





Citation: Hanamura Y, Mantiri ROSE, Ohtsuka S (2014) A new species of Gastrosaccus Norman, 1868 (Mysida, Mysidae, Gastrosaccinae) from a sandy shore of Indonesia. ZooKeys 438: 17–32. doi: 10.3897/zookeys.438.8031
A new species of the mysid crustacean genus Gastrosaccus Norman, 1868 (Mysida, Mysidae, Gastrosaccinae) is reported from a sandy shore of Lombok Island, Indonesia. These specimens resemble G. sorrentoensis Wooldridge & McLachlan, 1986 and G. yuyu Bamber and Morton, 2012 by the possession of an articulated process on the fifth abdominal somite together with a fringe of spine-like filaments on the posterodorsal margin of the carapace. The Lombok population differs from the known congeners by having comparatively fewer numbers of carpopropod segments on the endopod of the third to eighth thoracic limbs and the conformation in the telson and in the male third pleopod. Hence, G. lombokiensis sp. n. is proposed herein as a third species of “G. sorrentoensis” species group.
Mysid shrimp, taxonomy, new species, Lombok Island, Indonesia
The mysid genus Gastrosaccus Norman, 1868 (Crustacea, Mysida) currently comprises 24 species (
During a recent survey of coastal crustaceans in Indonesia, which aimed in part to clarify the nature of planktonic crustacean fisheries in the country (
The sampling site, Padak Guar beach is located on the north-east coast of Lombok Island, Indonesia. The beach substratum is predominately fine sand particles.
The material used in this paper was collected by the second author (RM) using a push net (2 m in mouth width, 3 m long, and 1 mm mesh openings), which was towed along the beach on the evening of June 6th, 2010. Mysid specimens were fixed in 70−99% ethanol immediately upon collection and preserved prior to analysis.
Body size of mysids (TL: mm) was measured between the distance from the apex of the rostrum to the posterior end of the telson excluding the apical spine-like setae. Description is based on adult specimens unless stated otherwise.
The type specimens are deposited in the National Museum of Nature and Science at Tsukuba, Japan (NSMT).
Mysis spinifera Goës, 1864; by monotypy.
Male (TL ca. 7.5 mm) (NSMT−Cr 22940), sandy beach, Padak Guar (08°25.665'S, 116°42.561'E), Lombok Island, push net, evening time (17:00), 6 June 2010, coll. R. Mantiri.
11 males (TL 4.0−ca. 7.5 mm), 3 females (TL 4.0−ca.7.5 mm), 3 juvs (TL 2.8–3.5 mm) (NSMT−Cr 22941), data same as for holotype.
Rostrum produced into sub-triangular fig. Posterodorsal edge of carapace bearing 7−12, commonly 9−11, spine-like filaments between dorsolateral slits and normally further 5−10 feeble denticulations/undulations on each side of posterolateral lobe posterior to slit. Fifth somite of abdomen with articulated triangular process at posteromedian end; pleuron of female first somite greatly expanded, fully covering marsupium. Telson typical of Gastrosaccinae form, with apical cleft occupying 1/8−1/6 length of telson, armed laterally with 8 or 9 robust spine-like setae, including posteriormost one. Uropod with exopod shorter than endopod, armed laterally with 13 or 14 spine-like setae; endopod with 6 or 7 spine-like setae on mesial margin. Labrum with single anteromedian tooth and additional smaller spines absent. Third pleopod of male with endopod multi-articulated; exopod greatly elongated, extending well beyond sixth abdominal somite, comprising 4 major segments, and basal segment without distinct sub-articles, distal segment 0.65–0.7 times length of penultimate one, armed with short sub-terminal seta and moderately long 2 terminal setae. First pleopod of female bi-lobed, second to fifth pleopods rudimentary, unsegmented lobe.
Male. Body (Fig. 1) moderately robust.
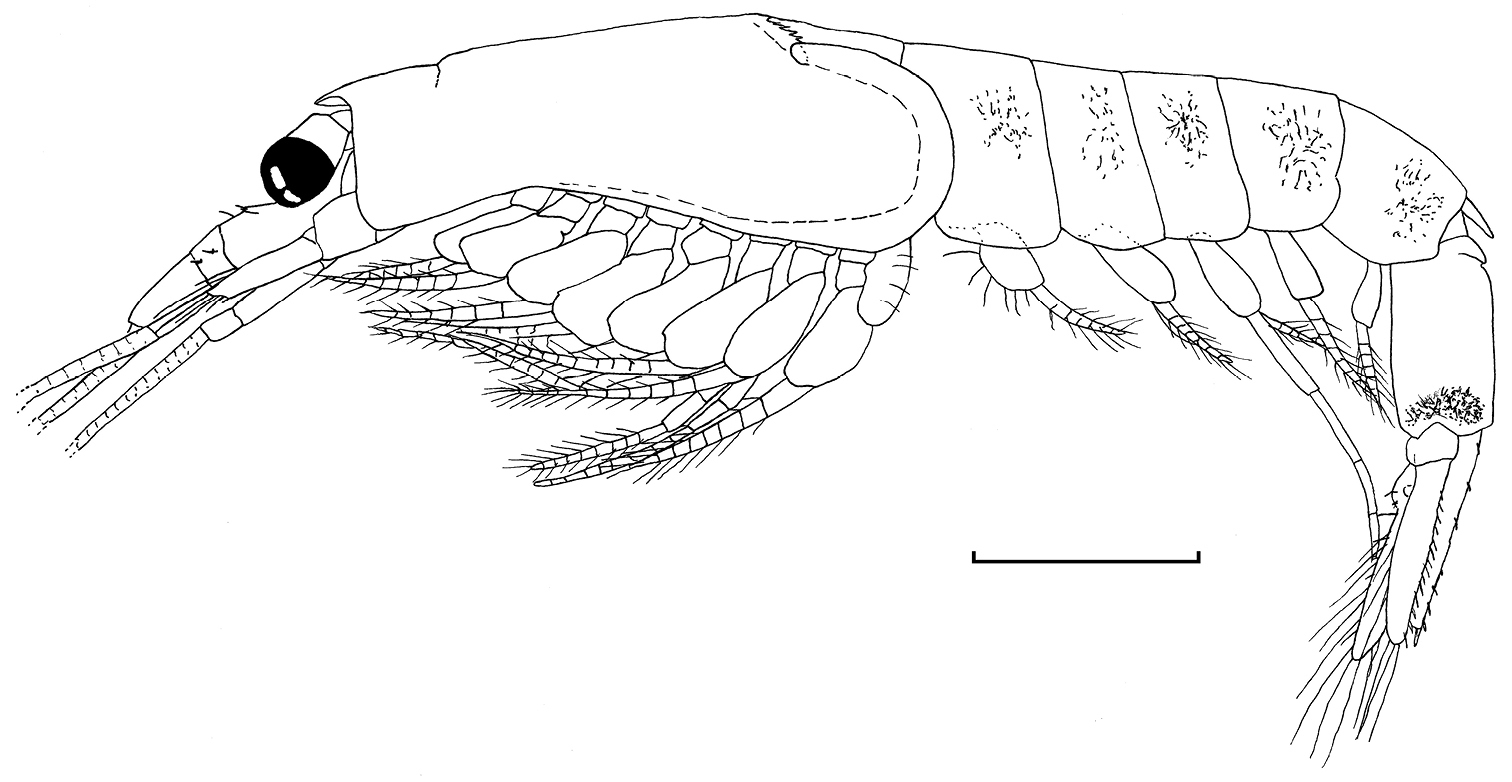
Gastrosaccus lombokiensis sp. n., entire body of paratype male (TL 6.9 mm) in lateral view. Scale = 1 mm.
Carapace (Figs 1, 2a, b) produced into sub-triangular rostrum with sub-acute apex; posterior dorsal margin deeply excavate, leaving exposed last thoracic somite, dorsolateral part of emargination forming slit with posterior lobe overlapping onto anterior one; lateral wing of carapace well developed, extending to anterior 1/3 of first abdominal pleuron; anterolateral part rounded; cervical sulcus marked dorsally anterior to anterior 1/3; posterior dorsal edge between dorsal slits bearing 7−12, most commonly 9−11, spine-like filaments, and further 5−10 feeble denticles/undulations on each margin posterior to slit (often hard to define exact numbers due to its poor development particularly in posterior part).
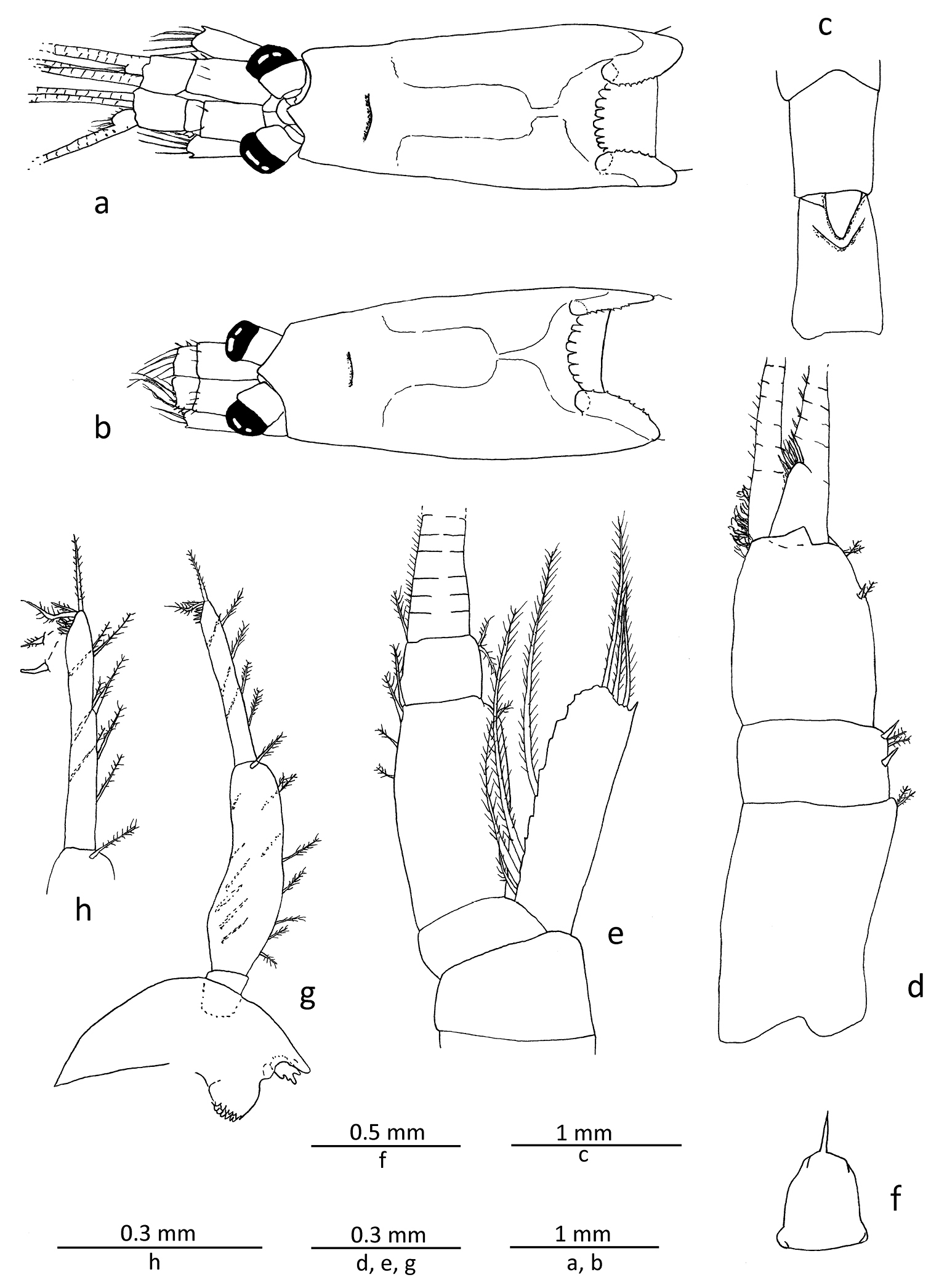
Gastrosaccus lombokiensis sp. n., paratype male (TL ca. 7.5 mm) (a, c−h) and paratype female (TL ca. 7.5 mm) (b): a carapace and cephalic appendages in male, dorsal view b same in female c fifth and sixth abdominal somites, dorsal view d right antennule, dorsal view e right antenna, dorsal view f labrum, ventral view g right mandible, external view h same, distal article of mandibular palp enlarged.
Abdomen (Figs 1, 2c) with anterior 4 somites rounded dorsally, sub-equal in length; fifth somite rounded dorsally, 1.3−1.4 times longer than precedents, with remarkable articulated sub-triangular process on posterodorsal margin; sixth somite very slightly longer than fifth, with transverse carinate ridge on anterodorsal part; brown to dark-brown melanophores on lateral surface of first to fifth somites and also in posterolateral part of sixth somite in ethanol preserved specimens.
Antennule (Fig. 2d) with basal segment longest, very slightly shorter than length of 2 anterior segments combined; second segment shortest, about 0.55 times longer than wide, with 2 spine-like setae, one at mid-length and another one at anterior end of lateral margin; third segment about twice the length of second, with short spine-like seta at distal 1/3 of lateral margin; lateral flagellum slightly widened at mesiobasal part, forming somewhat small male lobe bearing moderately long sensory setae; mesial flagellum more slender than lateral one. Antenna (Fig. 2e) with scale proportionately short, barely reaching or falling slightly short of anterior end of second antennular segment, slightly more than 3 times as long as wide, lateral margin smooth, distolateral spine not extending beyond anterior margin of lamella; antennal peduncle long, extending slightly beyond scale, penultimate segment elongated, approximately 3 times as long as distal one.
Labrum (Fig. 2f) with single sharp anteromedian tooth, additional teeth absent.
Mandibular palp (Fig. 2g, h) composed of 3 segments, distal segment about 0.8 times length of penultimate, with 2 long terminal setae, of which proximal one curved around its mid-length, often forming sub-chelate structure with terminal segment, and another seta moderately long at distal end, and also about 3 short spine-like setae as well as several short obtuse setae, all confined to distal part, forming comb-like structure. Mouthparts (maxillule, maxilla and second thoracopod) general form of genus (Fig. 3a, b, e). First thoracopod (Fig. 3c, d) with basal fig of exopod about 1.6 times length of its width.
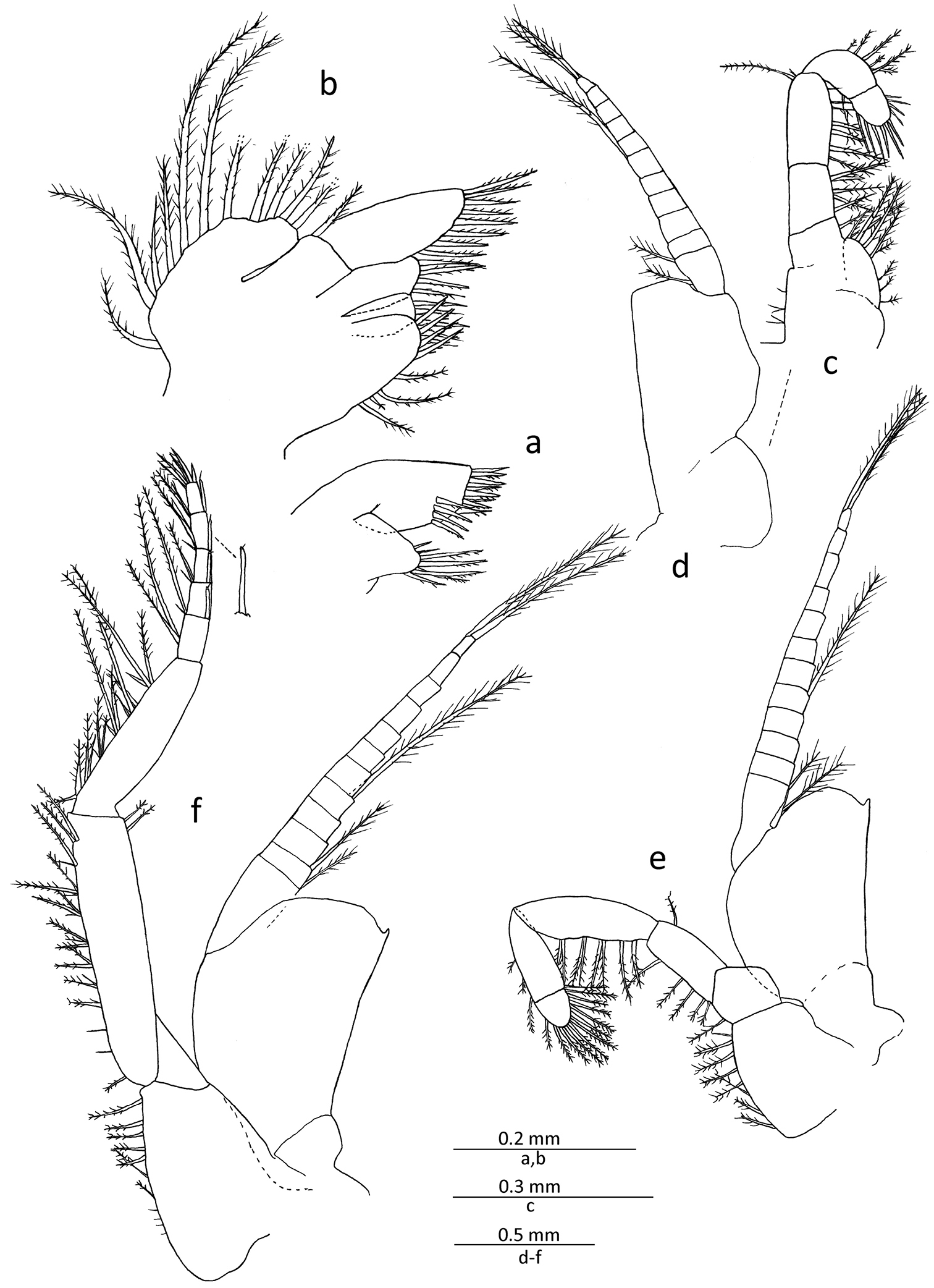
Gastrosaccus lombokiensis sp. n., paratype male (TL ca. 7.5 mm): a right maxillule, external view b right maxilla, external view c right endopod of first thoracic limb, external view d right exopod of first thoracic limb, external view e right second thoracic limb, internal view f right third thoracic limb, posterior view.
Third to eighth thoracopods (Figs 3f, 4a, b) similar in basic structure; exopods with basal fig smooth on lateral margin, and small process at distolateral corner except for eighth limb, and also with 11–14 segmented flagellar part; third to eighth endopods with carpopropodus composed of 5−9 segments, progressively increasing in numbers posteriorly, in which eighth endopod is divided into 8 or 9 segments.
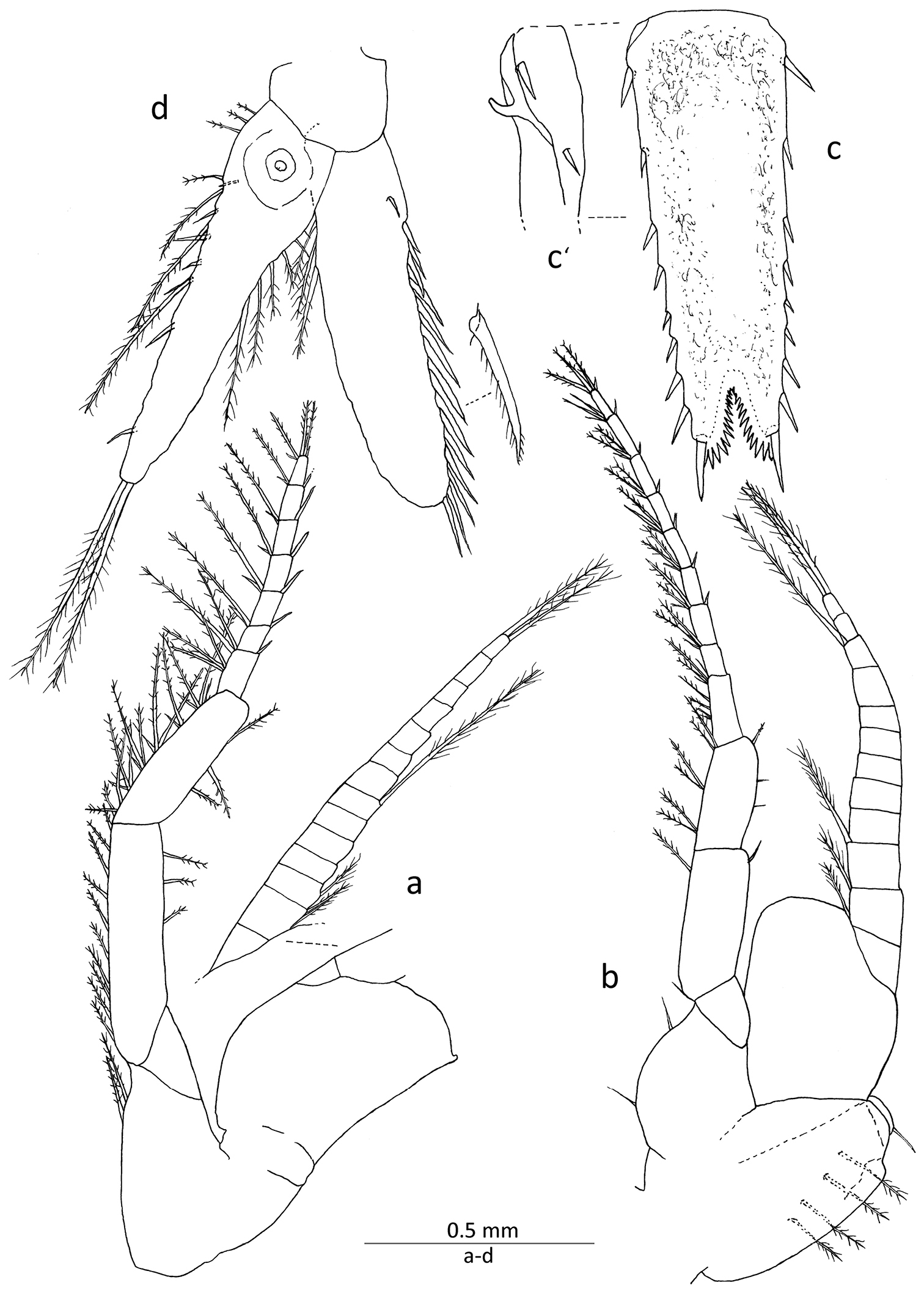
Gastrosaccus lombokiensis sp. n., paratype male (TL ca. 7.5 mm): a right sixth thoracic limb, posterior view b left eighth thoracic limb and penis, anterior view c telson, dorsal view c’ basal part of telson, lateral view d right uropod, dorsal view.
Penis (Fig. 4b) about 1.7 times as long as wide, with short terminal seta directed posteriorly and 4 or 5 long setae along lateral surface.
All pleopods of male biramous (Figs 1, 5a–e). First pleopod with sympod bearing several long setae along anterior margin; endopod rudimentary, unsegmented lobe possessing several marginal setae distally; exopod articulated to about 8 segments. Second pleopod with endopod articulated to about 7 segments; exopod articulated to about 8 segments, slightly longer than endopod. Third pleopod with endopod articulated to 6 or 7 segments; exopod greatly elongated, extending beyond posterior end of abdomen fully by distal segment, composed of 4 major segments, basal segment markedly compressed and broader than distal series of segments, sub-equal length or very slightly longer than second one, without distinct sub-segments but with slight indication of 2 incipient articulations, second segment tubular and unarmed, penultimate segment bearing short spine-like sub-distal seta, distal segment shortest, 0.65−0.70 times length of penultimate, armed distally with 2 moderately long spine-like setae with subsidiary setules on its distal half. Fourth and fifth pleopods similar in basic form, endopod unsegmented, bearing a few setae on distal margin; exopod articulated to about 8 segments.
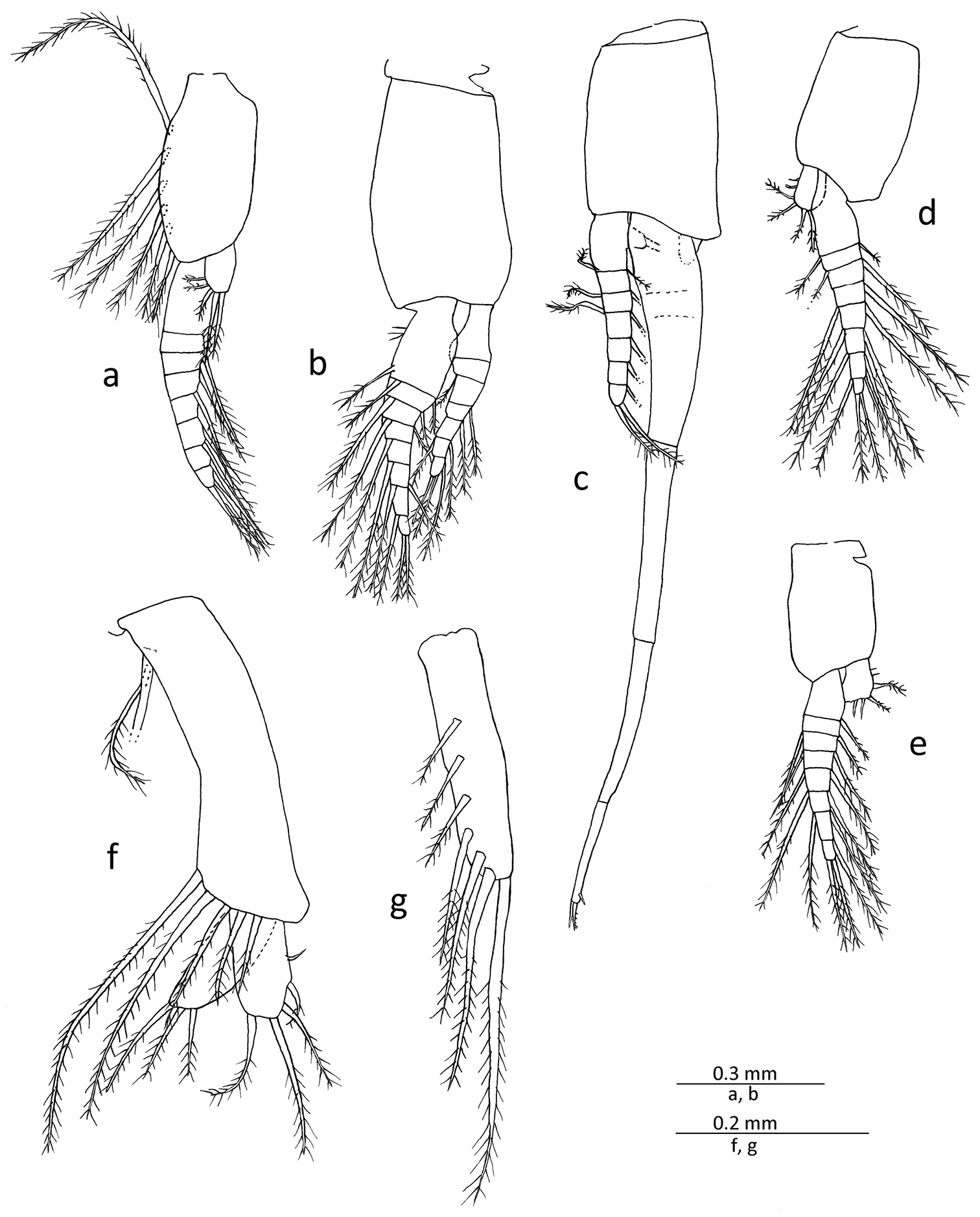
Gastrosaccus lombokiensis sp. n., paratype male (TL ca. 7.5 mm) (a−e) and paratype female (TL ca. 7.5 mm) (f, g): a right male first pleopod, mesial view b right male second pleopod, mesial view c right male third pleopod, lateral view d right male fourth pleopod, lateral view e right male fifth pleopod, mesial view f right first female pleopod, mesial view g right female third pleopod, mesial view.
Uropod (Fig. 4d) with exopod shorter than endopod, lateral margin armed with 13 or 14 robust spine-like setae, length increasing posteriorly, each spine-like seta with subsidiary setules on posterior margin and also short terminal protrusion; endopod extending beyond telson and reaching as far as end of distal telson setae, commonly bearing 7, occasionally 6, spine-like setae along entire mesial margin, showing general tendency towards increased setae length distally.
Telson (Fig. 4c) 1.1–1.2 times as long as sixth abdominal somite and 2.6−2.8 times as long as wide, apical cleft occupying about 1/7 length of telson, bearing approximately 15 dentations along each mesial margin; lateral margin with 8 or 9 spine-like setae of varying length, including longest apical seta, penultimate seta distinctly shorter than apical and located noticeably posterior to level of anterior edge of apical cleft, proximal lateral seta sub-equal in length to penultimate one, posterior series of lateral setae spaced equidistantly and unarmed gap between apical and penultimate setae subequal in distance with those of others; procurved sharp process (Fig. 4c’) present at proximal part of ventral side. Melanophores widely distiributed on dorsal surface of telson but not along mesiodorsal axis and often distal part of apical lobes in both sexes.
Female. Sexual dimorphism as common for genus.
Pleuron of first abdominal somite greatly expanded, fully covering marsupium.
Antennule (Fig. 2b) might differ slightly from that of male but is damaged in the present material and not dissected.
First pleopod (Fig. 5f) biramous, with unsegmented endopod and exopod, sub-equal in length and markedly shorter than sympod, exopod and endopod each with several long plumose setae on distal part; sympod with a few long setae near anterobasal part and further several long setae along anterior distal margin. Second to fifth pleopods (Fig. 5g) similar in shape and size, all rudimentary, unarticulated rod-shaped lobe, bearing several long setae on lateral surface and also on distal margin.
The name “lombokiensis” refers to the type locality, Lombok Island.
This species is remarkable among the members of Gastrosaccus in having an articulated process at the posterodorsal end of the fifth abdominal somite throughout its life stages, coupled with a fringe of spine-like filaments on the posterodorsal margin of the carapace. The combination of these intriguing characters is shared by Gastrosaccus sorrentoensis Wooldridge & McLachlan, 1986 from Western Australia (see also
Specimens from Lombok Island have the carpopropodus of the thoracic endopods comprising fewer numbers of segments, and at most 8 or 9 segments in the eighth limb compared to 13 or more segments in the previously known species (Table 1). There is a slight possibility that the paucity in the segment counts in Lombok specimens is due to comparatively smaller body sizes, thus showing a condition of younger stages. Although ovigerous females were not available in our collection, the largest female has a well-developed first abdominal pleuron, which is fully covering the marsupium lamellae. Also larger males (> ca. 6.5 mm) were considered as adult because there third pleopods are fully developed, extending well beyond the posterior margin of the abdomen at least by length of distal article. Furthermore, the posterior part of the vas deferens of several males exhibited whitish colour just above the ejaculatory duct, common in mature males.
Comparison of the “Gastrosaccus sorrentoensis Wooldridge & McLachlans, 1986” species group.
| Item/species | Gastrosaccus sorrentoensis Wooldridge & McLachlan, 1986 | Gastrosaccus yuyu Bamber & Morton, 2012 | Gastrosaccus lombokiensis sp. n. |
|---|---|---|---|
| Rostrum | wide triangular protrusion with sub-acute apex | tongue-like protrusion with widely round apex | sub-triangular protrusion with sub-acute apex |
| Posterior margin of carapace | about 25 spine-like filaments between dorsolateral slits and 14–25 filaments or undulations on dorsolateral edge posterior to slits (fewer number of filaments in smaller specimens) | 6–12 spine-like filaments between dorsolateral slits and 6–12 filaments on dorsolateral edge posterior to slits | 7–12, commonly 9–11, spine-like filaments between dorsolateral slits and 5–10 very feable denticles or undulations on dorsolateral edge posterior to slits |
| Mandibular palp | distal segment 0.8 times length of penultimate, terminally with long slender seta and long robust undulate seta directed laterally | distal segment 0.9 times length of penultimate, terminally with 2 slender setae not distinctly directed laterally | distal segment 0.8 times length of penultimate, terminally with long slender seta, long undulate seta directed laterally and one moderately long seta |
| First thoracic limb | basal fig of exopod about twice length of width | basal fig of exopod about 1.5 times length of width | basal fig of exopod about 1.6 times length of width |
| Third to eighth thoracic limbs | carpopropodus of endopod with 8–16 segments, and 14–16 on eighth | carpopropodus of endopod with 12–14 segments, and probably 13 or 14 on eighth | carpopropodus of endopod with 5–9 segments, and 8 or 9 on eighth |
| Female first pleopod | endopod slightly longer than exopod; sympod about 4 times length of width | endopod distinctly longer than exopod; sympod about 4 times length of width | endopod slightly longer than exopod; sympod about 3 times length of width |
| Male third pleopodal exopod | basal segment noticeably longer than second, divided into distinct 3 short sub-segments; distal segment 0.75 times length of penultimate | basal segment noticeably longer than second, divided into 2 or 3 sub-segments; distal segment 0.8 times length of penultimate | basal segment not divided into distinct sub-segments but with slight sign of possible 2 indicipent segmentations; distal segment 0.65–0.70 times length of penultimate |
| Uropodal endopod | 5 or 6 irregularly spaced mesial setae along entire margin | 7 or 8 irregularly spaced mesial setae confined to basal 2/3 | usually 7, occasionally 6, irregularly spaced mesial setae along entire margin |
| Telson | sharp process present at proximalventral part; about 2.5 times as long as wide; apical cleft 1/6 length of telson; 7 or 8 robust lateral setae, penultimate seta arising around level of anterior end of apical cleft, apical spine distinctly longer than penultimate | sharp process absent at proximal ventral part; about 2.8 times as long as wide, apical cleft 1/8 length of telson; 10 robust lateral setae, penultimate seta arising anterior to anterior end of apical cleft, apical spine very slightly longer than penultimate | sharp procurved process present at proximal ventral part, 2.6–2.8 times as long as wide, apical cleft 1/8–1/7 length of telson; 8 or 9 robust lateral setae, penultimate seta arising distinctly posterior to anterior end of apical cleft, apical spine distinctly longer than penultimate |
| Body size | male 7.0–9.3 mm, female 8.5–11.3 mm | male holotype 8.4 mm, female 9–13 mm | male ca. 6.5–7.5 mm, female ca. 7.5 mm |
| Occurrence | surf zone of sandy beach, Perth, Western Australia | surf zone of sandy beach, Java, Indonesia | surf zone of sandy beach, Lombok, Indonesia |
| Data source | present study |
The male third pleopodal exopod of this group basically has four major segments. In Gastrosaccus sorrentoensis and Gastrosaccus yuyu, the basal article is divided further into two or three short sub-segments instead of an entire basal article, without distinct short sub-segments. In the Lombok population, we do not find distinct short sub-articulations. However, a slight indication of incipient sub-segmentations was observed in the internal muscular part; but an associated segmented exoskeleton could not be found. Also the shape of the telson is noticeably different among the three populations and these features are regarded as size independent.
Compared to those of Gastrosaccus sorrentoensis, the Lombok population tended to have larger number of spine-like setae in the uropodal endopod and telson despite their smaller body size. The female telson of Gastrosaccus sorrentoensis bears a pair of pigmented spots at the inner side of the fifth lateral setae (
According to
Our observation suggested that Gastrosaccus lombokensis starts to develop external secondary sexual characteristics at a size around TL 3.5−4 mm in both sexes.
This species is known only from the sandy shore of Lombok Island, Indonesia.
The possession of an articulated process on the fifth abdominal somite together with spine-like filaments on the carapace is a remarkable feature among the 25 species of Gastrosaccus known to date, although the latter character is shared by several species of Gastrosaccus as well as Eurobowmaniella Murano, 1995 (see
Iiella kojimaensis (Nakazawa, 1910) has been believed to possess a posterodorsal process on the fifth somite (
Similarly, Eurobowmaniella simulans (Tattersall, 1915) has been reported to bear a mid-dorsal apophysis on the fifth abdominal somite in its early life stage. We have re-examined specimens of Eurobowmaniella simulans (TL 2.3−7.1 mm) collected from the northern Malacca Strait, but have failed to observe an articulated process, even in the smallest specimens. However, an anterodorsal ridge of the sixth abdominal somite is present throughout the observed size ranges (see also
The East Australian species Haplostylus brisbanensis (Băcescu & Udrescu, 1982) appears to have the same type of articulated process that is found in Chlamydopleon and the “Gastrosaccus sorrentoensis” species group. Haplostylus brisbanensis, however, has no spine-like filaments on the carapace and the endopod of the third male pleopod is reduced to a non-articulated lobe, a typical form for Haplostylus (see
The character distribution among the genera of Gastrosaccus affinities is an admixture of character states (Table 2). The articulated abdominal process is shared by small members of this animal group that are found in the geographically isolated Pacific and Atlantic coasts of American continents and Indonesia-Western Australian coasts. This remarkable morphological structure is most probable to have been acquired independently in different lineages rather than it is regarded as a plesiomorphic character and lost in the majority of gastrosaccini descents, although its phylogenetic significance is still not very clear. The taxonomic position of the Indonesia–Australian species of Gastrosaccus (and also Haplostylus brisbanensis) that bear the articulated abdominal process would be a subject of future consideration.
Comparison between Gastrosaccus and related genera.
| Item/genera | Archaeomysis Czerniavsky, 1882 | Chlamydopleon Ortmann, 1893 | Coifmanniella Heard & Price, 2006 | Eurobowmaniella Murano, 1995 | Gastrosaccus Norman, 1892 | Haplostylus Kossman, 1888 | Iiella Băcescu, 1968 |
|---|---|---|---|---|---|---|---|
| Posterior dorsal margin of carapcace | no median concavity; no fringe of spine-like filaments | median concavity and protruded median lobes; no fringe of spine-like filaments but with minute undulations | median concavity and protruded medianlobes; no fringe of spine-like filaments but minute undulations | no median concavity; fringe of spine-like filaments | median concavity and protruded median lobes present/absent; fringe of spine-like filaments present or absent | median concavity and protruded median lobes present/absent; no fringe of spine-like filaments | no median concavity; no fringe of spine-like filaments |
| Labrum | single long anteromedian spine | long anteromedian and several additional smaller spines | long anteromedian and several additional smaller spines | long anteromedian and several additional smaller spines | single long anteromedian spine | single long anteromedian spine | long anteromedian and several additional smaller spines |
| Abdomen (fifth somite) | no movable process at posterodorsal margin | movable process present at posterodorsal margin | no movable process at posterodorsal margin | no movable process at posterodorsal margin | no movable process at posterodorsal margin (except Gastrosaccus sorrentoensis and its closest affinities) | no movable process at posterodorsal margin (except H. brisbanensis) | no movable process at posterodorsal margin |
| Male second pleopod | exopod multi-segments; endopod multi-segments | exopod multi-segments; endopod uni-segment | exopod multi-segments; endopod uni-segment | exopod multi-segments; endopod multi-segments | exopod multi-segments; endopod multi-segments | exopod multi-segments; endopod uni-/multi-segments | exopod multi-segments; endopod multi-segments |
| Male third pleopod | exopod styliform; endopod multi-segment | exopod greatly complicated; endopod uni-segment |
exopod greatly complicated; endopod uni-segment |
exopod moderately modified; endopod multi-segments |
exopod styliform; endopod multi-segments |
exopod styliform; endopod uni-segment |
exopod styliform; endopod multi-segments |
| Female pleopods | first pleopod biramous; second-fifth pleopods with rudimentary exopod |
first pleopod biramous; second-fifth pleopods rudimentary rod-shaped lobe |
first pleopod biramous; second-fifth pleopods rudimentary rod-shaped lobe |
first pleopod biramous; second-fifth pleopods rudimentary rod-shaped lobe |
first pleopod biramous; second-fifth pleopods rudimentary rod-shaped lobe |
first pleopod biramous; second-fifth pleopods rudimentary rod-shaped lobe |
first pleopod uniramous; second-fifth pleopods rudimentary rod-shaped lobe |
| Geographical range | North Pacific, from Hong Kong to California, through subarctic Pacific islands | tropical and subtropical coasts of East Pacific as well as West Atlantic | tropical coasts of East Pacific and tropical and subtropical coasts of West Atlantic | Northeastern Indian Ocean, from India to northwestern Malay Peninsula | tenperate to tropical waters of East Atlantic through Indian Ocean to Australia and New Zealand | Mediterranean Sea through Indian Ocean to western Pacific | Northwestern Pacific, from Singapore to Japan and Korea |
Data sources:
The finding of Gastrosaccus lombokiensis, as a third species of the “Gastrosaccus sorrentoensis” group, implies possible further diversification of this species group in the eastern Indian Ocean. Despite a rather intensive survey of the beach mysids across the South East Asia, other species of gastrosaccini mysids could not be found except for Eurobowmaniella simulans, which was sampled in sandy beaches of Langkawi Island, off the north-east coast of Peninsular Malaysia (
| 1 | Sharp projection absent at proximal ventral part of telson, apical telson seta very slightly longer than penultimate one | Gastrosaccus yuyu Bamber & Morton, 2012 |
| – | Sharp projection present at proximal ventral part of telson, apical telson seta distinctly longer than penultimate one | 2 |
| 2 | Posteromedian margin of carapace with as many as 25 spine-like filaments; endopod of eighth thoracic limb composed of 14−16 segments; basal article of exopod of male third pleopod divided into 2 or 3 sub-segments; telson with pigmented spots at inner side of fifth lateral setae in female | Gastrosaccus sorrentoensis Wooldridge & McLachalan, 1998 |
| – | Posteromedian margin of carapace with less than 12 spine-like filaments; endopod of eighth thoracic limb composed of 8 or 9 segments; basal article of exopod of male third pleopod without distinct sub-segments; telson with melanophore widely spread along margins in both sexes | Gastrosaccus lombokiensis sp. n. |
We are grateful to Dr Masaaki Murano for reading and valuable suggestions on the earier version of the masuscript. We also thank Prof Ephrime B. Metillo of the Mindanao State University, Iligan City, the Philippines, for his review of the manuscript. Thanks are also extended to Ms Hiroko Sato for technical assistance. The MS was improved by several constructive comments/suggestions from referees of the journal. This study was partially supported by a grant-in-aid from the Japan Society of the Promotion of Science (awarded to SO: No. 25304031).