





Citation: Huang J, Yoshitake H, Zhang R, Ito M (2014) Taxonomic revision of the East Asian genus Scleropteroides Colonnelli 1979 (Coleoptera, Curculionidae, Ceutorhynchinae). ZooKeys 437: 45–86. doi: 10.3897/zookeys.437.6563
The genus Scleropteroides Colonnelli, 1979 (Ceutorhynchinae: Scleropterini) was revised on the basis of detailed morphological observations. The genus was redefined to include three species from East Asia: S. hypocrita (Hustache, 1916) is redescribed and recorded from northeastern China and northern Korea for the first time; S. horridulus (Voss, 1958) is redescribed with new records from southern Korea; S. insularis Voss, 1971 was moved from synonymy with S. hypocrita to that with S. horridulus (syn. n.), and S. longiprocessus Huang & Yoshitake, sp. n. is described as new, sympatric with S. hypocrita in Japan. All the species are associated with woody Rubus species (Rosaceae). A key to species, habitus photographs, illustrations of important characters, and distribution maps are provided for each species.
Rubus, new record, new species, Scleropterus, taxonomy
Scleropteroides Colonnelli, 1979 is placed in the tribe Scleropterini, subfamily Ceutorhynchinae, and is characterized mainly by a rostrum expanded from the base to apex, a visible scutellar shield, subtriangular elytra, strongly convex humeri, acute squamate granules on the elytral intervals, a deep rostral channel extending to the metaventrite, and dentate femora (
The genus Scleropteroides was established for Ceuthorrhynchidius hypocrita Hustache, 1916 from Japan (
Presently, Scleropteroides is in need of revision. Apart from morphological differences associated with hind-wing reduction in Scleropterus, the distinction between the two genera is still insufficient because more than half of the defining characteristics of Scleropteroides are common to Scleropterus or even incorrectly described. Moreover, the taxonomic status of Rhytidosomus insularis should be revised because it shows remarkable differences from Scleropteroides hypocrita despite Colonnelli’s synonymy.
In this study, we revise the genus Scleropteroides on the basis of detailed examinations of specimens collected from various localities in East Asia. In addition, we provide distributional and ecological information on Scleropteroides species. The systematic position of the genus, relationships among species, and the geographic distribution and host plant association of each species are discussed.
Specimens preserved in the following institutions and private collections were examined for this study: Canadian Museum of Nature, Ottawa/Gatineau (CMN); Entomological Laboratory, Faculty of Agriculture, Kyushu University, Fukuoka, Japan (ELKU); Laboratory of Environmental Entomology, Faculty of Agriculture, Ehime University, Matsuyama, Japan (EUMJ); Institute of Zoology, Chinese Academy of Sciences, Beijing, China (IZCAS); K. Izawa collection, Tajimi, Japan (KI); Muséum National d'Histoire Naturelle, Paris, France (MNHN); National Institute for Agro-Environmental Sciences, Tsukuba, Japan (NIAES); Natural History Museum Vienna, Wien, Austria (NHMV); S. Miyakawa collection, Kyushu University Museum, Fukuoka, Japan (SM); and Y. Shiozaki collection, Kawasaki, Japan (YS). All the descriptive work in this study was completed by J. Huang and H. Yoshitake.
External structures were observed under a Leica MZ95 stereoscopic microscope. Measurements of body parts are defined and abbreviated as follows: LB = body length, from the apex of the pronotum to the apices of elytra; LR = rostrum length, from a lateral view; WP = maximum width of pronotum; LP = pronotum length, from the base to the apex along the midline; WE = maximum width of elytra; and LE = length of elytra, from the base of humeri to the apex of elytra. All measurements are in millimeters. Habitus photographs were taken using a Nikon Coolpix995 digital camera attached to a Nikon SMZ1500 stereoscopic microscope. To examine terminalia, the specimens were macerated in hot water and dissected under a stereoscopic microscope. The abdomen was removed from the body and then cleaned in hot 10% KOH solution for 5–10 min. Terminalia were extracted from the abdomen and mounted on slides with glycerol (male) or pure water (female), examined using a Nikon Eclipse 55i optical microscope, and illustrated in detail using a camera lucida. Scale bars were calibrated using a Nikon objective micrometer. Details of some external structures were observed with a scanning electron microscope (Hitachi 3000-N). Plant nomenclature follows
Ceuthorrhynchidius hypocrita Hustache, 1916.
This genus is very similar to Scleropterus in having a six-segmented antennal funicle, elytra bearing acute squamate granules (usually in a row on each interval), and a rostral channel extending to the metaventrite. However, it is easily distinguishable from Scleropterus mainly by the bisinuate basal margin of the pronotum (Figs 10, 34, 36, 38), the well-developed scutellar shield (Figs 11, 34–39), prominent elytral humeri (Figs 11, 35, 37, 39), even elytral intervals (Figs 11, 35, 37, 39), dentate femora (Fig. 15), and simple fore tibiae that are not incurved apically (Fig. 15).
Dark brown in general appearance. Head, rostrum, prothorax (except apical part), venter, and pygidium black; antennae, apical part of prothorax, and tarsi paler.
Vestiture. Body surface evenly covered with ochreous pollinosity in life. Head mainly covered with brown clavate scales, mixed with white scales; vertex with scales directed medially; forehead with scales directed basally; basal margin and median carina fringed with white ovate recumbent scales. Rostrum covered with clavate scales on basal 2/3; scales directed basally, gradually becoming smaller toward apex, replaced by hair-like scales on apical 1/3. Prothorax mainly covered with very similar scales as those on head, with longitudinal stripe of white ovate recumbent scales on median and lateral parts; each scale directed basally. Elytra (Fig. 11) bearing row of white and brown clavate scales on each interval; scales directed apically. Legs (Fig. 15) densely covered with white and brown scales; femora mostly covered with clavate semirecumbent scales, mingled with feather-like scales along inner margin; tibiae mainly covered with clavate scales, except semirecumbent hair-like scales along inner margin; corbel of each tibia fringed with brown setae. Lateral pieces of meso- and metathorax mainly covered with white ovate recumbent scales. Sterna mainly covered with white lanceolate to ovate scales; meso- and metaventral receptacles densely covered with white aciculate scales. Venter mainly covered with white lanceolate to ovate recumbent scales, mingled with white and brown clavate scales; ventrites III and IV nearly naked on disc, with only transverse row of clavate scales; ventrite V bearing fine brown scales in median concavity and white ovate scales on sides. Pygidium mainly sparsely covered with short clavate semirecumbent scales, mingled with hair-like recumbent scales; scales brown, directed ventrally.
Head (Figs 7–8) reticulately punctured; each puncture moderate in size; vertex with median carina from base to apex; carina becoming obscure apically; forehead shallowly depressed, with apex slightly broader than base of rostrum and then strongly widened basally; eyes medium-sized, rounded triangular, weakly convex. Rostrum (Figs 7–8) slender, evenly curved; dorsum densely rugosely punctured except apex; punctures moderate in size from base to level between antennal insertions, then becoming smaller, sparser, and shallower toward apex; sides subparallel in basal half, more or less widened before antennal insertions, then subparallel in apical part; antennal scrobes well separated on ventral surface. Antennae (Figs 8–9) inserted before middle of rostrum; scape moderate in length, evidently clavate, round and fringed with 3–4 setae at apex, slightly shorter than funicle; funicle six-segmented; club lanceolate, finely pubescent except basal part.
Pronotum (Figs 10–13) slightly wider than long; dorsum densely coarsely punctured, simple, lacking tubercle or prominence; punctures smaller in apical and basal parts; basal margin bisinuate, smooth, not serrate; apical margin weakly raised, with shallow median incision. Scutellar shield subovate.
Elytra (Figs 11–12) subcordate, nearly as long as wide, widest just behind humeri; suture evidently bent leftward; interval I of right elytron slightly broader than that of left elytron; all intervals subequal in width and height, nearly three times as wide as striae, each with row of more or less small and acute squamate granules; striae less marked, nearly naked, lacking conspicuous scales or hair, finely punctured; each puncture round, separated by distance more than three times as long as its diameter. Hind wings (Fig. 19) well-developed.
Legs (Fig. 15) slender; femora slightly clavate, each armed with small tooth, bearing minute squamate granules; no jumping organ present (Fig. 16); tibiae bearing minute squamate or setiferous granules; protibiae simple, lacking mucro and not curved in at apex; meso- and metatibiae moderately mucronate; corbels short, simple, not dilated outward; tarsi (Figs 17–18) moderate in length; claws free, slender, appendiculate with sharp teeth; each tooth slender, extending from base to middle of each claw.
Underside. Prosternum coarsely and moderately densely punctured; mesoventrite densely and finely punctured; metaventrite with dense medium-sized punctures on disc and with sparser and coarser punctures on sides; lateral pieces of meso- and metasterna sparsely coarsely punctured. Rostral channel (Fig. 13) long, extending far beyond level between posterior margins of mesocoxal cavities, with dense minute punctures; mesoventral receptacle deep, laterally costate; costae short, subparallel; metaventral receptacle very deep, terminating in steep wall and U-shaped margin; lateral walls of meso- and metaventral receptacles interrupted by inner margins of mesocoxae; metaventrite more or less prominent ventrally along apico-lateral margin of metaventral receptacle. Metendosternite as in Fig. 14. Venter coarsely and more or less densely punctured; ventrites III and IV nearly polished on disc, with only transverse row of coarse and sparse punctures; ventrite V with subtriangular median concavity along basal margin; concavity faintly sulcate along midline. Tergum as in Figs 20–21; tergite VII with pair of minute setiferous plectral tubercles near base.
Pygidium transverse-pentagonal, flattened, very coarsely and reticulately punctured; bottom of each puncture opaque due to dense minute punctations; upper flange arcuate downward on each side.
Rostrum (Figs 24–25) slightly more slender. Antennae inserted just behind middle of the rostrum. Tibiae simple, not mucronate on all legs. Ventrites I and II moderately inflated, sparsely punctured, lacking prominence. Ventrite V simple or only with longitudinal median sulcus, lacking concavity. Pygidium smaller, sectorial, mainly covered with hair-like scales. Otherwise as in male.
East Asia (China including Taiwan, Korea, and Japan; Figs 133–134).
This species is characterized by the following characters: prothorax moderately constricted in the subapical part (Fig. 34); elytra with weakly prominent humeri, gently convergent toward the subapical calli (Fig. 35); scales in a row on elytral intervals, semirecumbent, evidently shorter than width of each interval (Fig. 35); apical half of the male rostrum slightly widened (Fig. 22); male metaventrite weakly prominent ventrally along the apico-lateral margin of the metaventral receptacle; male ventrite I with a median prominence along the apical margin; penis with apical projection blunt, rounded at the apex (Figs 41, 51, 61); basal part of the endophallus with a pair of fig-like sclerites (Figs 40, 50, 60); female sternite VIII with slender arms that are arcuate apically (Figs 47, 57, 67); posterior part of the bursa copulatrix densely covered with minute coniform spicules (Figs 45, 55, 65).
LB: 2.18–2.66 (mean, 2.41). LR: 0.96–1.11 (mean, 1.03). WP: 0.94–1.08 (mean, 1.01). LP: 0.78–0.97 (mean, 0.86). WE: 1.48–1.79 (mean, 1.61). LE: 1.50–1.82 (mean, 1.64). N = 10 for all measurements. Habitus as shown in Figs 1–2.

Habitus of Scleropteroides spp., males. 1–2 Scleropteroides hypocrita (Hustache) 3–4 Scleropteroides longiprocessus Huang & Yoshitake, sp. n. 5–6 Scleropteroides horridulus (Voss) 1, 3, 5 Dorsal habitus 2, 4, 6 Lateral habitus. Scale: 0.50 mm.
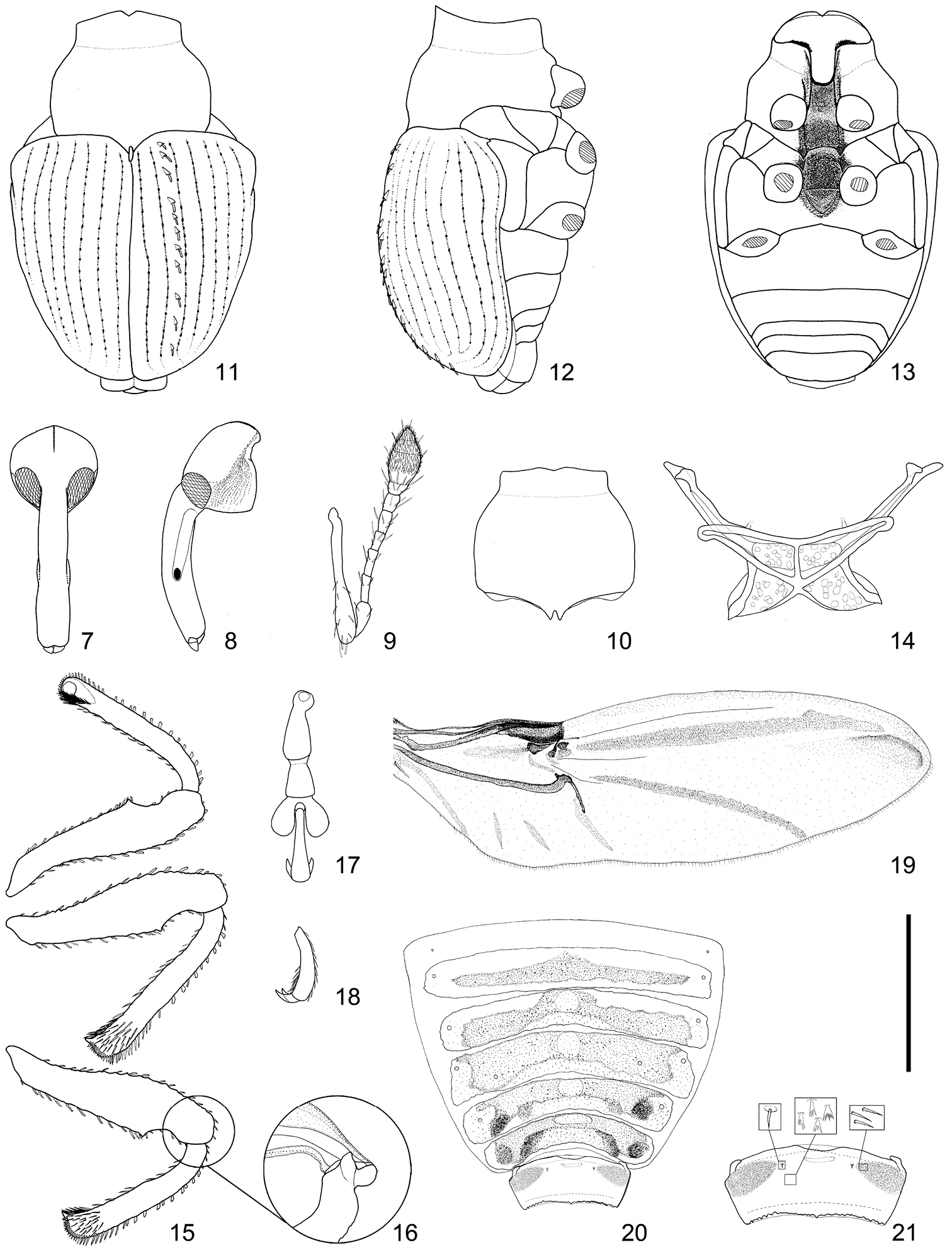
Diagnostic characteristics of Scleropteroides. 7 Head, dorsal view 8 Head, lateral view 9 Antenna 10 Prothorax 11 Body, dorsal view 12 Body, lateral view 13 Body, ventral view 14 Metendosternite 15 Legs 16 Base of hind-tibia, showing extensor and flexor tendons 17 Tarsus 18 Tarsomere V, lateral view 19 Hind wing 20 Tergites 21 Tergite VII. Scale: 1.00 mm for 7, 8, 10–13, 16, 19–20; 0.75 mm for 15, 20; 0.50 mm for 9, 14, 17–18, 21.
Vestiture. Clavate scales short and semirecumbent on head, basal 2/3 of rostrum, and pronotum. Hair-like scales fine and semirecumbent on apical 1/3 of rostrum. Scales on elytral intervals (Fig. 35) semirecumbent, short, 0.4–0.8 × as long as interval width. Clavate scales on tibiae semierect.
Rostrum (Figs 22–23) slender, 1.15–1.24 × as long as pronotum; apical part of sides slightly widened and 1.14 × as long as basal part. Antennae (Fig. 23) with length ratio of funicular segments I: II: III: IV: V: VI = 1.89: 1.44: 1.36: 1.00: 1.17: 1.00 and width ratio = 1.71: 1.00: 1.21: 1.29: 1.50: 1.64.
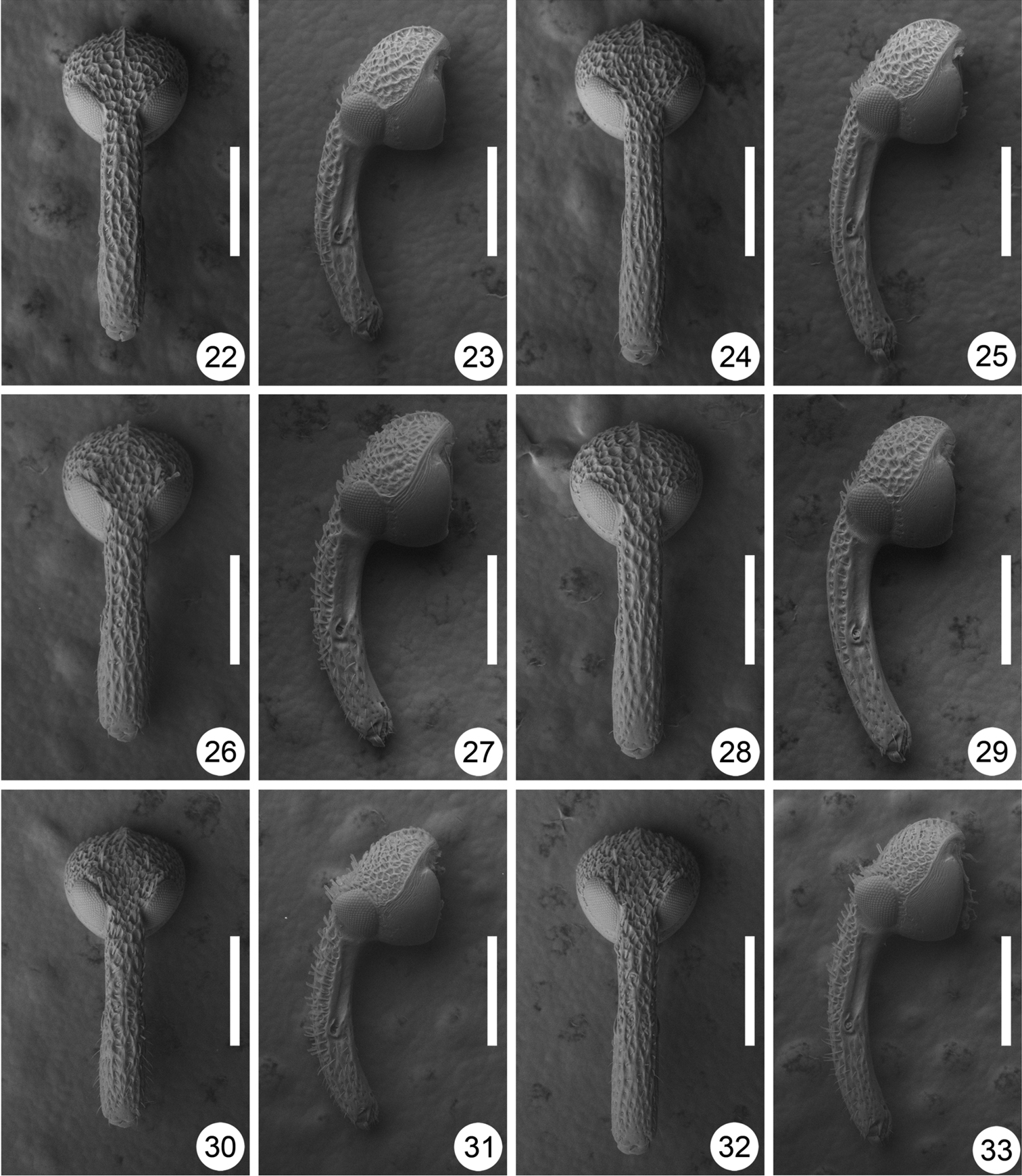
Heads of Scleropteroides spp. 22–25 Scleropteroides hypocrita (Hustache) 26–29 Scleropteroides longiprocessus Huang & Yoshitake, sp. n. 30–33 Scleropteroides horridulus (Voss) 22, 23, 26, 27, 30–31 Male 24, 25, 28, 29, 32–33 Female 22, 24, 26, 28, 30, 32 Dorsal view 23, 25, 27, 29, 31, 33 Lateral view. Scale: 0.50 mm.
Pronotum (Fig. 34) 1.12–1.21 × as long as wide, 0.51–0.54 × as long as and 0.60–0.66 × as wide as elytra; subapical constriction moderate; sides slightly widened from base to basal 1/3, faintly narrowed to middle, then gradually convergent toward apex.

Pronotum and right elytron of Scleropteroides spp., male. 34–35 Scleropteroides hypocrita (Hustache) 36–37 Scleropteroides longiprocessus Huang & Yoshitake, sp. n. 38–39 Scleropteroides horridulus (Voss) 34, 36, 38 Pronotum 35, 37, 39 Right elytron. Scale: 0.50 mm.
Elytra 0.98–1.07 × as long as wide, 1.87–1.97 × as long as and 1.51–1.66 × as wide as pronotum, gently convergent toward subapical calli; humeral calli weakly prominent; subapical calli weakly prominent.
Underside. Metaventrite weakly prominent ventrally along apico-lateral margin of metaventral receptacle. Venter coarsely and moderately densely punctured; ventrite I with median prominence along apical margin; prominence weak, semicircular, densely and finely punctured; ventrite V with more or less shallow median concavity; length ratio of ventrites I: II: III: IV: V = 4.18: 2.00: 1.00: 1.14: 1.82 and width ratio = 1.96: 1.75: 1.32: 1.18: 1.00.
Pygidium transverse-pentagonal.
Terminalia and genitalia. Sternite VIII (Figs 44, 54, 64) diminished into pair of eye-shaped sclerites; spiculum gastrale robust, evidently longer than penis or its apodeme, bent leftward. Tegmen (Figs 43, 53, 63) with apodeme more or less stout, nearly 0.8 × as long as the diameter of the tegminal ring, more or less widened toward apex. Penis (Figs 40–42, 50–52, 60–62) broad, relatively thin in profile, more or less moderately curved downward in the basal 2/3, then slightly bent upward in apical 1/3; sides faintly narrowed from base to basal 1/3, slightly broadened from basal 1/3 to apical 1/5, then strongly convergent apically; apical projection blunt, rounded at apex. Basal part of endophallus (Figs 40, 50, 60) with pair of fig-like sclerites, numerous rounded spicules in median part, and moderately dense minute spicules in apical part.
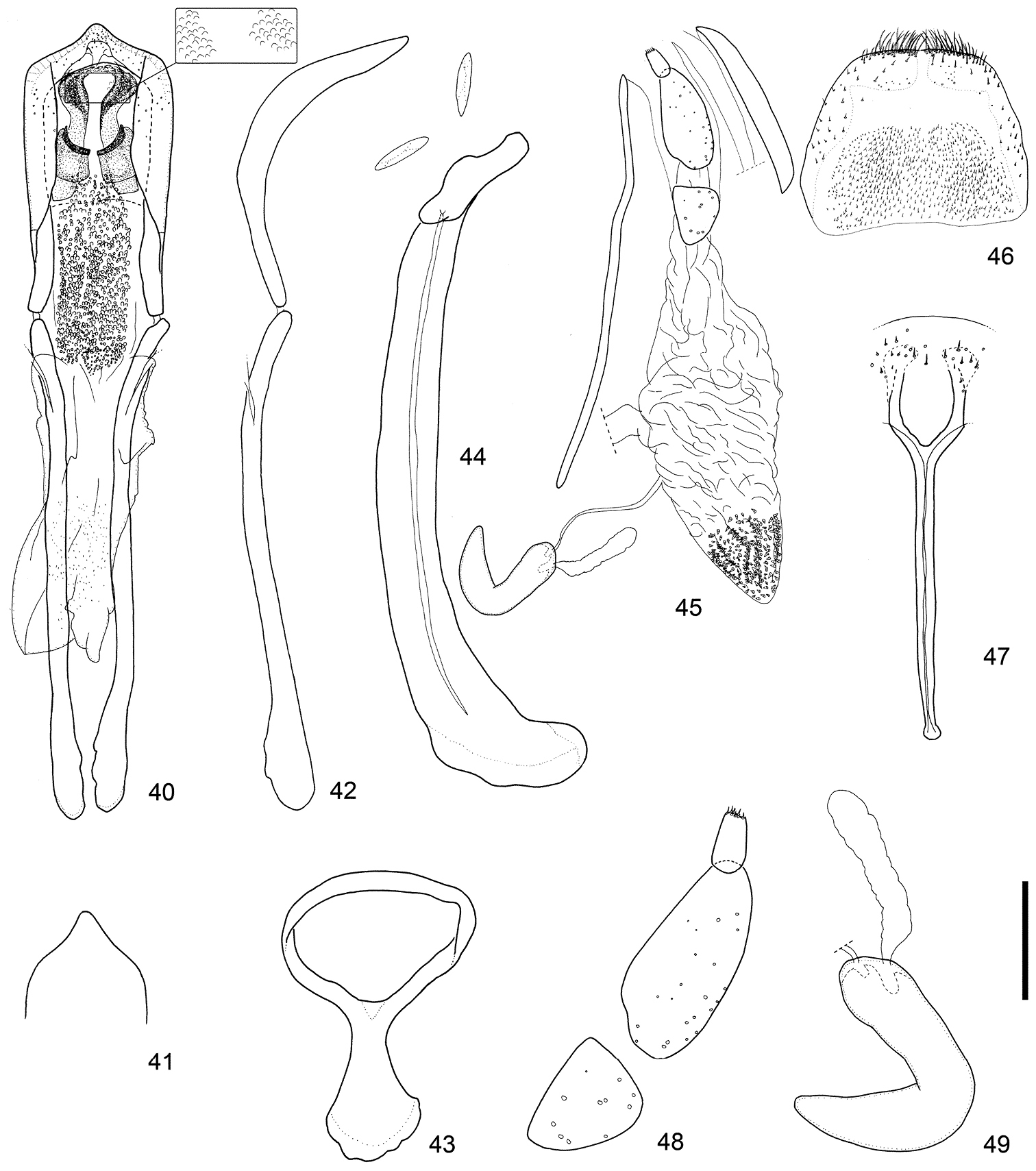
Male and female terminalia of Scleropteroides hypocrita (Hustache) from Takaosan, Japan. 40 Aedeagus, dorsal view 41 Apex of the penis, dorsal view 42 Aedeagus, lateral view 43 Tegmen 44 Sternites VIII and IX, male 45 Female terminalia and genitalia, lateral view. 46 Tergite VIII, female 47 Sternite VIII, female 48 Coxite and stylus. 49 Spermatheca. Scale: 0.20 mm for 40–47, 0.10 mm for 48–49.
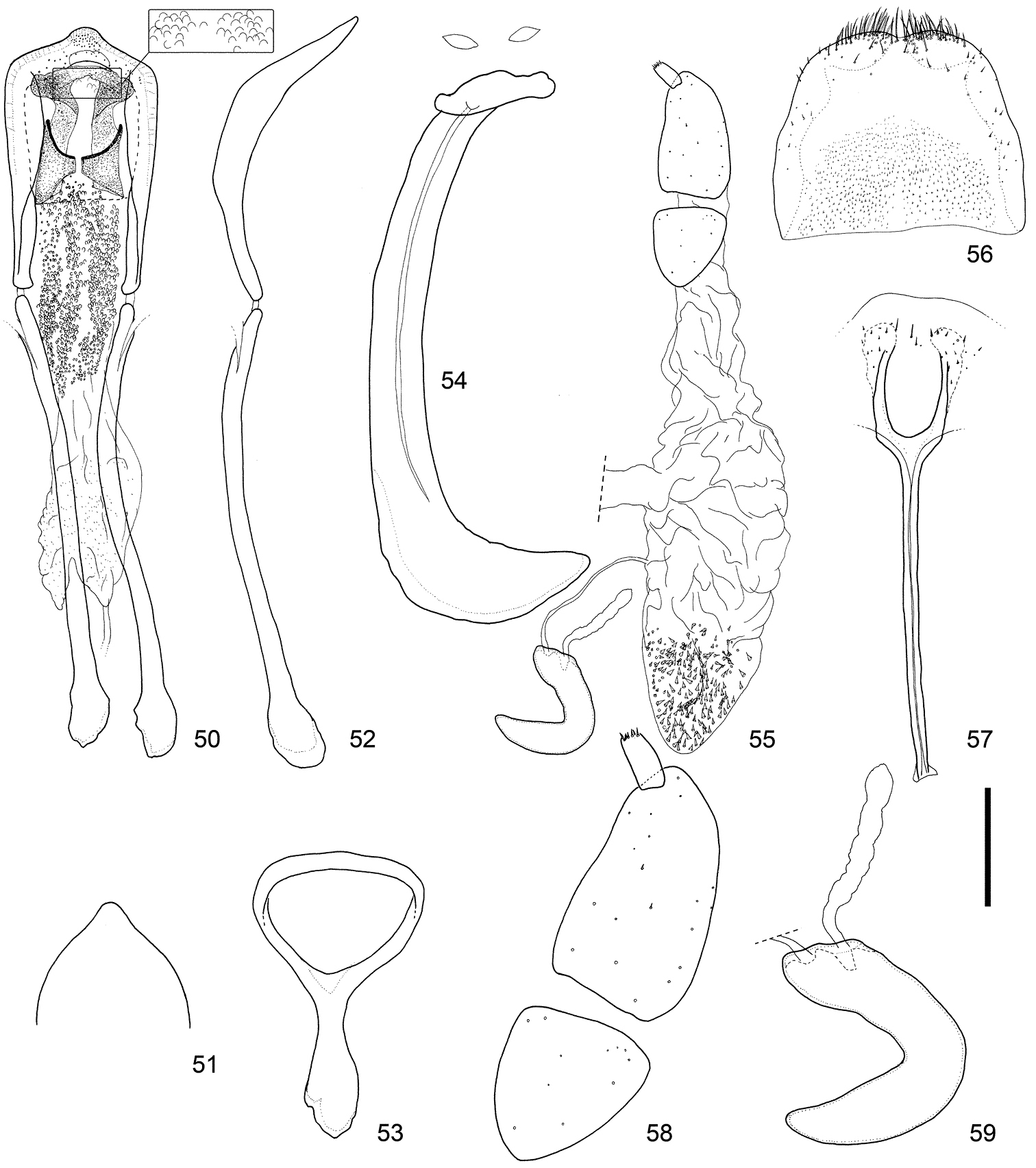
Male and female terminalia of Scleropteroides hypocrita (Hustache) from Mt. Jirisan, Korea. 50 Aedeagus, dorsal view 51 Apex of the penis, dorsal view 52 Aedeagus, lateral view 53 Tegmen 54 Sternites VIII and IX, male 55 Female genitalia, lateral view 56 Tergite VIII, female 57 Sternite VIII, female 58 Coxite and stylus 59 Spermatheca. Scale: 0.20 mm for 50–57, 0.10 mm for 58–59.
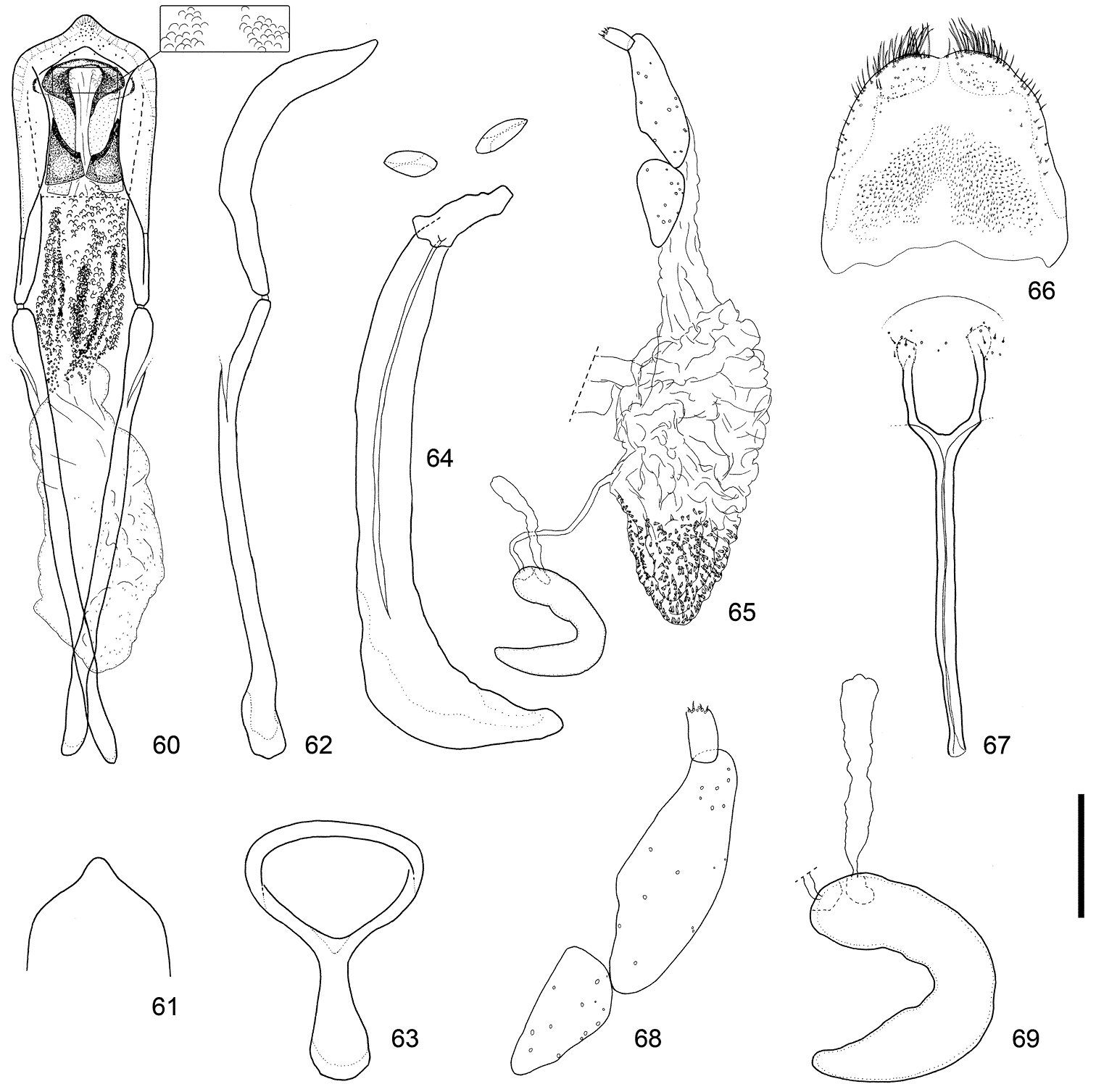
Male and female terminalia of Scleropteroides hypocrita (Hustache) from Heilongjiang, northeastern China. 60 Aedeagus, dorsal view 61 Apex of the penis, dorsal view 62 Aedeagus, lateral view 63 Tegmen 64 Sternites VIII and IX, male 65 Female genitalia, lateral view 66 Tergite VIII, female 67 Sternite VIII, female 68 Coxite and stylus 69 Spermatheca. Scale: 0.20 mm for 60–67, 0.10 mm for 68–69.
LB: 2.28–2.67 (mean, 2.50). LR: 1.05–1.31 (mean, 1.19). WP: 0.95–1.11 (mean, 1.03). LP: 0.80–0.97 (mean, 0.89). WE: 1.54–1.89 (mean, 1.72). LE: 1.57–1.82 (mean, 1.72). N = 11 for all measurements.
Rostrum (Figs 24–25) slightly more slender, 1.27–1.36 × as long as pronotum.
Pronotum 1.11–1.20 × as wide as long.
Elytra 0.97–1.03 × as long as wide.
Underside. Metaventrite more strongly prominent ventrally along apical margin of metaventral receptacle. Ventrites I and II moderately inflated, sparsely punctured, lacking prominence. Ventrite V lacking concavity, with only longitudinal median sulcus that is sometimes obscure.
Pygidium smaller, fan-shaped.
Terminalia and genitalia. Tergite VIII (Figs 46, 56, 66) with pair of combs of dense, long setae along apical margin. Sternite VIII (Figs 47, 57, 67) with pair of patches of several minute setae near apex; arms relatively slender, nearly 0.8 × as long as apodemes, nearly half as long as coxite and stylus combined, slightly basally fused, and apically arcuate; apodemes slender, moderately divergent near apex. Bursa copulatrix (Figs 45, 55, 65) with dense minute coniform spicules in posterior part. Coxites (Figs 48, 58, 68) robust, subdivided into two pieces, nearly 7.0 × as long as styli; styli apicolaterally inserted, moderate in length, nearly 2.0 × as long as wide. Spermatheca (Figs 49, 59, 69) with cornu slender, strongly curved and slightly attenuate; collum slightly convex; ramus indistinct; insertions of duct and gland close to each other.
Otherwise as in male.
HOLOTYPE: 1 female (MNHN), “Mont Takao/près Hachiôji/Japon 30-5-08/Edme Gallois; MUSEUM PARIS/NIPPON MOYEN/E. GALLOIS 1912; Ceuthorrhynchidius/hypocritus/Hust./type” (hand written). HOLOTYPE OF RHYTIDOSOMUS (RHYTIDOSOMUS) HOLDHAUSI WAGNER, 1944: 1 male (NHMV), "Rorelz/1875/Süd/Japan" (hand-written on grayish card); "♂" (typed on white card); "Typus" (typed on orange card); "Hÿshichosoma/det.H.Wagner/Holdhausi m./Tÿpe! ♂" (hand-written in red on white card, partially typed); "TYPUS" (typed on red card); "Holdhausi/Jap. Wagn." (hand-written on grayish card); "Scleropteroides/hypocrita (Hustache, 1916)/E. Colonnelli det. 1991" (hand-written on white card, partially typed). CHINA: Liaoning. 1 male and 1 female, Benxi, 41°17'N, 123°44'E, 3-VI-1963, H. Li (IZCAS, IOZ(E)896779 and 896770). 1 male, Anshan, Qianshan, 41°05'N, 123°06'E, 9-VI-1963, H. Li (IZCAS, IOZ(E)896773). 1 male and 1 female, Fengcheng, Tongyuanpu, 40°17'N, 123°55'E, 30-VI-1963, H. Li (IZCAS, IOZ(E)896771–896772). Heilongjiang. 2 males and 2 females, Ercengdianzi, 45°26'N, 127°07'E, 15–22-VI-1941 (IZCAS, IOZ(E)896474–896477). D.P.R. KOREA: Pyonganbukdo. 1 female, Mt. Myohyang-San, Around Habiro, 200–550 m, 29-VI- 2009, C. Han (NIAES). Pyeongyangjikhalsi. PyongYang-City, around Mt. RyongAk-San, near Suna-river: 1 male and 2 females, 18-V-2012, C. Han (NIAES); 1 female, 19–22-V-2012, C. Han (NIAES). R. KOREA: Gangwondo. 3 males and 5 females, Chuncheongun, Dongmyeon, Gamjeongri, 21-V-1992, K. Morimoto (ELKU). 3 males and 1 female, Chuncheongun, 11-VI-1997, K. Morimoto (ELKU). Gyeonggido. Pocheongun, Kwangnung: 2 males and 3 females, 16-V-1984, K. Morimoto (ELKU); 1 male and 1 female, 14–19-V-1992, M. T. Chujo (ELKU). Gyeongsangbukdo. 1 male and 1 female, Yeonggi, 14-VI-1997, K. Morimoto (ELKU). Gyeongsangnamdo. 1 female, Mt. Jirisan, Daesungri, 8-VI-1991, J. D. Bae (ELKU). 1 female, Mt. Jirisan, Bycjum Valley, 9-VI-1983, Lee Lab (ELKU). 1 female, Mt. Jirisan, Yeongsinbong–Samsinbong, 17-VI-1994, H. Kojima (ELKU). 1 female, Mt. Jirisan, Piagol, 30-V-2000, H. Yoshitake (NIAES). 1 male, Mt. Jirisan, Simweon, 31-V-2000, S. Kamitani (NIAES). 1 male, Mt. Jirisan, 31-V-2000, H. Yoshitake (NIAES). Hamyanggun, Macheonmyeon, Samjeongri, Mt. Jirisan: 2 males and 2 females, 9-V-1991, J. D. Bae (ELKU); 1 male, 9-V-1991, K. Morimoto (ELKU); 3 females, 11-V-1991, J. D. Bae (ELKU); 2 males and 3 females, 15-V-1991, J. D. Bae (ELKU); 4 males and 1 female, 6-VI-1991, J. D. Bae (ELKU); 2 males and 2 females, Samjeongrurak, 7-VI-1991, J. D. Bae (ELKU); 1 male, 14-VII-1991, K. Morimoto (ELKU); 13 males and 17 females, 15-V-1991, K. Morimoto (ELKU); 6 males and 6 females, 14-VI-1994, H. Kojima (NIAES); 5 females, 516 m, 35°21'09.2"N, 127°38'58.8"E, 5-V-2005, H. Yoshitake, on Rubus crataegifolius (NIAES). Jeollabukdo. 1 male and 1 female, Namwongun, Manbok Valley, 1-VI-1991, J. D. Bae (ELKU). 1 male and 5 females, Namwongun, Deongdong Valley, 7-VI-1991, J. D. Bae (ELKU). 1 male, Namwongun, Nogodan, 12-VII-1991, J. D. Bae (ELKU). 3 males and 1 female, Namwongun, Deongdongri, 19-VI-1994, H. Kojima (ELKU). Namwongun, Sannaemyeon, Simwon Valley: 1 female, 10-V-1991, K. Morimoto (ELKU); 1 male and 3 females, 13-V-1991, J. D. Bae (ELKU). Namwongun, Sannaemyeon, Jeonglyongchy: 2 females, 12-V-1991, K. Morimoto (ELKU); 1 male and 1 female, 14-V-1991, K. Morimoto (ELKU); 1 male and 1 female, 5-VI-1991, J. D. Bae; 1 male, 12-VI-1991, J. D. Bae (ELKU). JAPAN: Hokkaido. 1 male, Hidaka, 11-VII-83, Y. Shiozaki (YS). 8 males and 4 females, Taisei, Hirahama, 7-VI-2003, T. Miyata (NIAES). 1 male and 1 female, Kamiiso, Tomigawa, Hosokomatazawa, 27-VI-1993, T. Miyata (NIAES). 1 female, Fukushima, Mitake, 13-VI-1998, H. Yoshitake (NIAES). 1 male and 2 females, Assabuchou, Shimizu, 14-VI-1998, H. Yoshitake (NIAES). Honshu. AOMORI. Shimokita-hantou Peninsula, Oohata, Yunomata: 1 female, 16-VII-1956, K. Morimoto (ELKU); 1 female, 26-VII-1956, K. Morimoto (ELKU). 1 male and 1 female, Misawa, Yachigashira, 7-VI-2007, K. Morimoto (NIAES). 1 male, Hachinohe, Tanesashi-kaigan, 10-VI-2007, K. Morimoto (NIAES). IWATE. 1 male and 2 females, Miyako, Kamegamori, 15-VI-1986, K. Emoto (NIAES). 2 females, Sawauchi, Yasugasawa, 31-V-1998, H. Yoshitake (NIAES). 2 males and 1 female, Hanamaki, Toyosawa-rindou, 338–313 m, 39°28'36.2"–39°29'08.6"N, 140°56'15.7"–140°56'32.9"E, 8-VI-2007, H. Yoshitake, on R. crataegifolius (NIAES). 1 male and 1 female, Kawai, Yoshibezawa, 9-VI-2007, K. Morimoto (NIAES). 1 male, Kuji, Ootsuki-touge Pass, 10-VI-2007, K. Morimoto (NIAES). 1 male and 3 females, Kawai, Tsuchisaka-touge Pass, 12-VI-2007, J. Kantoh (NIAES). 1 male, Matsukusa, 800 m, 21-VI-1989, M. J. Sharkey (CMN). 3 males and 2 females, “Mt. Hyacinthe” (most probably an error in writing Mt. Hayachine), 500 m, 21-VI-1989, M. J. Sharkey (CMN). MIYAGI. 1 male and 3 females, Marumori, Fudou, 5-V-1994, H. Yoshitake (NIAES). 1 female, Shichigashuku, Inago, 20-V-1994, H. Yoshitake (NIAES). 1 female, Shiroishi, Mizubashounomori, 27-VI-1994, H. Yoshitake (NIAES). AKITA. Lake Tazawako: 2 females, 4-VI-1969, S. Miyakawa (SM); 3 males, 12-VI-1971, S. Miyakawa (SM); 1 male and 1 female, 15-VI-1971, S. Miyakawa (SM). 3 males and 2 females, Karabuki-shitsugen, Kuroyu, 12-VI-1974, S. Miyakawa (SM). 2 females, Hiyagata–Kunimi-onsen, 16-VI-1974, S. Miyakawa (SM). 3 males and 2 females, Senboku, Nishiki, Ainai, 10-VI-1983, S. Miyakawa (SM). YAMAGATA. 1 female, Yonezawa, Kariyasu, 15-VI-1975, S. Miyakawa (SM). 5 males and 4 females, Mt. Zaousan, Yoshikari-rindou, 18-VI-1983, S. Miyakawa (SM). 1 male, Oguni, Kotamagawa, 300 m, 6-VI-2005, H. Hirano (NIAES). 1 female, Oguni, Tamagawa, 120–150 m, 6-VI-2005, H. Hirano (NIAES). FUKUSHIMA. 1 female, Kamitoriwata, 20-VI-1976 (NIAES). Iwaki, Eda: 2 males and 1 female, 25-V-1980, Y. Shiozaki (YS). 3 males and 3 females, Minamiaizu, Funamata-rindou, 20-V-1983, S. Miyakawa (SM). 2 males, Namie, 30-IV-1994, H. Yoshitake (NIAES). Haranomachi, Yokokawa, Akane-rindou: 1 male, 15-VI-1984, K. Kinugasa (NIAES); 1 male, 31-V-1987, H. Ebihara (NIAES). Haranomachi, Kozikiishi-rindou: 11 males and 28 females, 4-VI-1988, S. Miyakawa (SM); 1 male, 4-VI-1988, S. Miyakawa (NIAES); 14 males and 9 females, 5-VI-1988, S. Miyakawa (SM). Hinoemata: 1 male, 14-VI-1980, Y. Shiozaki (YS); 1 female, 20-VI-1990, H. Kojima (ELKU); 2 females, Mikawa-rindou, 24-VI-1990, K. Yoshihara (SM). 1 male and 1 female, Nishigou, Yukiuribashi, 3-V-2002, S. Mizunoya (NIAES). 1 female, Nishigou, Daijou, 22-V-2002, S. Mizunoya (NIAES). Nishigou, Mabune: 1 male and 2 females, 650 m, 3-VI-2005 (NIAES), H. Hirano; 1 male and 1 female, 770 m, 3-VI-2005, H. Hirano (NIAES); 6 males and 8 females, 3-VI-2005, H. Yoshitake, on Rubus sp. (NIAES). IBARAKI. 3 males and 7 females, Mt. Yamizosan, Shimokitazawa, 28-V-1988, Y. Kurosawa (SM). 1 female, Takahagi, 29-V-1996, H. Takizawa (NIAES). TOCHIGI. 1 female, Mt. Sukaizan, 4–5-VII-1971, H. Takizawa (NIAES). 1 male and 1 female, Nishinasuno, 8-V-1977, S. Miyakawa (SM). 4 males and 3 females, Nikko, Kashiwagi, 16-V-1982, S. Miyakawa (SM). 1 male, Nikko, Nebazawadani, 19-VII-1940, S. Miyakawa (SM). 1 male, Fujiwara, Midorizawa-rindou, 21-V-1989, S. Miyakawa (SM). 1 female, Nasu, Toyahara, 13-VII-1996, H. Takizawa (NIAES). 1 female, Nasu, Mt. Minamigassan, 20-VII-1996, H. Takizawa (NIAES). 2 males and 1 female, Nasu-kougen, 1200–1500 m, 7-VI-2005, H. Hirano (NIAES). 4 males and 3 females, Nishinasuno, N. G. R. I., 500 m, 10-VIII-1989, M. J. Sharkey (CMN). GUNMA. 1 male and 1 female, Utsunomiya, 6-V-1968, H. Takizawa (NIAES). 2 females, Mt. Hotakayama, 13–15-VII-1975, H. Irie (ELKU). 1 female, Akagi, 21-V-1967 (SM). 1 male, Akagi, Akagi-shizenen, 9–10-V-1988, S. Saito (NIAES). 1 female, Akagi, Mt. Akagisan, 3-VI-1997, T. Ishikawa (NIAES). 1 male, Oku-tone, Lake Fujiwarako, 13-V-1990, S. Tsuyuki (NIAES). 4 males and 5 females, Niiharu, Amemi-rindou, 4-V-1998, S. Arai (NIAES). 5 males and 5 females, near Mt. Aneyama, 19-V-2001, S. Arai (NIAES). Matsuida, Kirizumi-onsen: 1 female, 27-V-1989, K. Matsumoto (NIAES); 1 male and 1 female, 27-V-1989, H. Kojima (NIAES); 1 male and 1 female, 17-V-1998, S. Arai (NIAES). SAITAMA. 2 males and 2 females, Shoumaru-touge Pass, 9-V-1976, S. Miyakawa (SM). 1 male, Minano, Shimohinozawa, Sawabe, 1-VI-1984, S. Miyakawa (SM). 1 female, Ootaki, near Taiyouji, 4-VII-1999, S. Arai (NIAES). CHIBA. 4 males and 2 females, Mt. Kiyosumiyama–Kenminnomori, 31-III-1979, J. Okuma (SM). 1 female, Bousou-hantou Peninsula, Mt. Kiyosumiyama, 30-IV-1991, S. Tsuyuki (NIAES). TOKYO. 1 male, Mitakadai, 30-V-1941, S. Miyakawa (SM). 1 male and 1 female, Mitaka, 25-V-1948, S. Miyakawa (SM). Mt. Oodakeyama: 1 male and 2 females, 21-V-1942, S. Miyakawa (SM); 4 males and 2 females, 24-V-1942, S. Miyakawa (SM). 1 female, Mt. Mitakesan, 15-V-1966 (SM). 1 female, Itsukaichi, Kariyose, 20-IV-1968, Y. Hasegawa (SM). Mt. Mitousan: 1 female, 4-V-1968, H. Takizawa; 1 male and 1 female, 27-V-1968, H. Takizawa (NIAES). 2 males and 2 females, Mt. Jinbasan, 6-V-1974, S. Miyakawa (SM). 1 male, Hinohara, Yazawa-rindou, 18-V-1997, H. Yoshitake (NIAES). Hachiouji: 3 males and 1 female, 1-V-1938, S. Miyakawa (SM); 1 male and 1 female, 1-V-1966 (SM); 2 males, 1-V-1966, S. Miyakawa (SM). Hachiouji, Uratakao, Kogesawa-rindou: 1 male, 30-IV-1938, S. Miyakawa (SM); 2 males and 2 females, 350–550 m, 35°38'30"–35°39'27" N, 139°11'57"–139°14'30" E, 17-V-2005, H. Yoshitake, on R. crataegifolius (NIAES). Okutama: 1 female, 9-V-1968, Hasegawa (SM); 1 male, 29-IV-1980, Y. Shiozaki (YS). 3 males and 4 females, Okutama, Kurasawadani, 6-V-1948, S. Miyakawa (SM). 2 males and 5 females, Okutama, Nippara, 24-V-1970, K. Unno (SM). 1 male and 1 female, Okutama, Unazawa, 7-V-1982, K. Shiozaki (YS). 2 males and 1 female, Okutama, Asamaone, 2-V-1992, K. Matsumoto (NIAES). KANAGAWA. 1 male, Yokohama, Motomachi, 29-VI-1970, S. Miyakawa (SM). 2 females, Hakone, Mt. Daigatake, Kozuka, 24-V-1979, S. Miyakawa (SM). Hadano, Mizunashigawa: 1 male, 29-IV-1983, K. Shiozaki (YS); 1 male, 29-IV-1983, Y. Shiozaki (YS). 1 female, Hadano, Yabitsu-touge Pass, 30-V-1983, K. Shiozaki (YS). 1 male, Yamakita, Mikuni-touge Pass, 23-V-1994, Y. Notsu (NIAES). 1 female, Tsukui, Kaminokawa-rindou, 21-V-1995, Y. Goto (NIAES). NIIGATA. 1 male, Kamikawa, 3-VI-2000, K. Takahashi (NIAES). FUKUI. 2 males and 5 females, Kanazu, Mt. Kariyasuyama, 20-V-1989, S. Inoue (ELKU). YAMAMASHI. 1 female, Mt. Mishotaisan, 17-VII-1940, S. Miyakawa (SM). 1 male and 1 female, Shirosu, 7-VI-1958 (ELKU). 1 female, Uenohara, 17-IV-1968 (SM). 1 male, Kawamura, Akiyama, 4-V-1969, S. Miyakawa (SM). 1 male, Lake Saiko, 27-V-1970, K. Unno (SM). 1 female, Ootsuki, Tomioka, 17-V-1976, S. Miyakawa (SM). 14 males and 19 females, Ootsuki, Koganezawa-rindou, 30-V-1980, J. Okuma (SM). 1 female, Mt. Fujisan, Subaru-line, 2000 m, 3-VI-1977, S. Miyakawa (SM). 1 female, Mt. Yatsugatake, 26-V-1979, A. Seki (NIAES). 16 males and 14 females, Koganezawa, 2-VI-1980, S. Miyakawa (SM). 1 female, Daibosatsu-touge Pass, 8-VI-1968, Y. Hasegawa (SM). Enzan, Hikawa-rindou: 1 male, 1-VII-1982, M. Sawai (SM); 1 male and 1 female, 2-VII-1982, M. Sawai (SM). 1 female, Mt. Kushigatayama, Maruyama-rindou, 23-IV-1984, S. Tsuyuki (NIAES). 2 males, Akeno, Manjuu-touge Pass, 6-VI-1986, S. Miyakawa (SM). 1 male, Kanayama, 1-VII-1989, T. Nonaka (NIAES). Ashiyasu, Hirogawara: 1 male, 24-V-1969, Y. Hasegawa (SM); 1 female, 8-VI-1989, H. Kojima (NIAES); 1 male and 1 female, 1529 m, 17-VI-1990, S. Miyakawa (SM); 1 female, 20-VI-1993, T. Horiguchi (NIAES). 1 male and 1 female, Nirasaki, Mt. Kayagatake, 16-V-1996, S. Miyakawa (SM). 1 female, Tabayama, Sanjounoyu, 20-V-1999 (NIAES). NAGANO. 1 female, Mt. Nyuukasayama, 6-VI-1942, S. Miyakawa (SM). 1 male and 3 females, Nagano, Uematsu, VI-1957, K. Matsuo (SM). 1 male and 1 female, Ina, Onasawa, 6-VI-1962, K. Oshima (ELKU). 1 male, Shimajimadani, VI-1970, N. Nino (SM). 1 female, Tobira-onsen, 26-VI-1974, H. Hayakawa (SM). 1 female, Kisofukushima, Higashiyama, 19-VII-1986, K. Matsui (SM). 1 female, Kisofukushima, 21-VIII-1988, K. Matsui (SM). 1 female, Kisofukushima, Uyama, 29-VIII-1988, K. Matsui (SM). 1 male, Kisofukushima, Kibio, 18-IX-1988, K. Matsui (SM). 1 male and 1 female, Kisokoma-kougen, Kibio, 21-VIII-1989, K. Matsui (SM). 1 male and 2 females, Nakakawa, 1-V-1998, H. Yoshitake (NIAES). 1 female, Hara, Pension Village, 2–4-VII-1998, H. Yoshitake (NIAES). GIFU. 1 male, Mt. Hakusan, 29-VI-2002, H. Hirano (NIAES). 1 female, Takane, Hiwada-kougen, 1-X-1989, K. Matsui (SM). 6 exs., Toki, 12-V-2010, K. Izawa, on R. trifidus (KI). SHIZUOKA. 1 female, Shinfuji, 30-IV-1989, N. Niwa (NIAES). 1 female, Honkawane, Sessokyou, 9–10-V-1992, T. Kishimoto (NIAES). Izu-hantou Peninsula: 2 males and 1 female, Kyuu-amagi-touge Pass, 3-V-1972, S. Miyakawa (SM); 2 males and 3 females, Mt. Nekkodake, 19-V-1974, S. Miyakawa (SM); 21 males and 16 females, Okuhara-rindou, 22-VI-1977, J. Okuma (SM); 2 females, Mt. Toogasayama, 17-V-1980, J. Okuma (SM); 1 male, Izu Okawa, 3-VI-1989, H. Takizawa (NIAES). AICHI. 8 males and 6 females, Kasugai, Hosono, 1-V-1991, H. Kojima (ELKU). Toyota, on R. trifidus: 5 exs., Oobora, Nikake-rindou, 3-V-2010, K. Izawa (KI); 6 exs., Nishiichinonochou, 8-V-2013, K. Izawa (KI); 7 exs., Asugawachou, Asugawa-hoan-rindou, 8-V-2013, K. Izawa (KI). MIE. Misugi, Hirakura: 1 male, 2-VI-1985, T. Imamura (NIAES); 3 females, 600–700 m, 15–17-V-2005, H. Hirano, on R. crataegifolius (NIAES). 1 male, Fujigawa, Mt. Suzugadake, 600 m, 3-V-2002, S. Arai (NIAES). 4 exs., Komonochou, Komono, 15-V-2013, K. Izawa, on R. microphyllus (KI). SHIGA. 1 male and 1 female, Mt. Ibukiyama, 220 m, 3-VI-1990, S. Miyakawa (SM). KYOTO. 1 male, Sugi-touge Pass, 29-V-1977, M. Sawai (SM). HYOGO. 1 male, Haga, Akasai-keikoku, 3-V-1993, S. Tsuyuki (NIAES). NARA. 1 female, Gose, Mt. Kongousan, 1100 m, 4-VII-1998, T. Kishimoto (NIAES). WAKAYAMA. 1 male and 1 female, Mt. Gomadanzan, 7–8-VI-1997, T. Ito (NIAES). 1 female, Nakahechi, Mizukami, 28-IV-2003, H. Hirano (NIAES). 1 male, Arida, Kamiyukawa, 19-V-1973, I. Matoba (ELKU). TOTTORI. 1 male and 1 female, Wakasa, Mt. Hyounosen, 7-VI-1987, K. Yoshihara (SM). SHIMANE. 1 female, Okinoshima I., Mt. Takuhiyama, 13-V-1975, J. Okuma (SM). OKAYAMA. 4 males and 1 female, Yubara, Yubara-onsen, 21-V-1991, H. Nakamura (NIAES). 1 male, Hokubou, Kamimizuta, 8-V-1982, K. Yoshihara (SM). 1 female, Iwayadani, 4-V-1981, K. Yoshihara (SM). 1 male and 1 female, Kawakami, Myouren-keikoku, 27-VI-1982, K. Yoshihara. HIROSHIMA. 1 female, Kouzan, Bessako, 10-V-1996 (ELKU). 1 female, Yoshiwa, 11-V-1976, K. Baba (ELKU). Shikoku. TOKUSHIMA. 1 male, Sanuki Mts., Mt. Ryuuousan, 17-VIII-1989, K. Matsumoto (NIAES). EHIME. 5 males and 5 females, Omogokei, 18–19-V-1968, K. Hatta (EUMJ). 1 male, Iyomishima, Tomisato, 3–5-VIII-1974, G. Tokihiro (EUMJ). 4 males and 6 females, Houjou, Kukawa, 22-IV-1977, A. Oda (EUMJ). Kyushu. FUKUOKA. Mt. Hikosan: 1 male, 18-VI-1965, K. Takeno (ELKU); 1 female, 12-VI-1965, S. Kimoto (ELKU); 1 female, 17-V-1971, M. T. Chujo (ELKU); 2 males and 1 female, 30-V-1995, K. Morimoto (ELKU). 1 male, Mt. Tachibanayama, 23-IV-1997, H. Kojima (ELKU). SAGA. 1 female, Sefuri Mts., 28-IV-2002, H. Hirano (NIAES). 16 males and 13 females, Mt. Kusenbuyama, Hakotani-rindou, 8-V-2005, H. Yoshitake, on R. crataegifolius (NIAES). NAGASAKI. Tsushima I.: 1 female, Mt. Ooboshiyama, 16-VII-1981, Y. Syouno (ELKU); 1 male, Izuhara, Mt. Ariakeyama, 5–9-V-1997, H. Yoshitake (NIAES). 2 males and 1 female, Tashirobaru, 29-IV-1979, S. Imasaka (NIAES). 1 male, Taradake Mts., Mt. Gokaharadake, 12-VI-1979, S. Imasaka (NIAES). Shimabara, Sanbuki: 1 male and 1 female, 7-V-1976, S. Imasaka (NIAES); 1 male, 10-V-1976, S. Imasaka (NIAES); 6 females, 17-V-1976, S. Imasaka (NIAES); 1 female, 21-V-1976, S. Imasaka (NIAES); 1 male and 2 females, 22-V-1976, S. Imasaka (NIAES). Shimabara, Akamatsudani: 3 females, 24-V-1980, S. Imasaka (NIAES); 1 male, 27-V-1980, S. Imasaka (NIAES). 2 females, Sasebo, Mt. Hattendake, 23-V-1982, S. Miyakawa (SM). 3 males and 2 females, Kawatana, Kobagou, 27-V-1984, J. Okuma (SM). OITA. Kuju Mts., Mt. Kurodake: 1 female, 29-VII-1979, S. Imasaka (NIAES); 1 male, 9-VI-1985, K. Konishi (ELKU); 2 males, Oike-enchi, 7-VI-2002, H. Hirano (NIAES). 1 female, Kuju Mts., Jizoubaru, 28-V-1987, K. Morimoto (ELKU). 1 male, Kuju Mts., Bougatsuru, 27-V-1988, K. Morimoto (ELKU). 1 male and 2 females, Kuju Mts., Choujabaru, 22-V-1999, K. Morimoto (ELKU). MIYAZAKI. 1 male, Ebino, Shiratori-onsen, 30-IV-1991, K. Morimoto (ELKU).
China (Liaoning and Heilongjiang – new record), Korea (North – new record, Central, South, and Ulreungdo Island), Japan (Hokkaido, Honshu, Izu Islands, Shikoku, Kyushu, and Tsushima Island) (Fig. 133).
Adults of this common species were collected from Rubus idaeus L. subsp. melanolasius Focke f. concolor (Kom.) Ohwi in Korea (
http://zoobank.org/C8AC65A6-71E7-4CD0-9387-CFA00B598B09
The species is very similar to Scleropteroides hypocrita but is distinguished from it by the following characteristics: subapical part of the prothorax strongly constricted (Fig. 36); elytra with strongly prominent humeri, straightly convergent toward the subapical calli, (Fig. 37); scales in a row on elytral intervals semierect, nearly as long as the width of each interval (Fig. 37); apical half of the male rostrum slightly more widened (Fig. 26); male metaventrite more prominent ventrally along the apico-lateral margin of the metaventral receptacle; male ventrite I with no prominence; penis with a longer apical projection (Figs 71, 81); endophallus with a pair of narrow falcate sclerites in the basal part (Figs 70, 80); female sternite VIII with arms that are divergent apically (Figs 77, 87); bursa copulatrix without minute coniform spicules in the posterior part (Figs 75, 85).
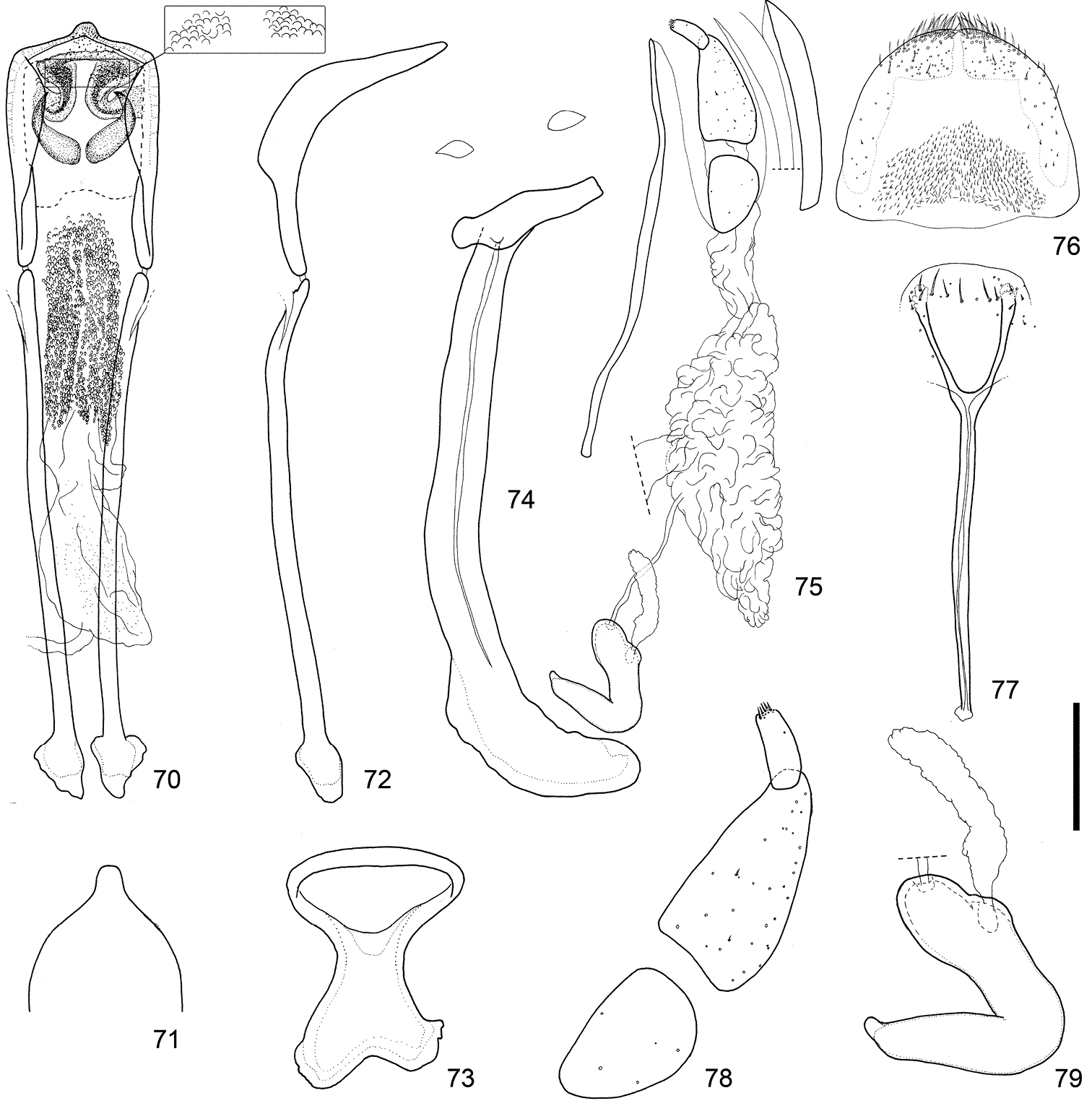
Male and female terminalia of Scleropteroides longiprocessus Huang & Yoshitake, sp. n. from Mt. Kusenbuyama, Kyushu, Japan. 70 Aedeagus, dorsal view 71 Apex of the penis, dorsal view 72 Aedeagus, lateral view 73 Tegmen 74 Sternites VIII and IX, male 75 Female terminalia and genitalia, lateral view 76 Tergite VIII, female 77 Sternite VIII, female 78 Coxite and stylus 79 Spermatheca. Scale: 0.20 mm for 70–77, 0.10 mm for 78–79.
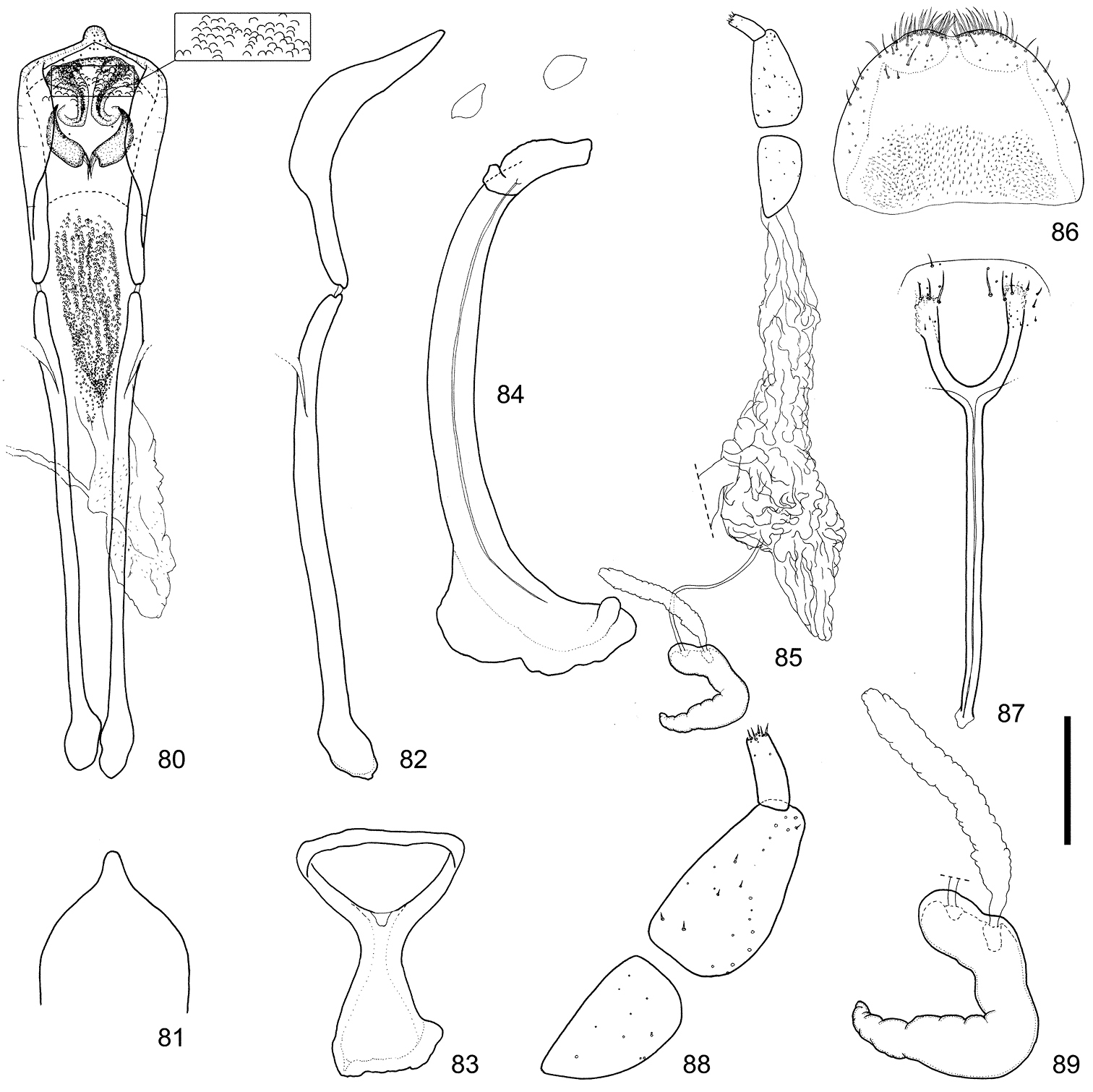
Male and female terminalia of Scleropteroides longiprocessus Huang & Yoshitake, sp. n. from Tochigi, Honshu, Japan. 80 Aedeagus, dorsal view 81 Apex of the penis, dorsal view 82 Aedeagus, lateral view 83 Tegmen 84 Sternites VIII and IX, male 85 Female genitalia, lateral view 86 Tergite VIII, female 87 Sternite VIII, female 88 Coxite and stylus 89 Spermatheca. Scale: 0.20 mm for 80–87, 0.10 mm for 88–89.
LB: 2.27–2.63 (mean, 2.39). LR: 0.97–1.08 (mean, 1.04). WP: 0.95–1.00 (mean, 0.97). LP: 0.76–0.85 (mean, 0.82). WE: 1.49–1.62 (mean, 1.57). LE: 1.53–1.66 (mean, 1.62). N = 10 for all measurements. Habitus as shown in Figs 3–4.
Vestiture. Clavate scales slightly longer and semierect on head, basal 2/3 of rostrum, and pronotum. Scales on elytral intervals (Fig. 37) semierect, slightly longer, 0.9–1.2 × as long as interval width.
Rostrum (Figs 26–27) slender, 1.21–1.32 × as long as pronotum; sides more strongly widened in apical part, which is 1.23 × as wide as basal part. Antennae (Fig. 27) with length ratio of funicular segments I: II: III: IV: V: VI = 1.88: 1.54: 1.12: 1.00: 0.96: 0.96 and width ratio = 1.73: 1.00: 1.36: 1.45: 1.45: 1.55.
Pronotum (Fig. 36) 1.13–1.27 × as wide as long, 0.47–0.52 × as long as and 0.59–0.65 × as wide as elytra; subapical constriction stronger; sides subparallel along basal half, then rapidly convergent toward apex.
Elytra 1.01–1.04 × as wide as wide, 1.92–2.14 × as long as and 1.54–1.69 × as wide as pronotum, straightly convergent toward subapical calli; humeral calli slightly more prominent; subapical calli moderately prominent.
Underside. Metaventrite moderately prominent ventrally along apico-lateral margin of metaventral receptacle. Ventrite I weakly flattened on disc, lacking prominence; length ratio of ventrites I: II: III: IV: V = 4.33: 2.00: 1.00: 1.13: 2.22 and width ratio = 1.89: 1.70: 1.23: 1.11: 1.00.
Genitalia. Tegmen (Figs 73, 83) with apodeme strongly widened toward apex. Penis (Figs 70–72, 80–82) thick in profile; sides more or less weakly widened from base to apical 1/5, strongly convergent apically; apical projection more slender. Endophallus (Figs 70, 80) with pair of narrow falcate sclerites in basal part.
Otherwise as in Scleropteroides hypocrita.
LB: 2.08–2.53 (mean, 2.34). LR: 0.97–1.15 (mean, 1.09). WP: 0.90–1.07 (mean, 0.97). LP: 0.74–0.86 (mean, 0.81). WE: 1.43–1.64 (mean, 1.54). LE: 1.46–1.68 (mean, 1.59). N = 10 for all measurements.
Rostrum (Figs 28–29) slightly more slender, 1.30–1.39 × as long as pronotum.
Pronotum 1.14–1.28 × as wide as long.
Elytra 1.01–1.08 × as long as wide.
Underside. Ventrite V with small median concavity, which is sometimes obscure. Sternite VIII (Figs 77, 87) with arms slightly more slender and apically divergent.
Genitalia. Bursa copulatrix (Figs 75, 85) lacking spicules. Coxites (Figs 78, 88) nearly 5.0 × as long as styli; styli more slender, nearly three times as long as wide. Spermatheca (Figs 79, 89) with collum more strongly convex; insertions of duct and gland slightly more distant from each other.
Otherwise showing almost the same sexual dimorphism as in Scleropteroides hypocrita.
HOLOTYPE: 1 male (NIAES), “[Japan: Kyushu]/Mts. Sefuri-sanchi/Mt. Kusenbu-yama/8-V-2005/Hiraku Yoshitake” (printed on white card); “On Rubus palmatus Thunb. (Rosaceae)/[JN: Nagaba-momiji-ichigo] with flowers” (print ed on white card); “[ HOLOTYPE ] Male/Scleropteroides/longiprocessus/Huang & Yoshitake, 2008” (typed on red card). PARATYPES. JAPAN: Honshu. IWATE. 1 male and 1 female, Miyako, Genbehdaira, 9-VI-2007, K. Morimoto (NIAES). 2 males and 6 females, Tsuchisaka-touge Pass, Kawai, 12-VI-2007, J. Kantoh (NIAES). MIYAGI. 1 male, Naruse, Miyatojima I., V-1995, M. Kodama (NIAES). 1 male and 2 females, Sendai, Mt. Banzan, 20-V-1995, H. Yoshitake (NIAES). AKITA. 1 male and 1 female, Lake Tazawako, 15-VI-1974, S. Miyakawa (SM). YAMAGATA. 2 males and 3 females, Mt. Zaousan, Yoshikari-rindou, 18-VI-1983, S. Miyakawa (SM). FUKUSHIMA. Haranomachi, Kozikiishi-rindou: 1 male, 30-V-1987, H. Ebihara (NIAES); 1 male, 5-VI-1988, S. Miyakawa (SM). 1 female, Haranomachi, Akanesawa-rindou, 5-VI-1988, S. Miyakawa (SM). 1 female, Namie, 30-IV-1994, H. Yoshitake (NIAES). 1 male, Iizaka, Moniwa, Yakematsu, 1-V-1997, S. Saito (NIAES). IBARAKI. Mt. Yamizosan, Shimokitazawa: 7 males and 1 female, 1022 m, 28-V-1988, S. Miyakawa (SM); 1 male and 2 females, 28-V-1988, Y. Kurosawa (SM). TOCHIGI. 1 male and 2 females, Nikko, Kashiwagi, 16-V-1982, S. Miyakawa (SM). 1 male and 1 female, Shiobara, Hikinuma, 19-V-1989, S. Miyakawa (SM). 3 males and 1 female, Mt. Shibakusayama, 18-V-1990, H. Kojima (ELKU). GUNMA. 1 male and 1 female, Matsuida, Kirizumi-onsen, 16-V-1998, S. Arai (NIAES). 2 males and 2 females, Niiharu, Amemi-rindou, 4-V-1998, S. Arai (NIAES). 1 female, Fujiwara, Nakanosawa-rindou, 19-V-1989, S. Miyakawa (SM). 6 males and 4 females, Fujiwara, Midorizawa-rindou, 29-V-1989, S. Miyakawa (SM). NIIGATA. Kurokawa: 1 female, 28-IV-1973, K. Baba (ELKU); 2 females, 26-V-1976, K. Baba (ELKU); 1 female, 24-V-1980, K. Baba (ELKU). 1 male and 1 female, Teradomari, Enjouji, 21–23-V-1996, K. Ishida (NIAES). 1 male, Sadogashima I., Nagaishihama, 22-VII-1970, K. Baba (ELKU). FUKUI. 2 males and 1 female, Obama, Shimonegouri, 6-V-1979, H. Sasaji (ELKU). YAMANASHI. 1 female, Mt. Fujisan, Fuji-rindou, 23-VI-1983, S. Miyakawa (SM). 1 male, Ootsuki, Nanaho, 9-V-1976, S. Miyakawa (SM). 1 female, Ootsuki, Koganezawa-rindou, 30-V-1980, J. Okuma (SM). NAGANO. 1 female, Tobira-onsen, 26-VI-1985, H. Hayakawa (SM). 5 males, Nagiso, 29-IV-1998, H. Yoshitake (NIAES). 2 males and 1 female, Iida, Hatouchi-rindou, 29–30-IV-1998, H. Yoshitake (NIAES). 2 males, Nakakawa, 1-V-1998, H. Yoshitake (NIAES). GIFU. Nakatsugawa, Kamisaka: 1 female, 13-VI-1987, S. Miyakawa (SM); 1 female, 13-VI-1987, K. Morimoto (ELKU). 5 exs., Nakatsugawa, Nenoue-kougen, 6-V-2013, K. Izawa, on R. palmatus var. coptophyllus (KI). SHIZUOKA. 1 female, Shinfuji, 30-IV-1989, N. Niwa (NIAES). 5 males and 6 females, Nakaizu, Mt. Amagisan, 3-V-1990, H. Kojima (ELKU). 1 male and 4 females, Mt. Ooyama, 5-V-1993, Y. Notsu (NIAES). 3 males and 1 female, Shizuoka, Umegashima, 15-V-1993, T. Kishimoto (NIAES). Izu-hantou Peninsula: 3 males and 1 female, Hacchouike Pond, Amagi-touge Pass, 28-V-1972, S. Miyakawa (SM); 1 female, Kamo, Oogusu, 11-V-1980, J. Okuma (SM); 1 female, Hacchouike Pond–Kantenbashi, 14-V-1980, J. Okuma (SM). 1 male, Umegashima, 15–16-V-1993, H. Sato (NIAES). 1 female, Shizuoka, Nakaizu, Amagi-kougen, 31-V-1987, S. Miyakawa (SM). AICHI. 6 exs., Shitara, Mt. Nishinagura-Iyama, 29-V-2010, K. Izawa, on R. palmatus var. coptophyllus (KI). Toyota, on R. palmatus var. coptophyllus: 5 exs., Nishiichinonochou, 8-V-2013, K. Izawa, (KI); 4 exs., Asugawachou, Asugawa-hoan-rindou, 8-V-2013, K. Izawa (KI); 2 exs., Inabuchou, Noirigawa, 22-V-2013, K. Izawa (KI). SAITAMA. 1 female, Mt. Kasayama, 26-V-1967, H. Takizawa (NIAES). 1 male, Saitama Yokote, 28-IV-1974, S. Miyakawa (SM). 1 female, Lake Miyazawako, 11-V-1975, S. Miyakawa (SM). 1 male, Shoumaru-touge Pass, 9-V-1976, S. Miyakawa (SM). CHIBA. 1 female, Mt. Kiyosumiyama–Kenminnomori, 31-V-1979, J. Okuma (SM). TOKYO. 1 female, Okutama, Kurasawadani, 19-VI-1938, S. Miyakawa (SM). Nishitama, Mt. Oodakeyama: 1 male, 21-V-1942, S. Miyakawa (SM); 2 males and 1 female, 15-V-1966, S. Miyakawa (SM). Hachiouji, Mt. Takaosan: 1 female, 30-V-1980, Y. Shiozaki (YS); 1 male and 1 female, 23-VI-1996, H. Yoshitake (NIAES); 5 exs., 22-V-2010, K. Izawa, on R. palmatus var. coptophyllus (KI). Hachiouji, Mt. Takaosan, Kogesawa-rindou: 3 males, 14-V-1944, S. Miyakawa (SM); 1 male and 1 female, 1–2-V-1999, H. Yoshitake (NIAES). 5 males and 5 females, Hachiouji, Mt. Takaosan, Path 6, 240–599 m, 35°37'30"–35°37'50"N, 139°14'36"–139°15'43"E, 15-V-2005, R. palmatus var. coptophyllus, H. Yoshitake (NIAES). 1 female, Hachiouji, Kitano, 14-IV-1968, S. Miyakawa (SM). 1 female, Inagi, Yomiuri Land, 17-V-1974, S. Miyakawa (SM). KANAGAWA. 1 male, Mikuni-touge Pass, 7-VI-1970, H. Takizawa (NIAES). 1 female, Mt. Ooyama, 2-VI-1981, S. Miyakawa (SM). 2 males, Yokohama, Nakayama, 3-V-1983, S. Miyakawa (SM). 1 female, Hakone, Oowakudani, 15-VI-1985, S. Tsuyuki (NIAES). Hadano: 1 female, Mt. Koubouyama, 24-IV-1983, Y. Shiozaki (YS); 1 male, 30-IV-1990, H. Takizawa (NIAES). 1 female, Tanzawa Mts., Mizunashigawa, 30-IV-1983, Y. Shiozaki (YS). Hadano, Yabitsu-touge Pass: 1 female, Yabitsu, 1-VII-1983, K. Shiozaki (YS); 1 female, 29-IV, K. Suzuki (ELKU). YAMANASHI. 1 female, Akaishi-sanmyaku Range, Norogawa-rindou, 5-VI-1983, Y. Shiozaki (YS). MIE. 1 female, Ninomata–Owase, 26-VI-1960, H. Ichihashi (NIAES). 1 male, Komono, Unbomine, 28-IV-1985, A. Amagasu (ELKU). 1 female, Shiga, Mt. Ibukiyama, 10-VI-1990, H. Takizawa (NIAES). 1 female, Toba, Mastuno, Mt. Aonomineyama, 19-V-1996, H. Yoshitake (NIAES). 8 males and 6 females, Toba, Toushijima I., 10-IV-1996, K. Ishida (NIAES). 1 male, Misugi, Hirakura, 7–8-VI-1997, H. Yoshitake (NIAES). NARA. 1 female, Kawakami, Mt. Wasamatayama, 8-VII-1995, A. Yoshida (NIAES). 1 male, Mt. Miyama, 7-VII-1989, S. Yamada (NIAES). OKAYAMA. 2 males and 1 female, Kamo, Kurami, 29-VI-1985, K. Watanabe (SM). 2 females, Tetta, Mt. Aratoyama, 16-V-1982, K. Yoshihara (SM). 1 male, Takahashi, Mt. Gagyuusan, 5-V-1984, K. Yoshihara (SM). Izu Islands. 1 female, Kouzushima I., Maehama–Tsuzukizawa, 9-V-1979, J. Okuma (SM). Shikoku. TOKUSHIMA. 1 female, Haranomachi, 8-VI-1980, K. Shiozaki (YS). 1 male and 1 female, 1-VI-1986, A. Watanabe (YS); 1 female, 5-VI-1986, A. Watanabe (YS). EHIME. 2 females, Imabaru, Mt. Takanawazan, 5-V-1976, S. Inoue (EUMJ). KOCHI. 1 male, Okunanokawa, 4-V-1967, H. Oonishi (EUMJ). Cape Murotomisaki: 1 male and 1 female, 7-VI-1959, M. Miyatake (EUMJ); 1 female, 8-V-1959, S. Hisamatsu (EUMJ); 2 females, 12-VII-1961, M. Miyatake (NIAES). Kyushu. FUKUOKA. 1 male, Chikugo, Mt. Kumadoyama, 1-VI-1958, Y. Miyaue (ELKU). 1 female, Munakata, Mt. Jouyama, 6-IV-1975, K. Kido (ELKU). 1 female, Itoshima, Himeshima I., 3-VI-1990, K. Kido (ELKU). 1 female, Shikanoshima I., 29-IV-1994, K. Morimoto (ELKU). 1 female, Fukuoka, Motooka, 2-VII-1994, K. Morimoto (ELKU). 1 male and 1 female, Mt. Sefurisan, 3-VI-2001, H. Hirano (NIAES). 1 male, Yabe, Mt. Shakagatake, 25-IV-2004, H. Hirano (NIAES). 3 males and 1 female, Nokonoshima I., 17-VI-2003, H. Hirano (NIAES). Mt. Hikosan: 2 males and 3 females, 17–19-V-1967, H. Takizawa (NIAES); 1 female, 4-V-1985, Y. Kajida (NIAES); 2 males and 4 females, 30-V-1995, K. Morimoto (ELKU); 1 female, 26-VI-2003, H. Hirano (NIAES). SAGA. 1 male, Okuhiratani, Mt. Taradake, 23-IV-1989, T. Yasunaga (ELKU). 2 males and 3 females, same data as holotype (NIAES). 2 males and 2 females, Mt. Kusenbuyama, Hakotani-rindou, 8-V-2005, H. Yoshitake, on R. crataegifolius (NIAES). NAGASAKI. 1 male, Shimabara, Taruki, 16-VII-1976, S. Imasaka (NIAES). 1 female, Shimabara, Akamatsudani Gorge, 8-VI-1979, S. Imasaka (NIAES). Mt. Unzendake: 1 female, 25-V-1976, S. Imasaka (NIAES); 1 female, 21-VII-1976, S. Imasaka (NIAES). 1 male and 2 females, Nomozeki, 13-V-1976, S. Imasaka (NIAES). 1 male, Tashirobaru, 21-IV-1977, S. Imasaka (NIAES). 15 males and 12 females, Sasebo, Mt. Eboshidake, 13-IV-1977, J. Okuma (SM). 4 males, Sasebo, Mt. Hattendake, 23-V-1982, J. Okuma (SM). Sasebo, Mt. Yahirodake: 1 male and 1 female, 18-IV-1977, J. Okuma (SM); 2 males, 6-IV-1981, J. Okuma (SM); 1 female, 21-V-1981, J. Okuma (SM); 1 female, 26-V-1982, J. Okuma (SM); 1 female, 29-III-1983, J. Okuma (SM); 1 female, 30-IV-1983, J. Okuma (SM); 1 male, 2-V-1983, J. Okuma (SM); 2 females, 15-V-1984, J. Okuma (SM). 1 male and 1 female, Hiradoshima I., Mt. Yasumandake, 10-V-1980, S. Imasaka (NIAES). KUMAMOTO. 1 male, Gokanoshou, 26–27-V-1978, K. Ohara (ELKU). Izumi, Mt. Shiratoriyama: 1 female, 1300 m, 6-VI-1980 (ELKU); 1 female, 7–8-VI-1989, T. Yasunaga (ELKU). 1 male and 2 females, Yabe, Naidaijin-rindou, 21-VI-1998, H. Kojima (ELKU). OITA. 1 male and 2 females, Kuju Mts., Bougatsuru, 27-V-1988, K. Morimoto (ELKU). KAGOSHIMA. Oosumi-hantou Peninsula, Cape Satamisaki: 1 male, 25-VI-1957, T. Saigusa (ELKU); 1 female, 8-VI-1958, H. Ueno (ELKU). 5 males and 6 females, Mt. Hoyoshidake, 14–15-IV-2002, H. Yoshitake (NIAES). 1 male, Mt. Kobadake, 28-VI-2003, H. Yoshitake (NIAES).
Japan (Honshu, Izu Islands, Shikoku, and Kyushu; Fig. 134).
The species name refers to the elongate apical projection of the penis, from Latin: longi-, from longus, -a, -um, meaning “long”, and processus, meaning “projection”. To be treated as a substantive in apposition.
In several localities in Honshu, a number of adults of this species were found on Rubus palmatus Thunb. var. coptophyllus (A.Gray) Kuntze ex Koidz. In addition, adults of this species were collected mostly from Rubus palmatus Thunb. and rarely from Rubus crataegifolius Bunge on Mt. Kusenbuyama, Kyushu.
This species differs from Scleropteroides hypocrita on the basis of the following characters: subapical part of the prothorax strongly constricted (Fig. 38); elytra with strongly prominent humeri, straightly convergent toward the subapical calli (Fig. 39); scales in a row on the elytral intervals erect, much longer than the width of each interval (Fig. 39); male metaventrite more strongly prominent ventrally along the apico-lateral margin of the metaventral receptacle; male ventrite I with no prominence; apical projection of the penis truncate at the apex (Figs 91, 101, 111, 114, 117, 120, and 123); basal part of the endophallus with a pair of weakly falcate sclerites (Figs 90, 100); female sternite VIII with apically divergent arms (Figs 97, 107); bursa copulatrix usually without minute coniform spicules (Figs 95, 105).
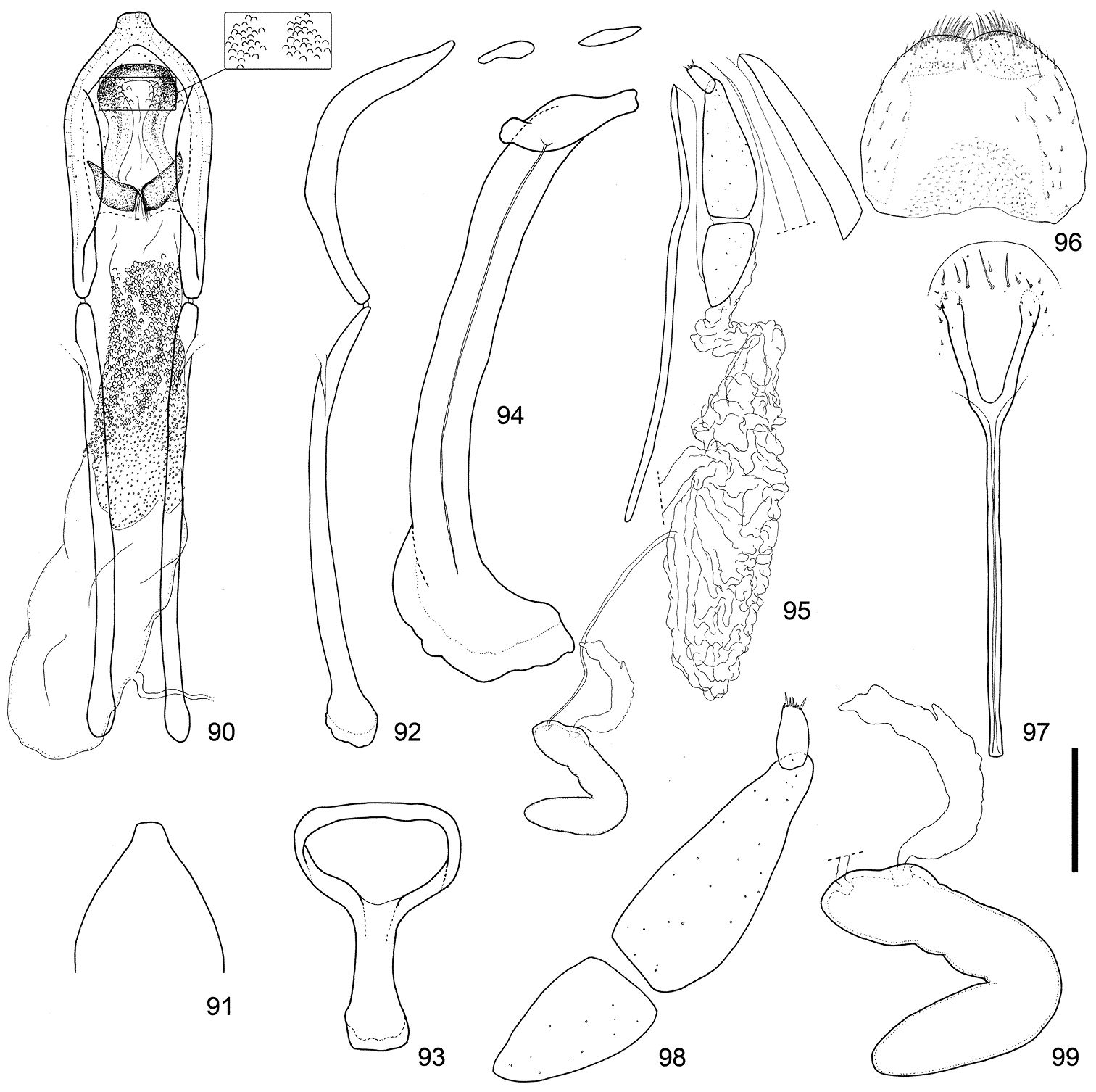
Male and female terminalia of Scleropteroides horridulus (Voss) from Amami-Oshima Island, the Ryukyus, Japan. 90 Aedeagus, dorsal view 91 Apex of the penis, dorsal view 92 Aedeagus, lateral view 93 Tegmen 94 Sternites VIII and IX, male 95 Female terminalia and genitalia, lateral view 96 Tergite VIII, female 97 Sternite VIII, female 98 Coxite and stylus 99 Spermatheca. Scale: 0.20 mm for 90–97, 0.10 mm for 98–99.
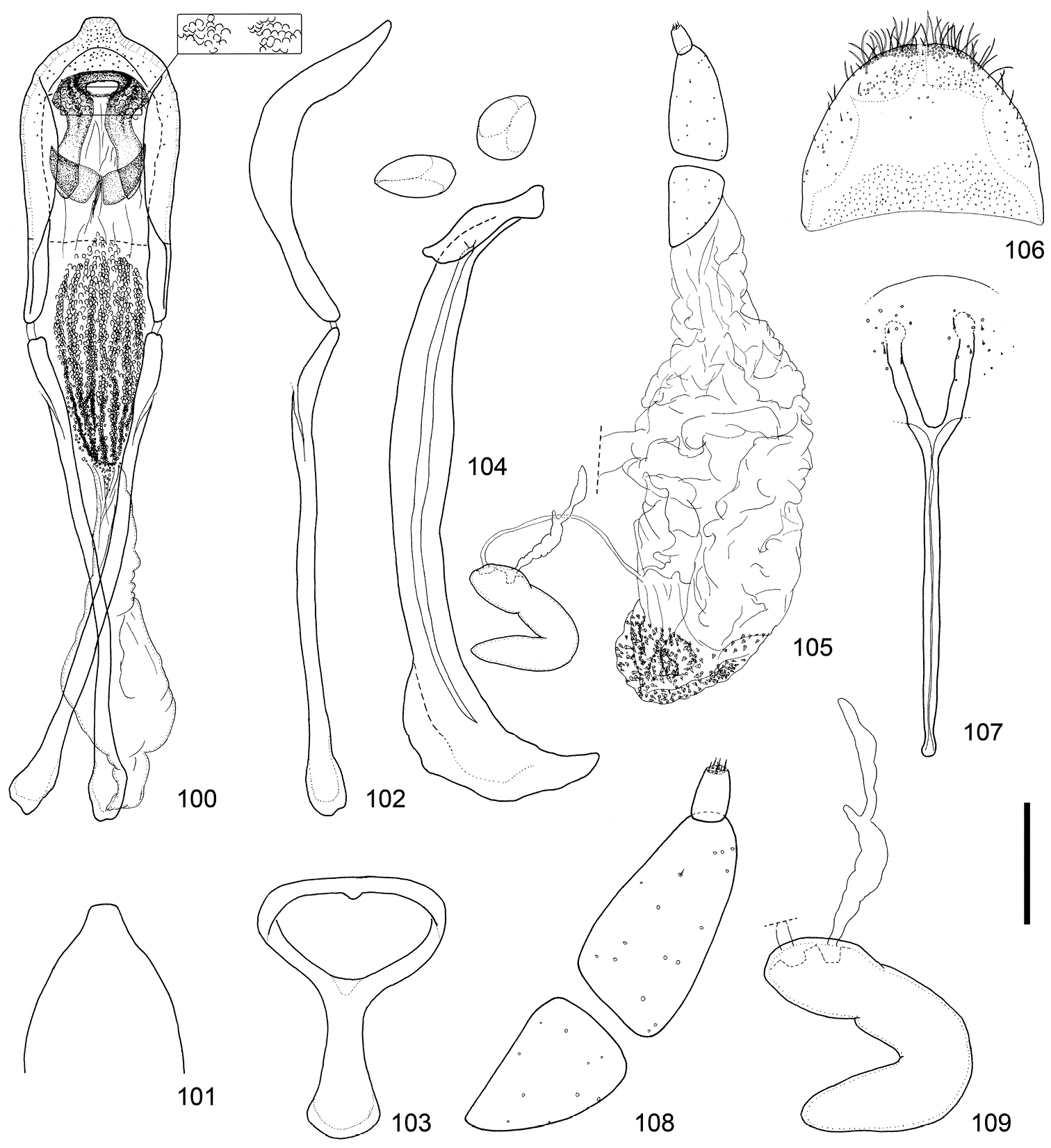
Male and female terminalia of Scleropteroides horridulus (Voss) from Fujian, southeastern China. 100 Aedeagus, dorsal view 101 Apex of the penis, dorsal view 102 Aedeagus, lateral view 103 Tegmen 104 Sternites VIII and IX, male 105 Female genitalia, lateral view 106 Tergite VIII, female 107 Sternite VIII, female 108 Coxite and stylus 109 Spermatheca. Scale: 0.20 mm for 100–107, 0.10 mm for 108–109.
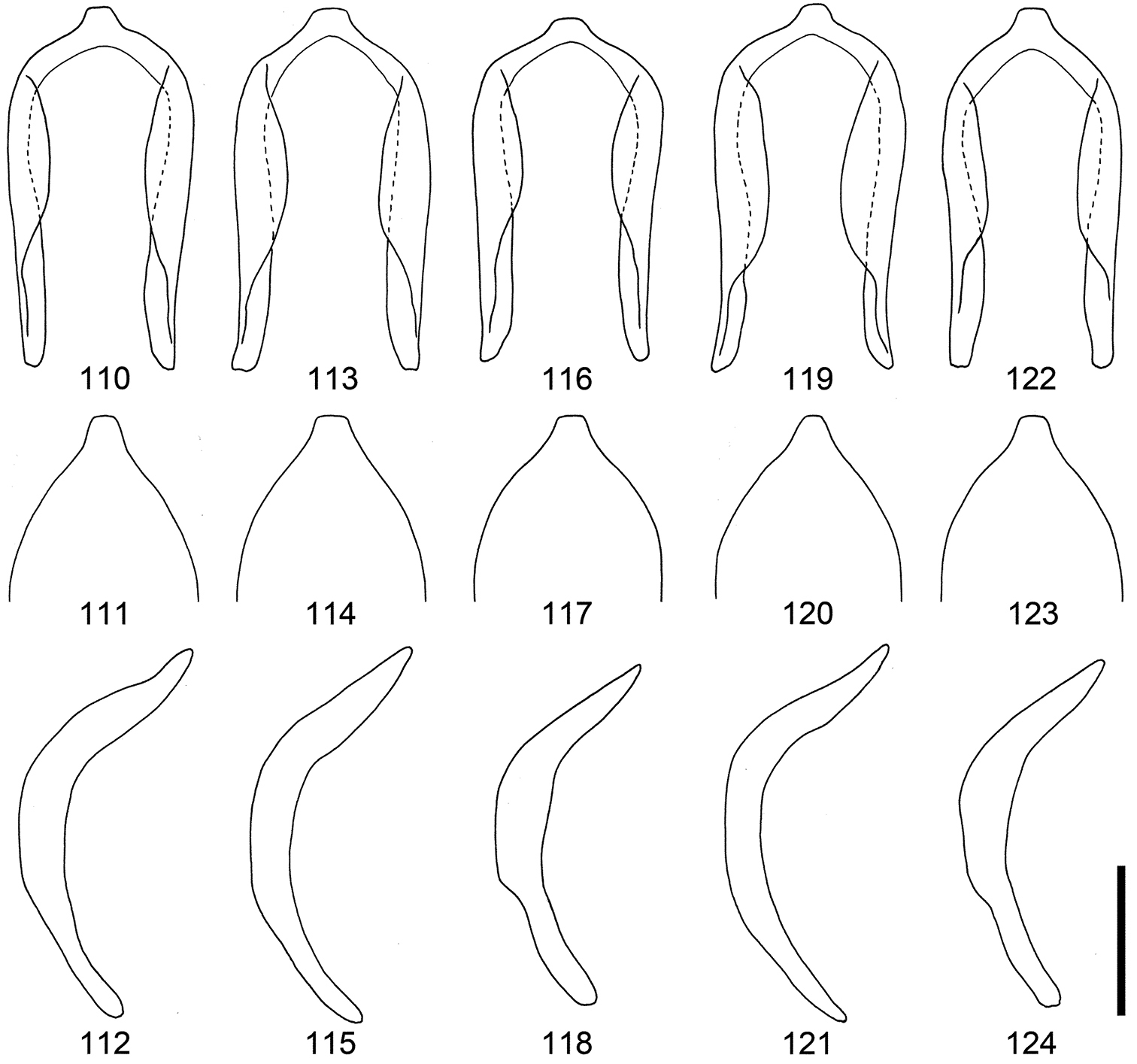
Male genitalia of Scleropteroides horridulus (Voss) from various localities. 110–112 Taiwan, China 113–115 Fujian, southeastern China 116–118 Hunan, central China 119–121 Yakushima Island, Nansei Islands, Japan 122–124 Mt. Jirisan, South Korea 110, 113, 116, 119, 122 Penis, dorsal view 111, 114, 117, 120, 123 Apex of the penis, dorsal view 112, 115, 118, 121, 124 Penis, lateral view. Scale: 0.20 mm.
LB: 2.40–2.71 (mean, 2.50). LR: 1.04–1.18 (mean, 1.12). WP: 0.94–1.10 (mean, 1.00). LP: 0.85–0.95 (mean, 0.90). WE: 1.61–1.91 (mean, 1.69). LE: 1.64–1.83 (mean, 1.69). N = 11 for all measurements. Habitus as shown in Figs 5–6.
Vestiture. Clavate scales long and erect on head, basal 2/3 of rostrum, and pronotum. Hair-like scales long and semierect on apical 1/3 of rostrum. Scales on elytral intervals (Fig. 39) erect, long, 1.3–1.5 × as long as interval width. Clavate scales on tibiae erect.
Rostrum (Figs 30–31) slender, 1.15–1.27 × as long as pronotum, sides slightly widened in apical part, which is 1.16× as wide as basal part. Antennae (Fig. 31) with length ratio of funicular segments I: II: III: IV: V: VI = 1.86: 1.50: 1.21: 1.00: 1.00: 0.93 and width ratio = 1.60: 1.00: 1.13: 1.20: 1.33: 1.47.
Pronotum (Fig. 38) 1.06–1.19 × as wide as long, 0.50–0.56 × as long as and 0.56–0.65 × as wide as elytra; subapical constriction strong; sides subparallel in basal 2/3, then strongly convergent toward apex.
Elytra 0.96–1.04 × as long as wide, 1.79–1.98× as long as and 1.54–1.79× as wide as pronotum, straightly convergent toward subapical calli; humeral calli strongly prominent; subapical calli moderately prominent.
Underside. Metaventrite moderately prominent ventrally along apico-lateral margin of metaventral receptacle. Venter coarsely and very densely punctured; ventrite I weakly flattened on disc, lacking prominence; ventrite V with more or less deeper median concavity; length ratio of ventrites I: II: III: IV: V = 3.56: 2.67: 1.00: 1.00: 2.56 and width ratio = 1.90: 1.76: 1.30: 1.20: 1.00.
Genitalia. Tegmen (Figs 93, 103) with apodeme slightly widened toward apex. Penis (Figs 90–92, 100–102, 110–124) with sides more or less weakly widened toward apical 1/5, then strongly convergent apically; apical projection truncate at apex. Basal part of endophallus (Figs 90, 100) with pair of weakly falcate sclerites.
Otherwise as in Scleropteroides hypocrita.
LB: 2.34–2.92 (mean, 2.68). LR: 1.06–1.36 (mean, 1.30). WP: 0.95–1.14 (mean, 1.09). LP: 0.88–1.04 (mean, 0.96). WE: 1.63–1.97 (mean, 1.82). LE: 1.62–2.03 (mean, 1.83). N = 16 for all measurements.
Rostrum (Figs 32–33) slightly more slender, 1.15–1.40 × as long as pronotum.
Pronotum 1.02–1.18 × as wide as long.
Elytra 0.98–1.05 × as long as wide.
Underside. Ventrite V simple, lacking median sulcus or concavity.
Terminalia and genitalia. Sternite VIII (Figs 97, 107) with arms slender and apically divergent. Bursa copulatrix (Figs 95, 105) usually simple, but sometimes with dense minute coniform spicules in posterior part. Spermatheca (Figs 99, 109, 126, 128, 130, 132) with collum more strongly convex; insertions of duct and gland slightly more distant from each other.
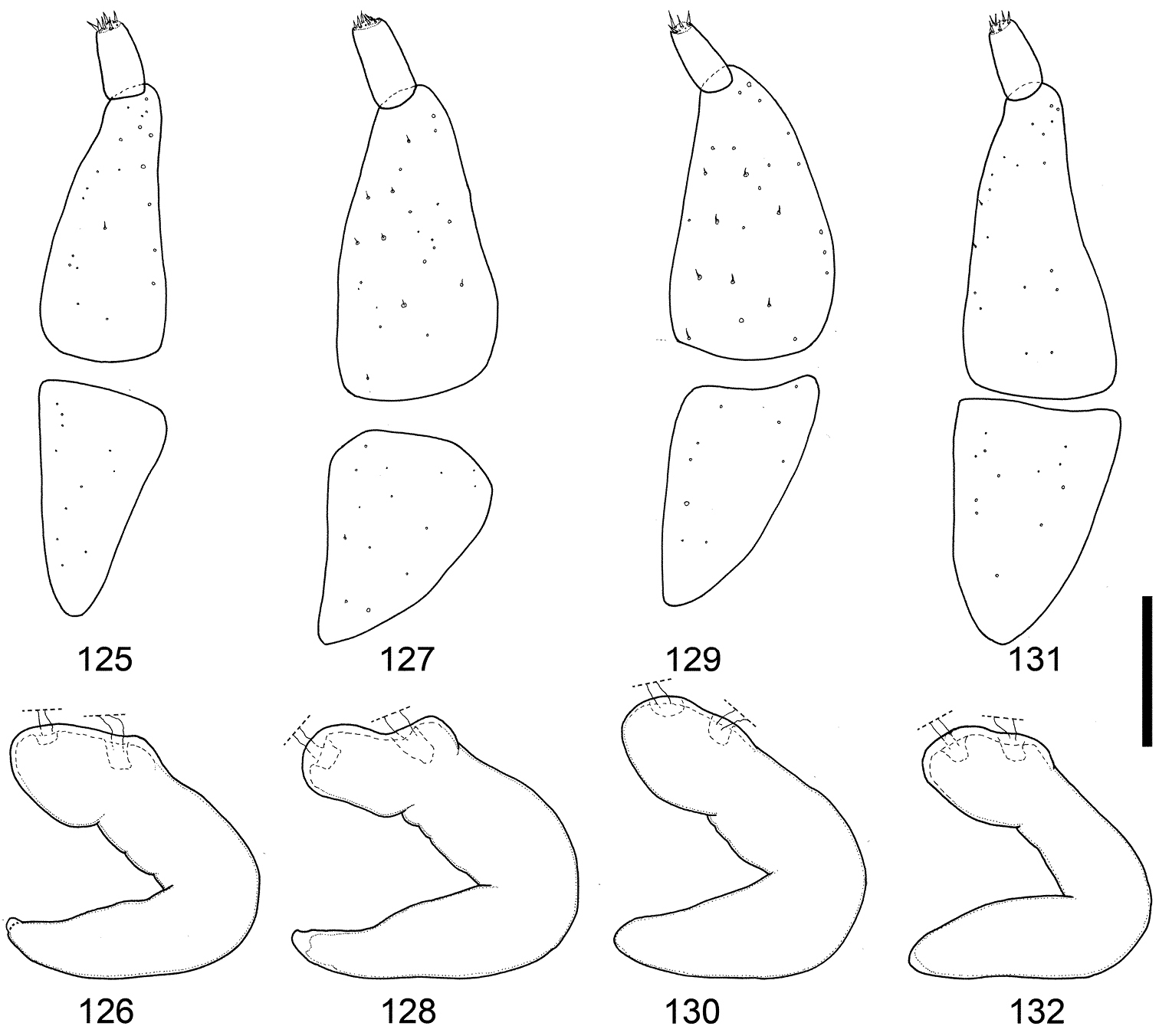
Female genitalia of Scleropteroides horridulus (Voss) from various localities. 125–126 Taiwan, China 127–128 Fujian, southeastern China 129–130 Yakushima Island, Nansei Islands, Japan 131, 132 Namwongun, South Korea. Scale bar: 0.10 mm.
Otherwise showing almost the same sexual dimorphism as in Scleropteroides hypocrita.
CHINA: Hunan. 1 male, Pingjiang, Mt. Mupushan, 1600 m, 1–12-VIII-2003, Li et al. (NIAES). Fujian. 2 males and 2 females, Jianyang, 27°20'N, 118°06'E, 18-IV-1965 (IZCAS, IOZ(E)896779, 896780, 896781, and 896786); 1 female, Jianyang, Aotou, Sanbanqiao, 27°35'N, 117°39'E, 18-IV-1965 (IZCAS IOZ(E)896788); 1 female, Jianyang, Aotou, Huangbaixi, 27°35'N, 117°39'E, 7-V-1965 (IZCAS IOZ(E)89679); 1 male, Jianyang, Aotou, Daoshui, 27°35'N, 117°39'E, 13-VI-1965 (IZCAS, IOZ(E)897690). 1 female, Shaowu, Tieyang, 27°10'N, 117°21'E, 15-V-1965 (IZCAS, IOZ(E)896789). 1 male, Wuyishan, Xingcun, Sangang, 752 m, 27°44.804'N, 117°40.600'E, 15-IV-2009, J. Huang (ZAFU). 1 female, Wuyishan, Xingcun, Sangang, 752 m, 27°44.804'N, 117°40.600'E, 15-IV-2009, S. Zhang (ZAFU). 1 male, Wuyishan, Xingcun, Qili, 899–912 m, 27°43.302'–27°43.042'N, 117°39.632'–117°39.334'E, 20-IV-2009, J. Tan (ZAFU). 1 female, Wuyishan, Xingcun, Qili, 899–912 m, 27°43.302'–27°43.042'N, 117°39.632'–117°39.334'E, 20-IV-2009, J. Huang (ZAFU). Chongqing. 1 female, Nanchuan, Yixiantian, 700 m, 29.05194°N, 107.12001°E, 12-VI-2010, R. Nie (IZCAS IOZ(E)1803625). TAIWAN. 1 male, Nantou, Sungkan–Meifeng, 25–26-V-1972, M. Sakai (EUMJ). 1 male and 1 female, Nantou, Nanshanchi, 20–23-III-1995, H. Kojima and M. Suehiro (NIAES). 1 male, Mt. Guandaoshan, 16-V-1986, I. Matoba (NIAES). 1 male, Taoyuan, Mt. Daguanshan, 4-IV-1991, H. Kojima (NIAES). KOREA: Gangwondo. 1 female, Chuncheongun, Dongmyeon, Gamjeongri, 21-V-1992, K. Morimoto (ELKU). Kyeongsangnamdo. 1 female, Mt. Jirisan, Samjeongri, 6-VI-1991, J. D. Bae (ELKU). Jeollabukdo. 1 male and 1 female, Namwongun, Nowondan, 5-VI-1991, J. D. Bae (ELKU). 1 female, Namwongun, Deongdongri, 19-VI-1994, H. Kojima (ELKU). JAPAN: Nansei Islands. YAKUSHIMA I. Nakama: 1 male, 1-IV-1985, A. Yoshida (NIAES); 1 male and 2 females, 25-III-1991, H. Kojima (ELKU); 4 males and 6 females, 30-III-2005, Y. Kubota (NIAES). AMAMI-OSHIMA I. 1 male, Sutaru-touge Pass, 23-III-1954, T. Mohri (EUMJ). Nishinakama: 1 female, 25-III-1954, T. Edashige (EUMJ); 1 female, 31-III-1968, M. Tomokuni (EUMJ); 1 male, 22-III-1990, T. Ueno (NIAES). Hatsuno: 1 male and 1 female, 24-III-1965, S. Fukuda (SM); 1 female, 12-IV-1971, M. Sakai (EUMJ); 1 female, 11-V-1976, J. Okuma (SM). 1 male, Daikuma, 10-IV-1971, M. Sakai (EUMJ). 1 female, Shinmura, 11-IV-1971, M. Sakai (EUMJ). Mt. Yuwandake: 1 male, 19-IV-1971, M. Sakai (EUMJ); 25 males and 18 females, 25–28-III-2003, H. Yoshitake, on Rubus croceacanthus (NIAES). 9 males and 8 females, Chuo-rindou, 18-III-1991, H. Kojima (ELKU). 2 males, Nangawa-rindou, 20-III-1991, H. Kojima (ELKU); 1 male and 2 females, 22-III-1991, H. Kojima (ELKU). 2 males and 5 females, Mt. Akatsuchiyama, 21-III-1991, H. Kojima (ELKU). UKEJIMA I. 1 male, 22-III-2004, K. Takahashi (NIAES). OKINAWAJIMA I. 1 female, Yona, 10-IV-1975, S. Imasaka (NIAES). 1 male, Oku, 15–18-V-1978, T. Tsutsumi (ELKU). 2 males and 1 female, Mt. Yonahadake, 16-IV-1991, H. Kojima (ELKU).
China (Chongqing and Fujian – new record, and Taiwan), Korea (Central and South – new record), Japan (Nansei Islands: Yakushima, Amami-Oshima, and Okinawajima Islands; Fig. 134).
A number of adults were collected from Rubus croceacanthus H.Lév. growing along the edge of an evergreen broad-leaved forest at the foot of Mt. Yuwandake, Amami-Oshima Island.
In the study, we could not examine the holotype of Rhytidosomus insularis Voss, which should be preserved in ELKU, but K. Morimoto (pers. comm., 2005) had examined it and confirmed its taxonomic identity. Also, our examination of a long series of Scleropteroides specimens from the type locality, Amami-Oshima Island, revealed that only one species of this genus occurs on the island.
Scleropteroides was established as a genus allied to Scleropterus in the tribe Scleropterini (
Previously, Scleropteroides was defined by
Morphologically, Scleropteroides is close to Scleropterus in sharing a rounded apex of the antennal scape, a six-segmented antennal funicle, a pronotum with neither tubercules nor prominences, elytral intervals bearing small and acute squamate granules usually in a row, a rostral channel extending to the metaventrite, and appendiculate claws. In addition, the close affinity between the two genera is emphasized by similarities in structures of the male and female genitalia.
On the basis of detailed morphological observations, we redefined Scleropteroides in this study as follows: 1) eyes convex (Figs 22–33); 2) antennal scape fringed with 3–4 linear scales at the apex, in addition to a few minute hairs; 3) basal margin of the pronotum bisinuate (Figs 10, 34, 36, 38); 4) pronotum lacking a sulcus (Figs 10, 34, 36, 38); 5) apical margin of the pronotum raised (Fig. 12); 6) scutellar shield well-developed, visible from above (Figs 11, 34–39); 7) hind wings well-developed (Fig. 19); 8) elytral humeri well-developed (Figs 11, 35, 37, 39); 9) elytral intervals subequal in width and height (Figs 11, 35, 37, 39); 10) femora dentate (Fig. 15); 11) protibiae simple, not incurved apically (Fig. 15); and 12) meso- and metatibiae mucronate only in males.
In contrast, Scleropterus has the following defining characteristics: 1) eyes flattened; 2) antennal scape fringed with 3–4 minute hairs at the apex; 3) basal margin of the pronotum nearly straight; 4) basal half of the pronotum with a weak longitudinal median sulcus; 5) apical margin of the pronotum slightly descending; 6) scutellar shield vestigial, usually invisible from above; 7) hind wings vestigial; 8) elytral humeri vestigial; 9) odd numbered intervals of the elytra wider and more prominent than even numbered intervals; 10) femora edentate; 11) protibiae incurved apically; and 12) meso- and metatibiae mucronate in both sexes.
The most apparent differences between the two genera are the structure of the hind wings (character 7) and associated characters (3, 6, and 8). Apart from these differences associated with hind-wing reduction in Scleropterus, however, Scleropteroides clearly differs from Scleropterus in having a pronotum lacking a sulcus, even elytral intervals, dentate femora, and simple protibiae. Additionally, Scleropteroides can also be distinguished from Scleropterus by the meso- and metatibiae lacking mucrones in females.
Our extensive and detailed examination of a large number of specimens revealed that Scleropteroides comprises three East Asian species – Scleropteroides hypocrita, Scleropteroides horridulus, and Scleropteroides longiprocessus. They are very similar in general appearance, as well as in general structures of male and female terminalia, but can be distinguished clearly by several consistent, taxonomically important morphological differences as shown in the following lines.
Scleropteroides hypocrita is characterized mainly by a moderately constricted prothorax in the subapical part (Fig. 34), gently convergent elytra toward the subapical calli (Fig. 35), moderately prominent humeri (Fig. 35), and elytral intervals with a row of semirecumbent scales that are evidently shorter than the interval width (Fig. 35). In addition, the species is also distinguished by sexual structures in males, such as a slightly widened rostrum in the apical half (Fig. 22), metaventrite that is weakly prominent along the apico-lateral margin of the metaventral receptacle, ventrite I bearing a median prominence along the apical margin, the penis with a blunt apical projection that is rounded at the apex (Figs 41, 51, 61), and a pair of fig-like sclerites on the basal part of the endophallus (Figs 40, 50, 60). In females, the arms of sternite VIII are apically arcuate (Figs 47, 57, 67) and the posterior part of the bursa copulatrix is densely covered with minute coniform spicules (Figs 45, 55, 65). The length of scales in a row on the elytral intervals is variable, from 0.4 to 0.8 times as long as the interval width, mostly ranging from 0.5 to 0.6 times as long. We examined specimens from various localities in detail, but no significant geographical variation was observed in the external or genital structures of either sex.
Scleropteroides longiprocessus shows character states intermediate between Scleropteroides hypocrita and Scleropteroides horridulus in having a strong constriction in the subapical part of the prothorax (Fig. 36), strongly prominent humeral calli (Fig. 37), and elytral intervals with semierect scales, each of which is nearly as long as the interval width (Fig. 37). This species was previously confused with Scleropteroides hypocrita due to their close resemblance in general appearance and the lack of a detailed morphological examination. However, Scleropteroides longiprocessus differs from Scleropteroides hypocrita by having a stouter rostrum (Fig. 26), more rapidly convergent elytra toward the subapical calli (Fig. 37), and ventrite I lacking a median prominence. Moreover, Scleropteroides longiprocessus is distinguished by male sexual traits, such as a metaventrite that is more prominent ventrally along the apico-lateral margin of the metaventral receptacle, an elongate projection at the apex of the penis (Figs 71, 81), and the paired falcate sclerites in the basal part of the endophallus (Figs 70, 80). With regard to female sexual traits in Scleropteroides longiprocessus, the arms of sternite VIII are apically divergent (Figs 77, 87) and the bursa copulatrix lacks spicules (Figs 75, 85). This species shows non-geographical intraspecific variation in the length of scales on the elytral intervals. Most specimens examined had scales slightly longer than the interval width, but some specimens bore stouter scales that were slightly shorter than the interval width.
Additionally, Scleropteroides longiprocessus resembles Scleropteroides horridulus in having straightly convergent elytra toward the subapical calli (Figs 37, 39), a metaventrite that is moderately prominent ventrally along the apico-lateral margin of the metaventral receptacle in males, ventrite I lacking a median prominence in both sexes, and the apically divergent arms of female sternite VIII (Figs 77, 87, 97, 107). However, Scleropteroides longiprocessus is easily distinguished from Scleropteroides horridulus by the more strongly widened rostrum in males (Fig. 26), the pronotum with subparallel sides in the basal half and a weaker subapical constriction, less-developed elytral humeri, less erect and shorter scales on the elytral intervals (Fig. 37), a penis with a slender apical projection that is rounded at the apex (Figs 71, 81), narrower sclerites in the basal part of the endophallus (Figs 70, 80), and an unarmed bursa copulatrix (Figs 75, 85).
The affinity of the three Scleropteroides species is still uncertain and a phylogenetic analysis of the genus is necessary to clarify their relationships.
Rubus is a cosmopolitan genus and contains an estimated 900–1000 species worldwide (
Presently, ecological information on Scleropteroides weevils is limited and their host plant associations are not clear (Table 2). However, the available data indicate that Scleropteroides weevils are associated with woody Rubus species belonging to the subgenus Idaeobatus and growing in and around woodlands at least in the adult stage because adults were collected from Rubus shrubs and observed feeding on leaves and flowers of the plants on many occasions. With regard to adult feeding habits, Scleropteroides hypocrita widely utilizes at least four Rubus species (Rubus crataegifolius, Rubus idaeus subsp. melanolasius f. concolor, Rubus microphyllus, and Rubus trifidus) in Korea and Japan, whereas Scleropteroides longiprocessus was observed feeding on Rubus palmatus in Honshu and Kyushu, and Scleropteroides horridulus on Rubus croceacanthus in the Ryukyus. In two localities in Toyota City, Honshu, Japan, adults of Scleropteroides hypocrita and Scleropteroides longiprocessus were found separately on Rubus microphyllus and Rubus palmatus var. coptophyllus, respectively (K. Izawa, pers. comm., 2014). Similarly, on Mt. Kusenbuyama, Kyushu, Japan, adults of Scleropteroides hypocrita and Scleropteroides longiprocessus were collected separately on Rubus crataegifolius and Rubus palmatus, respectively, except a few individuals of Scleropteroides longiprocessus collected simultaneously with Scleropteroides hypocrita on Rubus crataegifolius. These observations suggest that Scleropteroides species might show some differences in host use in localities where they occur sympatrically. As mentioned above, Scleropteroides hypocrita is sympatric with Scleropteroides longiprocessus in Japan and with Scleropteroides horridulus in South Korea (Table 1). Further studies are needed to explore the host plant range of each Scleropteroides species, especially in sympatric localities.
Localities where Scleropteroides species occur sympatrically.
| Species | Locality |
|---|---|
| Scleropteroides hypocrita and Scleropteroides horridulus | IN THE KOREAN PENINSULA: |
| GANGWONDO: Chuncheongun, Dongmyeon, Gamjeongri | |
| KYEONGSANGNAMDO: Mt. Jirisan, Samjeongri | |
| JEOLLABUKDO: Namwongun, Deongdongri | |
| Scleropteroides hypocrita and Scleropteroides longiprocessus | IN THE JAPANESE ARCHIPELAGO: |
| IWATE: Kawai, Tsuchisaka-touge | |
| AKITA: Lake Tazawako | |
| YAMAGATA: Mt. Zaousan, Yoshikari-rindou | |
| FUKUSHIMA: Haranomachi, Kozikiishi-rindou; Namie | |
| IBARAKI: Yamizosan, Shimokitazawa | |
| TOCHIGI: Fujiwara, Midorizawa-rindou | |
| GUMMA: Matsuida, Kirizumi-onsen | |
| SAITAMA: Shoumaru-touge Pass | |
| TOKYO: Okutama, Kurasawadani; Mt. Oodakeyama; Mt. Takaosan | |
| KANAGAWA: Tanzawa Mts., Yabitsu-touge Pass; Tanzawa Mts., Mizunashigawa; Yamakita, Mikuni-touge Pass | |
| CHIBA: Mt. Kiyosumiyama–Kenminnomori | |
| NAGANO: Nakakawa; Tobira-onsen | |
| SHIZUOKA: Shinfuji | |
| AICHI: Toyota, Nishiichinonochou; Toyota, Asugawachou, Asugawa-hoan-rindou | |
| MIE: Misugi, Hirakura | |
| SHIGA: Mt. Ibukiyama | |
| FUKUOKA: Mt. Hikosan | |
| NAGASAKI: Tashirobaru; Sasebo, Mt. Hattendake |
Summary of plant association data for Scleropteroides.
| Species | Host plant | Observation* | Locality | Author |
|---|---|---|---|---|
| Scleropteroides hypocrita | Rubus spp. | A | Japan | |
| Scleropteroides hypocrita | Rubus crataegifolius | A | Japan, Honshu: Mt. Takaosan | This study |
| Scleropteroides hypocrita | Rubus crataegifolius | A | Japan, Honshu: Iwate | This study |
| Scleropteroides hypocrita | Rubus crataegifolius | A | Japan, Kyushu: Mt. Kusenbuyama | This study |
| Scleropteroides hypocrita | Rubus sp. | A | Japan, Honshu: Fukushima | This study |
| Scleropteroides hypocrita | Rubus trifidus | A | Japan, Honshu: Gifu, Toki | This study |
| Scleropteroides hypocrita | Rubus trifidus | A | Japan, Honshu: Aichi, Toyota, Oobora, Nikake-rindou | This study |
| Scleropteroides hypocrita | Rubus microphyllus | A | Japan, Honshu: Aichi, Asugawachou, Asugawa-hoan-rindou | This study |
| Scleropteroides hypocrita | Rubus microphyllus | A | Japan, Honshu: Mie, Komonochou, Komono | This study |
| Scleropteroides hypocrita | Rubus idaeus subsp. melanolasius f. concolor | A | Korea | |
| Scleropteroides hypocrita | Rubus crataegifolius | A | Korea, South: Mt. Jirisan | This study |
| Scleropteroides longiprocessus | Rubus palmatus | A | Japan, Kyushu: Mt. Kusenbuyama | This study |
| Scleropteroides longiprocessus | Rubus palmatus var. coptophyllus | A | Japan, Honshu: Tokyo, Hachiouji, Mt. Takaosan | This study |
| Scleropteroides longiprocessus | Rubus palmatus var. coptophyllus | A | Japan, Honshu: Gifu, Nakatsugawa, Nenoue-kougen | This study |
| Scleropteroides longiprocessus | Rubus palmatus var. coptophyllus | A | Japan, Honshu, Aichi: Shitara, Mt. Nishinagura-Iyama | This study |
| Scleropteroides longiprocessus | Rubus palmatus var. coptophyllus | A | Japan, Honshu, Aichi, Toyota: Nishiichinonochou; Asugawachou, Asugawa-hoan-rindou; Inabuchou, Noirigawa | This study |
| Scleropteroides horridulus | Rubus croceacanthus | A | Japan, Ryukyus: Amami-Oshima I. | This study |
* A: Adults, feeding behavior was confirmed.
Generally, each species in the subfamily Ceutorhynchinae utilizes the same host plant in the adult and larval stages (e.g.,
The distribution of Scleropteroides is limited to East Asia (Figs 133–134), in contrast to Scleropterus, which is widely distributed from Central Europe through Central Asia to East Asia (
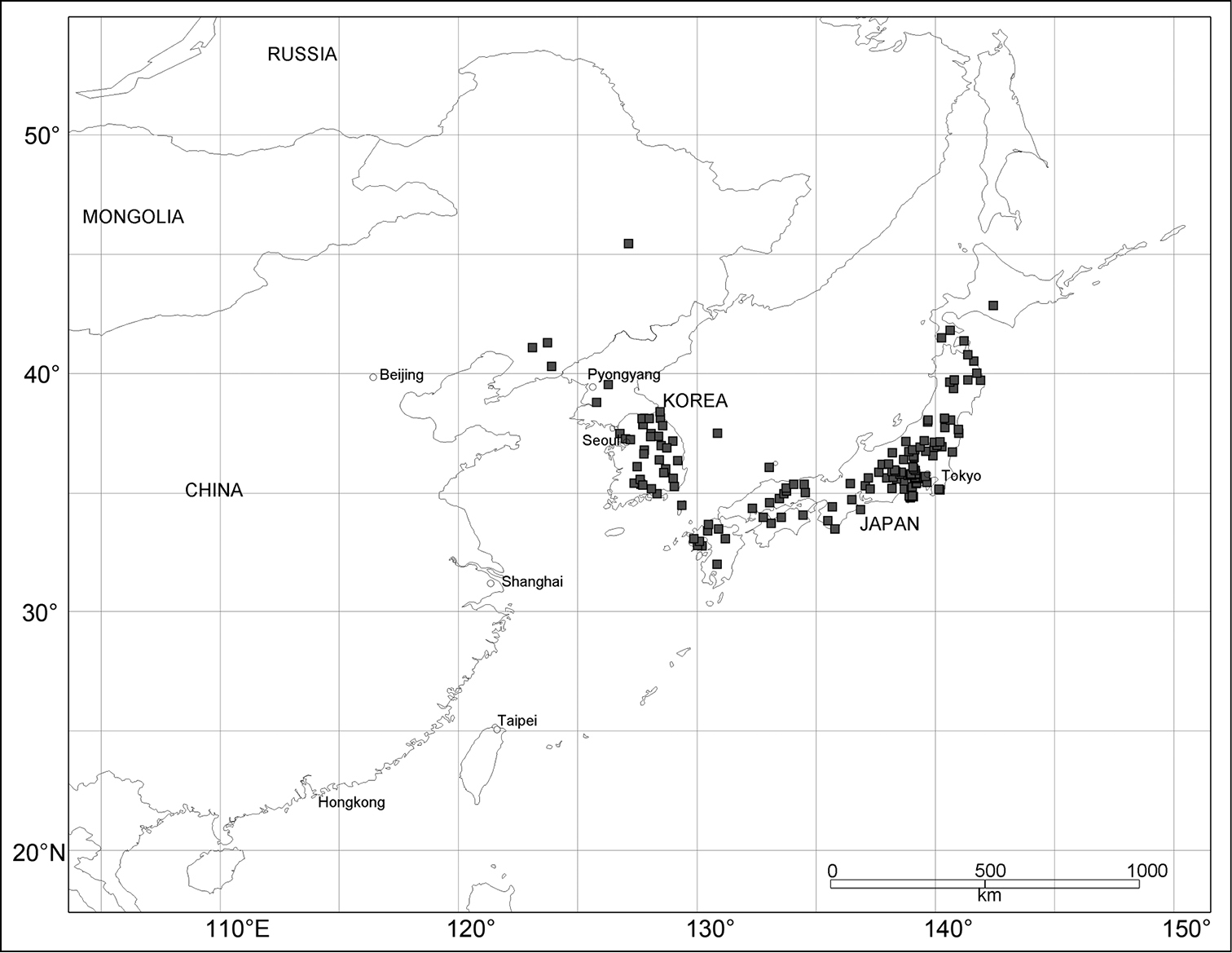
Geographic distribution of Scleropteroides hypocrita (Hustache) (■).
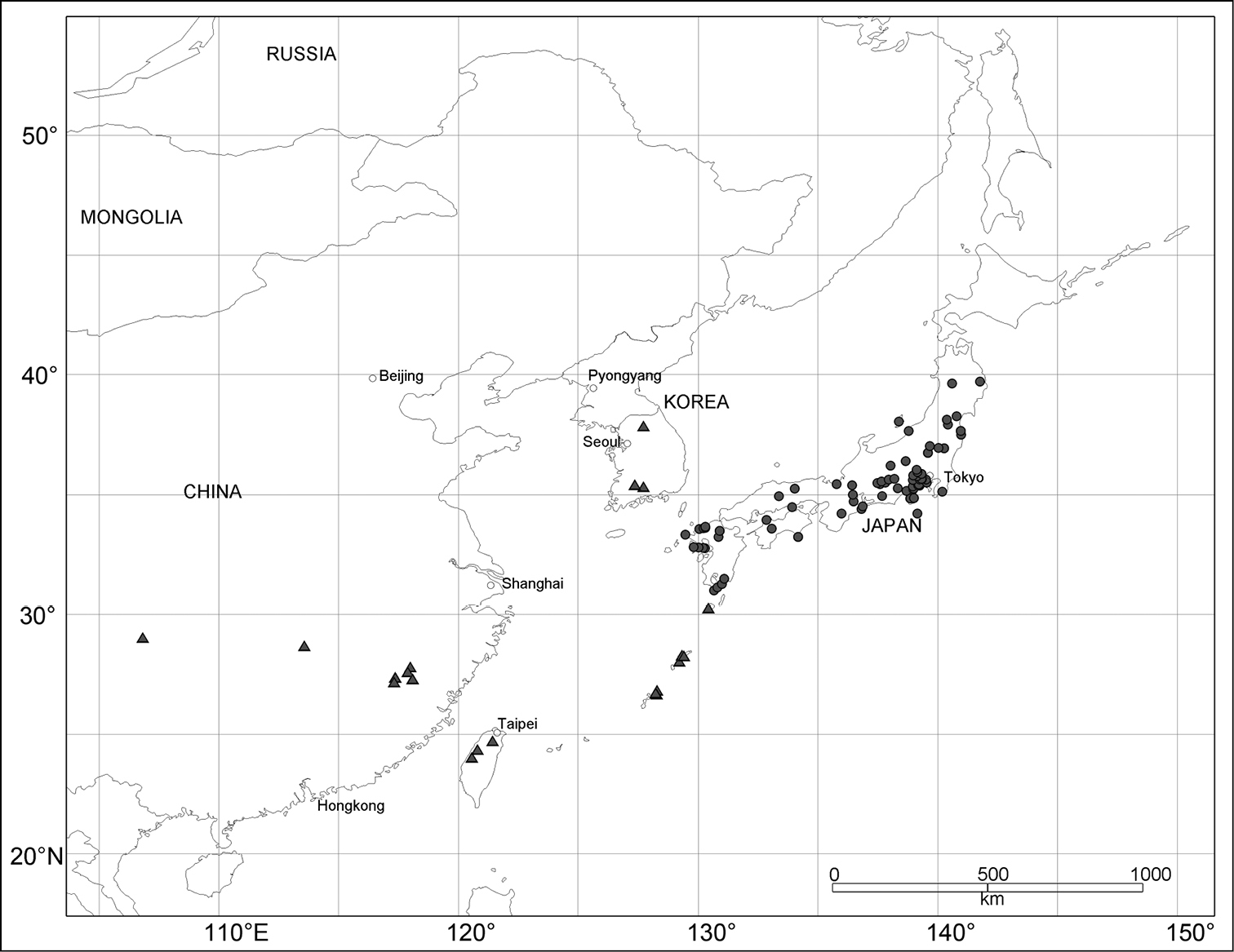
Geographic distributions of Scleropteroides longiprocessus Huang & Yoshitake, sp. n. (●) and Scleropteroides horridulus (Voss) (▲).

Scleropteroides hypocrita (Hustache). 135 Habitat in Toyosawa-rindou, Iwate, Honshu, Japan 136 Adult food plant, Rubus crataegifolius 137 Feeding traces of adults on a flower bud of Rubus crataegifolius (indicated with arrows) 138 Adult on a flower of Rubus crataegifolius.
Scleropteroides hypocrita is distributed in northeast China, North and South Korea, and Japan (Fig. 133), whereas Scleropteroides longiprocessus is restricted to Japan (Fig. 134). The two species occur sympatrically in many localities in Japan (Table 1). Further, Scleropteroides horridulus is distributed from Fujian through Taiwan to the Nansei Islands and in South Korea (Figs 133–134). This species is sympatric with Scleropteroides hypocrita at least in the Korean Peninsula (Table 1), but the distributional boundary of the two species is still unclear in continental China.
Currently, a considerable gap still remains in the distribution of Scleropteroides, especially in continental China and the Korean Peninsula. In addition, this genus has never been recorded from the southern part of the Russian Far East. Since the distribution of Scleropteroides strongly suggests its occurrence in these regions, further surveys are necessary to elucidate the range of the genus and that of each species.
| 1 | Subapical part of the prothorax moderately constricted (Fig. 34). Elytra gently convergent toward the subapical calli, with weakly prominent humeri. Scales on the elytral intervals semirecumbent, evidently shorter than the interval width (Fig. 35). Metaventrite weakly prominent ventrally along the apico-lateral margin of the metaventral receptacle in males. Ventrite I with a median prominence along the apical margin in males. Female sternite VIII with apically arcuate arms (Figs 47, 57, 67). [Rostrum slightly widened in the apical half in males. Male genitalia with a blunt projection at the apex of the penis (Figs 41, 51, 61) and a pair of fig-like sclerites in the basal part of the endophallus (Figs 40, 50, 60). Female genitalia with the bursa copulatrix bearing dense, minute, coniform spicules in the posterior part (Figs 45, 55, 65).] | Scleropteroides hypocrita |
| – | Subapical part of the prothorax strongly constricted (Figs 36, 38). Elytra straightly convergent toward the subapical calli, with strongly prominent humeri. Scales on elytral intervals more or less erect, evidently longer than or nearly as long as the interval width (Figs 37, 39). Metaventrite more strongly prominent ventrally along the apico-lateral margin of the metaventral receptacle in males. Ventrite I simple, lacking a median prominence in both sexes. Female sternite VIII with arms divergent apically (Figs 77, 87, 97, 107) | 2 |
| 2 | Apical half of the rostrum slightly widened in males (Fig. 30). Pronotum subparallel-sided in the basal 2/3, with a strong subapical constriction. Elytral humeri strongly prominent. Scales on elytral intervals erect, much longer than the width of each interval (Fig. 39). Apical projection of the penis blunt, truncate at the apex (Figs 91, 101, 111, 114, 117, 120, 123). Basal part of the endophallus with a pair of weakly falcate sclerites. Bursa copulatrix simple (Fig. 95) or with minute coniform spicules in the posterior part (Fig. 105) | Scleropteroides horridulus |
| – | Rostrum more strongly widened in the apical half in males (Fig. 26). Basal half of the pronotum subparallel-sided, with a weaker subapical constriction. Elytral humeri slightly less prominent. Scales on elytral intervals semierect, nearly as long as the interval width (Fig. 37). Apical projection of the penis sharp, rounded at the apex (Figs 71, 81). Basal part of the endophallus with a pair of narrow falcate sclerites (Figs 70, 80). Bursa copulatrix simple, lacking spicules (Figs 75, 85) | Scleropteroides longiprocessus |
We thank M. A. Alonso-Zarazaga, K.-J. Hong, U. Jinbo, K. Izawa, K. Morimoto, L. Ren, Z. Wang, H. Perrin, and O. Tadauchi for their help with this study. We also thank S. Arai, the late K. Emoto, C. Han, H. Hirano, S. Imasaka, T. Ishikawa, J. Kantoh, T. Kishimoto, H. Kojima, H. Li, K. Matsumoto, the late S. Miyakawa, Y. Notsu, S. Saito, M. Sakai, H. Schillhammer, H. Takizawa, and A. Yoshida for the loan or donation of specimens. Finally, we thank two reviewers for their helpful comments on this manuscript and R. S. Anderson for his kind help in English correction. This research was supported by NSFC programs 31172130/J0930004, ZJNSF program Y3090145, the China Scholarship Council (to J. Huang); National BioResource Project of MEXT Japan.