Checklist |
|
Corresponding author: Víctor Hugo González-Sánchez ( biologovhgs@gmail.com ) Corresponding author: Larry David Wilson ( bufodoc@aol.com ) Academic editor: Johannes Penner
© 2021 Víctor Hugo González-Sánchez, Jerry D. Johnson, David González-Solís, Lydia Allison Fucsko, Larry David Wilson.
This is an open access article distributed under the terms of the Creative Commons Attribution License (CC BY 4.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Citation:
González-Sánchez VH, Johnson JD, González-Solís D, Fucsko LA, Wilson LD (2021) A review of the introduced herpetofauna of Mexico and Central America, with comments on the effects of invasive species and biosecurity methodology. ZooKeys 1022: 79-154. https://doi.org/10.3897/zookeys.1022.51422
|
Abstract
Among the principal causes producing detrimental effects on global biodiversity are introductions of alien species. Very few attempts to control introduced amphibians and reptiles in Middle America (Mexico and Central America) can be identified, so listings are provided for 24 exotic species, 16 translocated species, and 11 species that were removed from the introduced species listing because of lack of substantiating evidence that they are from established populations. Biosecurity methods are also identified that can be applied for preventing, controlling, and managing introduced and especially invasive species.
Resumen
Entre las principales causas que producen efectos perjudiciales sobre la biodiversidad mundial se encuentran la introducción de especies exóticas. Se pueden identificar muy pocos intentos de controlar anfibios y reptiles introducidos en América Central (México y América Central), por lo que proporcionamos listas de 24 especies exóticas, 16 especies translocadas y 11 especies que eliminamos de la lista de especies introducidas debido a la falta de evidencia que corrobore que provienen de poblaciones establecidas. También identificamos métodos de bioseguridad que se pueden aplicar para prevenir, controlar y gestionar especies introducidas y especialmente invasoras.
Keywords
Biological invasion, exotic species, herpetofauna, introduced species, Middle America, translocated species
Palabras claves
Especies exóticas, especies introducidas, especies translocadas, herpetofauna, invasion biológica, mesoamérica
Introduction
Among the most important drivers for biotic extinctions are introduced invasive species (
A common misbelief is that the ecology of invasive species and the ecosystem alterations they produce are extensively documented (
Materials and methods
We compiled a list of the introduced reptiles and amphibians in Middle America by examining relevant literature for the region, complemented with records obtained from GBIF (GBIF.org 2018), iNaturalist (inaturalist.org 2018), Amphibian Species of the World (
Our study area comprises Mexico and all Central American countries (Belize, Guatemala, Honduras, El Salvador, Nicaragua, Costa Rica, and Panama), ordered by latitude. Our use of the term “Middle America” refers to the Central American countries plus Mexico. We do not use the term “Mesoamerica,” since this label is generally considered more relevant in an anthropological and historical context (
In this paper, we consider introduced species to be populations whose presence in an area is attributed to human activities that enabled them to overcome biogeographical barriers that they otherwise could not cross and become established. We prefer introduced as the universal term over some others, such as exotic, non-native, alien, or non-indigenous, since the word “introduced” is more easily associated with human intervention (
Introduced species are not all-encompassing on temporal and geographical bases, because not all introduced organisms manage to become “established;” i.e., surviving long enough to produce descendant lineages (
The “impact” of an introduced invasive species, either exotic or translocated, refers to how an introduced species distresses the physical, chemical, or biological environment, the effect of which might be evident at the genetic, individual, population, community, ecosystem, landscape, regional, or global levels (
Our taxonomic positions follow those discussed in
Results and discussion
Status of the exotic and translocated herpetofauna of Middle America
Currently, 40 species of herpetofaunal species are considered introduced to a region of Middle America, or indigenous to a region, but translocated to non-native areas. Of these, 24 are exotics (Tables
List of introduced and translocated herpetofauna in the countries from Middle America. Introduced herpetofauna in Middle America (Right total = number of countries in which the species has “exotic” or “translocated” status; Bottom total = number of exotic and translocated species in that country). Parentheses enclose the number of exotic and translocated species for that taxon.
| Taxa | Exotic (E) or translocated (T) | Mexico | Guatemala | Belize | Honduras | El Salvador | Nicaragua | Costa Rica | Panamá | Total | Main references |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Amphibians (10) | |||||||||||
| Anurans (10) | |||||||||||
| Eleutherodactylidae (4) | |||||||||||
| Eleutherodactylus antillensis | E | + | 1 |
|
|||||||
| Eleutherodactylus coqui | E | + |
|
||||||||
| Eleutherodactylus johnstonei | E | + | 1 |
|
|||||||
| Eleutherodactylus planirostris | E | + | + | + | + | + | 5 |
|
|||
| Hylidae (2) | |||||||||||
| Osteopilus septentrionalis | E | + | 1 |
|
|||||||
| Smilisca baudinii | T | + | 1 |
|
|||||||
| Pipidae (1) | |||||||||||
| Xenopus laevis | E | + | 1 |
|
|||||||
| Ranidae (3) | |||||||||||
| Lithobates berlandieri | T | + | 1 |
|
|||||||
| Lithobates catesbeianus | T | + | 1 |
|
|||||||
| Lithobates forreri | T | + | 1 |
|
|||||||
| Reptiles (31) | |||||||||||
| Crocodylia (1) | |||||||||||
| Crocodylidae (1) | |||||||||||
| Crocodylus moreletii | T | + | 1 | Alvarez-Romero (2008) | |||||||
| Squamata (23) | |||||||||||
| Dactyloidae (4) | |||||||||||
| Anolis allisoni | E | + | + | + | 3 |
|
|||||
| Anolis carolinensis | E | + | 1 |
|
|||||||
| Ctenonotus cristatellus | E | + | 1 |
|
|||||||
| Norops sagrei | E | + | + | + | + | + | + | 6 |
|
||
| Gekkonidae (9) | |||||||||||
| Gehyra mutilata | E | + | 1 | Álvarez-Romero (2008) |
|||||||
| Gekko gecko | E | + | 1 |
|
|||||||
| Hemidactylus frenatus | E | + | + | + | + | + | + | + | + | 8 |
|
| Hemidactylus garnotii | E | + | + | 2 |
|
||||||
| Hemidactylus haitianus | E | + | + | 2 |
|
||||||
| Hemidactylus mabouia | E | + | + | + | + | 4 | Álvarez-Romero (2008), |
||||
| Hemidactylus turcicus | E | + | + | 2 |
|
||||||
| Lepidodactylus lugubris | E | + | + | + | + | 4 |
|
||||
| Tarentola mauritanica | E | + | Ortíz-Mena et al. (2019) | ||||||||
| Iguanidae (5) | |||||||||||
| Ctenosaura conspicuosa | T | + | 1 |
|
|||||||
| Ctenosaura pectinata | T | + | 1 |
|
|||||||
| Ctenosaura similis | T | + | 1 |
|
|||||||
| Sauromalus hispidus | T | + | 1 |
|
|||||||
| Sauromalus varius | T | + | 1 |
|
|||||||
| Leiocephalidae (1) | |||||||||||
| Leiocephalus varius | E | + | 1 |
|
|||||||
| Phrynosomatidae (1) | |||||||||||
| Uta stansburiana | T | + | 1 |
|
|||||||
| Sphaerodactylidae (1) | |||||||||||
| Sphaerodactylus argus | E | + | + | + | 3 |
|
|||||
| Boidae (1) | |||||||||||
| Boa imperator | T | + | 1 |
|
|||||||
| Typhlopidae (1) | |||||||||||
| Indotyphlops braminus | E | + | + | + | + | + | + | 6 |
|
||
| Testudines (7) | |||||||||||
| Chelydridae (1) | |||||||||||
| Chelydra serpentina | E | + | 1 |
|
|||||||
| Emydidae (1) | |||||||||||
| Trachemys scripta | T | + | 1 |
|
|||||||
| Kinosternidae (1) | |||||||||||
| Kinosternon integrum | T | + | 1 |
|
|||||||
| Staurotypidae (1) | |||||||||||
| Staurotypus triporcatus | T | + | 1 |
|
|||||||
| Testudinidae (1) | |||||||||||
| Chelonoidis carbonarius | E | + |
|
||||||||
| Trionychidae (1) | |||||||||||
| Apalone spinifera | T | + | 1 |
|
|||||||
| Totals | 24E/16T | 29 | 4 | 5 | 9 | 2 | 6 | 9 | 10 | – |
Herpetofaunal introductions in Middle America can be traced back to colonial times and were associated with commercial routes between the Philippines and New Spain (mainly Acapulco, Mexico), or through the slave trade from Western Africa to the Caribbean and Antillean islands, and from there into Central America. More recent events involved the opening of the Panama Canal in 1914, the expansion of the irrigation infrastructure after the 1950’s in northern Mexico, and throughout Middle America due to the highly popular pet trade and agricultural practices. On the other hand, translocations have more obscure origins, and certainly some of those could have occurred in pre-Columbian times, like translocations of iguanid lizards onto several islands in the Sea of Cortes (also called the Gulf of California, or Mar de Cortés in Spanish) by the Seri society. Whereas it is often accepted in invasive species biology that 1492 is the cutoff date for delineation between native and non-native species, we are concerned at this point with translocated species, not non-native species. In addition, it is our opinion that the year 1492, as the time when Cristopher Columbus “discovered” the New World, is of disputable significance from a biological point of view. The matter of most significant concern, we think, is to what extent humans, whether from Spain or elsewhere, have had a hand in the movement of creatures around the world.
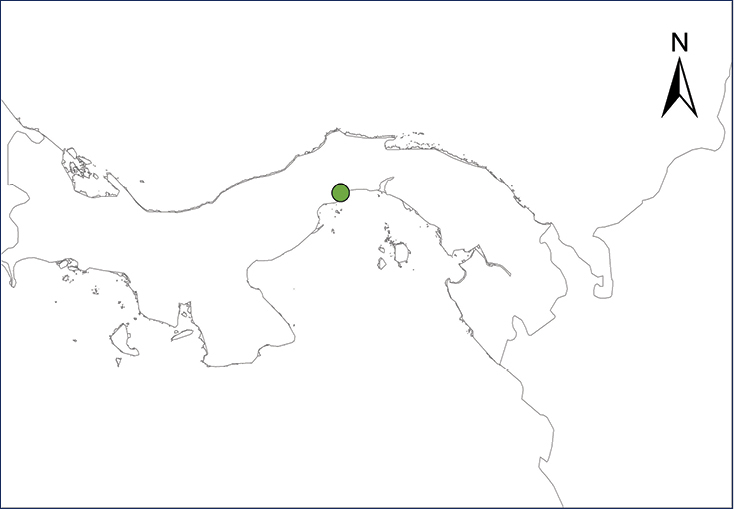
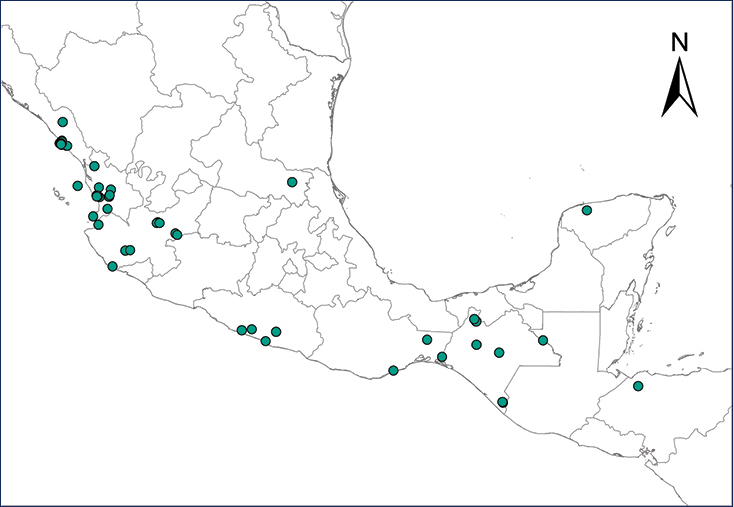
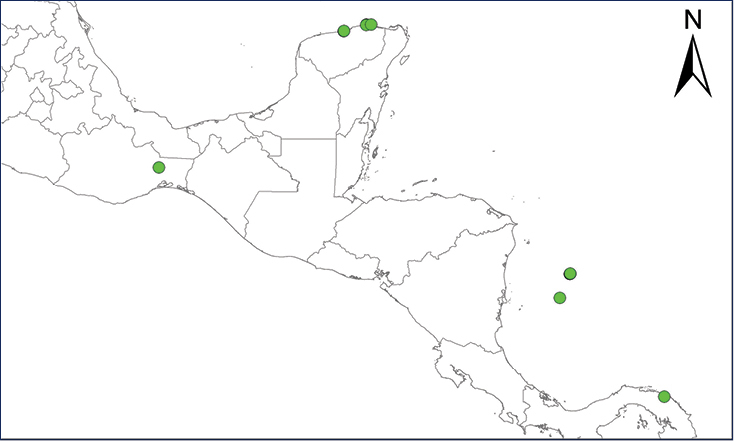
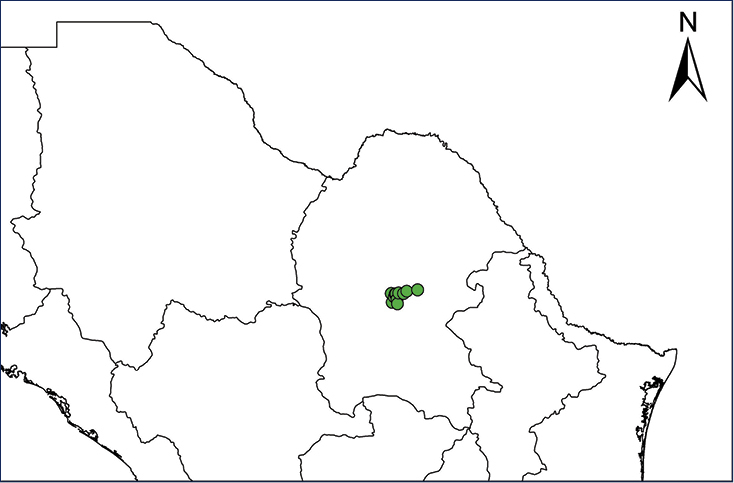
We recognize six major Middle American sites as “hotspots” for herpetofaunal invasions (four of which are depicted on Fig.

The four main Middle American sites considered as “hotspots” for herpetofaunal invasions: Northwestern Baja California and nearby Río Colorado delta in the Mexicali Valley, the Panama Canal, the Mexican Yucatan Peninsula, and The Papaloapan and Panuco basins in the Mexican state of Veracruz. The airports and seaports are not depicted due their ubiquity.
Finally, we recognize the following four major causes of introductions: 1) accidentals, mainly small species transported inadvertently by cargo vehicles, most frequently gekkonid lizards and anurans; 2) intentional releases, principally associated with pet trade and as food resources; most significantly chelonians and iguanid lizards, respectively; 3) escapees from the farming industry; mainly crocodiles and anurans, such as Morelet’s Crocodiles and American Bullfrogs; and 4) expanding invasion fronts when introduced naturalized species with high reproduction potential are well adapted to altered habitats. This fourth mechanism is especially relevant in anurans, such as those within the genus Eleutherodactylus and other species of original dispersers along invasion fronts.
The exotic herpetofauna of Middle America
Amphibia – Anura – Frogs
Family Eleutherodactylidae
Eleutherodactylus antillensis (Reinhardt & Lütken, 1863)
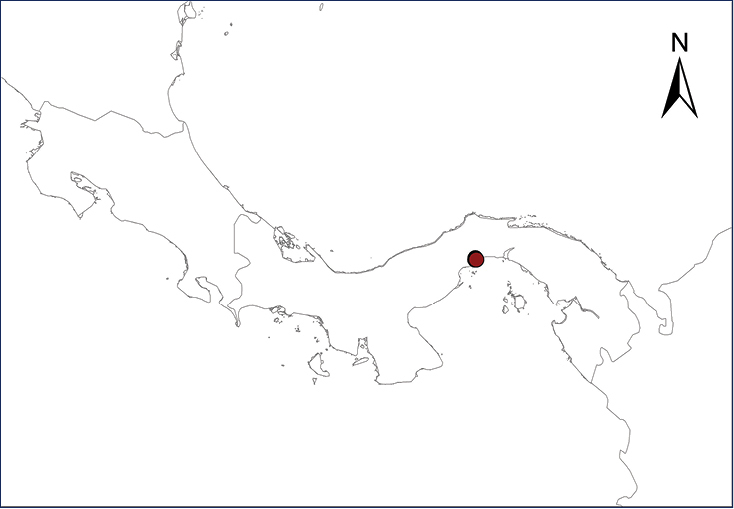
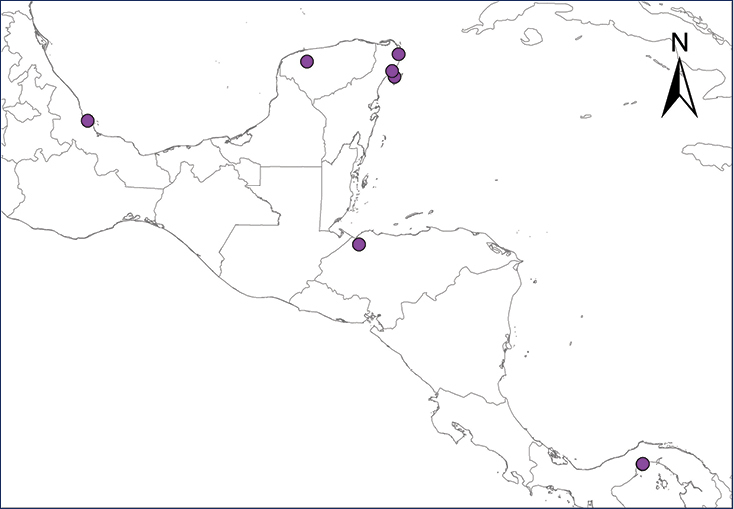
The Antilles Robber Frog is native to Puerto Rico, the Virgin Islands, and several associated islands and cays in that region. Numerous individuals apparently were introduced in the late 1950’s or early 1960’s into Panama City, probably through ornamental plants or intentionally released by a family after returning from a trip to Puerto Rico (
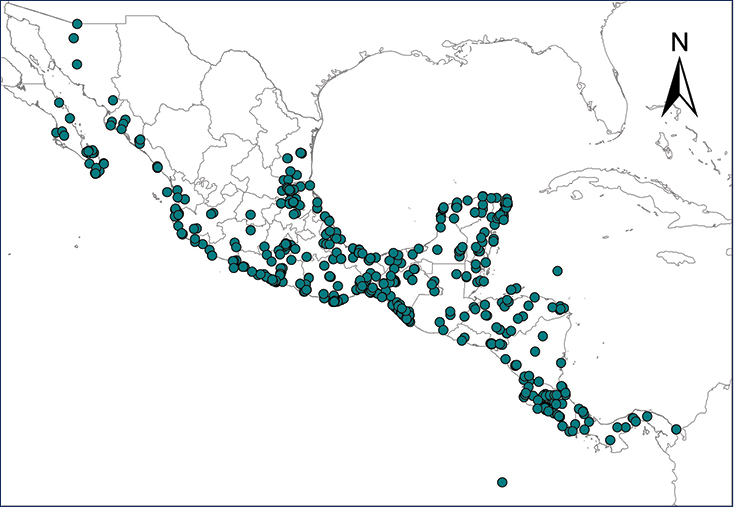
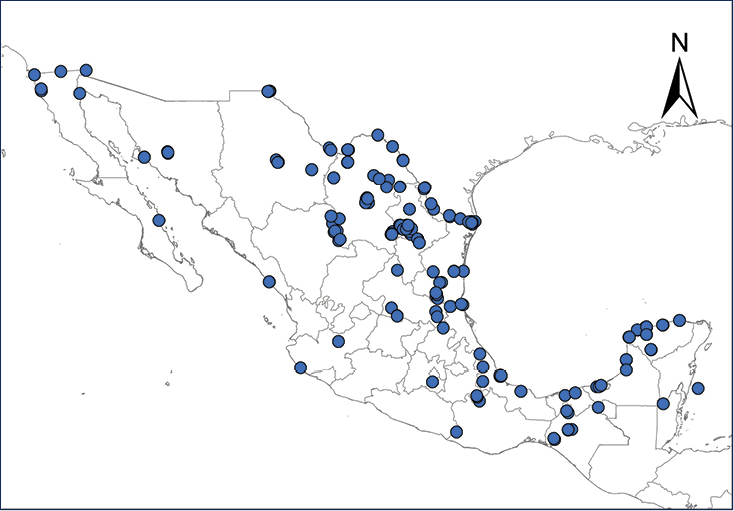
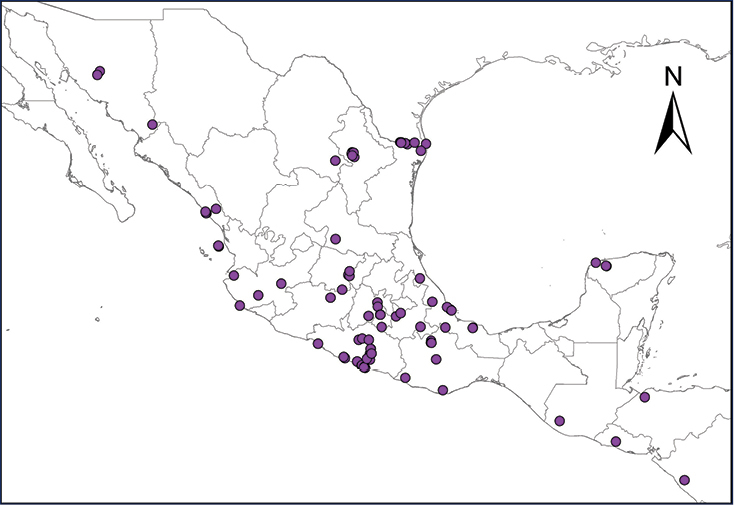
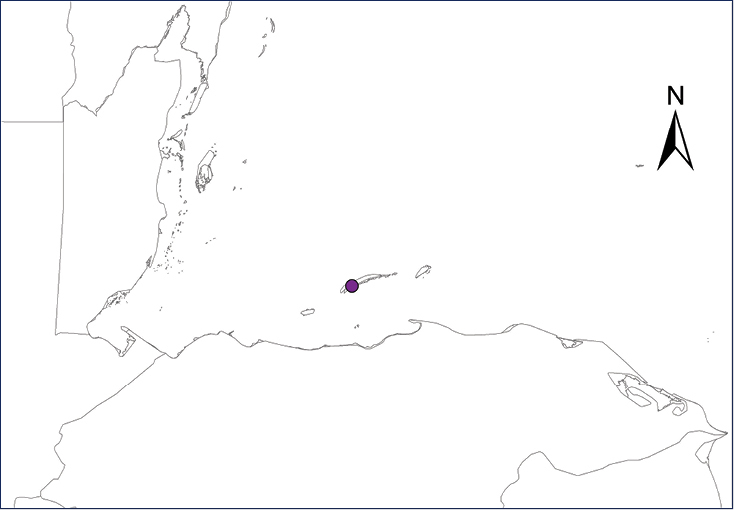
Distribution of introduced amphibians and reptiles in Mexico and Central American countries.
|
|
| Map 1. Eleutherodactylus antillensis. | Map 2. Eleutherodactylus coqui. |
|
|
| Map 3. Eleutherodactylus johnstonei. | Map 4. Eleutherodactylus planirostris. |
|
|
| Map 5. Osteopilus septentrionalis. | Map 6. Xenopus laevis. |
|
|
| Map 7. Chelonoidis carbonarius. | Map 8. Anolis allisoni. |
|
|
| Map 9. Anolis carolinensis. | Map 10. Ctenonotus cristatellus. |
|
|
| Map 11. Norops sagrei. | Map 12. Gekko gecko. |
|
|
| Map 13. Gehyra mutilata. | Map 14. Hemidactylus frenatus. |
|
|
| Map 15. Hemidactylus mabouia. | Map 16. Hemidactylus garnotii. |
|
|
| Map 17. Hemidactylus turcicus | Map 18. Lepidodactylus lugubris |
|
|
| Map 19. Tarentola mauritanica. | Map 20. Leiocephalus varius. |
|
|
| Map 21. Sphaerodactylus argus. | Map 22. Indotyphlops braminus. |
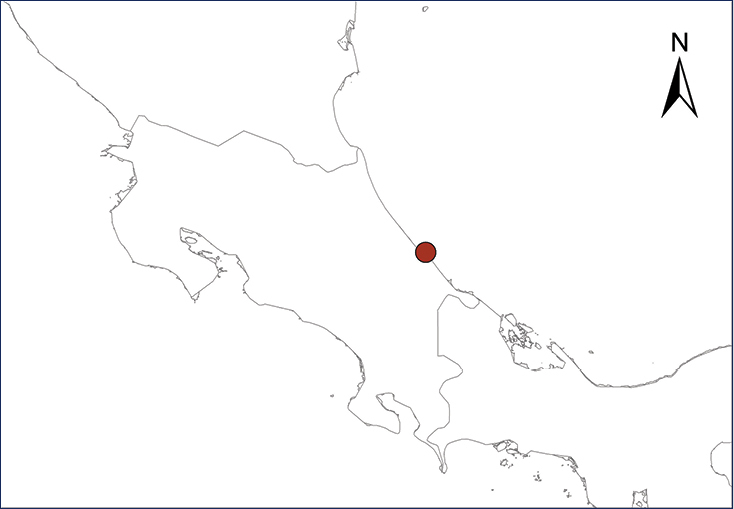
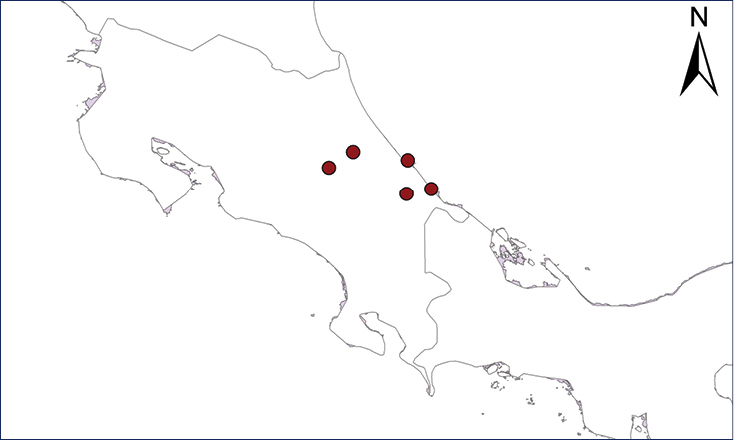
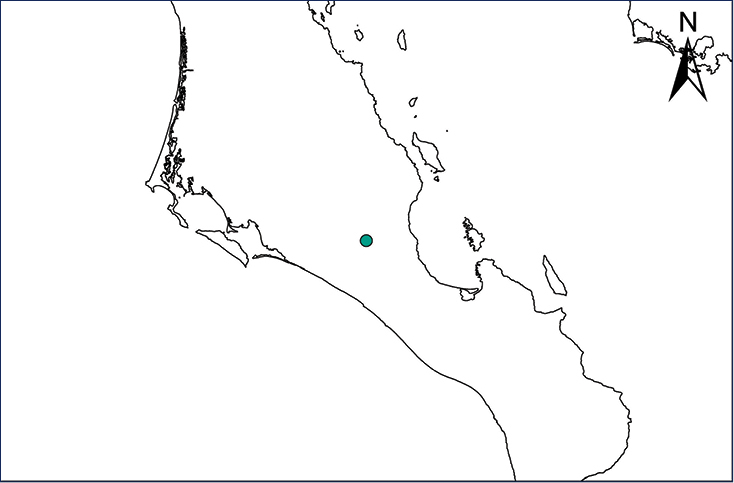
Eleutherodactylus coqui (Thomas, 1966)
Fig.
The Puerto Rican Coqui was first reported in Costa Rica by

Eleutherodactylus johnstonei (Barbour, 1914)
Johnstone’s Whistling Frog is native to the Lesser Antilles and has been introduced into several Caribbean areas, South America, the United States, and Europe (
Eleutherodactylus planirostris (Cope, 1862)
Fig.
The Greenhouse Frog is extremely small-sized (adults < 30 mm in length), is native to Cuba, Bahamas, and Cayman Islands (

Family Hylidae
Osteopilus septentrion,alis (Duméril & Bibron, 1841)
The Cuban Tree Frog is native to Cuba, Bahamas, and the Cayman Islands (
Family Pipidae
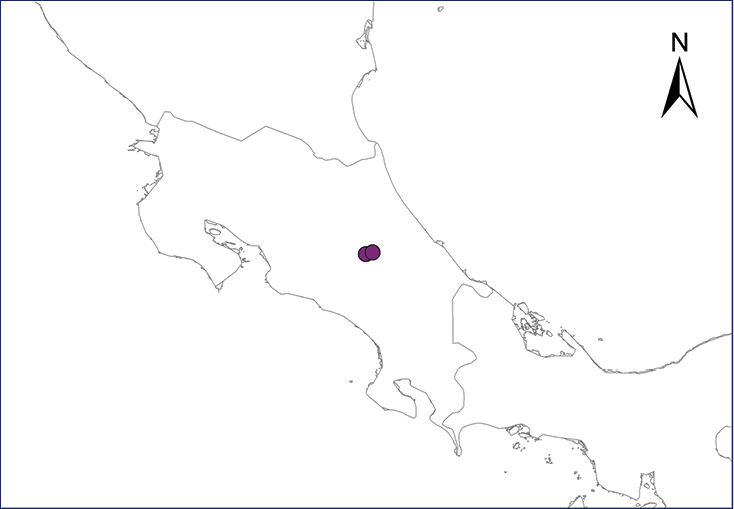
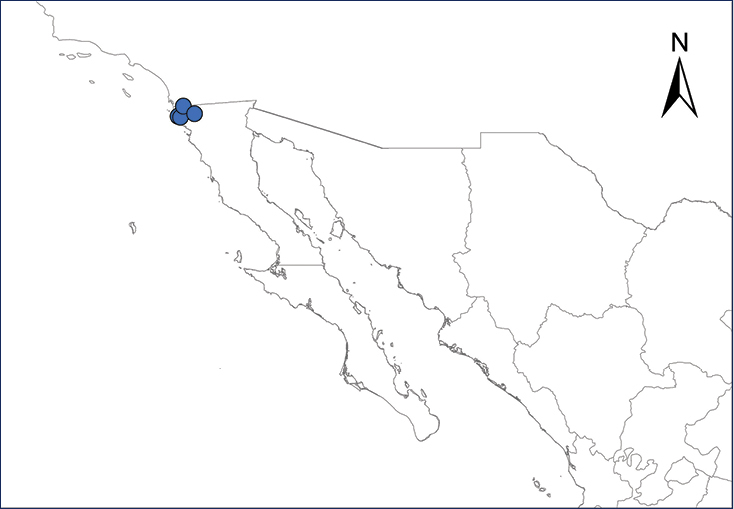
Xenopus laevis (Daudin, 1802)
Fig.
The African Clawed Frog is native to “extreme southern Angola…south to Cape Region of Rep. South Africa thence east and north in savanna habitats through Zimbabwe and southeastern Zambia to Malawi” (

Reptiles – Testudines – Turtles
Family Chelydridae
Chelydra serpentina (Linnaeus, 1758)
The Eastern Snapping Turtle is mentioned in the “Lista de las Especies Exóticas Invasoras para México” as present in the northern Mexican states of Coahuila and Chihuahua (
Family Testudinidae
Chelonoidis carbonarius (Spix, 1824)
The Red-footed Tortoise´s original distribution range extends from central Panama, through Colombia and the Atlantic versant of the Amazonas in Brazil, as far south as Paraguay and northern Argentina (
Reptiles – Squamata – Lizards
Family Dactyloidae
The anole family Dactyloidae contains the second highest number of introduced species (four species) in Middle America, and has exotic members distributed in all countries except El Salvador and Nicaragua. Norops sagrei is the most widespread anole in Middle America.
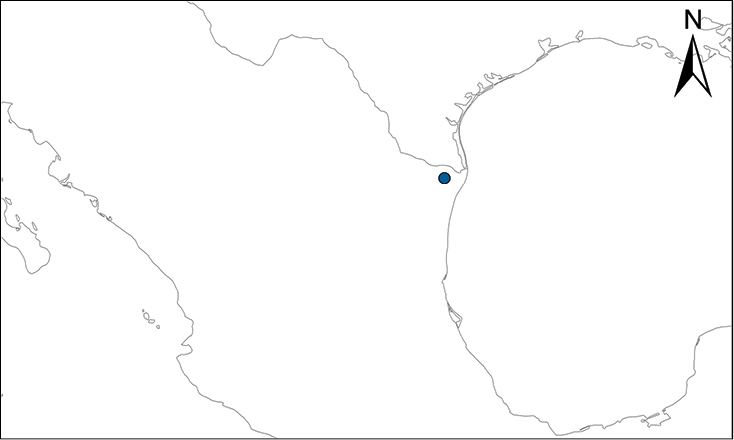
Anolis allisoni (Barbour, 1928)
Fig.
Allison’s Anole is listed by

Recent evidence (
Anolis carolinensis (Voigt, 1832)
Presence of the Green Anole in Mexico was suggested by
In Belize, the Green Anole is only known from a single specimen collected in 1966 on Half-Moon Cay (UF 23924); no other records were reported thereafter (
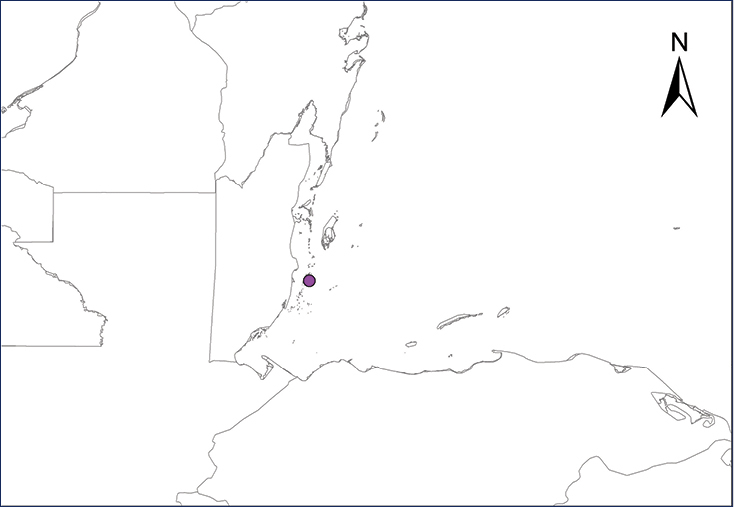
Ctenonotus cristatellus (Duméril & Bibron, 1837)
Fig.
The Crested Anole is native to Puerto Rico and the Virgin Islands (

In Mexico, C. cristatellus was reported from states on the Yucatan Peninsula, first by a single record from Cozumel, Quintana Roo, although
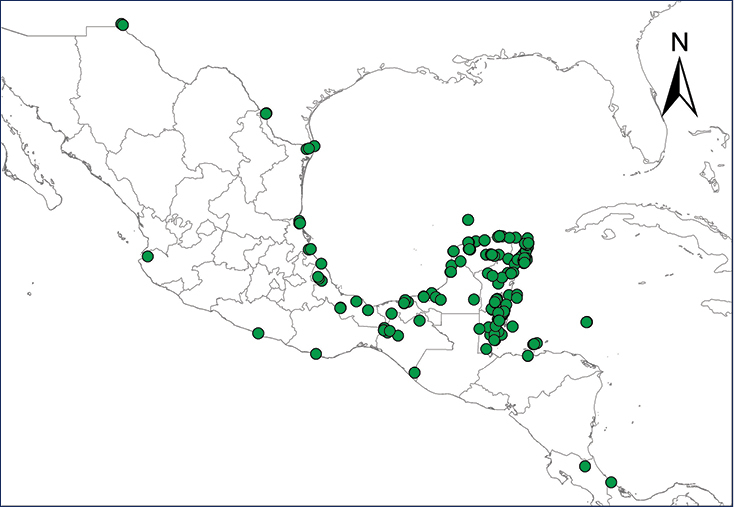
Norops sagrei (Duméril & Bibron, 1837)
The Cuban Brown Anole is native to Cuba, the Bahamas, and Cayman Islands, but it is unclear if all populations in Jamaica were introduced, or if some were native (
The naturalized distributional range in Mexico of N. sagrei comprises all the inland regions and several insular systems off the Yucatan Peninsula (
In Central America, the Cuban Brown Anole occurs throughout Belize (
Family Gekkonidae
Not surprisingly, the geckos contain the highest number of introduced species (10 species) for the region, distributed among two families (i.e., Gekkonidae and Sphaerodactylidae) in Middle America (Table
Hemidactylus is the most speciose genus, comprising five introduced species (H. frenatus, H. garnotii, H. haitianus, H. mabouia, and H. turcicus). At least one of these species occurs in each country of Middle America, but only H. frenatus is present in all seven (Table
Gekko gecko (Linnaeus, 1758)
Fig.
The Tokay Gecko is very well known due to its use in scientific research (

Gehyra mutilata (Wiegmann, 1834)
The Stump-toed Gecko is native to the Pacific basin region of Southeast Asia, where it has dispersed among Indian and Oceanic islands since the times of pre-Polynesian navigators (
The date for the introduction of G. mutilata into Mexico is unknown, although
Hemidactylus frenatus (Duméril & Bibron, 1836)
Fig.
The Common House Gecko is a well-known successful colonizer of urban environments (

It is believed that H. frenatus arrived on the North American continent around the 16th century by means of maritime commerce between the Philippines and Acapulco, Mexico (
Hemidactylus garnotii (Duméril & Bibron, 1836)
The Indo-Pacific or Garnot’s House Gecko, is native to the Indo-Pacific basin, and is widespread on several islands in Oceania, Asia, and the Pacific Ocean. In Middle America, it was introduced at several ports and urban centers in Costa Rica (
Hemidactylus haitianus (Meerwarth, 1901)
The Haitian House Gecko has had a complex and unclear taxonomic history. Traditionally, it was considered two different taxa: the Middle American and West Indian populations of H. brooki haitianus, type locality marked as “Haití, Port-au-Prince” (
Hemidactylus mabouia (Moreau De Jonnès, 1818)
Moreau’s Tropical House Gecko has no clear-cut geographic origin. The type locality is marked as “Antilles,” and restricted to St. Vincent Island by
Hemidactylus turcicus (Linnaeus, 1758)
The Mediterranean House Gecko is native to coastal areas of the Mediterranean, where it is widespread across southern Europe, the Levant, and more sporadically in North Africa (
Lepidodactylus lugubris (Duméril & Bibron, 1836)
Fig.
The Mourning Gecko is native to southeast Asian and Indo-Australian regions, and currently is distributed worldwide in the tropics from sea level up to 700 m elevation (

Tarentola mauritanica (Linnaeus, 1758)
Fig.
The Moorish Gecko is native to the European and North African Mediterranean basin (

Family Leiocephalidae
Leiocephalus varius (Garman, 1887)
The Cayman Curly-tailed Lizard is native to the Grand Cayman Islands (
Family Sphaerodactylidae
Sphaerodactylus argus (Gosse, 1850)
The Ocellated Dwarf Gecko is native to Jamaica, Cuba, and adjacent islands in the Bahamas and Antilles, including the Colombian Isla de San Andrés (
Records of S. argus from the northern coast of Yucatan (
Reptiles – Squamata – Snakes
Family Typhlopidae
Indotyphlops braminus (Daudin, 1803)
Fig.
The Brahminy Blindsnake is the most widespread alien reptile in the world (

The type locality is reportedly the region of Coromandel, southeast India (
The translocated herpetofauna of Middle America
Several species of amphibians and reptiles found in a region are translocated when introduced into areas of the same region outside their native ranges. We will consider only those with sound evidence of having been translocated by human activities and that have established populations (Table
Amphibia – Anura – Frogs
Family Hylidae
Smilisca baudinii (Duméril & Bibron, 1841)
The Mexican Treefrog ranges from “Extreme southern Texas (USA) and southern Sonora and southwestern Chihuahua (Mexico) south (including the Balsas Depression of Mexico) in tropical lowlands to Costa Rica on the Pacific slope; including the Tres Marias Islands off the coast of Nayarit, Mexico” (Frost, 2020). This species also can be found on other Mexican Pacific Islands (
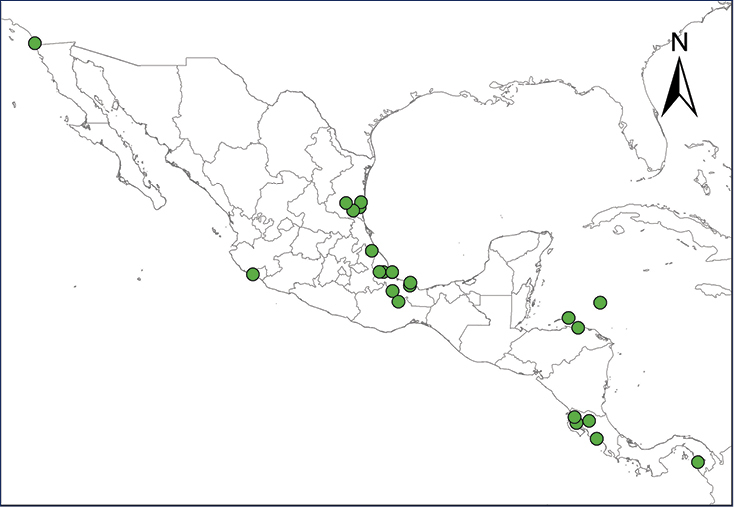
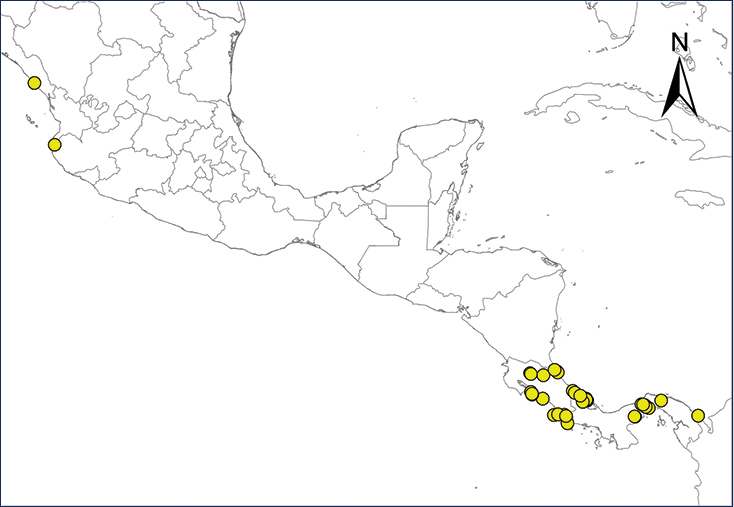
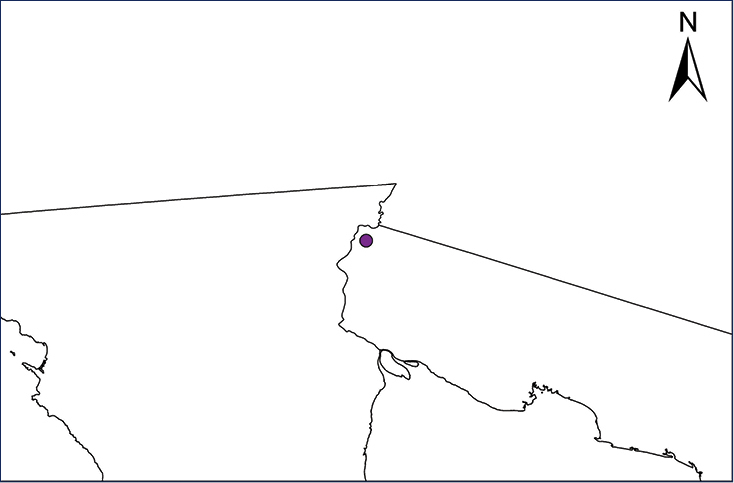
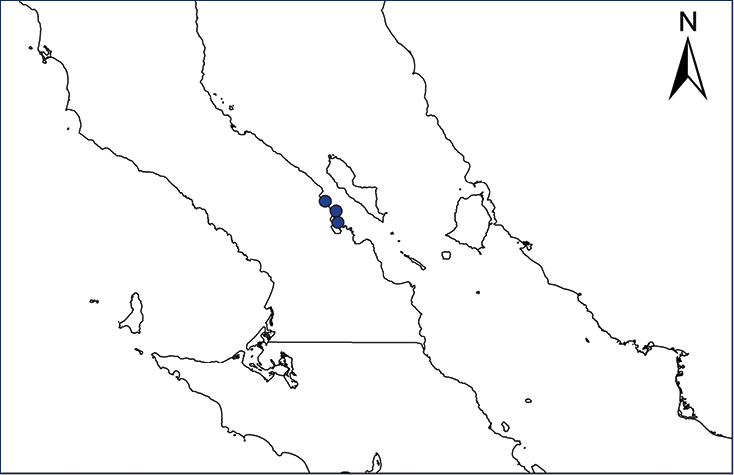
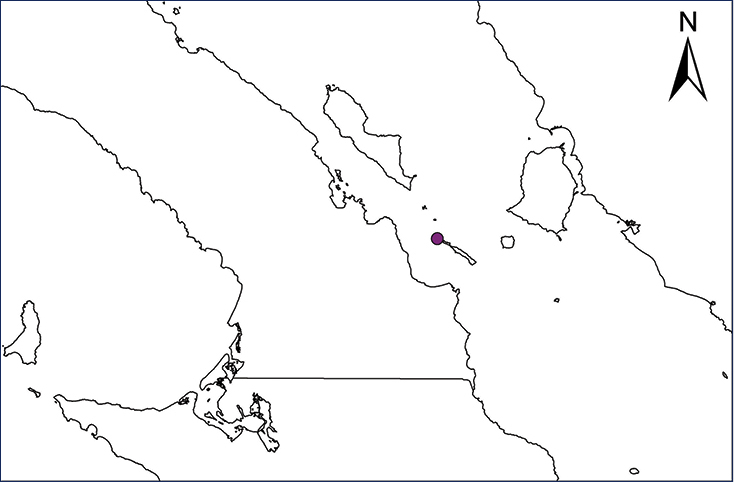
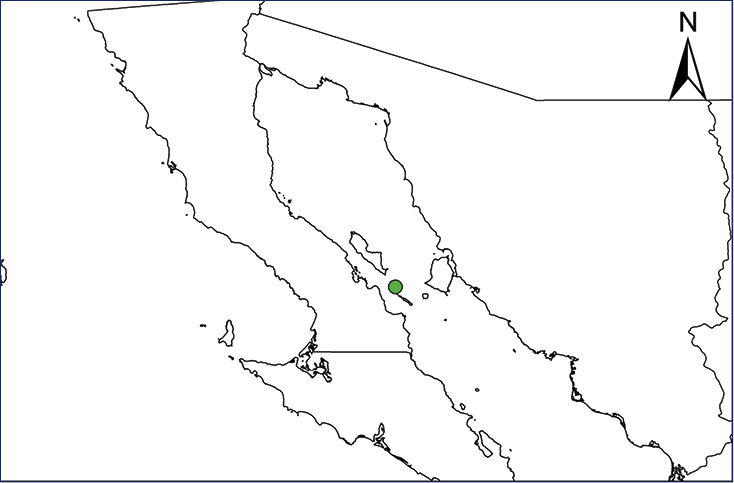
Non-native distribution of translocated amphibians and reptiles in Mexico and Central America.
|
|
| Map 23. Smilisca baudinii | Map 24. Lithobates berlandieri |
|
|
| Map 25. Lithobates forreri | Map 26. Apalone spinifera |
|
|
| Map 27. Ctenosaura conspicuosa | Map 28. Ctenosaura pectinata |
|
|
| Map 29. Ctenosaura similis | Map 30. Sauromalus hispidus |
|
|
| Map 31. Sauromalus varius | Map 32. Uta stansburiana |
|
|
| Map 33. Boa imperator | |
Family Ranidae
Lithobates berlandieri (Baird, 1859)
The native range of the Rio Grande Leopard Frog extends from southeastern New Mexico and central Texas southward into Mexico (
Lithobates catesbeianus (Shaw, 1802)
Fig.
The American Bullfrog originally ranged from southeastern Canada and central and eastern United States into northeastern Mexico (

Due to a high reproductive rate and generally destructive behaviors, L. catesbeianus has great potential for being harmful to native species, and has already been associated with declining and disappearing populations of native amphibians around the world (
Lithobates forreri (Boulenger, 1883)
Forrer’s Leopard Frog’s native distribution was considered to be on the mainland Pacific versant of Mexico from Sonora (
Reptiles – Crocodylia – Crocodiles
Family Crocodylidae
Crocodylus moreletii (Duméril & Bibron, 1851)
Fig.
Morelet’s Crocodile originally ranged only along the Atlantic lowlands of Middle America, from Tamaulipas, Mexico, to northern Guatemala and adjacent Belize (

The invasion of aquatic habitats by C. moreletii might have serious consequences for native biotic communities, since it is a large top predator (
Reptiles – Testudines – Turtles
Family Emydidae
Trachemys scripta (Thunberg in Schoepff, 1792)
Fig.
A common misbelief in Mexico is that the Pond Slider is native to Japan, thus the common name “Tortugas japonesas” (Japanese turtles) is used frequently. The species’ native geographic range, as presently understood, includes the southeastern United States and adjacent lowlands of northeastern Mexico (
Any review of literature covering the distribution of this introduced turtle should be made with special care, since the name Trachemys scripta, until recently, included almost every Pond Turtle population ranging throughout Middle America, unfortunately, listed as subspecies of T. scripta (
Our review of introduced species will only cover what has been called the Red-eared Slider, T. scripta elegans (Wied, 1838), which is listed among the 100 most dangerous invasive species, according to the Invasive Species Specialist Group (
Established introduced populations of the Elegans pattern class of T. scripta are disconnectedly distributed in several parts of Mexico, such as in Baja California, within several northern states, along the southern Pacific slopes, and on the Yucatan Peninsula (
In Honduras, T. scripta has been observed in Río Llanitos, Santa Bárbara, and Isla Guanaja in the Islas de la Bahía (= Bay Islands) (

Trachemys venusta (Gray, 1855)
The Mesoamerican Slider, as discussed elsewhere, was involved in the taxonomic dispute associated with T. ornata (
Family Kinosternidae
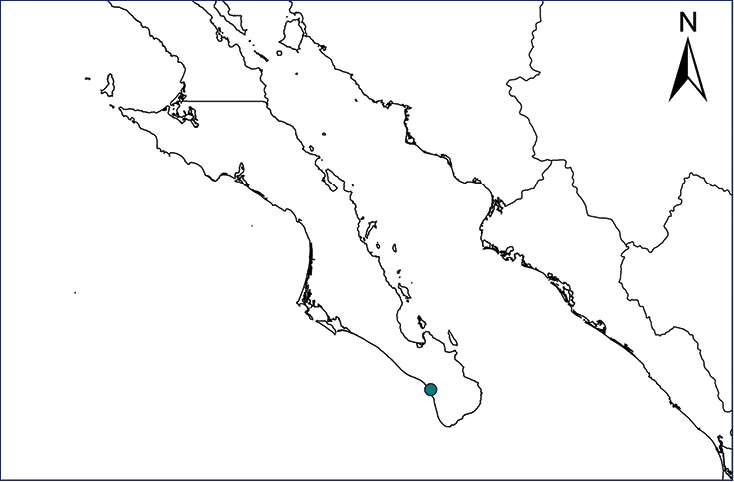
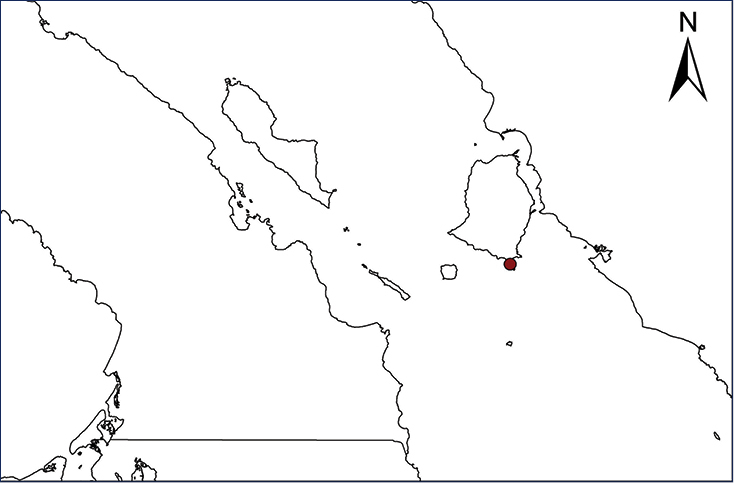
Kinosternon integrum (Le Conte, 1854)
A single individual of the Mexican Mud Turtle was captured 29 March 2007 in a perennial pool at the bottom of a rocky canyon in the Sierra La Laguna, Baja California Sur (
Family Trionychidae
Apalone spinifera (Le Sueur, 1827)
The Spiny Softshell has a limited native range in Mexico, principally in drainage systems associated with the Rio Grande, which is the border with Texas and a very small segment of New Mexico near El Paso, with neighboring states in Mexico (Chihuahua, Coahuila, Nuevo León, and Tamaulipas), then continuing along the Gulf lowlands to approximately Soto la Marina, Tamaulipas (
In the Cuatro Ciénegas area of Coahuila, Mexico, A. spinifera arrived at some local water sources when irrigation channels were opened from the northeast in the 1880’s (
Outside of northern Mexico, a single record exists for A. spinifera from Jalisco in an artificial pond in Puerto Vallarta (
There are, however, documented localities in Guerrero for A. spinifera from along the Río Balsas drainage, one from the vicinity of Colonia Valerio Trujano, near the Mezcala Bridge, Municipality of Edwardo Neri, that seems to be from an established population. Local residents indicated that it had been present there since the 1950’s (
Reptiles – Squamata – Lizards
Family Iguanidae
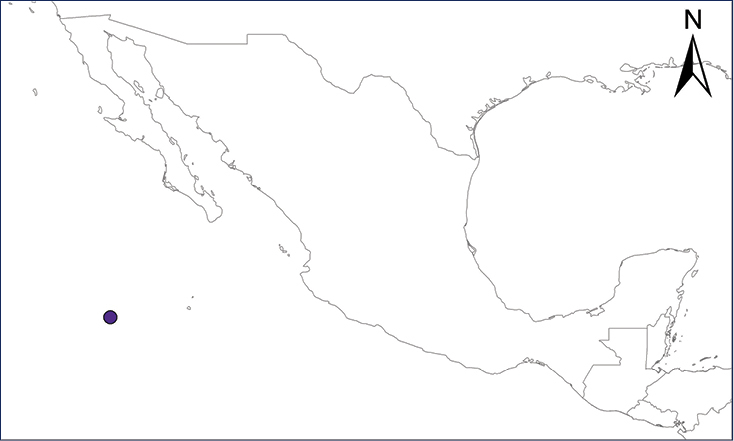
Ctenosaura conspicuosa (Dickerson, 1919)
The Isla San Esteban Spiny-tailed Iguana only exists on Cholludo and San Esteban islands, in the Sea of Cortes, located in close proximity to the coast of Sonora, Mexico. The cultural evidence suggests that C. conspicuosa populations on both islands could be due to a prehistoric introduction of C. nolascensis from Isla San Pedro Nolasco by the Seri culture (
Ctenosaura pectinata (Wiegmann, 1834)
The Western Spiny-tailed Iguana naturally occurs in low to intermediate elevations primarily on the Pacific versant of Mexico from Sinaloa into Chiapas, including subhumid interior basins and valleys and offshore islands (Uetz et al. 2020). This iguana was introduced on the remote Isla Clarion sometime in the mid-1990´s (
Ctenosaura similis (Gray, 1831)
Fig.
The Black Iguana naturally occurs on the Atlantic and Pacific versants from Mexico, below the Isthmus of Tehuantepec, through all countries in Central America (

Sauromalus hispidus (Stejneger, 1891)
The Spiny Chuckwalla inhabits several islands within the Gulf of California, including: Angel de La Guarda, Alcatraz, Cabeza de Caballo, Flecha, Granito, Mejía, Piojo, Pond, San Lorenzo Norte, San Lorenzo Sur, and numerous islands in Bahía de Los Ángeles (
Sauromalus varius (Dickerson, 1919)
The Piebald Chuckwalla is only known from the islands of San Esteban and Roca Lobos in the Sea of Cortes (
Family Phrynosomatidae
Uta stansburiana (Baird & Girard, 1852)
The Side-blotched Lizard is a common widespread generalist, occurring in the western United States, northern Mexico, and along the Baja California Peninsula and many of its associated islands (
Reptiles – Squamata – Snakes
Family Boidae
Boa imperator (Daudin, 1803)
The Central American Boa Constrictor, formerly a subspecies of Boa constrictor (
Reported introduced species not on our list of established populations in Middle America
Taricha torosa (Rathke, 1833)
Gopherus agassizii (Cooper, 1861)
The taxonomic history of the G. agassizii species complex of Desert Tortoises (Testudinidae) generally had been unresolved (
At this point, we do not think that the population around La Paz has been identified definitively as being G. morafkai, G. evgoodei, G. agassizii, or one of hybrid origin. We also consider any inhabitants that will be found in northeastern Baja California will probably fall within the native range of G. morafkai, unless they can be shown positively to be part of translocated G. agassizii, G. evgoodei, or hybrid populations. Thus, it will not be appropriate at this time to list any population of the G. agassizii species group of Desert Tortoises as being introductions within northwestern Mexico.
Gopherus berlandieri (Agassiz, 1857)
A single specimen of the Texas Tortoise (Testudinidae) was found in a city park at Puerto Vallarta, Jalisco, Mexico (
Staurotypus triporcatus (Wiegmann, 1828)
Until recently, the Mexican Giant Musk Turtle (Staurotypidae) was considered native to the Atlantic lowlands from central Veracruz, Mexico, through the southern Yucatan Peninsula, and on to the western Caribbean lowlands of Honduras (
Trachemys ornata (Gray, 1831)
The Trachemys scripta species group (Emydidae) has had a confusing taxonomic history in Middle America, especially those populations occurring in tropical latitudes (
Cnemidophorus ruatanus (Barbour, 1928)
The Ruatan Whiptail (Teiidae) was reported (as Cnemidophorus lemniscatus) by
Gonatodes albogularis (Duméril & Bibron, 1836)
Fig.
Identifying the native distribution of the Yellow-headed Gecko (Sphaerodactylidae) is problematical because of its wide range in parts of Middle America, northern South America, and on many islands in the West Indies (Uetz et al. 2020). A pertinent question is whether its native range is restricted to Middle and South America and it is introduced in the West Indies, or vice versa.

On the mainland, G. albogularis occurs from the Pacific slopes of the Soconusco and Sierra Madre regions in Chiapas, Mexico, through Central America into northern Colombia and Venezuela (
Phyllodactylus nocticolus (Dixon, 1964)
The occurrence of Peninsular Leaf-toed Gecko (Phyllodactylidae) on Isla Tiburón could be due to natural over-water island hopping, accidental introduction (
Phyllodactylus xanti (Cope, 1863)
Sauromalus obesus (Baird, 1859)
A chance exists that the Western Chuckwalla (Iguanidae) might have been translocated to several islands in the Sea of Cortes, based on the same reasons as those for S. hispidus and S. varius (
Varanus exanthematicus (Bosc, 1792)
The Savanna Monitor (Varanidae) is known in Mexico from a single record in Puerto Vallarta, Jalisco (
Considerations for management and control measures for introduced species
The main difficulty for management and control of the introduced invasive herpetofauna of Middle America is the lack of documentation. Most conservation managers or agencies have little to no tradition of publishing in the refereed scientific literature, so, for the most part, successes and failures of invasive species are found in grey literature sources (
A key component of any successful control or eradication program is early detection of an invasion and a quick response (
Another key component of control and eradication programs involves the correct identification of the presumed introduced species by qualified specialists and their training of non-expert volunteers so to avoid misidentification of native species as introduced ones. These specialists also need to participate in the removal programs. These lessons have come from the work of Rick Shine and his colleagues on the Cane Toad or Marine Toad (Rhinella marina) in Australia, as reported in numerous publications, including the book Cane Toad Wars (
A common challenging situation emerges in cases where some translocated species are under legal protection, but are exotic or even invasive in other parts of the country. This situation keeps conservation managers from performing effective eradication or control measures on those invasive populations (
The first obvious method for controlling introduced species of amphibians and reptiles is direct capture/sacrifice and trapping methodology. Some biologists, however, are opposed to the sacrificing such introduced creatures, for ethical reasons. This position is entirely understandable and is complex enough to require adequate discussion elsewhere. Several capture techniques include using nooses, pitfall traps, funnel traps, sticky traps, rubber bands, firearms, blowguns, and road cruising, among others, whose effectiveness are well known to herpetologists, thus we will not detail them here. Less common, but potentially successful procedures are discussed below.
Chemical control has a long tradition for managing invasive mammals, but its use has been employed infrequently for herpetofaunal control. The most publicized instances involved Brown Tree Snakes (Boiga irregularis) on Guam, where
Invasive anuran control programs in Australia included spraying lethal chemicals in water sources (
A key aspect for the success of some invasive anurans are potent chemical defenses that most vertebrate predators cannot tolerate. Nonetheless, several invertebrate groups are immune to those toxins, and many of them, such as dragonfly nymphs, fishing spiders, water beetles, ants, crabs, and crayfish, voraciously consume tadpoles or early stage metamorphs. Thus, introducing native invertebrate predators can be a biological control option when the only species in the water source is an alien anuran (
A major advantage for invasive species when reaching a new area is access to a parasite-free space (
A successful molecular tool for early detection and monitoring aquatic and semiaquatic invader species is testing water sources for their waterborne environmental DNA (eDNA), as described by
A key feature for managing introduced species, especially in large areas, is to identify important sites to focus control and eradication efforts, as well as to prevent invasions and/or reinvasions before they occur. Environmental niche modelling (ENM) has been a helpful tool for identifying potential corridors among the sources and areas vulnerable to invasions (
Decision makers often require methods that help them justify and decide where, when, and on which species to target conservation and/or control programs. For these reasons, there exist protocols that can be used to determine if a species is potentially at risk and deserves attention. Generally, these protocols consist of a questionnaire that must be answered by a specialist or by a panel of experts; examples are revealed in “Método de Evaluación Rápida de Invasividad (MERI) para Especies Exóticas en México” (
Finally, and most important of all, is the human component, which is pivotal for the success of any management program governing actions associated with invasive species (
Conclusions
As noted previously, interest in introduced invasive species is taxonomically skewed toward other vertebrates, such as mammals, birds, or fishes. Thus, with a few exceptions, the ecological influences and damages caused by introduced invasive amphibians and reptiles are unknown. This lack of knowledge can have ominous consequences, such as taxonomic uncertainty, causing voids in legislation, and omissions of reptiles and amphibians in many biosecurity protocols or practices. Additionally, a great proportion of the literature on monitoring or control programs corresponds to technical reports not easily accessible to other researchers and/or managers. A first barrier to overcome is to encourage managers and researchers to identify results of monitoring and control programs on the invasive herpetofauna and to have that information published in accredited journals.
For conservationists to influence protocols, it is imperative that they promote stricter legislation on damaging practices, such as the pet trade, which should be discouraged, especially in those species with high potential of being harmful to Middle American ecosystems, even if they are already present, like Trachemys spp., or those that are common in the pet trade of a region, but not yet reported in the wild, such as Varanus exanthematicus and Python molurus. Stricter legislation should not be limited to vertebrate species, but also extended to pathogens associated with amphibians (or reptiles), such Batrachochytrium dendrobatidis, Saprolegnia parasitica, and Ranavirus spp. We are aware that such listing of potential harmful pathogens exists for Mexico (
Finally, the ever-growing trade of goods on a global scale, the increasing interest by people for keeping exotic pets, and the human persistence for environmental degradation will continue to favor arrival and settlement of invasive species. Regrettably, the frequency, scope, and intensity of biological invasions are expected to increase during the ensuing decades. Thus, the study and management of introduced amphibians and reptiles in Middle America is a topic that offers a wide spectrum of opportunities for career development associated with young researchers, conservationists, and other professionals dealing in ecological restoration.
We consider biological conservation as a human value, which includes a series of moral codes and behaviors that transcend time and culture and define us as a species. In this way, and like other values, it contributes to building prosperity and free coexistence among societies. It is also obvious that invasive species control and management is controversial, since many people put a high value on any single living organism, independent of its origin. If they accept invasive species to be a legitimate part of our ecological footprint, however, it is clear that we have a moral and ethical responsibility to educate them on the negative impacts invasive species have on the overall well-being of our biosphere. At the same time, it is also our responsibility to help maintain biosecurity and ecological restoration measures as advocates to prevent, mitigate, and remediate damages caused by invasive species until the majority of humanity accepts the fact that being good stewards of our living spaces is the right thing to do. Conservation professionals also must understand what motivates people’s different attitudes towards invasive species (e.g., why they transport them, whether they perceive them as harmful or not, or whether they are willing to accept control methods or not) in order to develop meaningful programs that discourage harmful behaviors and promote more responsible attitudes. Therefore, integration of the human and technical component is fundamental for accepting biosecurity as one more principle guiding societal behavior.
Acknowledgements
We thank Lizbeth E. Lara-Sánchez for her review and comments on the control methods section. We are also thankful to Rubén Alonso Carbajal-Márquez, Luis Díaz-Gamboa, Julio Gutiérrez-Ramírez, Javier Ortiz-Medina, Louis W. Porras, Javier Sunyer, and Anny Peralta for allowing us to use their photographic material to illustrate this paper. We also are grateful to Vicente Mata-Silva for assistance with corrections to one of the distribution maps and to Louis W. Porras for supplying us with information concerning the references.
References
- Abarca J, Monge AM (2007) Geographic distribution: Costa Rica, San José: Hemidactylus mabouia. Herpetological Review 38(3): 351–352.
- Aguirre-Léon G, Matías-Ferrer N (2017) Islas Revillagigedo: sus reptiles y el redescubrimiento de una especie. La Jornada Ecológica 209: 16–18.
- Ahumada-Carrillo IT, Weatherman GN (2018) A confirmed established population of Lepidodactylus lugubris (Duméril & Bibron, 1836) (Squamata: Gekkonidae) in Mexico. Mesoamerican Herpetology 5(1): 179–180.
- Álvarez-Romero J, Medellín RA, Gómez de Silva H, Oliveras de Ita A (2005) Boa constrictor. Vertebrados Superiores Exóticos en México: Diversidad, Distribución y Efectos Potenciales. Instituto de Ecología, Universidad Nacional Autónoma de México. Bases de datos SNIB-CONABIO. Proyecto U020. México. D.F.
- Álvarez-Romero JG, Medellín RA, Oliveras de Ita A, Gómez de Silva H, Sánchez O (2008) Animales Exóticos en México: Una Amenaza para la Biodiversidad, México D.F. , Comisión Nacional para el Conocimiento y Uso de la Biodiversidad, Instituto de Ecología, UNAM, Secretaría del Medio Ambiente y Recursos Naturales, 502 pp.
- Arriaga A, Ramírez-Bautista A (2008) A Checklist of Reptiles from the Parque Nacional Isla Contoy, México. Boletín de la Sociedad Herpetológica Mexicana 16(2): 36–40.
- Auth DL (1994) Checklist and bibliography of the amphibians and reptiles of Panama. Smithsonian Herpetological Information Service 98: 1–59. https://doi.org/10.5479/si.23317515.98.1
- Avila-Villegas H, Rodríguez-Olmos LP, Lozano-Román LF (2007) Rana toro (Lithobates catesbeianus): anfibio introducido en Aguascalientes, México. Boletín de la Sociedad Herpetológica Mexicana 15(1): 16–17.
- Barker BS, Rodríguez-Robles JA (2017) Origins and genetic diversity of introduced populations of the Puerto Rican Red-Eyed Coqui, Eleutherodactylus antillensis, in Saint Croix (U.S. Virgin Islands) and Panamá. Copeia 105(2): 220–228. https://doi.org/10.1643/CG-16-501
- Barquero MD, Araya MF (2016) First record of the Greenhouse Frog, Eleutherodactylus planirostris (Anura: Eleutherodactylidae), in Costa Rica. Herpetology Notes (9): 145–147.
- Barrantes-Madrigal J (2017) El Coquí Común (Eleutherodactylus coqui) en Costa Rica: Selección de Hábitat, Etapa de Invasiòn y Aspectos Sociales Antrópicos que Podrían Influir en su Dispernsión y Manejo. M. Sc. Thesis. Universidad Nacional Costa Rica, 81 pp.
- Barrantes-Madrigal J, Parallada MS, Alvarado G, Chaves-Acosta VJ (2019) Distribution and invasion progress of Eleutherodactylus coqui (Anura: Eleutherodactylidae) introduced in Costa Rica. Phyllomedusa: Journal of Herpetology 18(1): 101–107. https://doi.org/10.11606/issn.2316-9079.v18i1p101-107
- Batista A, Ponce M, Garcés O, Lassiter E, Miranda M (2019) Silent pirates: Anolis sagrei Duméril & Bibron, 1837 (Squamata, Dactyloidae) taking over Panama City, Panama. Check List 15(3): 455–459. https://doi.org/10.15560/15.3.455
- Bauer AM, Jackman TR, Greenbaum E, Giri VB, de Silva A (2010) South Asia supports a major endemic radiation of Hemidactylus geckos. Molecular Phylogenetics and Evolution 57(1): 343–352. https://doi.org/10.1016/j.ympev.2010.06.014
- Beard KH, Price EA, Pitt WC (2009) Biology and Impacts of Pacific Island invasive species. 5. Eleutherodactylus coqui, the Coqui Frog (Anura: Leptodactylidae). Pacific Science 63(3): 297–316. https://doi.org/10.2984/049.063.0301
- Beard KH, Vogt KA, Kulmatiski A (2002) Top-down effects of a terrestrial frog on forest nutrient dynamics. Oecologia 133(4): 583–593. https://doi.org/10.1007/s00442-002-1071-9
- Blair C, Méndez de La Cruz F, Ngo A, Lindell J, Lathrop A (2009) Molecular phylogenetics and taxonomy of leaf-toed geckos (Phyllodactylidae: Phyllodactylus) inhabiting the peninsula of Baja California. Zootaxa 2027(1): 28–42. https://doi.org/10.11646/zootaxa.2027.1.2
- Böehm S (2013) Records of invasive Trachemys scripta elegans (Wied-Neuwied, 1839), in cenotes of the Yucatan Peninsula, Mexico. Herpetozoa 26: 98–100.
- Bohmann K, Evans A, Gilbert MTP, Carvalho GR, Creer S, Knapp M, Yu DW, de Bruyn M (2014) Environmental DNA for wildlife biology and biodiversity monitoring. Trends in Ecology & Evolution 29(6): 358–367. https://doi.org/10.1016/j.tree.2014.04.003
- Bonin F, Devaux B, Dupré A (2006) Turtles of the World. The Johns Hopkins University Press, Baltimore, 416 pp.
- Brattstrom BH (1955) Notes on the Herpetology of the Revillagigedo Islands, Mexico. The American Midland Naturalist 54(1): 219–229. https://doi.org/10.2307/2422189
- Brattstrom BH (1990) Biogeography of the Islas Revillagigedo, Mexico. Journal of Biogeography 17(2): 177–183. https://doi.org/10.2307/2845325
- Brooks JE, Savarie PJ, Johnston JJ (1998) The oral and dermal toxicity of selected chemicals to brown tree snakes (Boiga irregularis). Wildlife Research 25(4): 427–435. https://doi.org/10.1071/WR97035
- Buckley LJ, de Queiroz K, Grant TD, Hollingsworth BD, Iverson JB, Pasachnik SA, Stephen CL (2016) A checklist of the iguanas of the world (Iguanidae; Iguaninae). Herpetological Conservation and Biology 11(Monograph 6): 4–46.
- Bury RB, Germano DJ, Van Devender TR, Martin BE (2002) The desert tortoise in Mexico. Distribution, ecology, and conservation. In: Van Devender TR (Ed.) The Sonoran Desert Tortoise. Natural History, Biology and Conservation. University of Arizona Press, Tucson, 86–108. https://doi.org/10.2307/j.ctvfjcx1x.8
- Cabrera-Guzmán E, Crossland MR, Shine R (2012) Predation on the eggs and larvae of invasive cane toads (Rhinella marina) by native aquatic invertebrates in tropical Australia. Biological Conservation 153: 1–9. https://doi.org/10.1016/j.biocon.2012.04.012
- Cabrera-Guzmán E, Crossland MR, Shine R (2015) Invasive Cane Toads as Prey for Native Arthropod Predators in Tropical Australia. Herpetological Monographs 29(1): 28–39. https://doi.org/10.1655/HERPMONOGRAPHS-D-13-00007
- Cadotte MW (2006) Darwin to Elton: early ecology and the problem of invasive species. In: Cadotte MW, McMahon SM, Fukami T (Eds) Conceptual Ecology and Invasion Biology: Reciprocal Approaches to Nature (Vol. 1). Springer, New York, 15–33. https://doi.org/10.1007/1-4020-4925-0_2
- Campbell III E, Rodda GH, Fritts T, Bruggers R (1999) An integrated management plan for the Brown Treesnake (Boiga irregularis) on Pacific islands. In: Rodda GH, Sawai Y, Chiszar D, Tanaka H (Eds) Problem Snake Management: the Habu and the Brown Treesnake. Cornell University Press, Ithaca, New York, 423–435.
- Campbell JA (1998) Amphibians and Reptiles of Northern Guatemala, the Yucatán, and Belize. University of Oklahoma Press, Norman, 380 pp.
- Capinha C, Seebens H, Cassey P, García-Díaz P, Lenzner B (2017) Diversity, biogeography and the global flows of alien amphibians and reptiles. Diversity and Distributions 23(11): 1313–1322. https://doi.org/10.1111/ddi.12617
- Card DC, Schield DR, Adams RH, Corbin AB, Perry BW, Andrew AL, Pasquesi GIM, Smith EN, Jezkova T, Boback SM, Booth W, Castoe TA (2016) Phylogeographic and population genetic analysis reveal multiple species of Boa and independent origins of insular dwarfism. Molecular Phylogenetics and Evolution 102: 104–116. https://doi.org/10.1016/j.ympev.2016.05.034
- Carlton JT (1996) Biological invasions and cryptogenic species. Ecology 77(6): 1653–1655. https://doi.org/10.2307/2265767
- Carranza S, Arnold EN (2006) Systematics, biogeography, and evolution of Hemidactylus geckos (Reptilia:Gekkonidae) elucidated using mitochondrial DNA sequences. Molecular Phylogenetics and Evolution 38(2): 531–545. https://doi.org/10.1016/j.ympev.2005.07.012
- Casas-Andreu G, Aguilar X, Cruz RA (2001) La introducción y el cultivo de la rana toro (Rana catesbeiana) ¿Un atentado a la biodiversidad de México? Ciencia Eergo-sum 8(1): 62–67.
- Castro-Franco R, Bustos-Zagal MG (2004) Additional records and range extensions of reptiles from Morelos, México. Herpetological Review 35(2): 196–197.
- Cedeño-Vázquez JR, González-Vázquez J, Martínez-Arce A, Canseco-Márquez L (2014) First record of the invasive greenhouse frog (Eleutherodactylus planirostris) in the Mexican Caribbean. Revista Mexicana de Biodiversidad 85(2): 650–653. https://doi.org/10.7550/rmb.43234
- Cedeño-Vázquez JR, Platt SG, Thorbjarnarson J (2012) Crocodylus moreletii. The IUCN Red List of Threatened Species 2012. [Accessed 6 February 2021]
- Cedeño-Vázquez JR, Rodriguez D, Calmé S, Ross JP, Densmore LD, Thorbjarnarson JB (2008) Hybridization between Crocodylus acutus and Crocodylus moreletii in the Yucatan Peninsula: I. evidence from mitochondrial DNA and morphology. Journal of Experimental Zoology Part A: Ecological Genetics and Physiology 309(10): 661–673. https://doi.org/10.1002/jez.473
- Charruau P, Díaz de la Vega Pérez AH, Méndez de la Cruz FR (2015) Reptiles of Banco Chinchorro: updated list, life history data, and conservation. The Southwestern Naturalist 60(4): 299–312. https://doi.org/10.1894/0038-4909-60.4.299
- Clarke GS, Crossland MR, Shine R (2016) Can we control the invasive cane toad using chemicals that have evolved under intraspecific competition? Ecological Applications 26(2): 463–474. https://doi.org/10.1890/14-2365
- Clark L, Savarie PJ (2012) Efficacy of aerial broadcast baiting in reducing brown treesnake numbers. Human-Wildlife Interaction 6(2): 212–221.
- Clark L, Savarie PJ, Shivik JA, Breck SW, Dorr BS (2012) Efficacy, effort, and cost comparisons of trapping and acetaminophen-baiting for control of brown treesnakes on Guam. Human-Wildlife Interactions 6(2): 222–236. https://www.jstor.org/stable/24874096
- Cole NC, Jones CG, Harris S (2005) The need for enemy-free space: The impact of an invasive gecko on island endemics. Biological Conservation 125(4): 467–474. https://doi.org/10.1016/j.biocon.2005.04.017
- Colston TJ, Barão-Nóbrega JAL, Manders R, Lett A, Wilmott J, Cameron G, Hunter S, Radage A, Littlefair E, Williams RJ, Lopez Cen A, Slater K (2015) Amphibians and reptiles of the Calakmul Biosphere Reserve, México, with new records. Check List 11(5): 1–7. https://doi.org/10.15560/11.5.1759
- CONABIO (2018) Método de Evaluación Rápida de Invasividad (MERI) para especies exóticas en México Boa constrictor Linnaeus, 1758. https://www.gob.mx/cms/uploads/attachment/file/222314/Boa_constrictor_A.pdf [Accessed 6 February 2021]
- CONANP (2018) Programa de Manejo Parque Nacional Revillagigedo, Mexico. Comisión Nacional de Areas Naturales Protegidas, 328 pp.
- Conant R, Collins JT (1998) A Field Guide to Reptiles and Amphibians of Eastern and Central North America, Third edition. Houghton Mifflin, Boston, Massachusetts, 450 pp.
- Contreras-Calvario AI, Mora-Reyes A, Parra-Olea G, Mendoza AM (2018) New records of the introduced Eleutherodactylus planirostris (Anura: Eleutherodactylidae), in the state of Veracruz, México. Herpetological Journal 28(2): 96–99.
- Crawford AJ, Alonso R, Jaramillo CA (2011) DNA barcoding identifies a third invasive species of Eleutherodactylus (Anura: Eleutherodactylidae) in Panama City, Panama. Zootaxa 2890(1): 65–67. https://doi.org/10.11646/zootaxa.2890.1.6
- Crooks JA (2002) Characterizing ecosystem level consequences of biological invasions: the role of ecosystem engineers. Oikos 97(2): 153–166. https://doi.org/10.1034/j.1600-0706.2002.970201.x
- Crumly CR, Grismer LL (1994) Validity of the tortoise Xerobates lepidocephalus Ottley and Velazques in Baja California. In: Bury RB, Germano DJ (Eds) Biology of North American Tortoises. Fish and Wildlife Research 13: 33–37.
- Cruz-Sáenz D, Muñoz-Nolasco FJ, Mata-Silva V, Johnson JD, García-Padilla E, Wilson LD (2017) The herpetofauna of Jalisco, Mexico: composition, distribution, and conservation status. Mesoamerican Herpetology 4(1): 22–118.
- Cupul-Magaña FG (2012) Varanus exanthematicus (Bosc, 1792), Apalone spinifera emoryi (Le Sueur, 1827) y Gopherus berlandieri (Agassiz, 1857): reptiles exóticos en el área urbana de Puerto Vallarta, Jalisco, México. Cuadernos de Herpetología 26(1): 59–60. http://hdl.handle.net/10915/18207
- Cupul-Magaña FB, Rubio-Delgado A (2003) Distribución de dos especies de tortugas dulceacuícolas, Kinosternon chimalhuaca y Chelydra serpentina (Testudines), en Puerto Vallarta, Jalisco, México. Boletín de la Sociedad Herpetológica Mexicana 11: 49–50.
- Davis JL, Alford RA, Schwarzkopf L (2015) Some lights repel amphibians: implications for improving trap lures for invasive species. International Journal of Pest Management 61(4): 305–311. https://doi.org/10.1080/09670874.2015.1058991
- Davis MA (2009) Invasion Biology. Oxford University Press on Demand, Oxford, 244 pp.
- Davy CM, Méndez-de la Cruz FR, Lathrop A, Murphy RW (2011) Seri Indian traditional knowledge and molecular biology agree: no express train for island-hopping spiny-tailed iguanas in the Sea of Cortés. Journal of Biogeography 38(2): 272–284. https://doi.org/10.1111/j.1365-2699.2010.02422.x
- Dayton GH, Croll DA, Fuentes Cuevas S (2020) Geographic distribution: Mexico, Baja California Sur: Hemidactylus frenatus. Herpetological Review 51(2): e271.
- Dejean T, Valentini A, Miquel C, Taberlet P, Bellemain E, Miaud C (2012) Improved detection of an alien invasive species through environmental DNA barcoding: the example of the American bullfrog, Lithobates catesbeianus. Journal of Applied Ecology 49(4): 953–959. https://doi.org/10.1111/j.1365-2664.2012.02171.x
- de Sousa F, Arosemena F, Castillo JA, Mallorga HM (1989) Una nueva distribución geográfica de Eleutherodactylus antillensis (Reinhardt y Lütken, 1863) (Aamphibia: Anura: Leptodactylidae), identificación y hábitos ecológicos en la ciudad de Panamá. Sciencia (Panamá) 1989: 87–102.
- de Villa-Meza A, Avila-Flores R, Cuarón AD, Valenzuela-Galván D (2011) Procyon pygmaeus (Carnivora: Procyonidae). Mammalian Species 43(877): 87–93. https://doi.org/10.1644/877.1
- D’hondt B, Vanderhoeven S, Roelandt S, Mayer F, Versteirt V, Adriaens T, Ducheyne E, San Martin G, Grégoire J-C, Stiers I, Quoilin S, Cigar J, Heughebaert A, Branquart E (2015) Harmonia+ and Pandora+: risk screening tools for potentially invasive plants, animals and their pathogens. Biological Invasions 17(6): 1869–1883. https://doi.org/10.1007/s10530-015-0843-1
- Dixon JR (2013) Amphibians and Reptiles of Texas: With Keys, Taxonomic Synopses, Bibliography, and Distribution Maps. Texas A&M University Press, College Station, 443 pp.
- D.O.F. (2015) Proyecto de Modificación del Anexo Normativo III, Lista de especies en riesgo de la Norma Oficial Mexicana NOM-059-SEMARNAT-2010, Protección ambiental-Especies nativas de México de flora y fauna silvestresCategorías de riesgo y especificaciones para su inclusión, exclusión o cambio-Lista de especies en riesgo. Diario Oficial de la Federación, 78 pp.
- D.O.F. (2016) Acuerdo mediante el cual se dan a conocer en los Estados Unidos Mexicanos las enfermedades y plagas exóticas y endémicas de notificación obligatoria de los animales terrestres y acuáticos. Diario Oficial de la Federación, 61–76.
- Duellman WE (2001) The Hylid frogs of Middle America (Vol. 2). Society for the Study of Amphibians and Reptiles, Contibutions in Herpetology 18, Society for the Study of Amphibians and Reptiles, Ithaca, New York, 1158 pp.
- Echternacht A, Burton F, Blumenthal J (2011) The amphibians and reptiles of the Cayman Islands: Conservation issues in the face of invasions. Conservation of Caribbean Island Herpetofaunas 1: 129–147. https://doi.org/10.1163/ej.9789004194083.i-439.49
- Edwards T, Bonine KE, Ivanyi C, Prescott R (2005) The molecular origins of spiny-tailed iguanas (Ctenosaura) on the grounds of the Arizona-Sonora Desert Museum. Sonoran Herpetologist 18(11): 122–125.
- Edwards T, Jarchow CJ, Jones CA, Bonine KE (2010) Tracing genetic lineages of captive desert tortoises in Arizona. The Journal of Wildlife Management 7(4): 801–807. https://doi.org/10.2193/2009-199
- Edwards T, Karl A, Vaughn M, Rosen P, Meléndez Torres C, Murphy RW (2016) The desert tortoise trichotomy: Mexico hosts a third, new sister-species of tortoise in the Gopherus morafkai – G. agassizii group. ZooKeys 562: 131–158. https://doi.org/10.3897/zookeys.562.6124
- Farr WL (2011) Distribution of Hemidactylus frenatus in Mexico. The Southwestern Naturalist 56(2): 265–273. https://doi.org/10.1894/N06-FJRR-01.1
- Farr WL (2015) Herpetofauna of Tamaulipas. Chapter 8. In: Lemos-Espinal JA, Hollingsworth B, Mahrdt C, Brennan T, Babb R. and Painter C (Eds) Amphibians and Reptiles of the US-Mexico Border States/Anfibios y reptiles de los estados de la frontera México-Estados Unidos. Texas A&M University Press, 101–121.
- Finnerty PB, Shine R, Brown GP (2018) The costs of parasite infection: effects of removing lungworms on performance, growth and survival of free-ranging cane toads. Functional Ecology 32(2): 402–415. https://doi.org/10.1111/1365-2435.12992
- Fisher RN (1997) Dispersal and evolution of the Pacific Basin gekkonid lizards Gehyra oceanica and Gehyra mutilata. Evolution 51(3): 906–921. https://doi.org/10.1111/j.1558-5646.1997.tb03672.x
- Fitch HS (1975) Sympatry and interrelationships in Costa Rican anoles. Occasional Papers of the Museum of Natural History of the University of Kansas, University of Kansas 40: 1–60. https://doi.org/10.5962/bhl.part.17265
- Frankenberg E (1982) Social behaviour of the parthenogenetic Indo-Pacific Gecko, Hemidactylus garnotii. Zeitschrift für Tierpsychologie 59(1): 19–28. https://doi.org/10.1111/j.1439-0310.1982.tb00329.x
- Frankenberg E (1984) Interactions between two Species of colonizing house geckos, Hemidactylus turcicus and Hemidactylus garnotii. Journal of Herpetology 18(1): 1–7. https://doi.org/10.2307/1563665
- Fritz U, Stuckas H, Vargas-Ramírez M, Hundsdörfer AK, Maran J, Päckert M (2012) Molecular phylogeny of Central and South American slider turtles: implications for biogeography and systematics (Testudines: Emydidae: Trachemys. Journal of Zoological Systematics and Evolutionary Research 50(2): 125–136. https://doi.org/10.1111/j.1439-0469.2011.00647.x
- Frost DR (2020) Amphibian Species of the World: an online database. Version 6.1. American Museum of Natural History, New York, New York. Electronic Database accessible at https://amphibiansoftheworld.amnh.org/index.php [Accessed 6 February 2021]
- Fujita MK, Moritz C (2009) Origin and evolution of parthenogenetic genomes in lizards: current state and future directions. Cytogenetic and Genome Research 127(2–4): 261–272. https://doi.org/10.1159/000295177
- Gallardo B, Clavero M, Sánchez Marta I, Vilà M (2015) Global ecological impacts of invasive species in aquatic ecosystems. Global Change Biology 22(1): 151–163. https://doi.org/10.1111/gcb.13004
- Gamble T, Bauer AM, Colli GR, Greenbaum E, Jackman TR, Vitt LJ, Simons AM (2011) Coming to America: multiple origins of New World Geckos. Journal of Evolutionary Biology 24(2): 231–244. https://doi.org/10.1111/j.1420-9101.2010.02184.x
- García-Balderas CM, Cedeño-Vázquez JR, Mineros-Ramírez R (2016) Distribution Notes. Eleutherodactylus planirostris. Mesoamerican Herpetology 3(1): e173.
- García-Llorente M, Martín-López B, González JA, Alcorlo P, Montes C (2008) Social perceptions of the impacts and benefits of invasive alien species: Implications for management. Biological Conservation 141(12): 2969–2983. https://doi.org/10.1016/j.biocon.2008.09.003
- García-Llorente M, Martín-López B, Nunes PALD, González JA, Alcorlo P, Montes C (2011) Analyzing the social factors that influence willingness to pay for invasive alien species management under two different strategies: eradication and prevention. Environmental Management 48(3): 418–435. https://doi.org/10.1007/s00267-011-9646-z
- García-Rodríguez A, Chaves G, Wainwright M, Villegas A (2010) Geographic distribution: Costa Rica, Cartago, Turrialba District: Eleutherodactylus coqui. Herpetological Review 41(3): e375.
- García-Vázquez UO, Canseco-Márquez L, Gutiérrez-Mayén G, Trujano-Ortega M (2009) Actualización del conocimiento de la fauna herpetológica en el estado de Puebla, México. Boletín de la Sociedad Herpetológica Mexicana 17(1): 12–36.
- Garman S (1887) On the Reptiles and Batrachians of Grand Cayman. Proceedings of the American Philosophical Society 24(126): 273–277. https://www.jstor.org/stable/983076
- Garner TW, Perkins MW, Govindarajulu P, Seglie D, Walker S (2006) The emerging amphibian pathogen Batrachochytrium dendrobatidis globally infects introduced populations of the North American bullfrog, Rana catesbeiana. Biology Letters 2(3): 455–459. https://doi.org/10.1098/rsbl.2006.0494
- GBIF.org (2018) Global Biodiversity Information Facility. https://www.gbif.org [Accessed 6 February 2021]
- Glor RE, Losos JB, Larson A (2005) Out of Cuba: overwater dispersal and speciation among lizards in the Anolis carolinensis subgroup. Molecular Ecology 14(8): 2419–2432. https://doi.org/10.1111/j.1365-294X.2005.02550.x
- González-Martínez AI, Barrios-Caballero Y, de Jesús S (2017) Análisis de riesgo para especies invasoras en México. El impacto de las especies exóticas invasoras en México. Revista Legislativa de Estudios Sociales y de Opinión Pública 103: 24–29.
- González-Sánchez VH, Johnson JD, García-Padilla E, Mata-Silva V, DeSantis DL, Wilson LD (2017) The herpetofauna of the Mexican Yucatan Peninsula: composition, distribution, and conservation. Mesoamerican Herpetology 4(2): 264–380.
- Greenlees MJ, Harris S, White AW, Shine R (2018) The establishment and eradication of an extra-limital population of invasive cane toads. Biological Invasions 20(8): 2077–2089. https://doi.org/10.1007/s10530-018-1681-8
- Grismer LL (2002) Amphibians and Reptiles of Baja California, Including its Pacific Islands and the Islands in the Sea of Cortés. University of California Press, Berkeley, California, 409 pp. https://doi.org/10.1525/9780520925205
- Gutsche A, McCranie JR (2009) Geographic distribution: Honduras, Islas de la Bahía: Hemidactylus mabouia. Herpetological Review 4(1): 112–113.
- Guzmán-Guzmán S, Palma-Martínez M (2016) Geographic distribution: México, Veracruz, Municipality of Minatitlán: Gonatodes albogularis. Herpetological Review 47(3): e423.
- Hallowell E (1854) Description of new species of reptilia from western Africa. Proceedings of the Academy of Natural Sciences of Philadelphia 64: 62–65.
- Haramura T, Crossland MR, Takeuchi H, Shine R (2017) Methods for invasive species control are transferable across invaded areas. PLoS ONE 12(11): e0187265. https://doi.org/10.1371/journal.pone.0187265
- Harris DM, Kluge AG (1984) The Sphaerodactylus (Sauria: Gekkonidae) of Middle America. Occasional Papers of the Museum of Zoology, University of Michigan (706): 1–59. http://hdl.handle.net/2027.42/57142
- Henderson R, Villa J, Dixon JR (1976) Lepidodactylus lugubris (Reptilia: Gekkonidae): a recent addition to the herpetofauna of Nicaragua. Herpetological Review 7(4): e173.
- Hollingsworth B, Mahrdt CR, Grismer LL, Banta BH, Sylber CK (1997) The occurrence of Sauromalus varius on a satellite islet of Isla Salsipuedes, Gulf of California, México. Herpetological Review 28(1): 26–28.
- Hofman CA, Rick TC (2018) Ancient Biological Invasions and Island Ecosystems: Tracking Translocations of Wild Plants and Animals. Journal of Archaeological Research 26(1): 65–115. https://doi.org/10.1007/s10814-017-9105-3
- Hoogmoed MS, Avila-Pires TC (2015) Lepidodactylus lugubris (Duméril & Bibron 1836) (Reptilia: Gekkonidae), an introduced lizard new for Brazil, with remarks on and correction of its distribution in the New World. Zootaxa 4000(1): 90–110. https://doi.org/10.11646/zootaxa.4000.1.4
- Hulme PE (2014) Invasive species challenge the global response to emerging diseases. Trends in Parasitology 30(6): 267–270. https://doi.org/10.1016/j.pt.2014.03.005
- Hunter ME, Oyler-McCance SJ, Dorazio RM, Fike JA, Smith BJ (2015) Environmental DNA (eDNA) sampling Improves occurrence and detection estimates of Invasive Burmese Pythons. PLoS ONE 10(4): 1–17. https://doi.org/10.1371/journal.pone.0121655
- Ibañez R, Rand SA (1990) Geographic distribution: Panama, Province of Panama: Eleutherodactylus johnstonei. Herpetological Review 21(1): e37.
- Ineich I, Blanc CP (1987) Le peuplement herpétologique de Polynésie française. Adaptations et aléas. Bulletin de la Societe Zoologique de France 112(3–4): 381–400.
- Inaturalist.org (2018) Inaturalist.org [Accessed 6 February 2021]
- IUCN (2014) IUCN Red List of Threatened Species. Version 2014.2. www.iucnredlist.org [Accessed 6 February 2021]
- IUCN (2015) Lithobates catesbeianus [Online]. [Accessed 6 February 2021]
- Iverson JB, Young CA, Berry JF (1998) Kinosternon integrum LeConte (Mexican Mud Turtle). Society for the Study of Amphibians and Reptiles, Catalogue of American Amphibians and Reptiles (CAAR): 652.1–652.6.
- Jaramillo C, Wilson LD, Ibáñez R, Jaramillo F (2010) The herpetofauna of Panama: distribution and conservation status. In: Wilson LD, Townsend JH, Johnson JD (Eds) Conservation of Mesoamerican Amphibians and Reptiles. Eagle Mountain Publishing, LC, Eagle Mountain, Utah, 604–671.
- Jerde CL, Mahon AR, Chadderton WL, Lodge D (2011) “Sight-unseen” detection of rare aquatic species using environmental DNA. Conservation Letters 4(2): 150–157. https://doi.org/10.1111/j.1755-263X.2010.00158.x
- Jiménez-Arcos VH, Cruz-Padilla SS, Escalona-López A, Casas-Andreu G, Centenero-Alcalá E (2009) Geographic distribution: México, Guerrero, Municipality of Copalillo: Apalone spinifera. Herpetological Review 40(4): e448.
- Jiménez RR, Abarca JG (2014) Extensión de rango del gecko Lepidodactylus lugubris en la vertiente Caribe de Costa Rica. Herpetotropicos 10(1–2): 31–32.
- Johnson JD, Mata-Silva V, Ramírez-Bautista A (2010) Geographic distribution and conservation of the herpetofauna of southeastern Mexico. In: Wilson LD, Townsend JH, Johnson JD (Eds) Conservation of Mesoamerican Amphibians and Reptiles. Eagle Mountain Publishing, LC, Eagle Mountain, Utah, 322–369.
- Johnson JD, Mata-Silva V, Padilla EG, Wilson LD (2015a) The Herpetofauna of Chiapas, Mexico: composition, distribution, and conservation. Mesoamerican Herpetology 2(3): 272–329.
- Johnson JD, Mata-Silva V, Wilson LD (2015b) A conservation reassessment of the Central American herpetofauna based on the EVS measure. Amphibian & Reptile Conservation 9(2): 1–94.
- Kaiser H, Henderson RW (1994) The conservation status of Lesser Antillean frogs. Herpetological Natural History 2(2): 41–56.
- Kelehear C, Cabrera‐Guzmán E, Shine R (2012) Inadvertent consequences of community‐based efforts to control invasive species. Conservation Letters 5(5): 360–365. https://doi.org/10.1111/j.1755-263X.2012.00251.x
- Kluge AG (1969) The evolution and geographical origin of the New World Hemidactylus mabouia-brooki complex (Gekkonidae, Sauria). Miscellaneous Publications Museum of Zoology. University of Michigan (138): 1–78.
- Kôhler G (1999) The Amphibians and Reptiles of Nicaragua: A distributional checklist with keys. Courier Forschungsinstitut Senckenberg, Frankfurt a. M. 213: 1–121.
- Kôhler G (2001) Anfibios y Reptiles de Nicaragua. Herpeton, Verlag Elke Kôhler, Rohrstr. 22, 63075 Offenbach, Germany, 208 pp.
- Köhler G (2008) Reptiles of Central America (2nd Edn.). Herpeton Verlag Elke Köhler, Offenbach, Germany, 400 pp.
- Köhler G (2011) Amphibians of Central America. Herpeton, Verlag Elke Köhler, Offenbach, Germany, 379 pp.
- Kolby JE, Smith KM, Berger L, Karesh WB, Preston A, Pessier AP, Skerratt LF (2014) First evidence of amphibian chytrid fungus (Batrachochytrium dendrobatidis) and Ranavirus in Hong Kong amphibian trade. PLoS ONE 9(3): e90750. https://doi.org/10.1371/journal.pone.0090750
- Kraus F (2003) Invasion pathways for terrestrial vertebrates. In: Ruiz GM, Carlton JT (Eds) Bioinvasions: Pathways, Vectors, and Management Strategies. Island Press, New York, New York, 68–92.
- Kraus F (2009) Alien Reptiles and Amphibians: A Scientific Compendium and Analysis. Springer Science, New York, New York, 562 pp. https://doi.org/10.1007/978-1-4020-8946-6
- Kuchta SR (2005) Taricha torosa. In: Lannoo MJ (Ed.) Amphibian Declines: The Conservation Status of United States Species. University of California Press, Berkley, 904–908.
- Lavín PA, Lazcano D, Gadsden H (2014) Anfibios y reptiles exóticos y traslocados invasores. In: Mendoza R, Koleff P (Eds) Especies Acuáticas Invasoras en México, Comisión Nacional para el Conocimiento y Uso de la Biodiversidad (CONABIO), Mexico City, Mexico, 435–444.
- Lazcano-Barrero M (1993) Plan de acción para la conservación del cocodrilo americano (Crocodylus acutus) en Isla Contoy, Quintana Roo. Technical Report, Reserva Ecológica El Edén A. C. , Cancún, Quintana Roo, México, 9 pp.
- Lazcano D, Mendoza-Alfaro R, Campos-Múzquiz L, Lavin-Murcio PA, Quiñónez-Martínez M (2010) Notes on Mexican herpetofauna 15: the risk of invasive species in northeastern Mexico. Bulletin of the Chicago Herpetological Society 45(7): 113–117.
- Lee JC (1996) The Amphibians and Reptiles of the Yucatán Peninsula. Comstock Publishing Asociates, Cornell University Press, Ithaca, New York, 500 pp.
- Leets-Rodríguez L, López-Guevara H, Sunyer J (2019) First country records of the invasive Brahminy Blindsnake Indotyphlops braminus (Daudin, 1803) (Squamata, Typhlopidae) from Nicaragua. Revista Nicaragüense de Biodiversidad (45): 3–11.
- Legler JM, Vogt RC (2013) Turtles of Mexico: Land and Freshwater Forms, University of California Press. Berkeley, 402 pp. https://doi.org/10.1525/9780520956896
- Lemos-Espinal JA (2015) Amphibians and Reptiles of the US–Mexico Border States. Texas A&M University Press. College Station, 614 pp.
- Lemos-Espinal JA, Dixon JR (2013) Amphibians and Reptiles of San Luis Potosí. Eagle Mountain Publishing, LC, Eagle Mountain, Utah, 300 pp.
- Lemos-Espinal JA, Smith GR, Woolrich-Peña GA, Cruz A (2017) Amphibians and Reptiles of Chihuahua, Mexico, with comparisons with adjoining states. ZooKeys 658: 105–130. https://doi.org/10.3897/zookeys.658.10665
- Lemos-Espinal JA, Smith GR (2016) Amphibians and reptiles of the state of Coahuila, Mexico, with comparison with adjoining states. ZooKeys 593: 117–137. https://doi.org/10.3897/zookeys.593.8484
- Lemos-Espinal JA, Smith HM (2007a) Anfibios y Reptiles del Estado de Chihuahua, México. UNAM, CONABIO, Mexico, 613 pp.
- Lemos-Espinal JA, Smith HM (2007b) Anfibios y Reptiles del Estado de Coahuila, México. UNAM, CONABIO, México, 550 pp.
- Lemos-Espinal JA, Dixon JR (2013) Amphibians and Reptiles of San Luis Potosí. Eagle Mountain Publishing, Eagle Mountain, Utah, 300 pp.
- Lemos-Espinal JA, Webb RG, Smith HM (1999) Emory’s Softshell Turtle, Apalone spinifera emoryi, in Mexico. Bulletin of the Maryland Herpetological Society 35: 40–42.
- Liner EA, Casas-Andreu G (2008) Standard Spanish, English and Scientific names of the Amphibians and Reptiles of Mexico. Herpetological Circular 38, Society for the Study of Amphibians and Reptiles, Lawrence, Kansas, 162 pp.
- Lira-Noriega A, Ramírez H (2016) Análisis de riesgo detallado para cinco especies de reptiles con potencial invasor de alto riesgo para México. In: CONABIO (Ed.) Proyecto GEF Aumentar las Capacidades Nacionales para el Manejo de las Especies Exóticas Invasoras (EEI) a través de la Implementación de la Estrategia Nacional de EEI. Ciudad de México, México, 129 pp.
- López-González C (1991) Estudio Prospectivo de los Vertebrados Terrestres del Corredor Turístico Cancún-Tulum, Quintana Roo, Mexico. Tesis de Licenciatura, Universidad Nacional Autónoma de Mexico, 127 pp.
- Lorvelec O, Pascal M (2006) Les vertébrés de clipperton soumis à un siècle et demi de bouleversements écologiques. Revue D’écologie 61(2): 135–158. http://hdl.handle.net/2042/55675
- Lovich RE, Grismer LL, Danemann G (2009) Conservation status of the herpetofauna of Baja California, México and associated islands in the Sea of Cortez and Pacific Ocean. Herpetological Conservation and Biology 4(3): 358–378.
- Lovich RE, Mahrdt CR (2007) Herpetofauna terrestre. In: Danemann G, Ezcurra E (Eds) Bahía de Los Angeles: Recursos Naturales y Comunidad. Línea Base 2007. Pronatura Noroeste AC, Secretaría de Recursos Naturales y Medio Ambiente, Instituto Nacional de Ecología y Museo de Historia Natural de San Diego, 495–521.
- Lowe S, Browne M, Boudjelas S, De Poorter M (2000) 100 of the World’s Worst Invasive Alien Species: A Selection from the Global Invasive Species Database. Invasive Species Specialist Group, Auckland, New Zealand, 12 pp.
- Luja VH, Blázquez MC, Rodríguez-Estrella R (2007) Tortuga introducida: reporte de Kinosternon integrum (Leconte, 1854) en Baja California Sur. Boletín de la Sociedad Herpetológica Mexicana 15(2): 40–41.
- Machkour-M’Rabet S, Hénaut Y, Charruau P, Gevrey M, Winterton P, Legal L (2009) Between introgression events and fragmentation, islands are the last refuge for the American crocodile in Caribbean Mexico. Marine Biology 156(6): 1321–1333. https://doi.org/10.1007/s00227-009-1174-5
- MacIsaac H, Tedla R, Ricciardi A (2010) Patterns and Rate of Growth of Studies. In: Ricardson DM (Ed.) Fifty Years of Invasion Ecology: The Legacy of Charles Elton. Blackwell Publishing Ltd. , Hoboken, New Jersey, 456 pp. https://doi.org/10.1002/9781444329988.ch5
- Mack RN, Simberloff D, Mark Lonsdale W, Evans H, Clout M, Bazzaz FA (2000) Biotic invasions: causes, epidemiology, global consequences, and control. Ecological Applications 10(3): 689–710. https://doi.org/10.1890/1051-0761(2000)010[0689:BICEGC]2.0.CO;2
- Marr SR, Johnson SA, Hara AH, McGarrity ME (2010) Preliminary evaluation of the potential of the helminth parasite Rhabdias elegans as a biological control agent for invasive Puerto Rican Coquís (Eleutherodactylus coqui) in Hawaii. Biological Control 54(2): 69–74. https://doi.org/10.1016/j.biocontrol.2010.04.012
- Martínez-Hernández A, Pineda-Manzano U, Bauer AM, Vázquez-Villa BM (2017) First record of the Mediterranean House Gecko, Hemidactylus turcicus Linnaeus 1758 in the metropolitan area of San Luis Potosí, Mexico. IRCF Reptiles & Amphibians 24(2): 143–147.
- Martínez-Morales MA, Cuarón AD (1999) Boa constrictor, an introduced predator threatening the endemic fauna on Cozumel Island, Mexico. Biodiversity & Conservation 8(7): 957–963. https://doi.org/10.1023/A:1008815004072
- Mata-Silva V, Johnson JD, Wilson LD, Rocha A, García-Grajales J (2013) Geographic distribution: Mexico, Oaxaca: Hemidactylus frenatus. Herpetological Review 44(4): 625–626.
- Mayer GC (2010) Ecological distribution of the Puerto Rican crested anole, Anolis cristatellus cristatellus, in Cahuita, Costa Rica. Anolis Newsletter VI: 138–144.
- McCoy CJ (1970) Hemidactylus turcicus. Catalogue of American Amphibians and Reptiles (CAAR), 87.1–87.2.
- McCranie JR (2015) A checklist of the amphibians and reptiles of Honduras, with additions, comments on taxonomy, some recent taxonomic decisions, and areas of further studies needed. Zootaxa 3931: 352–386. https://doi.org/10.11646/zootaxa.3931.3.2
- McCranie JR (2018) The lizards, crocodiles, and turtles of Honduras: systematics, distribution, and conservation. Bulletin of the Museum of Comparative Zoology 15(1): 1–129. https://doi.org/10.5962/bhl.title.155244
- McCranie JR, Gutsche A (2009) Geographic distribution: Honduras, Atlántida: Anolis allisoni. Herpetological Review 40(1): e112.
- McCranie JR, Gutsche A (2014) Geographic distribution: Honduras, Department of La Paz: Eleutherodactylus (Euhyas) planirostris. Herpetological Review 45(4): e653.
- McCranie JR, Harrison A, Valdés-Orellana L (2017) Updated Population and Habitat Comments about the reptiles of the Swan Islands, Honduras. Bulletin of the Museum of Comparative Zoology 161(7): 265–284. https://doi.org/10.3099/MCZ34.1
- McCranie JR, Hedges SB (2012) Two new species of geckos from Honduras and resurrection of Sphaerodactylus continentalis Werner from the synonymy of Sphaerodactylus millepunctatus Hallowell (Reptilia, Squamata, Gekkonoidea, Sphaerodactylidae). Zootaxa 3492: 65–76.
- McCranie JR, Hedges SB (2013) A review of the Cnemidophorus lemniscatus group in Central America (Squamata: Teiidae), with comments on other species in the group. Zootaxa 3722(3): 301–316. https://doi.org/10.11646/zootaxa.3722.3.1
- McCranie JR, Köhler G (2015) The anoles (Reptilia: Squamata: Dactyloidae: Anolis: Norops) of Honduras. Systematics, Distribution, and Conservation. Bulletin of the Museum of Comparative Zoology 161(11): 1–280. https://doi.org/10.3099/0027-4100-14.1.1
- McCranie JR, Köhler F, Gutsche A, Valdés-Orellana L (2013) Trachemys grayi emolli (Testudines, Emydidae) in Honduras and its systematic relationships based on mitochondrial DNA. Zoosystematics and Evolution, Museum für Naturkunde 89(1): 21–29. https://doi.org/10.1002/zoos.201300003
- McCranie JR, Wilson LD, Köhler G (2005) Amphibians and Reptiles of the Bay Islands and Cayos Cochinos, Honduras. Bibliomania! Salt Lake City, Utah, 210 pp.
- McCranie JR, Townsend JH, Wilson LD (2006) The Amphibians and Reptiles of the Honduran Mosquitia. Krieger Publishing Company, Malabar, 291 pp.
- McCranie JR, Valdés-Orellana L (2014) New island records and updated nomenclature of amphibians and reptiles from the Islas de la Bahía, Honduras. Herpetology Notes 7: 41–49.
- McGaugh SE, Janzen FJ (2008) The status of Apalone atra populations in Cuatro Ciénegas, Coahuila, México: preliminary data. Chelonian Conservation and Biology 7(1): 88–95. https://doi.org/10.2744/CCB-0651.1
- McNeely JA (2001) An introduction to human dimensions of invasive alien species. In: McNeely JA (Eds) The Great Reshuffling. Human Dimensions of Invasive Alien Species. IUCN. Gland, Switzerland and Cambridge, United Kingdom, 5–22.
- Measey GJ, Rödder D, Green SL, Kobayashi R, Lillo F (2012) Ongoing invasions of the African clawed frog, Xenopus laevis: a global review. Biological Invasions 14(11): 2255–2270. https://doi.org/10.1007/s10530-012-0227-8
- Meerman J, Garel J (2002) Tokay gecko Gekko gecko. (L) established on South Water Caye, Belize. http://biological-diversity.info/tokay.htm [Accessed 6 February 2021]
- Mellink E (2002) Invasive vertebrates on islands of the Sea of Cortés. Invasive exotic species in the Sonoran region. University of Arizona Press, Tucson, 112–125.
- Mellink E, Ferreira-Bartrina V (2000) On the wildlife of wetlands of the Mexican portion of the Río Colorado Delta. Bulletin of the Southern California Academy of Sciences 99(3): 115–115. https://scholar.oxy.edu/scas/vol99/iss3/2
- Mertz WH, Costa-Campos CE, Valentim DSS, Furtado MFM (2015) Geographic distribution: México: Guerrero, Municipality of Zihuatanejo: Trachemys ornata. Herpetological Review 46(1): e58.
- Miller M (2006) Effectiveness of Cane Toad Eradication Methods: Arakwal National Park. Independent Study Project Collection, 324, 34 pp. https://digitalcollections.sit.edu/isp_collection/324.
- Miller RR (1946) The probable origin of the soft-shelled turtle in the Colorado River basin. Copeia 1946(1): e46. https://doi.org/10.2307/1438829
- Moll EO (1995) The turtle Trachemys scripta and the pet trade. Aliens 1: 1–3.
- Morales JR, Chang MJ, Acevedo ME (2017) First records of Hemidactylus garnotii Duméril & Bibron, 1836 (Squamata: Gekkonidae) in Guatemala. Mesoamerican Herpetology 4(4): 953–955.
- Muller BJ, Schwarzkopf L (2017) Relative effectiveness of trapping and hand-capture for controlling invasive cane toads (Rhinella marina). International Journal of Pest Management 64(2): 185–192. https://doi.org/10.1080/09670874.2017.1363443
- Muñoz-Alonso LA (2006) Actualización y enriquecimiento de las bases de datos del proyecto de evaluación y análisis geográfico de la diversidad faunística de Chiapas. El Colegio de la Frontera Sur. Informe final SNIB-CONABIO proyecto No. U014. México. http://www.conabio.gob.mx/institucion/proyectos/resultados/InfU014.pdf [Accessed 6 February 2021]
- Murphy RW (1983) Paleobiogeography and genetic differentiation of the Baja California herpetofauna. Occasional Papers of the California Academy of Sciences 137: 1–48.
- Murphy RW, Aguirre-León G (2002) The nonavian reptiles: origins and evolution. In: Case TJ, Cody ML, Ezcurra E (Eds) A New Island Biogeography of the Sea of Cortés. Oxford University Press, New York, 181–270. https://doi.org/10.1093/oso/9780195133462.003.0014
- Murphy RW, Berry KH, Edwards T, Leviton AE, Lathrop A (2011) The dazed and confused identity of Agassiz’s land tortoise, Gopherus agassizii (Testudines, Testudinidae) with the description of a new species, and its consequences for conservation. ZooKeys 113: 39–71. https://doi.org/10.3897/zookeys.113.1353
- Murphy RW, Méndez de la Cruz FR (2010) The Herpetofauna of Baja California and its associated islands; a conservation assessment and priorities. In: Wilson LD, Townsend JH, Johnson JD (Eds) Conservation of Mesoamerican Amphibians and Reptiles. Eagle Mountain Publishing, LC, Eagle Mountain, Utah, 238–273.
- Nabhan GP (2000) Cultural dispersal of plants and reptiles to the Midriff Islands of the Sea of Cortés: integrating indigenous human dispersal agents into island biogeography. Journal of the Southwest 42(3): 545–558. https://www.jstor.org/stable/40170213
- Nabhan GP (2002) Cultural dispersal of plants and reptiles. In: Case TJ, Cody ML, Ezcurra E (Eds) A New Island Biogeography of the Sea of Cortés. Oxford University Press, Oxford, 407–416. https://doi.org/10.1093/oso/9780195133462.003.0022
- Ochoa-Ochoa L, Flores-Villela O, García-Vázquez U, Correa-Cano M, Canseco-Márquez L (2006) Hemidactylus mabouia (Geco Casero Tropical) [Online]. CONABIO. http://www.conabio.gob.mx/informacion/gis/?vns=gis_root/biodiv/distpot/reptilia/sauria/hemi_mabugw [Accessed 6 February 2021]
- Ochoa-Ochoa L, Flores-Villela O, Ríos-Muñoz CA, Arroyo-Cabrales J, Martínez-Gordillo M (2017) Mexico’s ambiguous invasive species plan. Science 355(6329): 1033–1033. https://doi.org/10.1126/science.aam9400
- O’Hanlon SJ, Rieux A, Farrer RA, Rosa GM, Waldman B (2018) Recent Asian origin of chytrid fungi causing global amphibian declines. Science 360(6389): 621–627. https://doi.org/10.1126/science.aar1965
- Olson CA, Beard KH, Pitt WC (2012) Biology and impacts of Pacific island invasive species 8. Eleutherodactylus planirostris, the Greenhouse Frog (Anura: Eleutherodactylidae). Science 66(3): 255–270. https://doi.org/10.2984/66.3.1
- Ortiz-Medina JA, Nahuat-Cervera PE, Cabrera-Cen D, Vega-Marcín A, Cedeño-Vázquez JR (2017) Distribution Notes. First records of the invasive Greenhouse Frog, Eleutherodactylus planirostris (Cope, 1862) (Anura: Eleutherodactylidae) in the state of Yucatán, Mexico. Mesoamerican Herpetology 4(1): 191–193.
- Ortiz-Medina JA, Cabrera-Cen DI, Chan-Noh MM, Cedeño-Vázquez JR (2019) First record of the Moorish Gecko, Tarentola mauritanica (Linnaeus, 1758) (Squamata: Phyllodactylidae), in Mexico. Herpetology Notes (12): 971–974. https://www.biotaxa.org/hn/article/view/46810/56653
- Ottley JR, Velázquez-Solis VM (1989) An extant, indigenous tortoise population in Baja California Sur, Mexico, with the description of a new species of Xerobates (Testudines: Testudinidae). The Great Basin Naturalist 49(4): 496–502. https://doi.org/10.5962/bhl.part.22649
- Parham JF, Papenfuss TJ, van Dijk PP, Wilson BS, Marte C, Schettino LR, Simson WB (2013) Genetic introgression and hybridization in Antillean freshwater Turtles (Trachemys) revealed by coalescent analyses of mitochondrial and cloned nuclear markers. Molecular Phylogenetics and Evolution 67(1): 176–187. https://doi.org/10.1016/j.ympev.2013.01.004
- Parham JF, Papenfuss TJ, Buskirk JR, Parra-Olea G, Chen J-Y, Simison WB (2015) Trachemys ornata or not ornata: reassessment of a taxonomic revision for Mexican Trachemys. Proceedings of the California Academy of Science (Series 4) 62(4): 359–367.
- Parker IM, Simberloff D, Lonsdale WM, Goodell K, Wonham M, Kareiva PM, Williamson MH, Von Holle B, Moyle PB, Byers JE, Goldwasser L (1999) Impact: toward a framework for understanding the ecological effects of invaders. Biological Invasions 1(1): 3–19. https://doi.org/10.1023/A:1010034312781
- Pasachnik SA (2013) Growth, reproduction, and diet of Roatán Spiny-tailed Iguanas, Ctenosaura oedirhina, with notes on the status of the species. Herpetological Conservation and Biology 8(1): 191–198.
- Pejchar L, Mooney HA (2009) Invasive species, ecosystem services and human well-being. Trends in Ecology & Evolution 24(9): 497–504. https://doi.org/10.1016/j.tree.2009.03.016
- Peralta-García A, Valdez-Villavicencio JH, Galina-Tessaro P (2014) African clawed frog (Xenopus laevis) in Baja California: a confirmed population and possible ongoing invasion in Mexican watershEds The Southwestern Naturalist 59(3): 431–434. https://doi.org/10.1894/NBF-12.1
- Perrings C, Dalmazzone S, Williamson M (2005) The economics of biological invasions. In: Mooney HA, Mack R, McNeely JA, Neville LE, Schei PJ, Waage JK (Eds) Invasive Alien Species: A New Synthesis. Island Press, Washington, 16–36.
- Peterson TA (2003) Predicting the geography of species invasions via ecological niche modeling. The Quarterly Review of Biology 78(4): 419–433. https://doi.org/10.1086/378926
- Petren K, Case TJ (1997) A phylogenetic analysis of body size evolution and biogeography in chuckwallas (Sauromalus) and other iguanines. Evolution 51(1): 206–219. https://doi.org/10.1111/j.1558-5646.1997.tb02402.x
- Piaggio AJ, Engeman RM, Hopken MW, Humphrey JS, Keacher KL (2014) Detecting an elusive invasive species: a diagnostic PCR to detect Burmese Python in Florida waters and an assessment of persistence of environmental DNA. Molecular Ecology Resources 14(2): 374–380. https://doi.org/10.1111/1755-0998.12180
- Platt SG, Meerman JC, Rainwater TR (1999) Diversity, observations and conservation of the herpetofauna of Turneffe, Lighthouse and Glovers Atolls, Belize. Bulletin British Herpetological Society 66(1): 1–13.
- Platz JE, Clarkson RW, Rorabaugh JC, Hillis DM (1990) Rana berlandieri: recently introduced populations in Arizona and Southeastern California. Copeia 1990(2): 324–333. https://doi.org/10.2307/1446338
- Porras LW, Wilson LD, Schuett GW, Reiserer RS (2013) A taxonomic reevaluation and conservation assessment of the Common Cantil, Agkistrodon bilineatus (Squamata: Viperidae): a race against time. Amphibian & Reptile Conservation 7(1): 48–73.
- Powell R, Conant R, Collins JT (2016) Peterson’s Field Guide to Reptiles and Amphibians of Eastern and Central North America, Fourth Edition. Houghton Mifflin Harcourt Publishing Company, New York, 494 pp.
- Powell R, Maxey SA (1990) Hemidactylus brookii. Catalogue of American Amphibians and Reptiles (CAAR), Society for the Study of Amphibians and Reptiles 493.1–493.3.
- Powell R, Henderson RH, Adler K, Dundee HA (1996) An annotated Check list of West Indian amphibians and reptiles. In: Powell R, Henderson RW (Eds) Contributions to West Indian Herpetology: A Tribute to Albert Schwartz. Society for the Study of Amphibians and Reptiles, Ithaca, 51–93.
- Pyšek P, Richardson DM (2010) Invasive species, environmental change and management, and health. Annual Review of Environment and Resources 35: 25–55. https://doi.org/10.1146/annurev-environ-033009-095548
- Pyšek P, Richardson DM, Pergl J, Jarošík V, Sixtová Z (2008) Geographical and taxonomic biases in invasion ecology. Trends in Ecology & Evolution 23(5): 237–244. https://doi.org/10.1016/j.tree.2008.02.002
- Quijada-Mascareñas A, Canseco-Márquez L (2007) Geographic distribution: Mexico, Nayarit: Gehyra mutilata. Herpetological Review 38(4): e483.
- Ramírez-Bautista A, Hernández-Salinas U, García-Vázquez UO, Leyte-Manrique A, Canseco-Márquez L (2009) Herpetofauna del Valle de México: Diversidad y Conservación. Universidad Autónoma del Estado de Hidalgo (ICBI), CONABIO, México, D. F., 213 pp.
- Ramírez-Bautista A, Hernández-Salinas U, Cruz-Elizalde R, Berriozabal-Islas C, Lara-Tufiño D, Goyenechea Mayer-Goyeneche I, Castillo-Cerón JM (2014) Los Anfibios y Reptiles de Hidalgo, México: Diversidad, Biogeografía y Conservación. Sociedad Herpetológica Mexicana, A.C. Pachuca de Soto, Hidalgo, 387 pp.
- Ramos-Guerra S, Gatica A (2014) Ecología alimentaria de la rana toro Lithobates catesbeianus (Shaw, 1802) en el noroeste de Chihuahua, México. In: Mendoza R, Koleff P (Eds) Especies acuáticas invasoras en México. Comisión Nacional para el Conocimiento y Uso de la Biodiversidad, México, 411–429.
- Rato C, Harris DJ, Carranza S, Machado L, Perera A (2016) The taxonomy of the Tarentola mauritanica species complex (Gekkota: Phyllodactylidae): Bayesian species delimitation supports six candidate species. Molecular Phylogenetics and Evolution 94(A): 271–278. https://doi.org/10.1016/j.ympev.2015.09.008
- Rato C, Harris DJ, Perera A, Carvalho SB, Carretero MA (2015) A combination of divergence and conservatism in the niche evolution of the Moorish Gecko, Tarentola mauritanica (Gekkota: Phyllodactylidae). PLoS ONE 10(5): e0127980. https://doi.org/10.1371/journal.pone.0127980
- Ray DA, Dever JA, Platt SG, Rainwater TR, Finger AG (2004) Low levels of nucleotide diversity in Crocodylus moreletii and evidence of hybridization with C. acutus. Conservation Genetics 5(4): 449–462. https://doi.org/10.1023/B:COGE.0000041024.96928.fe
- Recuero E, Martínez-Solano I, Parra-Olea G, García-París M (2004) Smilisca baudinii (Anura: Hylidae) in Baja California Sur, México. Herpetological Review 35(3): e296.
- Reed RN, Kraus F (2010) Invasive reptiles and amphibians: global perspectives and local solutions. Animal Conservation 13(1): 3–4. https://doi.org/10.1111/j.1469-1795.2010.00409.x
- Reynolds RG, Niemiller ML, Revell LJ (2014) Toward a tree-of-life for the boas and pythons: multilocus species-level phylogeny with unprecedented taxon sampling. Molecular Phylogenetics and Evolution 71(1): 201–213. https://doi.org/10.1016/j.ympev.2013.11.011
- Reynoso F (1990a) Geographic distribution: Mexico, Baja Californias Sur: Gehyra mutilata. Herpetological Review 21(1): e22.
- Reynoso F (1990b) Geographic distriution: Mexico, Baja California Sur: Hemidactylus frenatus. Herpetological Review 21(1): e22.
- Rhodin AGJ, Iverson JB, Bour R, Fritz U, Georges A, Shaffer HB (2017) Turtles of the world: annotated checklist and atlas of taxonomy, synonymy, distribution, and conservation status. In: Rhodin AGJ, Iverson JB, Van Dijk PP, Saumure RA, Buhlmann KA, Prichard PCH, Mittermeier RA (Eds) Conservation Biology of Freshwater Turtles and Tortoises: A Compilation Project of the IUCN/SSC Tortoise and Freshwater Turtle Specialist Group, Chelonian Research Monograph 7: 1–292 https://doi.org/10.3854/crm.7.checklist.atlas.v8.2017
- Richardson DM, Pyšek P, Carlton JT (2011) A compendium of essential concepts and terminology in invasion ecology. In: Ricardson DM (Ed.) Fifty Years of Invasion Ecology: The Legacy of Charles Elton. Blackwell Publishing Ltd., Hoboken, 409–421. https://doi.org/10.1002/9781444329988.ch30
- Robinson MD (1972) Chromosomes, Protein Polymorphism, and Systematics of Insular Chuckwalla Lizards (Genus Sauromalus) in the Gulf of California, Mexico. Ph.D. Dissertation, University of Arizona, Tucson, 71 pp.
- Rocha S, Ineich I, Harris DJ (2009) Cryptic variation and recent bipolar range expansion within the Stumped-Toed Gecko Gehyra mutilata across Indian and Pacific Ocean islands. Contributions to Zoology 78(1): 1–8. https://doi.org/10.1163/18759866-07801001
- Rodda GH, Fritts TH, Clark CS, Gotte SW, Chiszar D (1999) A state of the art trap for the Brown Tree Snake. In: Rodda GH, Sawai Y, Chiszar D, Tanaka H (Eds) Problem Snake Management: the Habu and the Brown Tree Snake. Cornell University Press, Ithaca, New York, 268–305. https://doi.org/10.7591/9781501737688
- Rödder D, Solé M, Böhme W (2008) Predicting the potential distributions of two alien invasive housegeckos (Gekkonidae: Hemidactylus frenatus, Hemidactylus mabouia). North-Western Journal of Zoology 4(2): 236–246.
- Rodriguez D, Cedeño‐Vázquez JR, Forstner MRJ, Densmore LD (2008) Hybridization between Crocodylus acutus and Crocodylus moreletii in the Yucatan Peninsula: II. Evidence from microsatellites. Journal of Experimental Zoology Part A: Ecological and Integrative Physiology 309: 674–686. https://doi.org/10.1002/jez.499
- Roesler H, Bauer AM, Heinicke MP, Greenbaum E, Jackman T (2011) Phylogeny, taxonomy, and zoogeography of the genus Gekko Laurenti, 1768 with the revalidation of G. reevesii Gray, 1831 (Sauria: Gekkonidae). Zootaxa 2989: 1–50. https://doi.org/10.11646/zootaxa.2989.1.1
- Romero-Contreras AT, Ávila-Ramos L (1999) Mesoamérica: historia y reconsideración del concepto. Ciencia Ergo-sum 6: 233–242.
- Rorabaugh JC, King AD (2013) Geographic distribution: Mexico, Sonora, Municipality of Cananea: Apalone spinifera. Herpetological Review 44(1): 104–105.
- Rorabaugh JC, Lemos-Espinal JA (2016) A Field Guide to the Amphibians and Reptiles of Sonora, Mexico. ECO Herpetological Publishing and Distribution, Rodeo, New Mexico, 687 pp.
- Rorabaugh JC, Montoya ES, Gomez-Sapiens MM (2008) Geographic distribution: Mexico, Sonora: Apalone spinifera. Herpetological Review 39(3): e365.
- Rorabaugh JC, Servoss JM (2006) Geographic distribution: Mexico, Sonora. Rana berlandieri. Herpetological Review 37(1): e102.
- Rorabaugh JC, Sredl MJ, Miera V, Drost CA (2002) Continued invasion by an introduced frog (Rana berlandieri): southwestern Arizona, southeastern California, and Río Colorado, México. The Southwestern Naturalist 47(1): 12–20. https://doi.org/10.2307/3672797
- Rösler H, Glaw F (2010) Morphologische variation und taxonomie von Hemidactylus brookii Gray, 1845 und Hemidactylus angulatus Hallowell, 1854 sowie phänotypisch ähnlicher taxa. Spixiana 33(1): 139–160.
- Ruíz-Pérez GA (1996) Claves Preliminares para Reconocer a los Reptiles de Nicaragua. Centro de Derecho Ambiental y Promoción para al Desarrollo, Managua, Nicaragua, 88 pp.
- Sakai AK, Allendorf FW, Holt JS, Lodge DM, Molofsky J, With KA, Baughman S, Cabin RJ, Cohen JE, Ellstrand NC, McCauley DE, O’Neil P, Parker IM, Thompson JN, Weller SG (2001) The population biology of invasive species. Annual Review of Ecology and Systematics 32(1): 305–332. https://doi.org/10.1146/annurev.ecolsys.32.081501.114037
- Salazar-Saavedra M, Loza JC, Fernández M, Martínez-Fonseca J, Dwyer Q, Sunyer J (2015) Chelonoidis carbonarius (Spix, 1824): a member of the Nicaraguan herpetofauna. Mesoamerican Herpetology 2(4): 571–573.
- Sasa M, Chaves G, Porras LW (2010) The Costa Rican Herpetofauna: Conservation status and future perspectives. In: Wilson LD, Townsend JW, Johnson JD (Eds) Conservation of Mesoamerican Amphibians and Reptiles. Eagle Mountain Publishing, LC, Eagle Mountain, Utah, 510–603.
- Saucedo B, Serrano JM, Jacinto-Maldonado M, Leuven RSEW, Rocha García AA, Méndez Bernal A, Gröne A, Van Beurden SJ, Escobedo-Bonilla CM (2019) Pathogen risk analysis for wild amphibian populations following the first report of a Ranavirus outbreak in farmed American Bullfrogs (Lithobates catesbeianus) from northern Mexico. Viruses 11(1): 11–26. https://doi.org/10.1016/j.ympev.2010.06.014
- Savage JM (2002) The Amphibians and Reptiles of Costa Rica: a Herpetofauna Between Two Continents, Between Two Seas. University of Chicago Press, Chicago, 934 pp.
- Savage JM, Bolaños-Vives F (2005) A checklist of the amphibians and reptiles of Costa Rica: additions and nomenclatural revisions. Zootaxa 2005(1): 1–23. https://doi.org/10.11646/zootaxa.2005.1.1
- Schloegel LM, Picco AM, Kilpatrick AM, Davies AJ, Hyatt AD, Daszak P (2009) Magnitude of the US trade in amphibians and presence of Batrachochytrium dendrobatidis and Ranavirus infection in imported North American bullfrogs (Rana catesbeiana). Biological Conservation 142(7): 1420–1426. https://doi.org/10.1016/j.biocon.2009.02.007
- Schmidt KP (1941) The amphibians and reptiles of British Honduras. Field Museum of Natural History Publications in Zoology Series 22: 475–510. https://doi.org/10.5962/bhl.title.2963
- Schwartz A, Thomas R (1975) A Check-list of West Indian Amphibians and Reptiles. Carnegie Museum of Natural History Special Publication No. 1. Pittsburgh, Pennsylvania, 216 pp. https://doi.org/10.5962/bhl.title.123681
- Seidel ME (2002) Taxonomic observations on extant species and subspecies of slider turtles, genus Trachemys. Journal of Herpetology 36(2): 285–292. https://doi.org/10.2307/1566003
- SEMARNAT (2017) Lista de las Especies Exóticas Invasoras para México [Online]. Secretaría de Medio Ambiente y Recursos Naturales. https://www.gob.mx/semarnat/documentos/lista-de-reptiles [Accessed 6 February 2021]
- Serrano-Gómez SS, Guevara-Chumacero LM, Barriga-Sosa IDLA, Ullóa-Arvízu R, González-Guzmán S, Vázquez-Peláez CG (2016) Low levels of genetic diversity in Crocodylus acutus in Oaxaca and Guerrero, Mexico, and molecular-morphological evidence of the presence of C. moreletii. Biochemical Systematics and Ecology 69: 51–59. https://doi.org/10.1016/j.bse.2016.08.005
- Sexton OJ, Brown HM (1977) The reproductive cycle of an iguanid lizard, Anolis sagrei from Belize. Journal of Natural History 11: 241–250. https://doi.org/10.1080/00222937700770171
- Sharp RL, Larson LR, Green GT (2011) Factors influencing public preferences for invasive alien species management. Biological Conservation 144(8): 2097–2104. https://doi.org/10.1016/j.biocon.2011.04.032
- Shaw CE (1946) A new locality for the spiny chuckwalla, Sauromalus hispidus. Copeia 1946(4): 254–254. https://doi.org/10.2307/1438114
- Shine C, Williams N, Gündling L (2000) A guide to designing legal and institutional frameworks on alien invasive species. IUCN, Gland, Switzerland, Cambridge, United Kingdom, and Bonn, Germany, 138 pp.
- Shine R (2015) Reducing the Ecological Impact of Invasive Cane Toads. In: Canning-Clode J (Ed.) Biological Invasions in Changing Ecosystems: Vectors, Ecological Impacts, Management and Predictions. Warsaw: Walter de Gruyter. Warsaw, 301–317. https://doi.org/10.1515/9783110438666-019
- Shine R (2018) Cane Toad Wars. University of California Press, Oakland, 288 pp. https://doi.org/10.1525/9780520967984
- Simberloff D (2005) Non-native species DO threaten the natural environment! Journal of Agricultural and Environmental Ethics. Ethics 18(6): 595–607. https://doi.org/10.1007/s10806-005-2851-0
- Simberloff D (2009) We can eliminate invasions or live with them: successful management projects. In: Langor DW, Sweeney J (Eds) Ecological Impacts of Non-Native Invertebrates and Fungi on Terrestrial Ecosystems. Springer, Dordrecht, 149–157. https://doi.org/10.1007/978-1-4020-9680-8_11
- Slevin JR (1928) The amphibians of western North America. Occasional Papers of the California Academy of Sciences 16: 1–152.
- Smith HM, Burger L (1949) A new subspecies of Anolis sagrei from the Atlantic coast of tropical América. Anales del Instituto de Biología 20: 407–410.
- Smith HM, Grant C (1961) The Mourning Gecko in the Americas. Herpetologica 17(1): e68. https://www.jstor.org/stable/3889726
- Smith HM, Taylor EH (1948) An annotated checklist and key to the amphibians of Mexico. Smithsonian Institution, United States National Museum Bulletin 194, United States Government Printing Office, Washington, 118 pp. https://doi.org/10.5479/si.03629236.194
- Solís J, Wilson L, Townsend J (2014) An updated list of the amphibians and reptiles of Honduras, with comments on their nomenclature. Mesoamerican Herpetology 1(1): 123–144.
- Sosa-Tovar G, García-Padilla E, Wilson LD, Mata-Silva V (2019) Geographic distribution: Mexico, Tamaulipas, Municipality of Tampico: Hemidactylus mabouia. Herpetological Review 50(4): e746.
- Stafford PJ, Meyer JR (2000) A Guide to the Reptiles of Belize. The Natural History Museum, London, Academic Press, London, United Kingdom, 356 pp.
- Stafford PJ, Walker P, Edgar P, Pennj MG (2010) Distribution and conservation of the Herpetofauna of Belize. In: Wilson LD, Townsend JH, Johnson JD (Eds) Conservation of Mesoamerican Amphibians and Reptiles. Eagle Mountain Publishing, LC, Eagle Mountain, Utah, 370–404.
- Standfuss B, Lipovšek G, Fritz U, Vamberger M (2016) Threat or fiction: is the pond slider (Trachemys scripta) really invasive in central Europe? A case study from Slovenia. Conservation Genetics 17(3): 557–563. https://doi.org/10.1007/s10592-015-0805-2
- Stebbins RC (2003) A Field Guide to Western Reptiles and Amphibians. The Peterson Field Guide Series. Houghton Mifflin Company. New York, New York, 533 pp.
- Stejneger L (1904) Herpetology of Porto Rico. Annual Report of the U.S. National Museum. U.S. Government Printing Office, Washington, 551–724.
- Stuart LC (1955) A brief review of the Guatemalan lizards of the genus Anolis. Miscellaneous Publications of the Museum of Zoology University of Michigan (91): 1–31. http://hdl.handle.net/2027.42/56336
- Stuart LC (1963) A checklist of the herpetofauna of Guatemala. Miscellaneous Publications of the Museum of Zoology, University of Michigan (122): 1–148. http://hdl.handle.net/11154/99592
- Suárez-Atilano M, Rojas-Soto O, Parra J, Vázquez-Domínguez E (2017) The role of the environment on the genetic divergence between two Boa imperator lineages. Journal of Biogeography 44(9): 2045–2056. https://doi.org/10.1111/jbi.13006
- Sunyer J, Nicholson KE, Phillips JG, Gubler JA, Obando LA (2013) Lizards (Reptilia: Squamata) of the Corn Islands, Caribbean Nicaragua. Check List 9(6): 1383–1390. https://doi.org/10.15560/9.6.1383
- Sunyer J, Martínez-Fonseca J (2019) Fichas de las especies de anfibios y reptiles que han sido registradas para Nicaragua posterior a HerpetoNica (2015). Revista de Temas Nicaraguenses (139): 36–61.
- Terán-Juárez SA, García-Padilla E, García-Morales LJ (2015) New records and distributional range extensions for amphibians and reptiles from Tamaulipas, México. Mesoamerican Herpetology 2(2): 208–213.
- Terán-Juárez SA, García-Padilla E, Mata-Silva V, Johnson JD, Wilson LD (2016) The herpetofauna of Tamaulipas, Mexico: composition, distribution, and conservation. Mesoamerican Herpetology 3(1): 42–113.
- Tepos-Ramírez M, Acosta-Ramírez A, Jiménez-Velázquez G, Álvarez-Alvarado L (2019) Geographic distribution: Mexico, Querétaro, Municipality of Querétaro: Hemidactylus turcicus. Herpetological Review 50(3): e526.
- Thomas R (1975) The argus group of West Indian Sphaerodactylus (Sauria: Gekkonidae). Herpetologica 31(2): 177–195. https://www.jstor.org/stable/3891650
- Tingley R, Ward-Fear G, Schwarzkopf L, Greenlees MJ, Phillips BL, Brown G, Clulow S, Webb J, Capon R, Sheppard A, Strive T, Tizard M, Shine R (2017) New weapons in the toad toolkit: a review of methods to control and mitigate the biodiversity impacts of invasive cane toads (Rhinella marina). The Quarterly Review of Biology 92(2): 123–149. https://doi.org/10.1086/692167
- Toda M, Takahashi H, Nakagawa N, Sukigara N (2010) Ecology and control of the green anole (Anolis carolinensis), an invasive alien species on the Ogasawara Islands. In: Kawakami K, Okochi I (Eds) Restoring the Oceanic Island Ecosystem. Springer, Tokyo, 145–152. https://doi.org/10.1007/978-4-431-53859-2_22
- Torchin ME, Lafferty KD, Dobson AP, McKenzie VJ, Kuris AM (2003) Introduced species and their missing parasites. Nature 421: 628–630. https://doi.org/10.1038/nature01346
- Townsend CH (1890) Scientific results of explorations by the US Fish Commission steamer Albatross. No. XV. Reptiles from Clarion and Socorro Islands and the Gulf of California, with description of a new species. Proceedings of the United States National Museum 13: 143–144. https://doi.org/10.5479/si.00963801.800.143
- Townsend JH, Wilson LD (2010) Conservation of the Honduran herpetofauna: issues and imperatives. In: Wilson LD, Townsend JH, Johnson JD (Eds) Conservation of Mesoamerican Amphibians and Reptiles. Eagle Mountain Publishing, LC, Eagle Mountain, Utah, 186–705.
- Tyler MJ (2006) Cane toad pheromones. In: Molloy K, Henderson W (Eds) Science of Cane Toad Invasion and Control. Proceedings of the Invasive Animals CRC/CSIRO/Qld NRM&W Cane Toad Workshop. Brisbane, Queensland, 163–164.
- Uetz P, Hosek J, Hallerman J (2019) The Reptile Database. Zoological Museum, Hamburg. http://www.reptile-database.org [Accessed 6 February 2021]
- Upton DE, Murphy RW (1997) Phylogeny of the Side-Blotched Lizards (Phrynosomatidae: Uta) based on mtDNA sequences: support for a midpeninsular seaway in Baja California. Molecular Phylogenetics and Evolution 8(1): 104–113. https://doi.org/10.1006/mpev.1996.0392
- Vázquez-Domínguez E, Suárez-Atilano M, Booth W, González-Baca C, Cuarón AD (2012) Genetic evidence of a recent successful colonization of introduced species on islands: Boa constrictor imperator on Cozumel Island. Biological Invasions 14(10): 2101–2116. https://doi.org/10.1007/s10530-012-0217-x
- Velarde E, Hollingsworth B, Rebman J (2008) Geographic distribution: Mexico, Baja California: Sauromalus hispidus. Herpetological Review 39(3): e368.
- van Dijk PP, Hammerson G, Vazquez Diaz J, Quintero Diaz GE, Santos G, Flores-Villela O (2007) Kinosternon hirtipes (errata version published in 2016). The IUCN Red List of Threatened Species 2007: e.T63670A97381507, [Online]. [Accessed 6 February 2021]
- van Wilgen NJ, Roura-Pascual N, Richardson DM (2009) A quantitative climate-match score for risk-assessment screening of reptile and amphibian introductions. Environmental Management 44(3): 590–607. https://doi.org/10.1007/s00267-009-9311-y
- Vertnet.org (2018) Vertnet.org http://vertnet.org/ [Accessed 6 February 2021]
- Villa J, Wilson LD, Johnson JD (1988) Middle American Herpetology: A Bibliographic Checklist. University of Missouri Press, Columbia, 131 pp.
- Vitousek PM, D’Antonio CM, Loope LL, Rejmanek M, Westbrooks R (1997) Introduced species: a significant component of human-caused global change. New Zealand Journal of Ecology 21(1): 1–16. https://www.jstor.org/stable/24054520
- Vitt LJ, Caldwell JP (2014) Herpetology: An Introductory Biology of Amphibians and Reptiles, Academic Press, San Diego, California, 691 pp. https://doi.org/10.1016/B978-0-12-386919-7.00002-2
- Wallach V (2009) Ramphotyphlops braminus (Daudin): a synopsis of morphology, taxonomy, nomenclature and distribution (Serpentes: Typhlopidae). Hamadryad 34(1): 34–61.
- Wallach V, Williams KL, Boundy J (2014) Snakes of the World: A Catalogue of Living and Extinct Species, CRC Press, Boca Raton, Florida, 1237 pp. https://doi.org/10.1201/b16901
- Walsh JR, Carpenter SR, Vander Zanden MJ (2016) Invasive species triggers a massive loss of ecosystem services through a trophic cascade. Proceedings of the National Academy of Sciences 113(15): 4081–4085. https://doi.org/10.1073/pnas.1600366113
- Webb RG (1973) Trionyx atra Webb and Legler: Black Softshell Turtle. Catalogue of American Amphibians and Reptiles (CAAR): 137.1.
- Weiss AJ, Hedges SB (2007) Molecular phylogeny and biogeography of the Antillean geckos Phyllodactylus wirshingi, Tarentola americana, and Hemidactylus haitianus (Reptilia, Squamata). Molecular Phylogenetics and Evolution 45(1): 409–416. https://doi.org/10.1016/j.ympev.2007.01.006
- Weldon C, du Preez LH, Hyatt AD, Muller R, Speare R (2004) Origin of the amphibian chytrid fungus. Emerging Infectious Diseases 10(12): 2100–2105. https://doi.org/10.3201/eid1012.030804
- Weterings R, Vetter KC (2017) Invasive house geckos (Hemidactylus spp.): their current, potential and future distribution. Current Zoology 64(5): 559–573. https://doi.org/10.1093/cz/zox052
- Williams CF, Britton JR, Turnbull JF (2013) A risk assessment for managing non-native parasites. Biological Invasions 15(6): 1273–1286. https://doi.org/10.1007/s10530-012-0364-0
- Wilson LD, Johnson JD (2010) Distributional patterns of the herpetofauna of Mesoamerica, a biodiversity hotspot. In: Conservation of Mesoamerican Amphibians and Reptiles. Wilson LD, Townsend JH, Johnson JD (Eds) Eagle Mountain Publishing, LC, Eagle Mountain, Utah, 30–235.
- Wilson LD, Mata-Silva V, Johnson JD (2013a) A conservation reassessment of the Reptiles of Mexico based on the EVS measure. Amphibian & Reptile Conservation 7(1): 2–46.
- Wilson LD, Johnson JD, Mata-Silva V (2013b) A conservation reassessment of the amphibians of Mexico based on the EVS measure. Amphibian & Reptile Conservation 7(1): 98–127.
- Witmer GW, Snow NP, Moulton RS (2015) Efficacy of potential chemical control compounds for removing invasive American bullfrogs (Rana catesbeiana). SpringerPlus 4(1): 1–5. https://doi.org/10.1186/s40064-015-1319-6
- Woolrich-Peña GA, García-Padilla E, DeSantis DL, Johnson DJ, Mata-Silva V, Wilson LD (2017) The herpetofauna of Puebla, Mexico: composition, distribution, and conservation status. Mesoamerican Herpetology 4(4): 2–95.
- Woolrich-Piña GA, Ponce-Campos P, Loc-Barragán J, Ramírez-Silva JP, Mata-Silva V, Johnson JD, García-Padilla E, Wilson LD (2016) The herpetofauna of Nayarit, Mexico: composition, distribution, and conservation. Mesoamerican Herpetology 3(2): 376–448.
- Yáñez-Arenas C, Díaz-Gamboa L, Rodríguez-Pérez A, Salmerón-Flores A, Patrón-Rivero C (2016) Análisis de riesgo de tortugas con potencial invasor en México. In: CONABIO and PNUD (Eds) GEF 0089333 “Aumentar las Capacidades Nacionales para el Manejo de las Especies Exóticas Invasoras (EEI) a través de la Implementación de la Estrategia Nacional de EEI, ” Ciudad de México: Unidad Académica de Yucatan-UNAM, 84 pp.
- Young AM, Larson BMH (2011) Clarifying debates in invasion biology: a survey of invasion biologists. Environmental Research 111(2): 893–898. https://doi.org/10.1016/j.envres.2011.06.006
- Zaldı́var-Riverón A, León-Regagnon V, Nieto-Montes de Oca A (2004) Phylogeny of the Mexican coastal leopard frogs of the Rana berlandieri group based on mtDNA sequences. Molecular Phylogenetics and Evolution 30(1): 38–49. https://doi.org/10.1016/S1055-7903(03)00141-6
- Zamora-Abrego JG, García-Vázquez UO, Canseco-Márquez L, Nieto-Montes de Oca A (2006) Geographic Distribution: Mexico, Veracruz, Municipality of Minatitlán: Anolis sagrei. Herpetological Review 37(4): e493.
- Zavaleta ES, Hobbs RJ, Mooney HA (2001) Viewing invasive species removal in a whole-ecosystem context. Trends in Ecology & Evolution 16(8): 454–459. https://doi.org/10.1016/S0169-5347(01)02194-2
Supplementary material
Multipoint shapefiles, occurrence records for introduced and translocated reptiles and amphibians in Mexico and Central American Countries
Data type: species data