined veins. Legs very long, hind femur reaching beyond metasomal apex, length of tibia III plus tarsus III as long as body; spurs of tibia III longer than half metatarsal length; tarsal claws pectinate.
Males: similar to females; ramellus present.
CT 2–2.4; ML 0.7–0.9; IO 1.2–1.3; OO 1.6–2.1; Fl1 4.5–4.8; OT 0.2; B 7.7–11.5 mm; A 11–14 mm; F 9.2–10 mm.
Etymology
Named after our colleague, Pascal Rousse, who first noted this to be a new species.
Distribution
Namibia and South Africa.
Comments
In males, the ramellus on the fore wing is present, distinguishing the species from T. acaule and T. luteum. The wings of T. rosei are inter-locked; this character could not be compared.

Tetractenion pseudolutea , sp. nov.
Type material
Holotype
♀: Angola (A11), Bruco, 26.ii–2.iii.1972, Southern African Exp. B.M. 1972-1 (
Differential diagnosis
While the color pattern of Tetractenion pseudolutea is identical to T. luteum, it is distinguishable from T. luteum by having pectinate tarsal claw on the hind leg. The head is rounded behind the eyes, separating the species from T. acaule and T. pascali. The pronotal collar with no more than a wrinkle present distinguishes the species from T. acaule and T. rosei. Pectinate tarsal claws on the hind leg separates T. pseudolutea from T. acaule and T. luteum. Metasomal tergites II and III are longer than wide distinguishing T. pseudolutea from T. ibayaensis; and T. acaule, T. luteum, and T. ibayaensis, respectively. Sparse microtrichia on the wings distinguishes the species from T. luteum and T. pascali; yellowish-brown venation separates the species from T. acaule, T. luteum, T. ibayaensis and T. pascali; and a yellow pterostigma distinguishes the species from T. acaule and T. ibayaensis.
Description
The body color is the same as in Tetractenion luteum, except for density of microtrichia on the wings. Tetractenion pseudolutea has sparse microtrichia on the wings with yellow-brown venation, and the pterostigma is yellow.
Head is rounded behind eyes; occiput deeply and angularly excavated, occipital carina strong, extending to lower gena at base of mandible; eyes very large, malar space more than half as long as wide as base of mandible; face and clypeus finely and evenly punctate, background hardly shining; face with three lobes, tentorial pits deep; clypeus small, laterally convex with declivity, apically invaginated, clypeal edge convex; mandibular teeth triangular, lower tooth longer than upper tooth; antennae long, slender and apically tapered.
Mesosoma stout; mesoscutum deeply punctate, mesoscutal lobes hardly present, notauli posteriorly meeting before reaching the scutellum; pronotum finely punctate on a shiny background, no more than a wrinkle present on collar; mesopleuron and mesonotum finely punctate; mesopleuron higher than wide, epicnemial carina ending at anterior edge of mesopleuron; pits on mesopleuron and propodeum are shallow; propodeum weakly convex, finely punctate, posteriorly confluently grading into transverse wrinkles, posterior transverse carina reduced, lateral longitudinal carinae present but faint, spiracle small and circular-elliptical.
Metasoma indistinctly punctate on a shiny background; tergite I twice as long as wide, tapered anteriorly, sometimes weakly dorso-ventrally depressed in the medial region, glymma present, spiracle positioned in front of middle and protruding, especially dorsally; tergite II longer than wide, gastrocoeli indistinct; tergite III longer than wide; tergites IV–VIII moderately laterally compressed.
Fore wing with ramellus rarely present on Rs-M vein; areolet large, quadrate, with a short stalk receiving 2m-cu at the center. Hind wing with Cu1 shorter than cu-a such that Cu2 arises above the middle of these combined veins. Legs very long, hind femur reaching beyond metasomal apex, length of tibia III plus tarsus III as long as body, spurs of tibia III longer than half metatarsal length; tarsal claws pectinate.
CT 2.3; ML 0.6; IO 0.9–1.0; OO 1.7; Fl1 4.6–5.6; OT 0.1; B 9.1–10.7 mm; A 11.3–11.8 mm; F 8.6–9.8 mm.
Etymology
This species at first glance appears to be identical in coloration to T. luteum but has morphological differences.

Tetractenion rosei , sp. nov.
Type material
Holotype
♂: Cameroon, Yaoundé, 1953, C.I.E. Coll. 15098. Pres. by Com. Inst. Ent., B. M. 1962-1. Exetastes sp. ♀ det. J. F. Perkins (
Differential diagnosis
Tetractenion rosei is immediately distinguishable from other Tetractenion species by the reddish color of the head and pronotum in combination with a yellow body, completely yellow legs with venation on the wings also yellow. The head is not narrowed straight behind the eyes but rather rounded, distinguishing the species from T. acaule and T. pascali. The malar space nearly as long as the basal mandible width separates T. rosei from T. acaule, T. pseudolutea, and T. ibayaensis. Tetractenion acaule is the only other species besides T. rosei possessing a thickened and well-defined carina on the pronotal collar.
Pectinate hind tarsal claws separate the species from T. acaule and T. luteum. Sparse microtrichia on the wings distinguishes the species from T. luteum and T. pascali, and the pterostigma is yellow distinguishing the species from T. acaule and T. ibayaensis. Metasomal tergites II and III are longer than wide separating T. rosei from T. ibayaensis; and T. acaule, T. luteum, and T. ibayaensis, respectively.
Description
Color : head and pronotum reddish, black area restricted to region of ocelli. Body, legs, antennae yellow. Wings with sparse microtrichia, venation yellow, pterostigma yellow.
Head rounded behind eyes; occiput deeply and angularly excavated, occipital carina strong, extending to lower gena at mandibular base; malar space nearly as long as basal mandibular width; eyes very large; face and clypeus moderately and evenly punctate on a shiny background; face with three lobes, tentorial pits deep; clypeus small, laterally convex with declivity, apically invaginated, clypeal edge convex; mandibular teeth triangular, lower tooth longer than upper tooth; antenna long, slender and apically tapered.
Mesosoma stout with a shiny background; mesopleuron moderately punctate, epicnemial carina ending at anterior edge of mesopleuron; pits on the mesopleuron and propodeum shallow; mesonotum moderately punctate; pronotum sparsely and finely punctate on a shiny background with a well-defined carina on collar; mesoscutum deeply punctate, mesoscutal lobes hardly present, notauli posteriorly meeting before reaching the scutellum; propodeum weakly convex, deeply punctate posteriorly confluently grading into transverse wrinkles, posterior transverse carina indistinct, lateral longitudinal carinae present, spiracle small and round.
Metasoma indistinctly punctate on a shiny background; tergite I more than twice as long as wide, tapered anteriorly, slight dorso-ventral depression in medial region, glymma present, spiracle in front of middle and protruding; tergites II and III longer than wide; gastrocoeli on tergite II indistinct; tergites IV–VIII higher than wide.
Hind wing with Cu1 shorter than cu-a such that Cu2 arises above the middle of these combined veins. Legs very long, hind femur reaching beyond metasomal apex, length of tibia III plus tarsus III as long as body, spurs of tibia III longer than half metatarsal length; tarsal claws pectinate.
CT 1.6; ML 0.9; IO 1.4; OO 2.2; Fl1 3.5; B 9.3 mm; F 8.6 mm.
Etymology
Named because of the reddish color of the head and pronotal collar. Noun in apposition.
Distribution
Cameroon.
Comments
This is a rare species known only from one female specimen. Sampling in other areas of the Afrotropical region has so far not produced any further specimens. The wings are inter-locked in such a way that a useful diagnostic character of the wings cannot be seen, i.e., whether the ramellus is present or not.

Discussion
Since publication of the first key to genera of Banchinae in the Afrotropical region (
The general habitus and coloration of Tetractenion species suggest that this is possibly a nocturnal genus. A list of characters associated with being nocturnal or crepuscular includes a general brown-yellow color; long antennae; large eyes and large ocelli (
Although there have been recent comprehensive long-term inventory surveys conducted across many parts of Africa with many rich, recently collected bulk samples that still need to be sorted, in reality, comprehensive sampling of Ichneumonidae in the region has been relatively limited to specific areas (
Most of the species have the pro-, meso-, and meta-tarsal claws pectinate to the apex. Tetractenion luteum and T. acaule (a Madagascan endemic) are the only two species that possess simple tarsal claws on the hind leg. While the overall color patterns of T. pseudolutea are identical to T. luteum, it is readily distinguishable from T. luteum by having a pectinate tarsal claw on the hind leg. With most Tetractenion species having pectinate tarsal claws on the hind leg, it is plausible that this character state is the plesiomorphic condition. Based on the assumption that it is more parsimonious for evolutionary trajectories to proceed via the reduction of morphological characters, rather than evolution of more complex character states, T. luteum and T. acaule are most probably the more derived species within the genus, but this hypothesis requires corroboration with the addition of genetic evidence, and a thorough phylogenetic analyses based on both morphological and molecular characters.
The current revision has increased the species richness of the genus threefold. Further comprehensive sampling will undoubtedly uncover additional Tetractenion species in the Afrotropical region.
Acknowledgements
TRB received financial assistance provided by the South African National Research Foundation (NRF) through the Professional Development Program (PDP) and funding from the City of Cape Town. SvN was funded by South African NRF (National Research Foundation) grants: GUN 2068865; GUN 61497; GUN 79004; GUN 79211; GUN 81139; GUN 98115. Part of the South African field work conducted by SvN was funded by the National Science Foundation under PlatyPBI grant no. DEB-0614764 to N.F. Johnson and A.D. Austin. Cape Nature; the Eastern Cape Department of Environmental Affairs and the Northern Cape Department of Nature and Environmental Conservation provided collecting permits for South Africa. This study also benefited from generous contributions of banchine material from the following sources:
References
- Arnett RH, Samuelson GA, Nishida GM (1993) The Insect and Spider Collections of the World. Sandhill Crane Press, Gainsville, 316 pp.
- Bennett AMR, Cardinal S, Gauld ID, Wahl DB (2019) Phylogeny of the subfamilies of Ichneumonidae. Journal of Hymenoptera Research 71: 1–156. https://doi.org/10.3897/jhr.71.32375
- Broad GR (2014) A revision of Sachtlebenia Townes, with notes on the species of Townesion Kasparyan (Hymenoptera: Ichneumonidae: Banchinae). Proceedings of the Russian Entomological Society 85(1): 63–76. https://doi.org/10.47640/1605-7678_2014_85_1_63
- Broad GR, Sääksjärvi IE, Veijalainen A, Notton DG (2011) Three new genera of Banchinae (Hymenoptera: Ichneumonidae) from Central and South America. Journal of Natural History 45(21–22): 1311–1329. https://doi.org/10.1080/00222933.2011.552809
- Choi JK, Kang GW, Lee JW (2015) Two new species of Leptobatopsis Ashmead (Hymenoptera: Ichneumonidae: Banchinae) from South Korea and gynandromorphy in L. nigricapitis. Zootaxa 3964(2): 275–287. https://doi.org/10.11646/zootaxa.3964.2.7
- Fernandes LBDR, Dias Filho MM, Fernandes MA, Penteado-Dias AM (2010) Ichneumonidae (Hymenoptera) parasitoids of Lepidoptera caterpillars feeding on Croton floribundus Spreng (Euphorbiaceae). Revista Brasileira de Entomologia 54(2): 263–269. https://doi.org/10.1590/S0085-56262010000200009
- Fitton MG (1985) The ichneumon-fly genus Banchus (Hymenoptera) in the Old World. Bulletin of the British Museum (Natural History) (Entomology) 51: 1–60. https://www.biodiversitylibrary.org/part/14889#/summary
- Fitton MG (1987) A review of the Banchus group of ichneumon-flies, with a revision of the Australian genus Philogalleria (Hymenoptera: Ichneumonidae). Systematic Entomology 12(1): 33–45. https://doi.org/10.1111/j.1365-3113.1987.tb00545.x
- Gauld ID (1985) The phylogeny, classification and evolution of parasitic wasps of the subfamily Ophioninae (ichneumonidae). Bulletin of the British Museum (Natural History) 51: 1–185. https://www.biodiversitylibrary.org/part/14890#/summary
- Gauld ID (1991) The Ichneumonidae of Costa Rica 1. Memoirs of the American Entomological Institute 47: 1–589.
- Gauld ID, Huddleston T (1976) The nocturnal Ichneumonoidea of the British Isles. Including a key to the genera. Entomologist'S, Gazette 27(1): 35–49. https://www.cabdirect.org/cabdirect/abstract/19760533821
- Gauld ID, Gomez J, Godoy C (2002) Subfamily Banchinae. In: Gauld ID (Ed.) The Ichneumonidae of Costa Rica, 4. Memoirs of the American Entomological Institute 66: 263–746.
- Gauld ID, Mitchell P (1978) The Taxonomy, Distribution and Host Preferences of African Parasitic Wasps of the Subfamily Ophioninae (Hymenoptera: Ichneumonidae). Commonwealth Agricultural Bureau, Slough, 287 pp.
- Gauld ID, Wahl DB (2000) The Townesioninae: a distinct subfamily of Ichneumonidae (Hymenoptera) or a clade of the Banchinae? Transactions of the American Entomological Society 126(3): 279–292. https://www.jstor.org/stable/25078717?seq=1
- Gupta VK (2002) Glyptopimpla Morley (Hymenoptera: Ichneumonidae: Banchinae) – a valid genus with descriptions of new species from the Orient. Oriental Insects 36(1): 221–237. https://doi.org/10.1080/00305316.2002.10417332
- Greiner B (2006) Visual adaptations in the night active wasp Apoica pallens. Journal of Comparative Neurology 495(3): 255–262. https://doi.org/10.1002/cne.20882
- Herrera-Florez AF (2017) A new species of Sphelodon Townes (Hymenoptera: Ichneumonidae: Banchinae) from Colombia. Zootaxa 4277(2): 289–294. https://doi.org/10.11646/zootaxa.4277.2.11
- Hopkins T, Roininen H, van Noort S, Broad GR, Kaunisto K, Sääksjärvi IE (2019a) Extensive sampling and thorough taxonomic assessment of Afrotropical Rhyssinae (Hymenoptera, Ichneumonidae) reveals two new species and demonstrates the limitations of previous sampling efforts. ZooKeys 878: 33–71. https://doi.org/10.3897/zookeys.878.37845
- Hopkins T, Roininen H, Sääksjärvi IE (2019b) Extensive sampling reveals the phenology and habitat use of Afrotropical parasitoid wasps (Hymenoptera: Ichneumonidae: Rhyssinae). Royal Society Open Science 6: 190913. https://doi.org/10.1098/rsos.190913
- Kang GW, Kolarov J, Lee JW (2019) Cryptopimpla (Hymenoptera, Ichneumonidae, Banchinae) of South Korea, with description of two new species. ZooKeys 830: 99–109. https://doi.org/10.3897/zookeys.830.31974
- Kang GW, Kolarov J, Lee JW (2020) A review of South Korean Alloplasta Förster (Hymenoptera: Ichneumonidae:Banchinae) with description of a new species. Zootaxa 4763(2): 270–280. https://doi.org/10.11646/zootaxa.4763.2.9
- Kasparyan DR, Kuslitzky WS (2018) Contribution to the fauna of the ichneumon-wasp genus Rhynchobanchus Kriechbauer, 1894 (Hymenoptera, Ichneumonidae: Banchinae) in the Russian Far East. Entomological Review 98(6): 748–752. https://doi.org/10.1134/S001387381806012X
- Khalaim AI, Ruíz-Cancino E (2012) Mexican species of Exetastes (Hymenoptera: Ichneumonidae: Banchinae), with description of three new species. Revista Mexicana de Biodiversidad 83: 370–379. https://doi.org/10.22201/ib.20078706e.2012.2.953
- Klopfstein S, Santos BF, Shaw MR, Alvarado M, Bennett AM, Dal Pos D, Giannotta M, Herrera Florez AF, Karlsson D, Khalaim AI, Lima AR, Mikó I, Sääksjärvi IE, Shimizu S, Spasojevic T, van Noort S, Vilhelmsen L, Broad GR (2019) Darwin wasps: a new name heralds renewed efforts to unravel the evolutionary history of Ichneumonidae. Entomological Communications 1: ec01006. https://doi.org/10.37486/2675-1305.ec01006
- Li ZJ, Li T, Yan J, Sheng ML (2018) The genus Rhynchobanchus Kriechbaumer in China, with descriptions of a new species and first record of the genus from Oriental region (Hymenoptera, Ichneumonidae, Banchinae). ZooKeys 752: 125–136. https://doi.org/10.3897/zookeys.752.23884
- Morley C (1917) On some South African Ichneumonidae in the collection of the South African Museum. Annals of the South African Museum 17: 191–229.
- Penev L, Sharkey M, Erwin T, van Noort S, Buffington M, Seltmann K, Johnson N, Taylor M, Thompson C, Dallwitz M (2009) Data publication and dissemination of interactive keys under the open access model. ZooKeys 21: 1–17. https://doi.org/10.3897/zookeys.21.274
- Quicke DLJ (2015) The Braconid and Ichneumonid Parasitoid Wasps. Biology, Systematics, Evolution and Ecology. Wiley Blackwell, Oxford, 704 pp. https://doi.org/10.1002/9781118907085
- Reynolds Berry T (2019) Systematics of the parasitoid wasp subfamily Banchinae (Hymenoptera; Ichneumonidae) in the Afrotropical region, PhD thesis, Stellenbosch University, South Africa.
- Reynolds Berry T, van Noort S (2016) Review of Afrotropical Cryptopimpla Taschenberg (Hymenoptera, Ichneumonidae, Banchinae), with description of nine new species. ZooKeys 640: 103–137. https://doi.org/10.3897/zookeys.640.10334
- Seyrig A (1932) Les Ichneumonides de Madagascar. I Ichneumonidae Pimplinae. Mémoires de l’Académie Malgache. Fascicule 11, 183 pp.
- Seyrig A (1935) Mission scientifique de l’Omo. Tome III. Fascicule 18. Hymenoptera, II. Ichneumonidae: Cryptinae, Pimplinae, Tryphoninae et Ophioninae. Mémoires du Muséum National d’Histoire Naturelle, Paris 4: 1–100.
- Sharkey M, Yu D, van Noort S, Seltmann K, Penev L (2009) Revision of the Oriental genera of Agathidinae (Hymenoptera, Braconidae) with an emphasis on Thailand and interactive keys to genera published in three different formats. ZooKeys 21: 19–54. https://doi.org/10.3897/zookeys.21.271
- Sheng ML (2011) Five new species of the genus Cryptopimpla Taschenberg (Hymenoptera, Ichneumonidae) with a key to species known from China. ZooKeys 117: 9–49. https://doi.org/10.3897/zookeys.117.1302
- Sheng ML, Sun SP, Wang XN, Wu HW (2018) A new genus and species of subfamily Banchinae (Hymenoptera, Ichneumonidae) from China. Zootaxa 4413(3): 541–550. https://doi.org/10.11646/zootaxa.4413.3.8
- Shimizu S, Ogawa R (2019) Discovery of the subfamily Lycorininae Cushman and Rohwer, 1920 (Hymenoptera: Ichneumonidae) from Indonesia, based on Lycorina longicauda Shimizu, sp. nov., with a key to the Oriental Lycorina species. Austral Entomology 58(1): 148–155. https://doi.org/10.1111/aen.12347
- Takasuka K, Watanabe K, Konishi K (2011) Genus Cryptopimpla Taschenberg new to Sulawesi, Indonesia, with description of a new species (Hymenoptera, Ichneumonidae, Banchinae). Journal of Hymenoptera Research 23: 65–75. https://doi.org/10.3897/jhr.23.1595
- Townes HK (1969) Genera of Ichneumonidae, Part 3 (Lycorininae, Banchinae, Scolobatinae, Porizontinae). Memoirs of the American Entomological Institute 13: 1–307.
- Townes HK, Townes M (1973) A catalogue and reclassification of the Ethiopian Ichneumonidae. Memoirs of the American Entomological Institute 19: 1–416.
- Tschopp S, Riedel A, Kropf C, Nentwig W, Klopfstein S (2013) The evolution of host associations in the parasitic wasp genus Ichneumon (Hymenoptera: Ichneumonidae): convergent adaptations to host pupation sites. BMC Evolutionary Biology 13(1): 1–74. https://doi.org/10.1186/1471-2148-13-74
- van Noort S (2019) Assessing the status quo of Afrotropical ichneumonid knowledge. Zenodo. http://doi.org/10.5281/zenodo.3395821
- van Noort S (2020) WaspWeb: Hymenoptera of the Afrotropical region. www.waspweb.org [accessed on 20 Feb 2020]
- van Noort S, Buffington ML, Forshage M (2015) Afrotropical Cynipoidea (Hymenoptera). ZooKeys 493: 1–176. https://doi.org/10.3897/zookeys.493.6353
- Vas Z (2017) Data to the Vietnamese ichneumon wasp fauna with description of a new Teleutaea species (Hymenoptera: Ichneumonidae). Folio Entomologica Hungarica 78: 101–110. https://doi.org/10.17112/FoliaEntHung.2017.78.101
- Wahl DB (1988) A review of the mature larvae of the Banchini and their phylogenetic significance, with comments on the Stilbopinae (Hymenoptera: Ichneumonidae). In: Gupta VK (Ed.) Advances in Parasitic Hymenoptera Research. Brill, Leiden, 147–161.
- Wahl DB (1991) The status of Rhimphoctona, with special reference to the higher categories within Campopleginae and the relationships of the subfamily (Hymenoptera: Ichneumonidae). Society 117: 193–213.
- Wahl DB, Sharkey MJ (1993) Chapter 10. Superfamily Ichneumonoidea. In: Goulet H, Huber JT (Eds) Hymenoptera of the World: An Identification Guide to Families. Agriculture Canada, Ottawa, 358–509.
- Warrant EJ (2008) Seeing in the dark: vision and visual behaviour in nocturnal bees and wasps. Journal of Experimental Biology 211: 1737–1746. https://doi.org/10.1242/jeb.015396
- Warrant EJ, Kelber A, Gislén A, Greiner B, Ribi W, Wcislo WT (2004) Nocturnal vision and landmark orientation in a tropical halictid bee. Current Biology 14(15): 1309–1318. https://doi.org/10.1016/j.cub.2004.07.057
- Watanabe K (2017) Revision of the genus Amphirhachis Townes, 1970 (Hymenoptera, Ichneumonidae, Banchinae) from Japan. ZooKeys 685: 49–64. https://doi.org/10.3897/zookeys.685.13552
- Watanabe K (2020) Taxonomic study of the tribe Banchini (Hymenoptera: Ichneumonidae: Banchinae) from Japan. Japanese Journal of Systematic Entomology, Supplementary Series 2: 1–58.
- Watanabe K, Maeto K (2012) A new species of the genus Himertosoma from the Ryukyus, Japan, with a key to species from the Palaearctic and Oriental regions (Hymenoptera, Ichneumonidae, Banchinae). ZooKeys 234: 59–66. https://doi.org/10.3897/zookeys.234.3794
- Watanabe K, Maeto K (2014) Revision of the genus Apophua Morley, 1913, from Japan (Hymenoptera, Ichneumonidae, Banchinae). Zootaxa 3784(5): 501–527. https://doi.org/10.11646/zootaxa.3784.5.1
- Watanabe K, Sheng ML (2018) Taxonomic notes on Exetastes fukuchiyamanus Uchida, 1928 (Hymenoptera, Ichneumonidae, Banchinae), with description of a new species from japan and China. Zootaxa 4399(2): 281–288. https://doi.org/10.11646/zootaxa.4399.2.11
- Yu DSK, van Achterberg C, Hortsmann K (2020) Taxapad 2012, Ichneumonoidea 2011. www.taxapad.com [accessed on 20 Aug 2020]
Supplementary materials
Lucid Interchange Format version 3 (LIF3) for the key to the genera of Afrotropical Banchinae (Hymenoptera, Ichneumonidae)
Data type: Lucid Interchange Format version 3
Explanation note: The LIF3 file is an XML-based file that stores all the Lucid4 key data, allowing exchange of the key with other key developers.
Lucid Interchange Format version 3 (LIF3) and Lucid SDD files for the key to Tetractenion species (Hymenoptera, Ichneumonidae, Banchinae)
Data type: Lucid Interchange Format version 3 (LIF3) and Lucid SDD files
Explanation note: The LIF3 file is an XML-based file that stores all the Lucid4 key data, allowing exchange of the key with other key developers.