





Citation: Ermilov SG, Alvarado-Rodríguez O, Retana-Salazar OP (2014) Two new species of Pergalumna (Acari, Oribatida, Galumnidae) from Costa Rica, including a key to all species of the genus from the Neotropical region. ZooKeys 435: 7–23. doi: 10.3897/zookeys.435.8213
Two new species of oribatid mites of the genus Pergalumna (Oribatida, Galumnidae), P. elongatiporosa sp. n. and P. striatiprodorsum sp. n., are described from leaf litter of a secondary forest in Costa Rica. Pergalumna elongatiporosa sp. n. is most similar morphologically to P. horvathorum P. Balogh, 1997 and P. sura P. Balogh, 1997, however, it differs from both by the body size, body surface ornamentation and morphology of notogastral porose areas A1 and A3. Pergalumna striatiprodorsum sp. n. is most similar morphologically to P. hawaiiensis hawaiiensis (Jacot, 1934) and P. strigulata Mahunka, 1978, however, it differs from P. hawaiiensis by the length of interlamellar setae and surface ornamentation of the prodorsum; from P. strigulata by the surface of ornamentation of the notogaster, length of interlamellar setae and morphology of bothridial setae. An identification key to known species of Pergalumna from the Neotropical region is given.
Oribatid mites, new species, Pergalumna, key, Costa Rica, Neotropical region
Pergalumna is a large genus that was proposed by
In the course of taxonomic identification of Costa Rican oribatid mites collected in 2013, we found two new species of the genus Pergalumna. The main goal of our paper is to describe these species. Earlier, only three species were known from Costa Rica (
Three specimens (holotype: male; two paratypes: one male and one female) of Pergalumna elongatiporosa sp. n. and nine specimens (holotype: female; eight paratypes: five males and three females) of Pergalumna striatiprodorsum sp. n. are from: Costa Rica, 9°50'24"N, 83°53'17"W, Cartago, Dulce Nombre, Paraíso, Jardín Botánico Lankester, 1400 m a.s.l., in leaf litter in secondary forest, 14.V.2013, collected by O. Alvarado-Rodríguez and A.P. Retana-Salazar.
Holotypes and paratypes were mounted in lactic acid on temporary cavity slides for measurement and illustration. The body length was measured in lateral view, from the tip of the rostrum to the posterior edge of the ventral fig. The notogastral width refers to the maximum width in dorsal aspect. Lengths of body setae were measured in lateral aspect. All body measurements are presented in micrometers. Formulae for leg setation are given in parentheses according to the sequence trochanter–femur–genu–tibia–tarsus (famulus included). Formulae for leg solenidia are given in square brackets according to the sequence genu–tibia–tarsus. General terminology used in this paper follows that of Grandjean (summarized by
Body size: 332–352 × 246–266. Body surface and pteromorphs microgranulate. Rostral, lamellar and interlamellar setae well developed, barbed. Bothridial setae setiform, ciliate unilaterally. Anterior notogastral margin not developed. Three pairs of porose areas: Aa and A3 elongate triangular, A1 long, band-shaped, specifically curving, Aa located between notogastral alveoli la and lm. Median pore and postanal porose area absent. Aggenital and ano-adanal setae simple, short. Solenidion φ on tibia IV inserted in proximal part.
Measurements. Body length: 332 (holotype, male), 340–352 (two paratypes: one male and one female); notogaster width: 246 (holotype), 246–266 (two paratypes).
Integument. Body color brown to black-brown. Body surface and pteromorphs with dense microgranules (their diameter up to 2). Pteromorphs with poorly visible wrinkles.
Prodorsum. Rostrum broadly rounded. Rostral (ro, 24–28), lamellar (le, 49–53) and interlamellar (in, 69–77) setae setiform, barbed. Bothridial setae (ss, 90–102) setiform, densely ciliate unilaterally. Exobothridial setae absent. Lamellar and sublamellar lines distinct, parallel, curving backwards. Insertions of lamellar setae distanced from the lamellar lines. Porose areas Ad small, elongate oval (8–12 × 2–4), located latero-posteriorly to interlamellar setae.
Notogaster. Anterior notogastral margin not developed. Dorsophragmata (D) long. Notogastral setae represented by 10 pairs of alveoli. Three pairs of porose areas well visible, with distinct margins: Aa weakly triangular, transversally oriented (36–57 × 8–12), A1 long, band-shaped, specifically curving (57–69 × 8–13), A3 elongate, narrowly triangular (28–32 × 8–16). Porose areas Aa located between notogastral alveoli la and lm. Median pore absent. All lyrifissures distinct; im and opisthonotal gland openings (gla) located latero-anteriorly to A1.
Gnathosoma. Morphology of subcapitulum, palps and chelicerae typical for Pergalumna (see
Epimeral and lateral podosomal regions. Apodemes 1, 2, sejugal and 3 well visible. Six pairs of setiform epimeral setae observed; setal formula: 1–0–2–3. Setae 4a, 4b (4) thin, smooth, shorter than 1b, 3b, 3c, 4c (8–10), slightly barbed. Pedotecta II (Pd II) scale-like, rounded in ventral view. Discidia (dis) pointly triangular. Circumpedal carinae (cp) distinct, directed posterior of setae 3b.
Anogenital region. Six pairs of genital (g1, g2, 8; g3–g6, 4), one pair of aggenital (ag, 4), two pairs of anal (an1, an2, 4) and three pairs of adanal (ad1–ad3, 4) setae minute, thin, smooth. Anterior parts of genital figs with two setae. Adanal setae ad3 inserted laterally or antero-laterally to lyrifissures iad. Postanal porose area absent.
Legs. Morphology of leg segments, setae and solenidia typical for Pergalumna (see
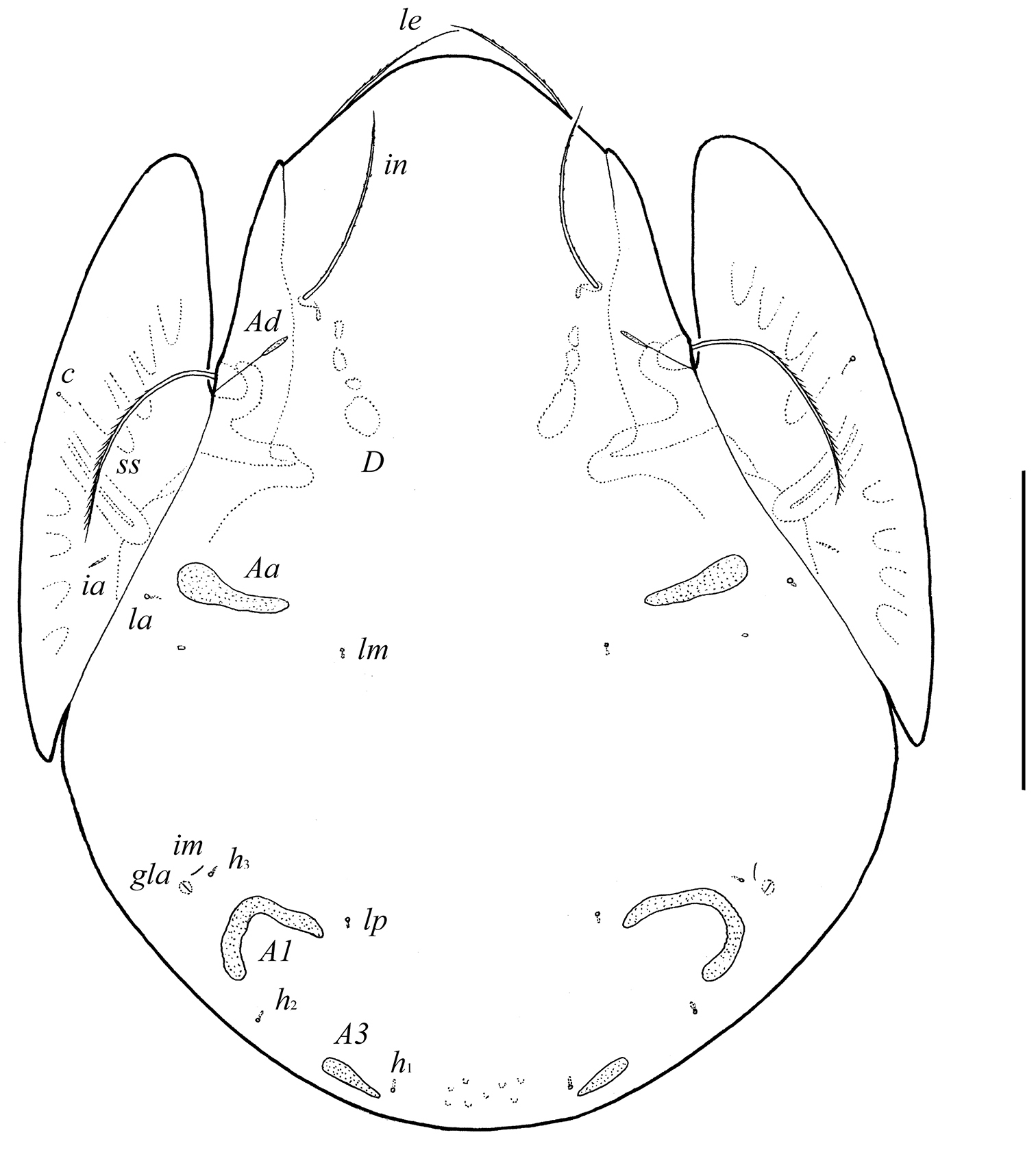
Pergalumna elongatiporosa sp. n.: dorsal view. Scale bar 100 μm.
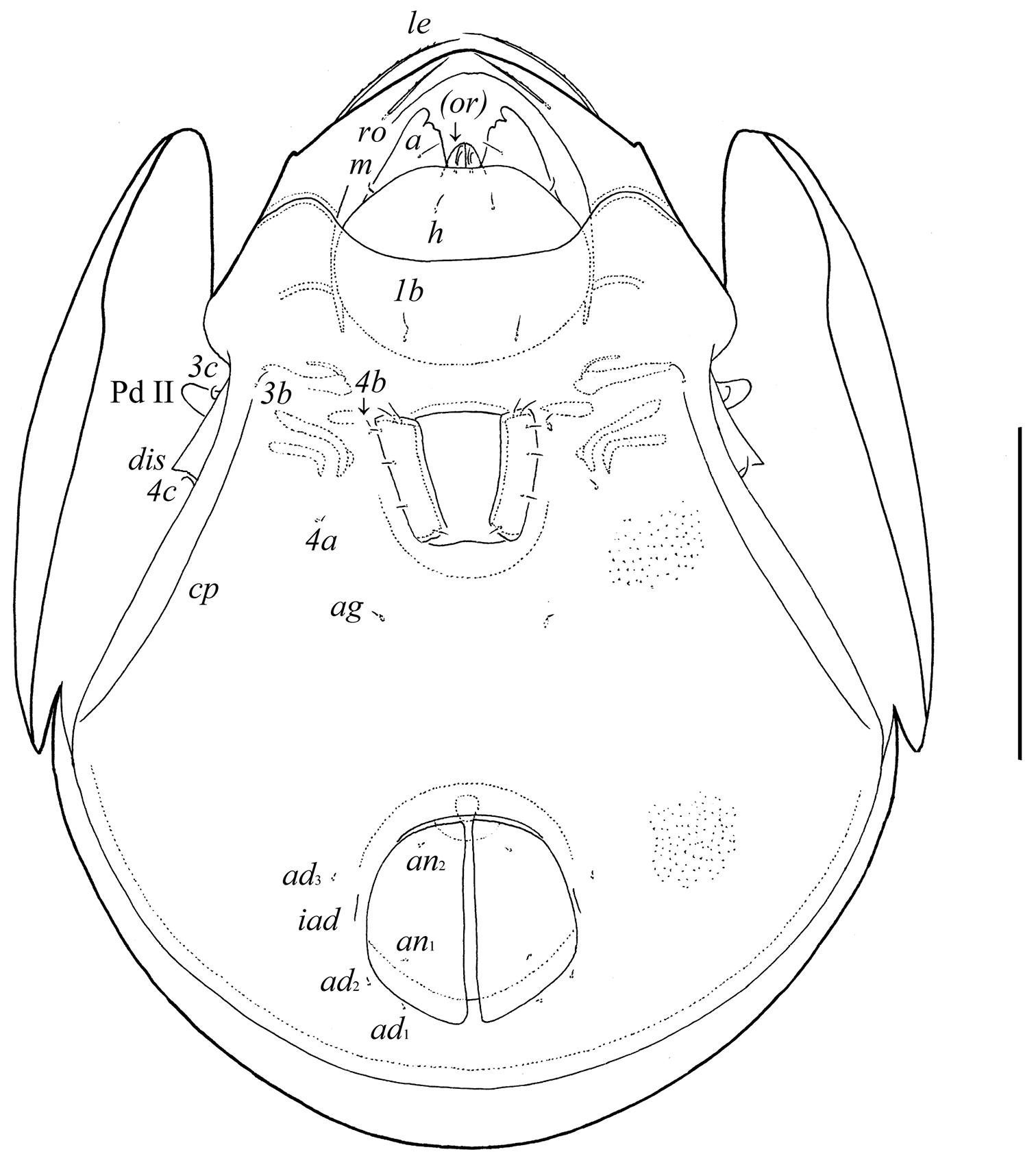
Pergalumna elongatiporosa sp. n.: ventral view (legs not illustrated). Scale bar 100 μm.
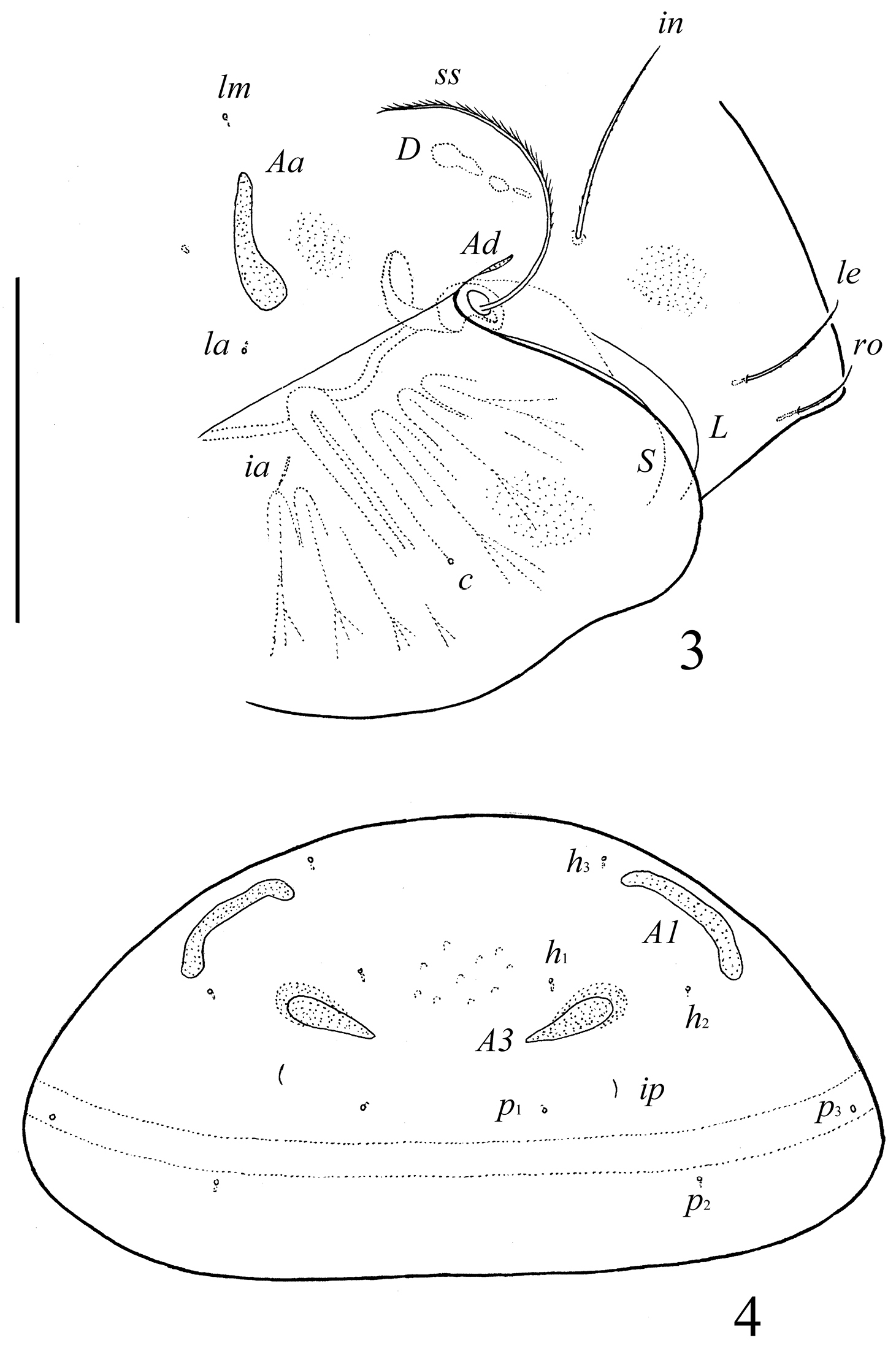
Pergalumna elongatiporosa sp. n.: 3 dorso-lateral view of prodorsum and anterior part of notogaster and pteromorph (gnathosoma and legs not illustrated) 4 posterior view of notogaster. Scale bars 100 μm.
Leg setation and solenidia of Pergalumna elongatiporosa sp. n. (same data for Pergalumna striatiprodorsum sp. n.).
| Leg | Trochanter | Femur | Genu | Tibia | Tarsus |
|---|---|---|---|---|---|
| I | v’ | d, (l), bv’’ | (l), v’, σ | (l), (v), φ1, φ2 | (ft), (tc), (it), (p), (u), (a), s, (pv), v’, (pl), l’’, ε, ω1, ω2 |
| II | v’ | d, (l), bv’’ | (l), v’, σ | (l), (v), φ | (ft), (tc), (it), (p), (u), (a), s, (pv), ω1, ω2 |
| III | v’ | d, ev’ | l’, σ | l’, (v), φ | (ft), (tc), (it), (p), (u), (a), s, (pv) |
| IV | v’ | d, ev’ | d, l’ | l’, (v), φ | ft’’, (tc), (p), (u), (a), s, (pv) |
Roman letters refer to normal setae (ε to famulus), Greek letters to solenidia. Single prime (’) marks setae on anterior and double prime (’’) setae on posterior side of the given leg segment. Parentheses refer to a pair of setae.
The holotype is deposited in the Zoological Institute of the Russian Academy of Sciences, St. Petersburg, Russia; one paratype is deposited in the collection of the Tyumen State University Museum of Zoology, Tyumen, Russia; one paratype is deposited in the collection of the Centro de Investigación en Estructuras, Microscópicas (CIEMIC), Ciudad de la Investigación, Universidad de Costa Rica, San José, Costa Rica.
This specific name “elongatiporosa” refers to the elongate porose areas A1.
In having the setiform bothridial setae, well developed interlamellar setae, absence of anterior notogastral margin and presence of three pairs of porose areas (Aa transversally elongated), Pergalumna elongatiporosa sp. n. is most similar to Pergalumna horvathorum P. Balogh, 1997 and Pergalumna sura P. Balogh, 1997 (see also
Body size: 630–697 × 448–514. Body surface and pteromorphs microgranulate; surface of prodorsum with numerous longitudinal stria. Rostral, lamellar and bothridial setae of medium size, setiform, barbed; interlamellar setae short, slightly thickened, barbed. Anterior notogastral margin not developed. Three pairs of porose areas oval; Aa located between notogastral alveoli la and lm, close to lm. Median pore absent. Aggenital and ano-adanal setae simple, short. Postanal porose area present, elongated.
Measurements. Body length: 697 (holotype, female), 630–697 (eight paratypes: five males and three females); notogaster width: 514 (holotype), 448–514 (eight paratypes).
Integument. Body color brown to black-brown. Body surface, pteromorphs and subcapitular mentum with dense microgranules (their diameter up to 2). Surface of prodorsum with numerous longitudinal stria. Pteromorphs with poorly visible wrinkles.
Prodorsum. Rostrum broadly rounded. Rostral (41–53), lamellar (57–69) and bothridial (106–114) setae setiform, barbed. Interlamellar setae short (12–16), setiform, slightly thickened, barbed. Exobothridial setae absent. Lamellar and sublamellar lines distinct, parallel, curving backwards. Insertions of lamellar setae distanced from the lamellar lines. Porose areas Ad elongate oval (20–28 × 4–8), located latero-posteriorly to interlamellar setae.
Notogaster. Anterior notogastral margin not developed. Dorsophragmata of medium size. Notogastral setae represented by 10 pairs of alveoli. Three pairs of porose areas well visible, with distinct margins: Aa rounded (14–16) or oval, weakly transversally oriented (14–20 × 12–16), A1 oval, weakly elongated diagonally (24–41 × 12–20), A3 rounded (12–16 )or oval (12–16 × 10–12). Porose areas Aa located between notogastral alveoli la and lm, but clearly closer to lm. Median pore absent. All lyrifissures distinct; im located latero-anteriorly to A1. Opisthonotal gland openings located laterally to A1.
Gnathosoma. Morphology of subcapitulum, palps and chelicerae typical for Pergalumna (see
Epimeral and lateral podosomal regions. Apodemes 1, 2, sejugal and 3 well visible. Six pairs of setiform epimeral setae observed; setal formula: 1–0–2–3. Setae 4a, 4b (6–8) thin, smooth, shorter than 3b (20–24) and 1b, 3c, 4c (32–36), slightly barbed. Pedotecta II scale-like, rounded in ventral view. Discidia pointly triangular. Circumpedal carinae distinct, directed posterior of setae 3b.
Anogenital region. Six pairs of genital (g1, 16–18; g2, 12–14; g3–g6, 8–10), one pair of aggenital (6–8), two pairs of anal (6–8) and three pairs of adanal (6–8) setae setiform, thin, smooth. Anterior parts of genital figs with two setae. Adanal setae ad3 inserted laterally to lyrifissures iad. Postanal porose area present, elongate oval (20–28 × 6–8).
Legs. Morphology of leg segments, setae and solenidia typical for Pergalumna (see
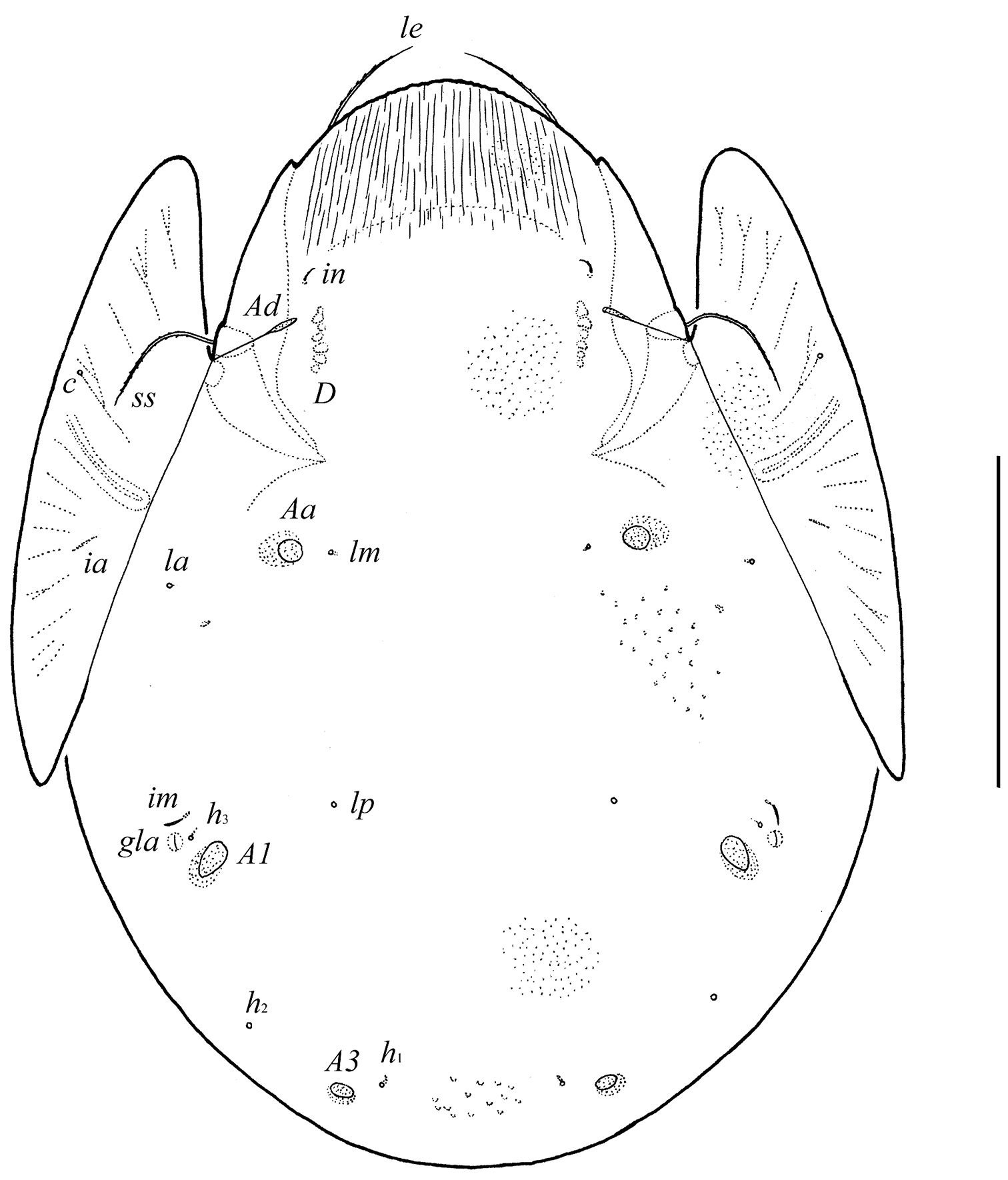
Pergalumna striatiprodorsum sp. n.: dorsal view. Scale bar 200 μm.
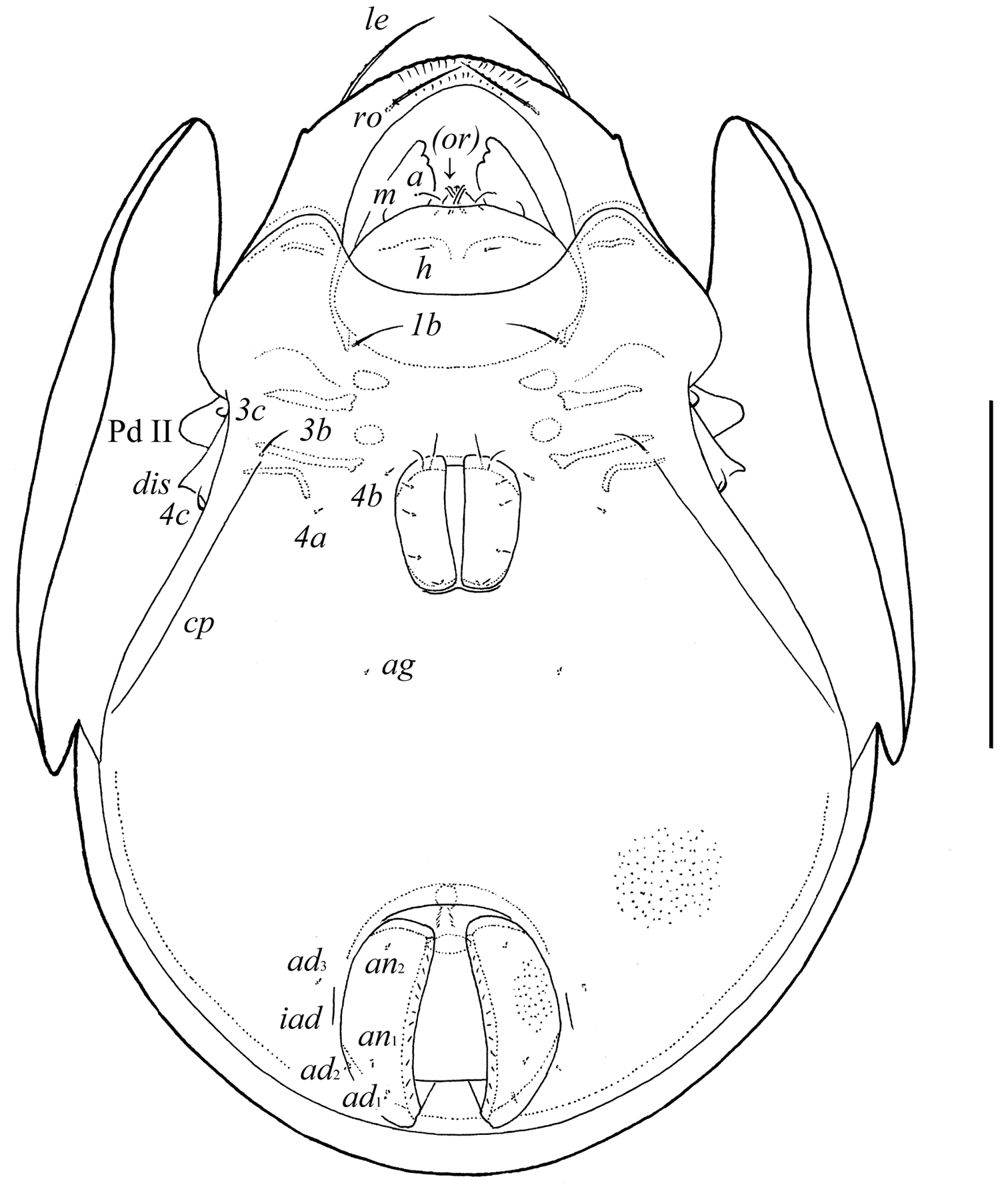
Pergalumna striatiprodorsum sp. n.: ventral view (legs not illustrated). Scale bar 200 μm.
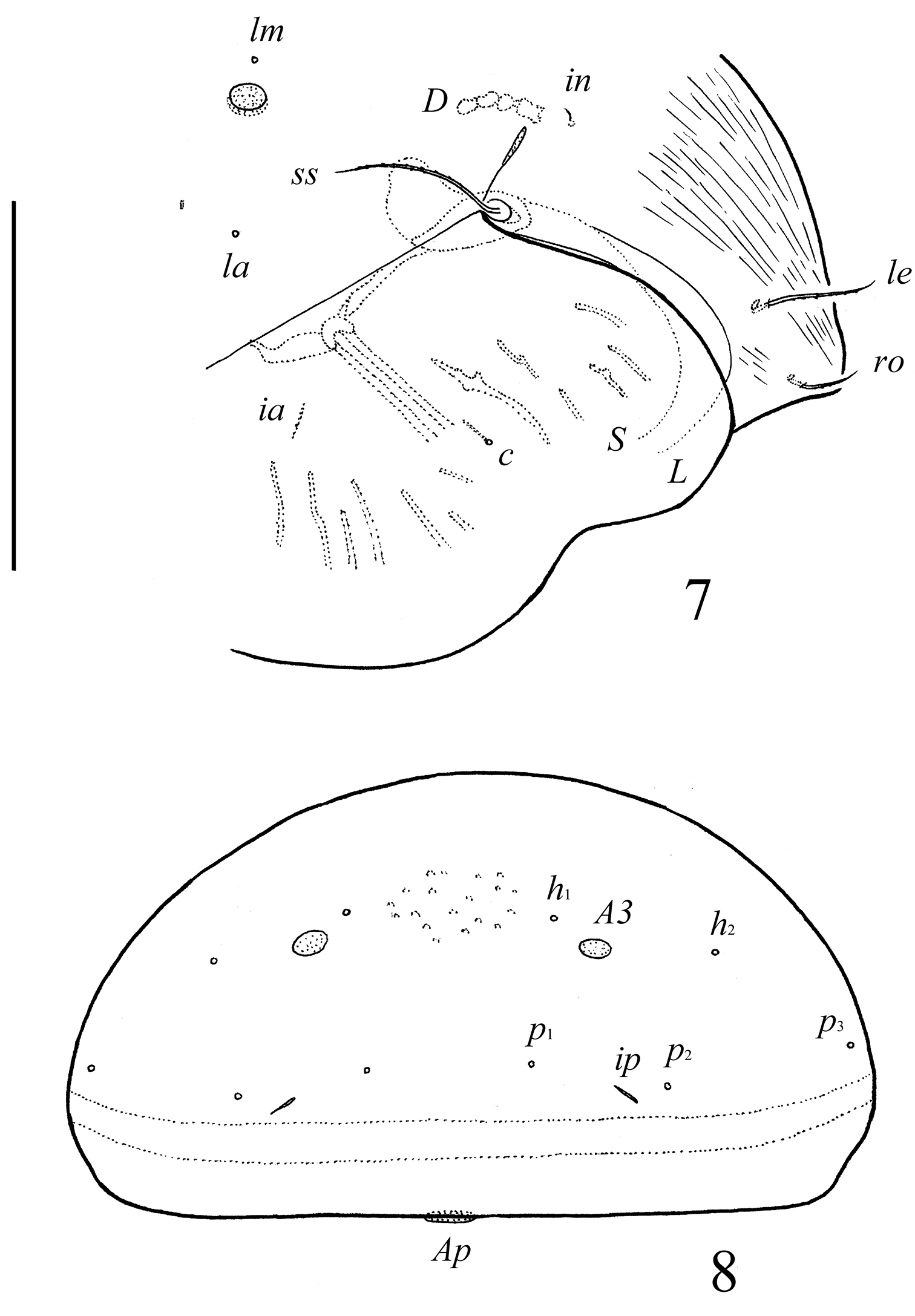
Pergalumna striatiprodorsum sp. n.: 7 dorso-lateral view of prodorsum and anterior part of notogaster and pteromorph (gnathosoma and legs not illustrated) 8 posterior view of notogaster. Scale bars 200 μm.
The holotype is deposited in the Zoological Institute of the Russian Academy of Sciences, St. Petersburg, Russia; seven paratypes are deposited in the collection of the Tyumen State University Museum of Zoology, Tyumen, Russia; one paratype is deposited in the collection of the Centro de Investigación en Estructuras, Microscópicas (CIEMIC), Ciudad de la Investigación, Universidad de Costa Rica, San José, Costa Rica.
This specific name “striatiprodorsum” refers to the striate prodorsum.
In having the striate prodorsum, setiform bothridial setae, absence of anterior notogastral margin and presence of three pairs of oval porose areas (Aa oval, rounded, located close to lm), Pergalumna striatiprodorsum sp. n. is most similar to Pergalumna hawaiiensis hawaiiensis (Jacot, 1934) from the Pacific Islands and Pergalumna strigulata Mahunka, 1978 from Mauritius. However, it differs from Pergalumna hawaiiensis by the short interlamellar setae (versus medium size in Pergalumna hawaiiensis) and striate prodorsum (versus only anterior part of prodorsum with stria in Pergalumna hawaiiensis); from Pergalumna strigulata by the absence of stria on notogaster (versus notogaster striate in Pergalumna strigulata), short interlamellar setae (versus medium size in Pergalumna strigulata) and barbed bothridial setae (versus smooth in Pergalumna strigulata).
Among Neotropical species Pergalumna striatiprodorsum sp. n. is most similar morphologically to Pergalumna decorata Balogh & Mahunka, 1977. However, it differs from the latter by the absence of anterior notogastral margin (versus anterior margin present in Pergalumna decorata), barbed bothridial setae (versus smooth in Pergalumna decorata), absence of stria on notogaster (versus notogaster striate in Pergalumna decorata) and microgranulate body surface and pteromorphs (versus with larger foveoles in Pergalumna decorata).
| 1 | Bothridial setae with clear head | 2 |
| – | Bothridial setae setiform | 13 |
| 2 | Anterior margin of notogaster developed | 3 |
| – | Anterior margin of notogaster not developed | 9 |
| 3 | Surface of prodorsum with numerous longitudinal stria or rugae | 4 |
| – | Surface of prodorsum without stria and rugae | 5 |
| 4 | Surface of notogaster striate; interlamellar setae of medium size; notogaster with four pairs of porose areas (A2 present); body length: 330–460 | Pergalumna striata (Pérez-Íñigo & Baggio, 1980) (see also |
| – | Surface of notogaster foveolate; interlamellar setae absent; notogaster with three pairs of porose areas (A2 absent); body size: 486–527 × 405–437 | Pergalumna complicata Balogh & Mahunka, 1978 (see Fig. 23A–B, not Fig. 24A–E, in |
| 5 | Centro-anterior part of notogaster with specific ornamentation (longitudinal line with lateral lineate branches); lamellar setae minute; notogastral porose areas Aa elongated transversally, very narrow; body size: 282–298 × 199–215 | Pergalumna ornamenta Ermilov, Starý, Sandmann, Marian & Maraun, 2013. Distribution: Ecuador |
| – | Centro-anterior part of notogaster without specific ornamentation; lamellar setae of medium size or long; notogastral porose areas Aa triangular, boot-shaped, rounded or oval | 6 |
| 6 | Interlamellar setae long; notogastral porose areas Aa triangular or boot-shaped | 7 |
| – | Interlamellar setae minute; notogastral porose areas Aa rounded or oval | 8 |
| 7 | Median pore located posterior to the virtual line connecting porose areas A2; notogastral porose areas A1 and A2 small; lyrifissures im located between setal alveoli lm and lp; body length: 470–600 | Pergalumna bryani bryani (Jacot, 1934) (see also |
| – | Median pore located little posterior to the virtual line connecting porose areas A1; notogastral porose areas A1 and A2 of medium size; lyrifissures im located laterally to A1; body size: 620 × 430 | Pergalumna comparanda (Berlese, 1920) (see also |
| 8 | Bothridial setae pointed distally; notogastral porose areas A3 elongated, longer than A1 and A2; body length: 820 | Pergalumna andicola Hammer, 1961. Distribution: Peru |
| – | Bothridial setae rounded distally; notogastral porose areas A1, A2 and A3 rounded, similar in size; body length: 730 | Pergalumna anellata Hammer, 1961. Distribution: Peru |
| 9 | Interlamellar setae long | 10 |
| – | Interlamellar setae minute or represented by alveoli | 11 |
| 10 | Bothridial head barbed medio-distally, stalk smooth; rostral setae inserted laterally; pteromorphal wrinkles indistinct; body size: 320–400 × 270 | Pergalumna numerosa (Sellnick, 1923) (see also see also |
| – | Bothridial head and stalk barbed; rostral setae inserted ventro-laterally; pteromorphal wrinkles clearly visible; body size: 338–392 × 258–320 | Pergalumna bellesii Pérez-Íñigo & Baggio, 1997. Distribution: Brazil |
| 11 | Basal part of prodorsum with longitudinal stria; notogaster with four pairs of porose areas; lamellar setae of medium size; body length: 600 | Pergalumna montana Hammer, 1961. Distribution: Peru and Venezuela |
| – | Basal part of prodorsum without longitudinal stria; notogaster with three pairs of porose areas; lamellar setae short | 12 |
| 12 | Notogastral porose areas Aa oval; median pore present; bothridial setae well barbed; body length: 492–576 | Pergalumna nasica Pérez-Íñigo & Baggio, 1980. Distribution: Brazil and Argentina |
| – | Notogastral porose areas Aa triangular; bothridial setae indistinctly barbed; median pore absent; body length: 468 × 360 | Pergalumna cardosensis Pérez-Íñigo & Baggio, 1986. Distribution: Brazil and Peru |
| 13 | Anterior margin of notogaster developed | 14 |
| – | Anterior margin of notogaster not developed | 17 |
| 14 | Surface of prodorsum and notogaster nearly smooth; body size: 520–676 × 502 | Pergalumna foveolata Hammer, 1973 (see also |
| – | Surface of prodorsum with longitudinal stria or large foveoles; surface of notogaster striate | 15 |
| 15 | Surface of prodorsum with longitudinal stria; interlamellar setae minute; notogastral porose areas Aa located closer to setal alveoli lm than to la; body size: 637–653 × 469–494 | Pergalumna decorata Balogh & Mahunka, 1977. Distribution: Neotropical region |
| – | Surface of prodorsum with large foveoles; interlamellar setae of medium size; notogastral porose areas Aa equal distanced from setal alveoli la and lm | 16 |
| 16 | Rostrum pointed; anal figs striate; body size: 810–860 × 780–810 | Pergalumna decoratissima Pérez-Íñigo & Baggio, 1986. Distribution: Neotropical region |
| – | Rostrum rounded, with lateral tooth on each side; anal figs not striate; body size: 780–962 × 630–747 | Pergalumna paradecoratissima Ermilov & Kalúz, 2012. Distribution: Ecuador |
| 17 | Surface of prodorsum with three striate bands (one basal, transverse and two dorso-lateral, longitudinal striate bands) | 18 |
| – | Surface of prodorsum without three striate bands | 19 |
| 18 | Rostrum pointed; posterior part of notogaster with striate bands; body size: 415–464 × 282–332 | Pergalumna boliviana Ermilov, 2013 (see |
| – | Rostrum rounded; posterior part of notogaster without striate bands; body size: 278 × 213 | Pergalumna passimpunctata Balogh & Mahunka, 1969 (see also |
| 19 | Surface of prodorsum with numerous longitudinal stria | 20 |
| – | Surface of prodorsum without stria | 21 |
| 20 | Whole surface of prodorsum striate; interlamellar setae minute; notogastral porose areas Aa rounded; body size: 630–697 × 448–514 | Pergalumna striatiprodorsum sp. n. Distribution: Costa Rica |
| – | Only basal part of prodorsum striate; interlamellar setae of medium size; notogastral porose areas Aa elongated transversally; body length: 750 | Pergalumna magnipora magnipora (Hammer, 1961). Distribution: Peru |
| 21 | Interlamellar setae of medium size or long | 22 |
| – | Interlamellar setae minute or represented by alveoli | 30 |
| 22 | Rostrum tridentate; body size: 384 × 360 | Pergalumna plumata Pérez-Íñigo & Baggio, 1986. Distribution: Neotropical region |
| – | Rostrum rounded | 23 |
| 23 | Notogastral porose areas A1 of specific structure | 24 |
| – | Notogastral porose areas A1 rounded or oval | 25 |
| 24 | Body surface microgranulate; notogastral porose areas Aa weakly triangular, A1 long, band-shaped, specifically curving; body size: 332–352 × 246–266 | Pergalumna elongatiporosa sp. n. Distribution: Costa Rica |
| – | Body surface smooth; notogastral porose areas Aa elongate oval, A1 with rounded anterior part (with distinct margins) and triangular posterior part (without distinct margins); body size: 448–498 × 315–348 | Pergalumna sura P. Balogh, 1997 (see also |
| 25 | Interlamellar setae longer than bothridial setae | 26 |
| – | Interlamellar setae shorter than bothridial setae | 27 |
| 26 | Adanal setae of medium size, ad3 inserted posteriorly to lyrifissures iad; surface of prodorsum with small tubercles; body size: 1062–1261 × 713–863 | Pergalumna paralongisetosa Ermilov & Kalúz, 2012. Distribution: Ecuador |
| – | Adanal setae minute, ad3 inserted laterally to lyrifissures iad; surface of prodorsum without tubercles; body size: 697–713 × 498–506 | Pergalumna ecuadorensis Ermilov & Kalúz, 2012. Distribution: Ecuador |
| 27 | Notogastral porose areas A1 clearly larger than other porose areas; body length: 400 | Pergalumna melloi Pérez-Íñigo & Baggio, 1994. Distribution: Brazil |
| – | Notogastral porose areas A1 not larger than other porose areas | 28 |
| 28 | Two pairs of notogastral porose areas (A1, A2) observed, A1 very narrowly band-shaped; body size: 394–410 × 295 | Pergalumna horvathorum P. Balogh, 1997. Distribution: Neotropical region |
| – | Three or four pairs of notogastral porose areas observed, A1 rounded or oval | 29 |
| 29 | Three pairs of notogastral porose areas (A3 not observed) present; median pore absent; body size: 390–420 × 342–360 | Pergalumna pauliensis Pérez-Íñigo & Baggio, 1991. Distribution: Brazil |
| – | Four pairs of notogastral porose areas (including A3) present; median pore present; body size: 410–490 × 315–365 | Pergalumna aequalis (Sellnick, 1923). Distribution: Neotropical region |
| 30 | Notogastral porose areas Aa rounded | 31 |
| – | Notogastral porose areas Aa elongated transversally | 32 |
| 31 | Rostrum pointed; notogastral porose areas A1 and A3 oval or rounded; body length: 620 | Pergalumna bifissurata Hammer, 1972. Distribution: Polynesia and Galapagos Islands |
| – | Rostrum rounded; notogastral porose areas A1 and A3 elongated; body length: 500–540 | Pergalumna australis Pérez-Íñigo & Baggio, 1980. Distribution: Brazil and Ecuador |
| 32 | Four pairs of notogastral porose areas present; interlamellar setae represented by alveoli; body size: 276–348 × 228–300 | Pergalumna parva Pérez-Íñigo & Baggio, 1986. Distribution: Brazil |
| – | Three pairs of notogastral porose areas present (A2 absent); interlamellar setae represented by microsetae | 33 |
| 33 | Lamellar setae longer than rostral setae; surface of anterior part of prodorsum granulate; body size: 863–1000 × 680–697 | Pergalumna silvatica Hammer, 1961 (see also |
| – | Lamellar setae shorter than rostral setae; surface of prodorsum not granulate; body size: 468 × 408 | Pergalumna aegra Pérez-Íñigo & Baggio, 1986. Distribution: Brazil |
1 We did not include two Neotropical species, Pergalumna obvia (Berlese, 1914) and Pergalumna curva ventralis (Willmann, 1931) (sensu
We cordially thank Dr. Elizabeth A. Hugo-Coetzee (National Museum, Bloemfontein, South Africa) and one anonymous reviewer for the valuable comments.