





Citation: Ermilov SG, Martens J, Tolstikov AV (2014) The genus Galumna in Nepalese oribatid mite fauna, with notes on systematic placement of some species (Acari, Oribatida, Galumnidae). ZooKeys 438: 33–44. doi: 10.3897/zookeys.438.8192
The oribatid mite genus Galumna (Oribatida, Galumnidae) is recorded for the first time in Nepal. A new species, Galumna tetraporosa sp. n., is described from soil of secondary mixed broadleaved forest. It is most similar morphologically to G. tokyoensis Aoki, 1966 and G. valida Aoki, 1994, however, it differs from both by the absence of interlamellar setae and the presence of two pairs of notogastral porose areas Aa. Galumna granalata Aoki, 1984 is redescribed on the basis of specimens from Nepal. Galumna floridae (Jacot, 1929) and G. hexagona Balogh, 1960 are transferred to the genus Notogalumna; G. mauritii Mahunka, 1978 is transferred to the genus Dimidiogalumna.
Oribatida, new species, supplementary description, new combination, Galumnidae, Galumna, Notogalumna, Dimidiogalumna, Nepal
Galumna is a genus that was proposed by
In the course of taxonomic identification of Nepalese Galumna, we discovered two species: Galumna granalata Aoki, 1984 and a new one. This genus is recorded for the first time in Nepal. The primary goal of this paper is to describe and illustrate a new species. The secondary goal is to make a supplementary description of Galumna granalata based on the Nepalese material. This species was described
Additionally, the systematic placement of three species of the genus Galumna, Galumna floridae (Jacot, 1929), Galumna hexagona Balogh, 1960 and Galumna mauritii Mahunka, 1978, are discussed.
Three specimens (holotype: male; two paratypes: one male and one female) of Galumna tetraporosa sp. n. are from: Nepal, 2450–2720 m a.s.l., Terhathum District, ridge Tinjura Dara, broadleaved forest, soil, 17.IX.1983, collected by J. Martens and B. Daams.
Eight specimens of Galumna granalata are from: Nepal, 1650–1800 m a.s.l., Taplejung District, valley of the Kabeli River, below village Yamputhin, secondary mixed broadleaved forest with bamboo, soil, 03–04.IX.1983, collected by J. Martens and B. Daams.
Specimens were mounted in lactic acid on temporary cavity slides for measurement and illustration. The body length was measured in lateral view, from the tip of the rostrum to the posterior edge of the ventral fig. The notogastral width refers to the maximum width in dorsal aspect. Lengths of body setae were measured in lateral aspect. All body measurements are presented in micrometers. Formulae for leg setation are given in parentheses according to the sequence trochanter–femur–genu–tibia–tarsus (famulus included). Formulae for leg solenidia are given in square brackets according to the sequence genu–tibia–tarsus. General terminology used in this paper follows that of Grandjean (summarized by
Body size: 913–979 × 730–763. Rostrum pointed. Rostral and lamellar setae of medium size. Interlamellar setae represented by alveolus. Bothridial setae long, clavate, barbed. Lamellar and sublamellar lines divergent in medio-distal part. Anterior notogastral margin not developed. Notogaster with five pairs of small, rounded porose areas, Aa divided into two parts. Median pore absent. Aggenital and ano-adanal setae minute. Postanal porose area present, elongated.
Measurements. Body length: 962 (holotype), 913–979 (two paratypes); notogaster width: 730 (holotype), 730–763 (two paratypes).
Integument. Body color black to dark brown. Body surface smooth. Pteromorphs with poorly visible radiate wrinkles.
Prodorsum. Rostrum with strong tooth (12–20). Rostral setae (ro, 53–65) setiform, barbed. Lamellar setae (le, 65–77) setiform, little thicker and less barbed than rostral setae. Interlamellar setae absent, represented alveolus. Bothridial setae (ss, 114–123) with long stalk and shorter, barbed clavate head. Exobothridial setae absent. Porose areas Ad present, elongate oval (24–32 × 12–16), but visible only in dissected specimen. Lamellar lines (L) curving backwards; sublamellar lines (S) parallel in basal part and divergent in medio-distal part to lamellar lines.
Notogaster. Anterior notogastral margin not developed. Dorsophragmata (D) of medium size, elongated. Notogastral setae represented by 10 pairs of alveoli. Five pairs of small, rounded porose areas with distinct borders: Aa divided into two porose areas – smaller lateral (8–16) and larger medial (20–28); A1 (12–16) and A2 (16–28) located close to each other; A3 (24–36) usually largest. Alveoli la inserted posteriorly to Aa. Lyrifissures im located anteriorly or antero-laterally to A1. Opisthonotal gland openings (gla) located antero-laterally to A2. Median pore absent.
Gnathosoma. Generally, morphology of subcapitulum, palps and chelicerae typical for most Galumnidae (for example, see
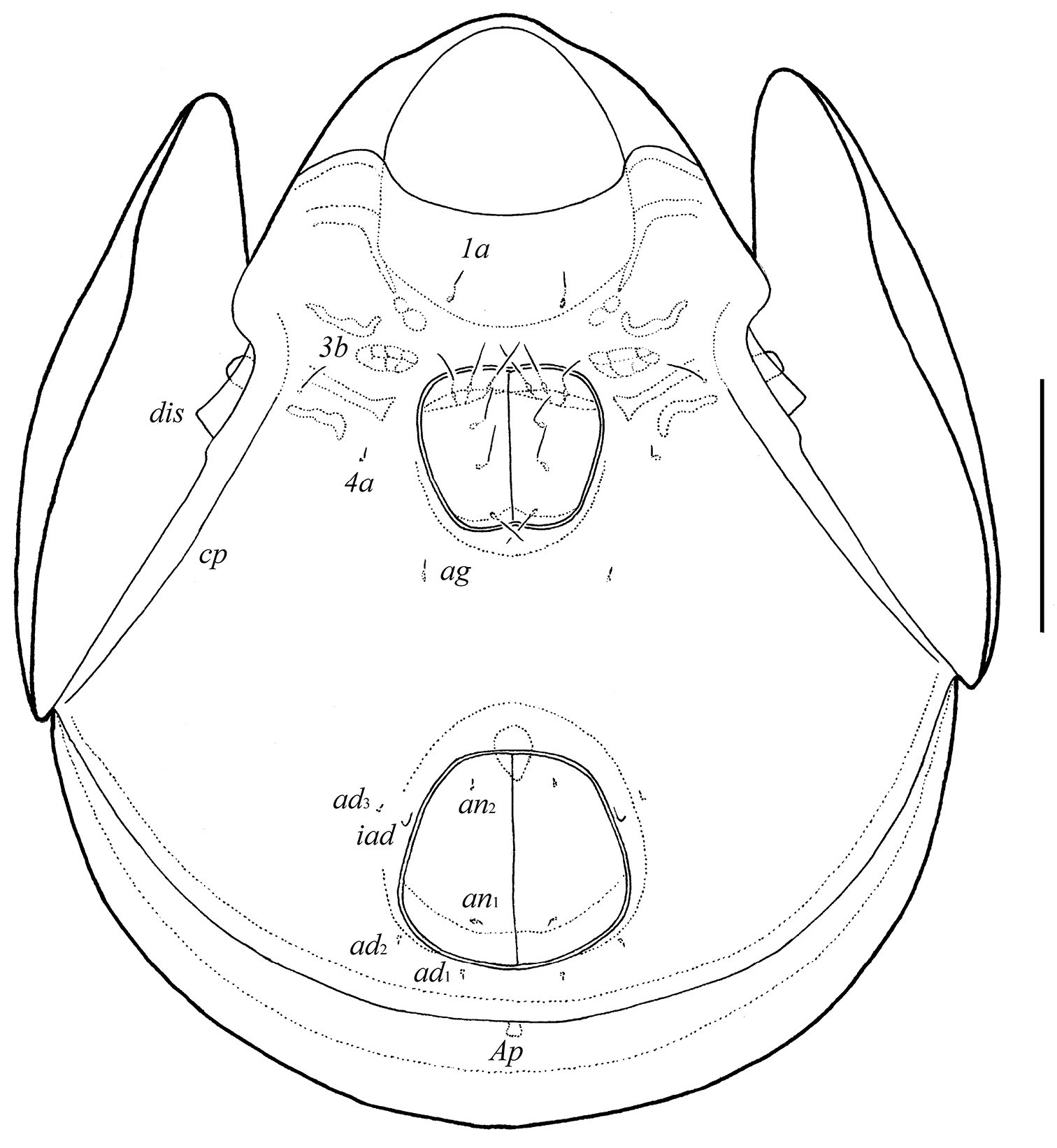
Epimeral and lateral podosomal regions. Apodemes (1, 2, sejugal, 3) well visible. Four pairs of epimeral setae (1b, 3b, 4a, 4b) observed ventrally, all setiform, thin, smooth, similar in length (24–32). Discidia (dis) triangular, circumpedal carinae (cp) distinct.
Anogenital region. Six pairs of genital setae (g1–g6, 24–28) setiform, thin, smooth. Anterior edge of genital figs with three setae. One pair of aggenital (ag), two pairs of anal (an1, an2) and three pairs of adanal (ad1–ad3) setae minute, similar in length (4). Adanal setae ad3 inserted laterally to adanal lyrifissures iad. Postanal porose area (Ap) elongated (36–49 × 8–12).
Legs. Generally, morphology of leg segments, setae and solenidia typical for most Galumnidae (for example, see
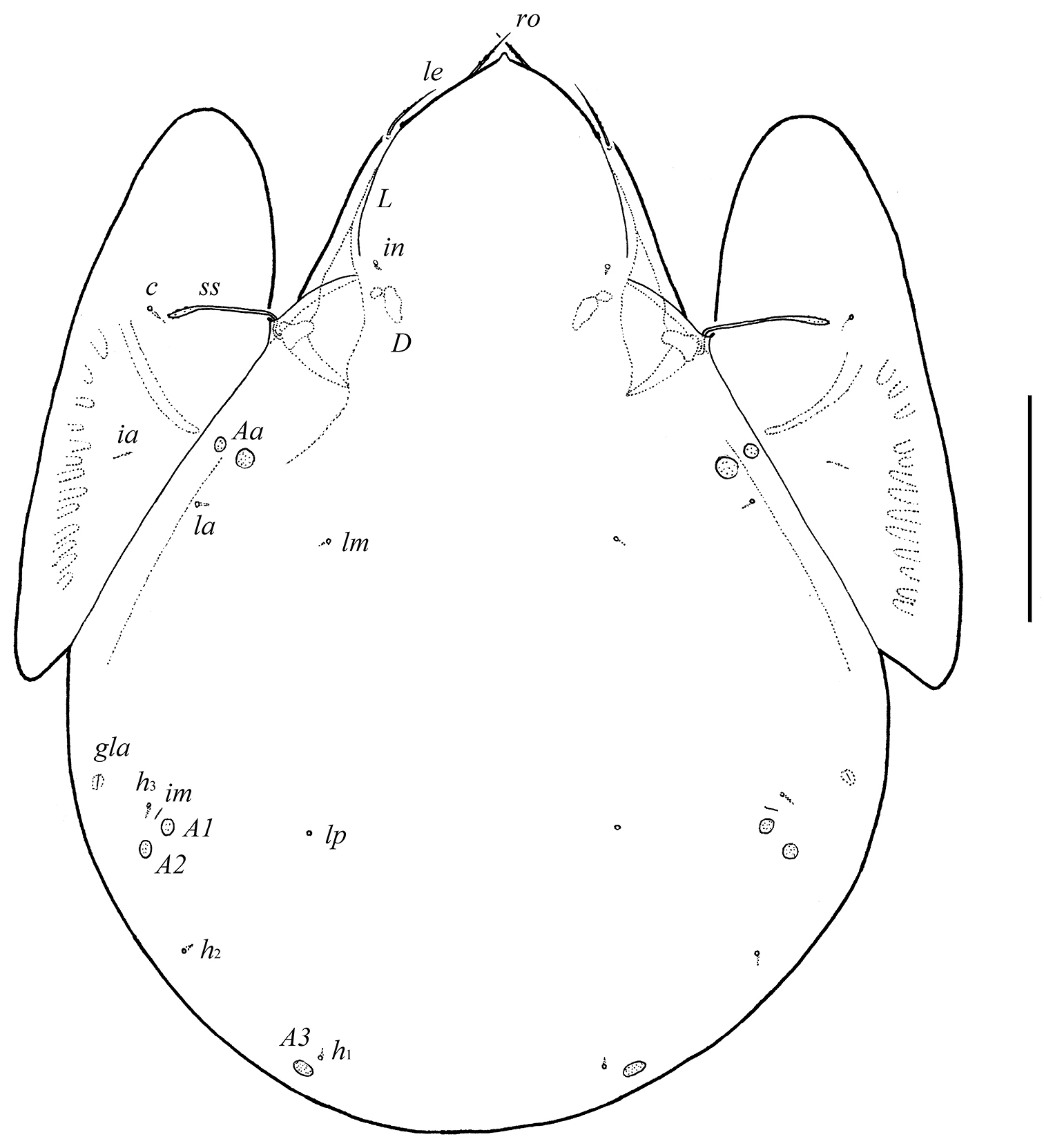
Galumna tetraporosa sp. n., adult: dorsal view. Scale bar 200 μm.
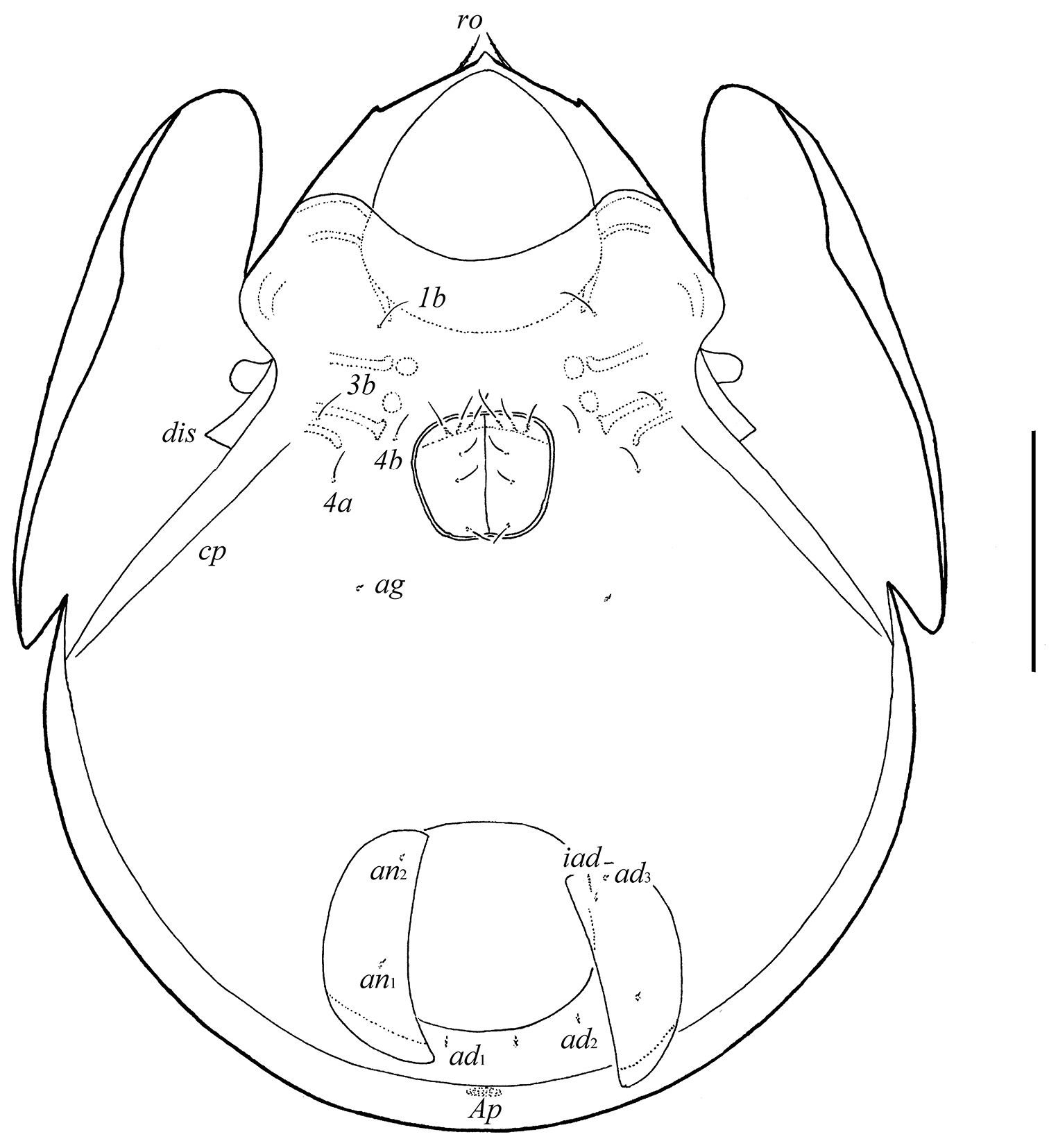
Galumna tetraporosa sp. n., adult: ventral view (gnathosoma and legs not illustrated). Scale bar 200 μm.
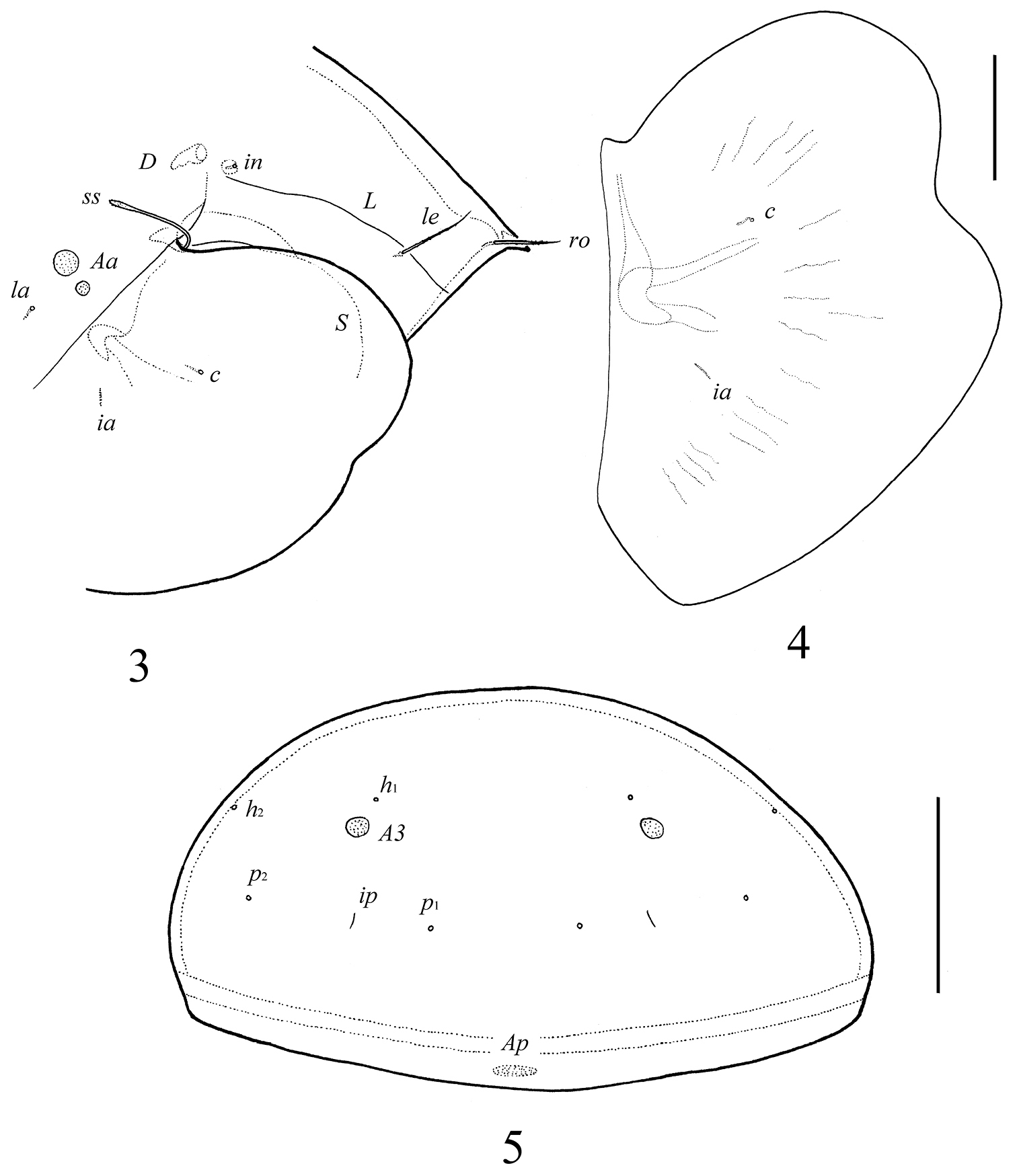
Galumna tetraporosa sp. n., adult: 3 dorso-lateral view of prodorsum and anterior part of notogaster and pteromorph 4 pteromorph 5 posterior view of notogaster. Scale bars 200 μm (3, 5), 100 μm (4).
Leg setation and solenidia of adult Galumna tetraporosa sp. n. (same data for Galumna granalata Aoki, 1984).
| Leg | Trochanter | Femur | Genu | Tibia | Tarsus |
|---|---|---|---|---|---|
| I | v’ | d, (l), bv’’ | (l), v’, σ | (l), (v), φ1, φ2 | (ft), (tc), (it), (p), (u), (a), s, (pv), v’, (pl), l’’, ε, ω1, ω2 |
| II | v’ | d, (l), bv’’ | (l), v’, σ | (l), (v), φ | (ft), (tc), (it), (p), (u), (a), s, (pv), ω1, ω2 |
| III | v’ | d, ev’ | l’, σ | l’, (v), φ | (ft), (tc), (it), (p), (u), (a), s, (pv) |
| IV | v’ | d, ev’ | d, l’ | l’, (v), φ | ft’’, (tc), (p), (u), (a), s, (pv) |
Roman letters refer to normal setae (ε to famulus), Greek letters to solenidia. Single prime (’) marks setae on anterior and double prime (’’) setae on posterior side of the given leg segment. Parentheses refer to a pair of setae.
The holotype and one paratype are deposited in the collection of the Senckenberg Institution Frankfurt, Germany; one paratype is deposited in the collection of the Tyumen State University Museum of Zoology, Tyumen, Russia.
The specific name “tetraporosa” refers to the four notogastral porose areas Aa.
In having the pointed rostrum and bothridial setae with well developed head, Galumna tetraporosa sp. n. is most similar to Galumna tokyoensis Aoki, 1966 from the eastern Palaearctic region and Galumna valida Aoki, 1994 from the Pacific Islands. However, it clearly differs from both by the absence of interlamellar setae (versus long in Galumna tokyoensis and Galumna valida) and the presence of two pairs of notogastral porose areas Aa (versus one pair in Galumna tokyoensis and Galumna valida).
Measurements. Body length: 431–464 (eight specimens); notogaster width: 332–356 (eight specimens).
Integument. Body color light brown. Body surface smooth. Pteromorphs with radiate wrinkles and small (length up to 12), elongate, grain-shaped tubercles. Wrinkles and tubercles very poorly visible in some specimens.
Prodorsum. Rostrum rounded. Rostral (28–32) and lamellar (16–20) setae setiform, thin, smooth, often not visible in dorsal view. Interlamellar setae minute (4). Bothridial setae (57–65) with long stalk and shorter, barbed, clavate head. Exobothridial setae absent. Porose areas Ad present, elongate oval (10–16 × 4–6). Lamellar and sublamellar lines parallel, curving backwards.
Notogaster. Anterior notogastral margin well developed, convex. Dorsophragmata of medium size, triangular. Notogastral setae represented by 10 pairs of alveoli. Four pairs of rounded porose areas with distinct borders: Aa (16–20) larger than A1 (12–16), A2 (8–10) and A3 (10–16). Alveoli la inserted posteriorly to Aa. Lyrifissures im located between lm and lp. Opisthonotal gland openings located laterally to A1. Median pore present, located little posterior to the virtual line connecting porose areas A1.
Gnathosoma. Generally, morphology of subcapitulum, palps and chelicerae typical for most Galumnidae (for example, see
Epimeral and lateral podosomal regions. Apodemes (1, 2, sejugal, 3) well visible. Three pairs of epimeral setae (1a, 3b, 4a) observed ventrally, all setiform, thin, smooth; 1a and 3b (12–16) longer than 4a (8). Discidia triangular, circumpedal carinae distinct.
Anogenital region. Six pairs of genital setae (12–16) setiform, thin, smooth. Anterior edge of genital figs with three setae. One pair of aggenital (8), two pairs of anal (4) and three pairs of adanal (4) setae minute. Adanal setae ad3 inserted laterally to adanal lyrifissures iad. Postanal porose area (Ap) very small, rounded (6–12). Ovipositor of typical morphology for Galumnidae (
Legs. Generally, morphology of leg segments, setae and solenidia typical for most Galumnidae (for example, see
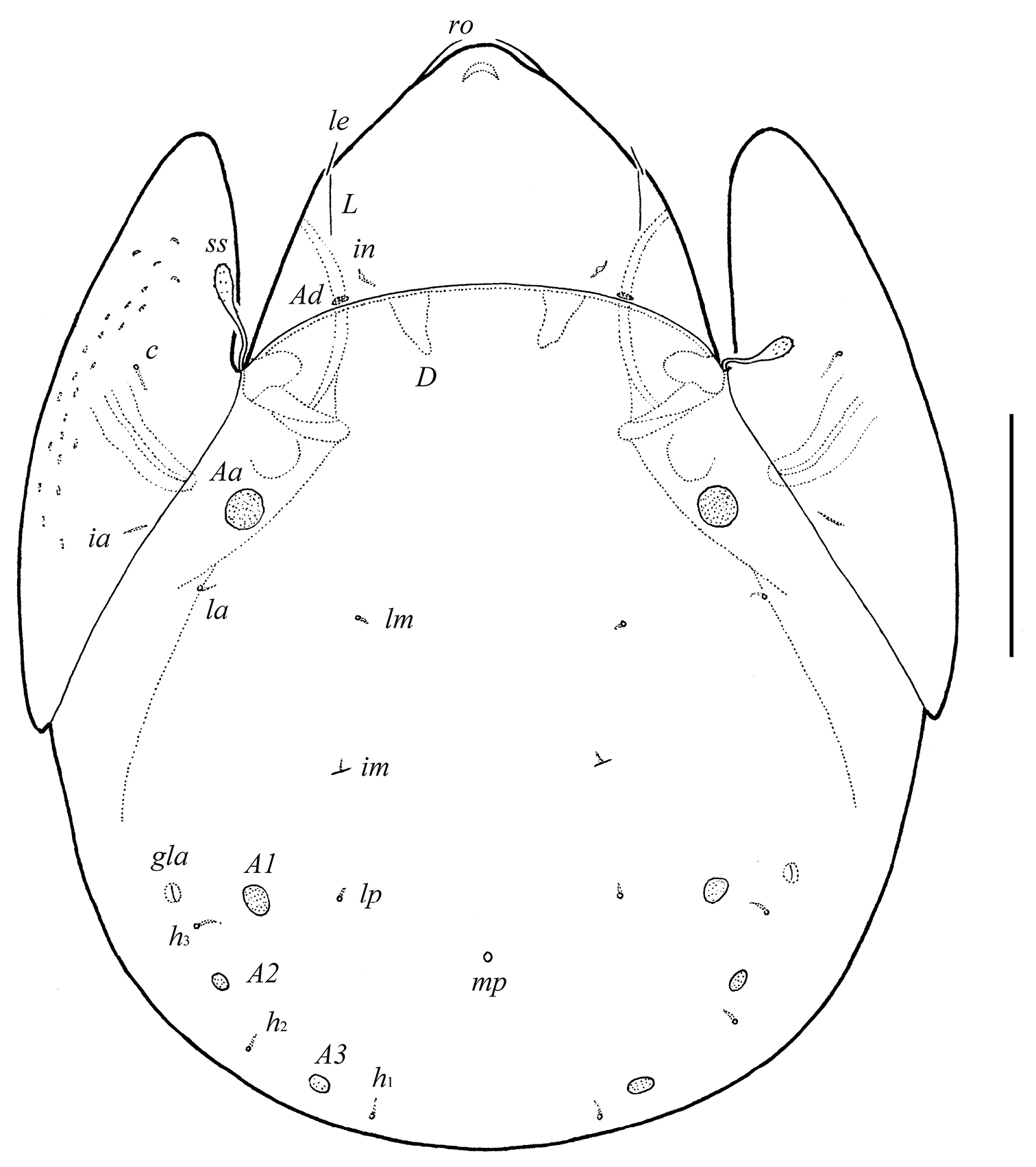
Galumna granalata Aoki, 1984, adult: dorsal view. Scale bar 100 μm.
Galumna granalata Aoki, 1984, adult: ventral view (gnathosoma and legs not illustrated). Scale bar 100 μm.
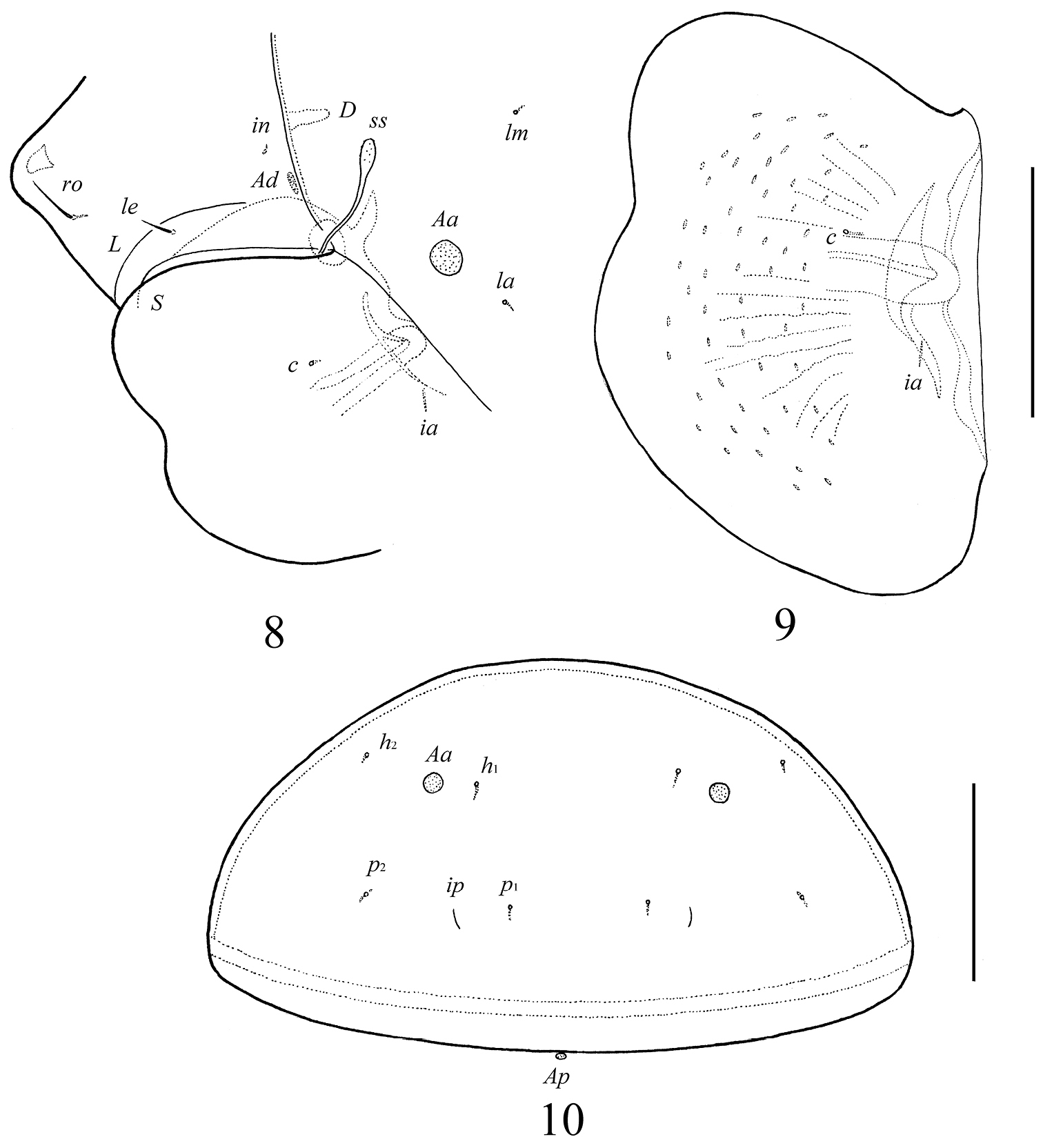
Galumna granalata Aoki, 1984, adult: 8 dorso-lateral view of prodorsum and anterior part of notogaster and pteromorph 9 pteromorph 10 posterior view of notogaster. Scale bar 100 μm.
Galumna granalata distinctly differs from other species of the genus Galumna by the presence of grain-shaped tubercles (granules in opinion of Aoki) on pteromorphs. The present Nepalese specimens of this species are morphologically and in general appearance similar to the Japanese specimens (
The analysis of literature on the Galumna-species has revealed an incorrect systematic placement of three species: Galumna floridae (Jacot, 1929), Galumna hexagona Balogh, 1960 and Galumna mauritii Mahunka, 1978.
Galumna floridae and Galumna hexagona were described by
Galumna mauritii was described by
We cordially thank Dr. Umukusum Shtanchaeva and Prof. Dr. Luis Subías (Universidad Complutense de Madrid, Madrid, Spain) for consultations, and two anonymous reviewers for the valuable comments. Jochen Martens thanks B. Daams for helpful companionship during the Nepalese expeditions, as well as the Feldbausch Foundation and the Wagner foundation at Fachbereich Biologie of Mainz University for their support of the author's field work in Asia.