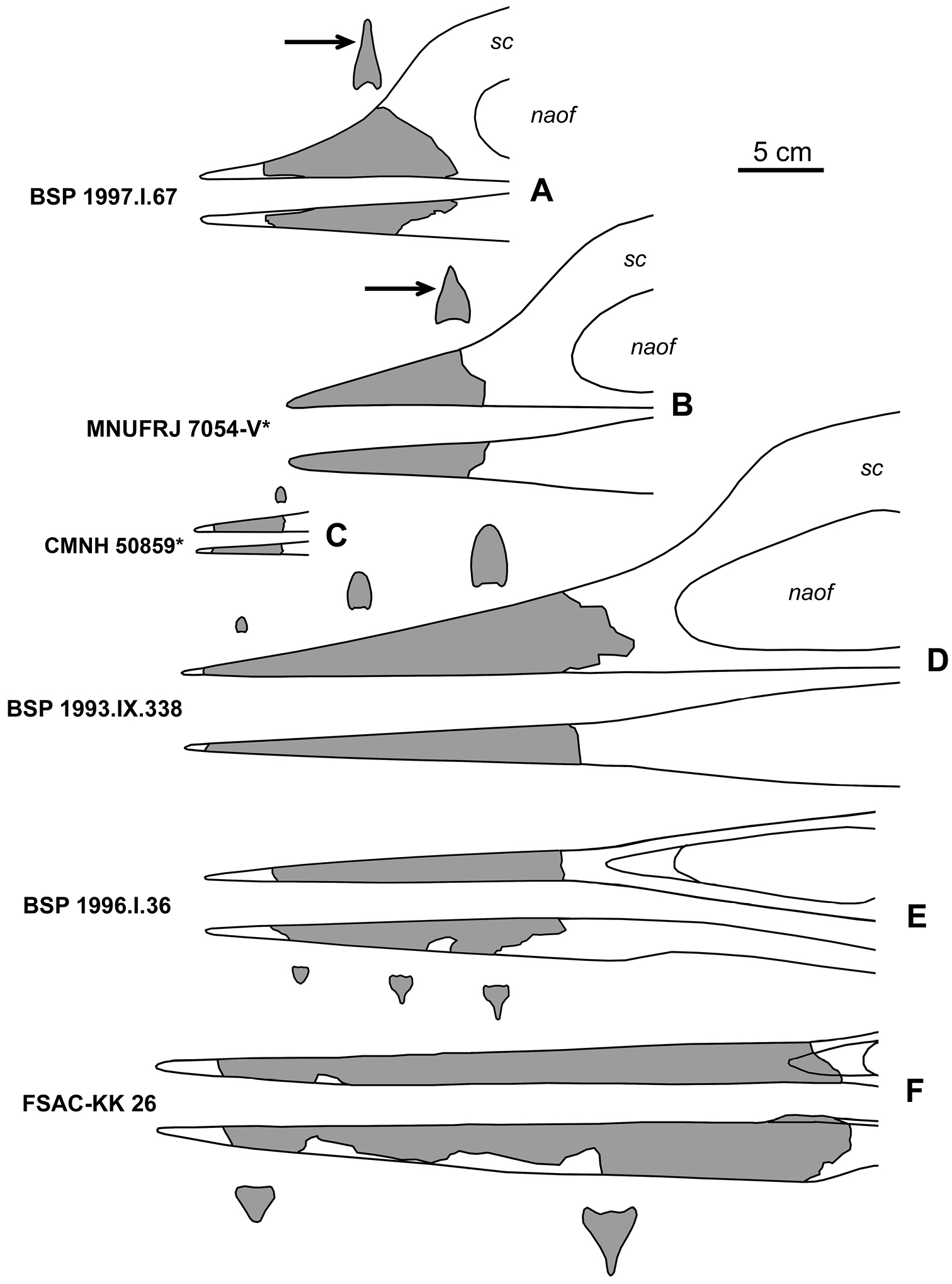
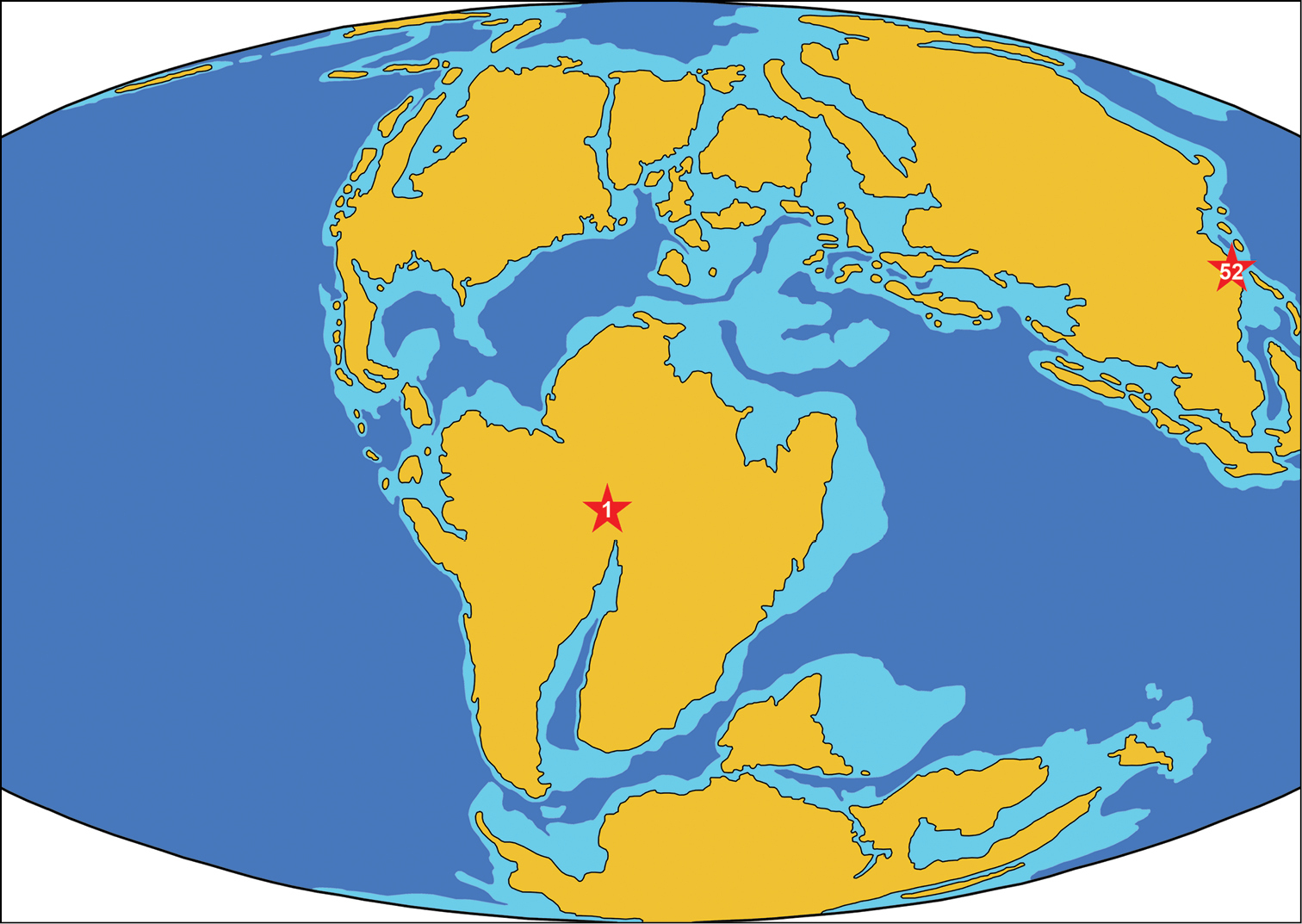
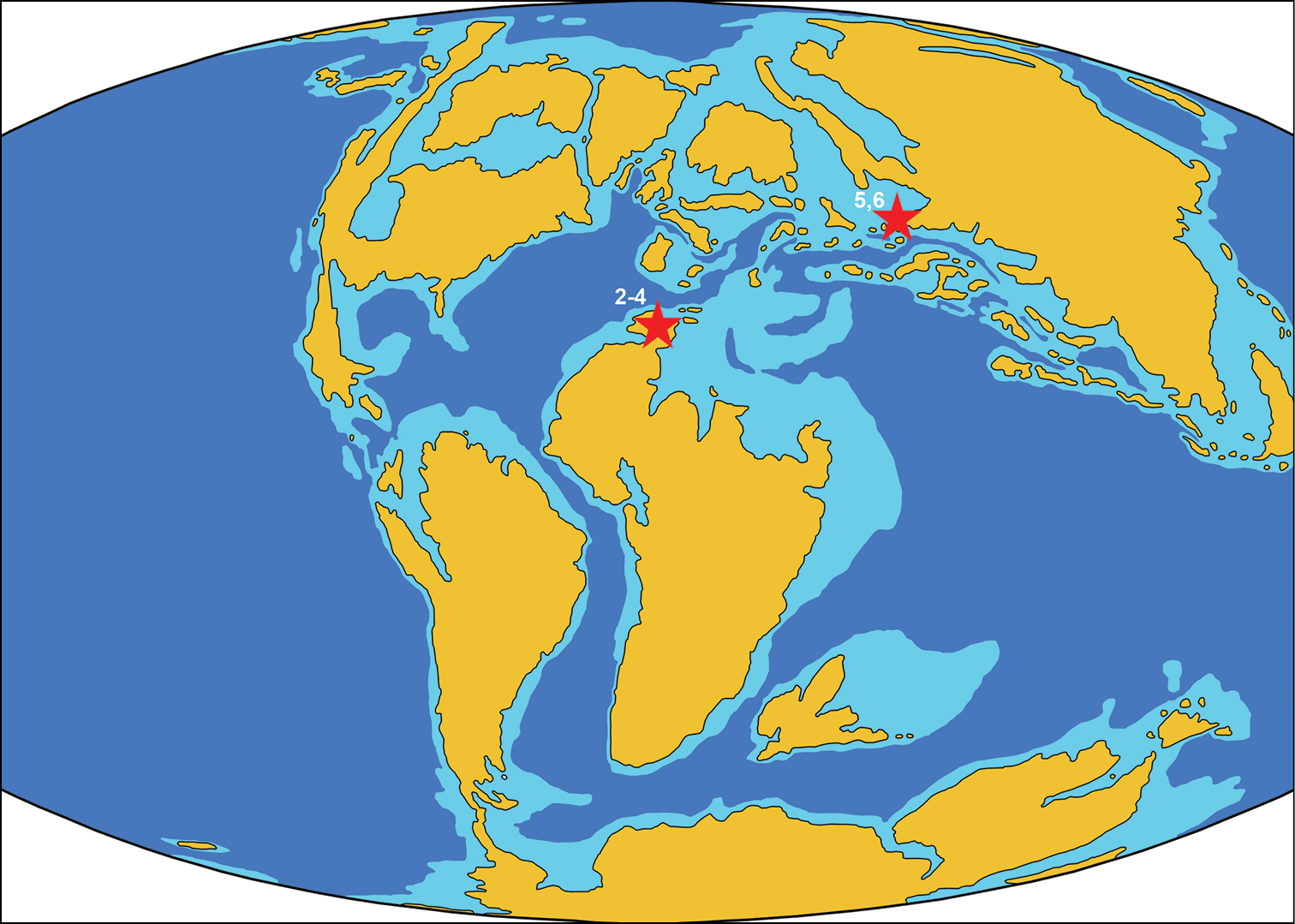
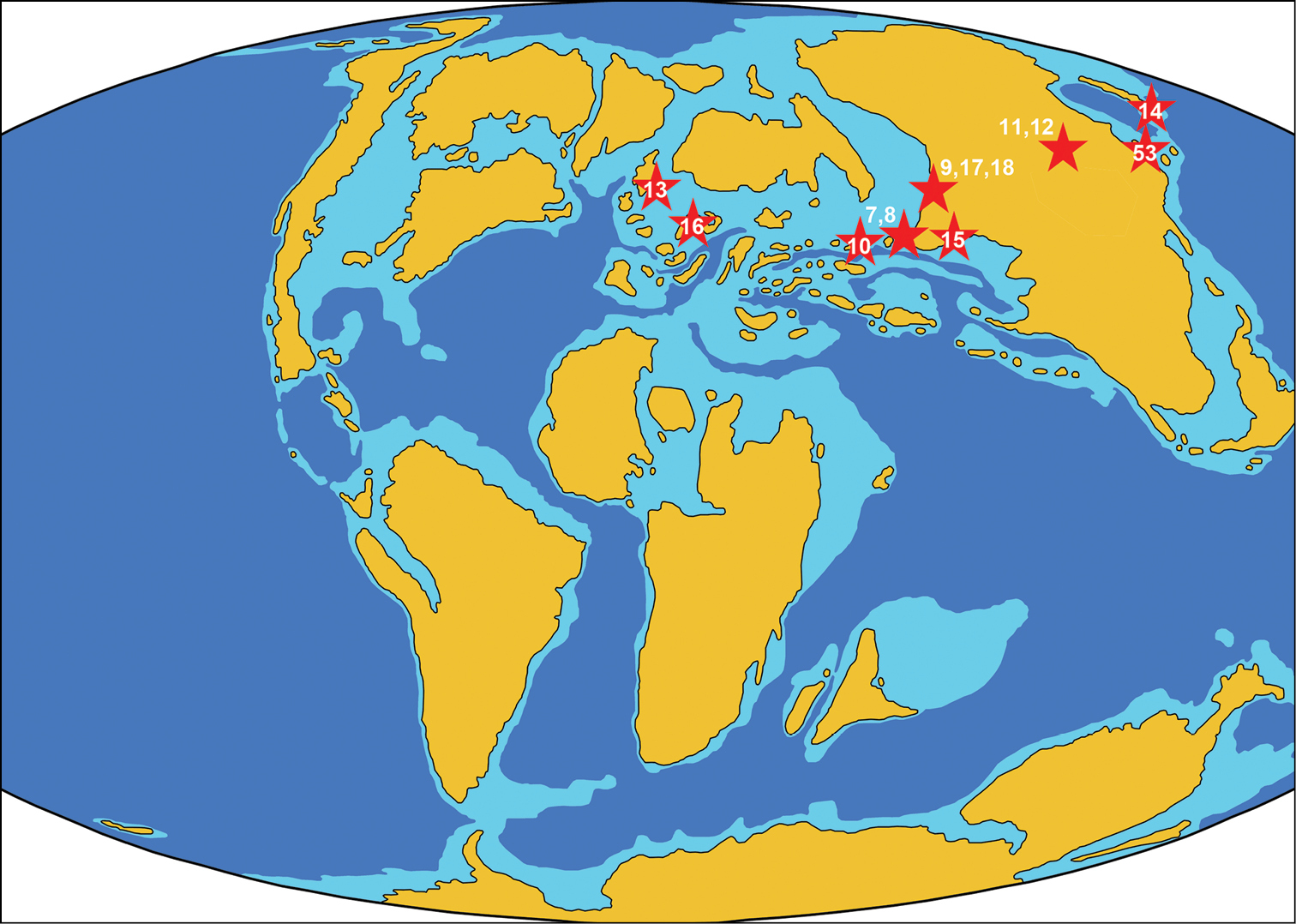
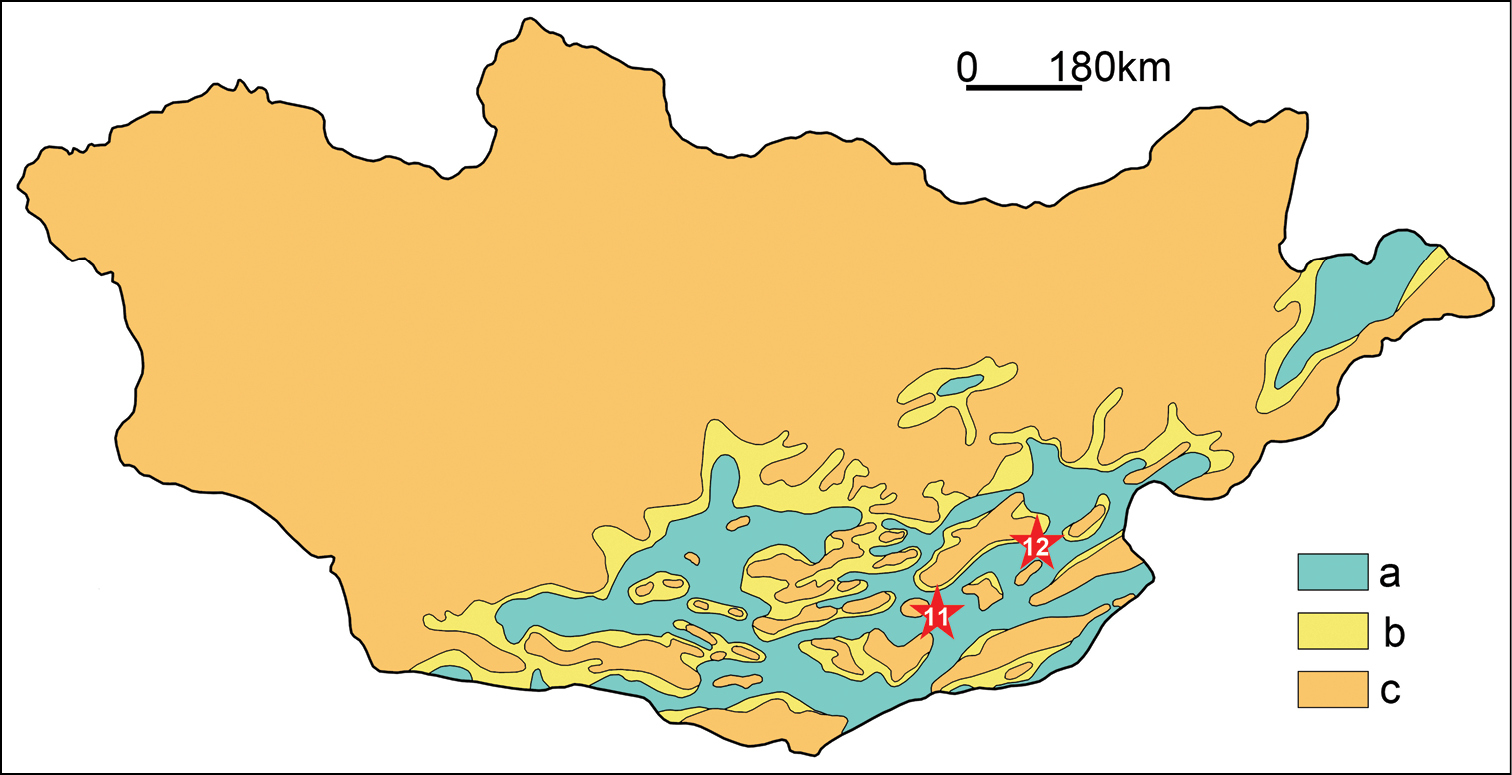
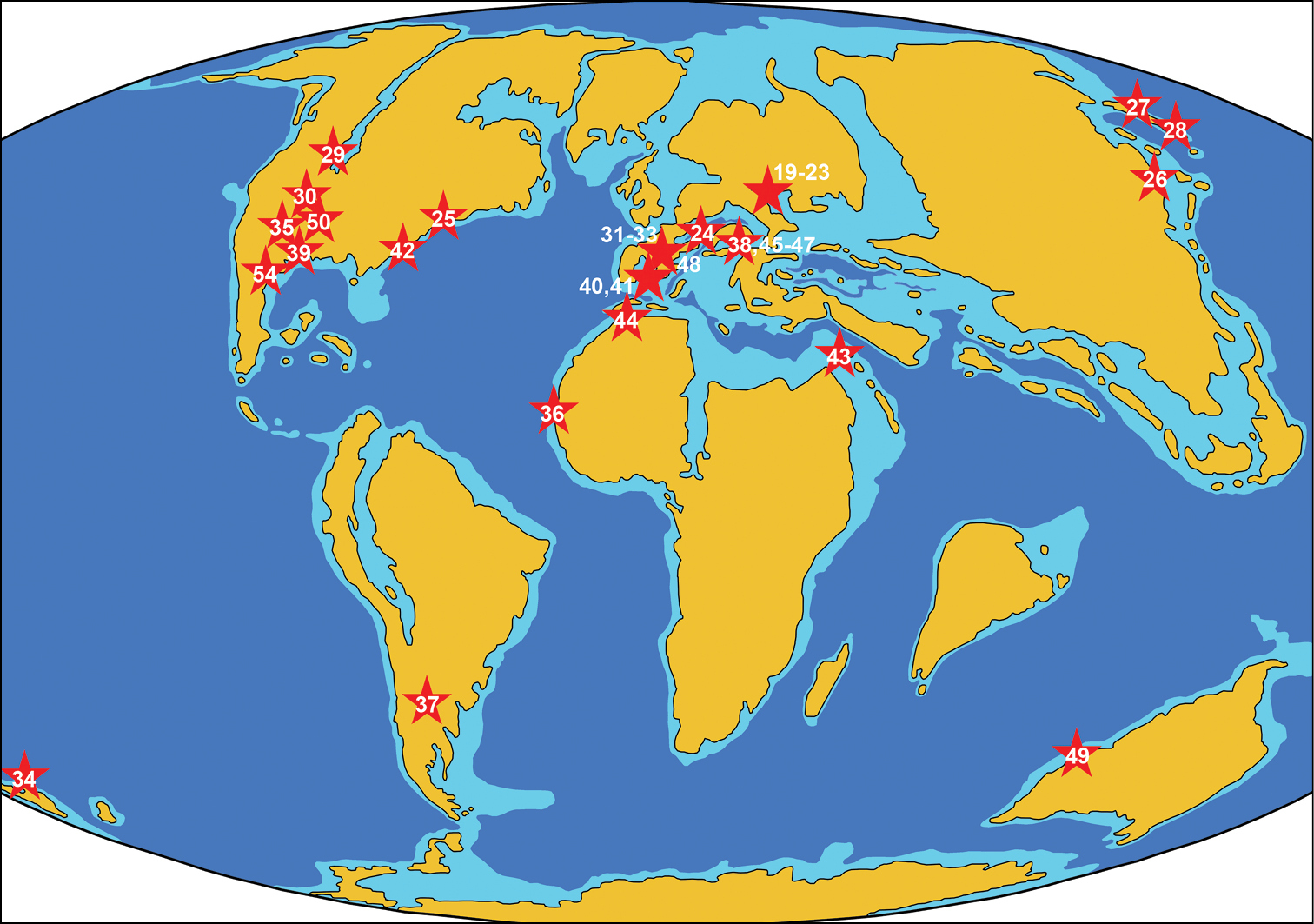
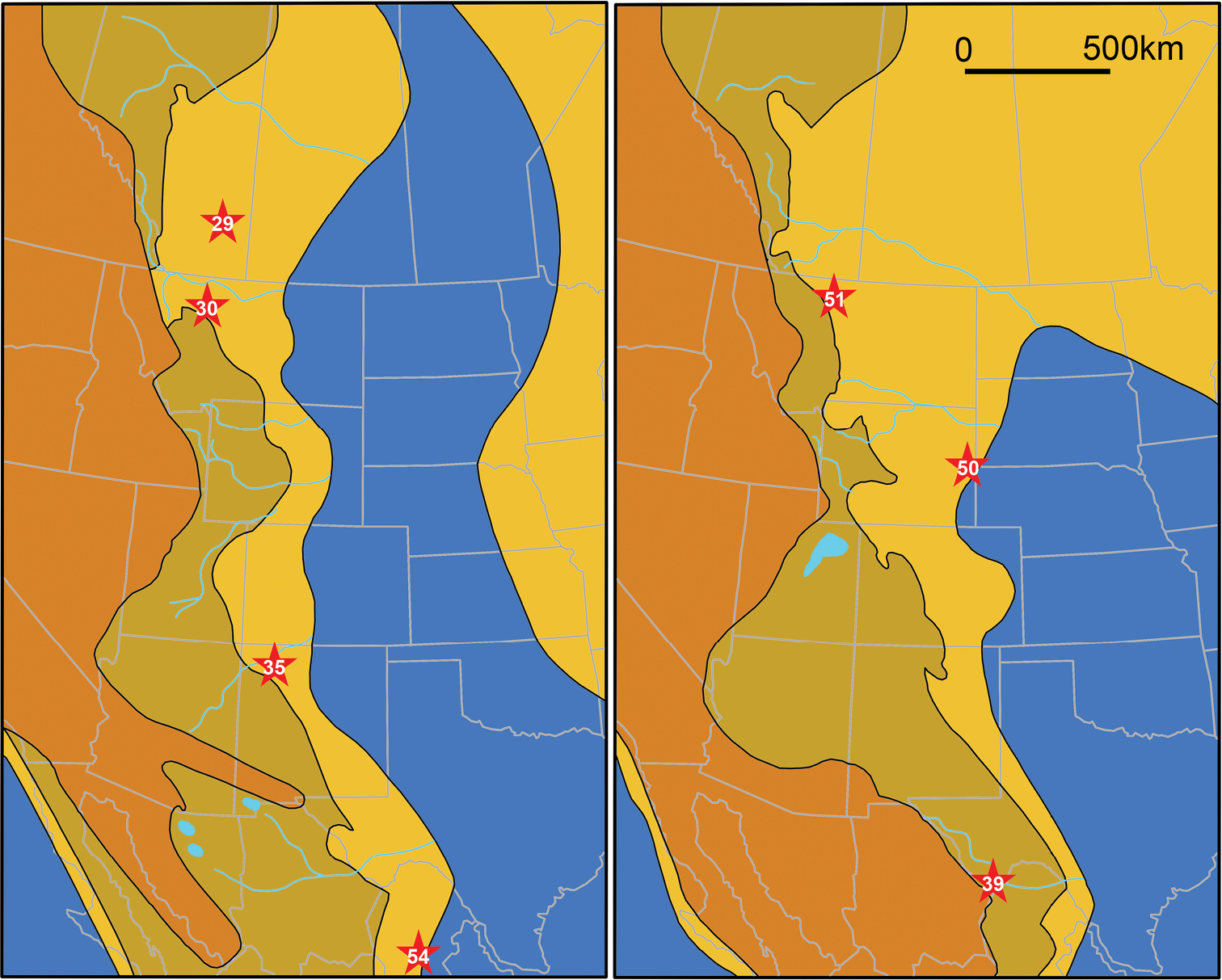
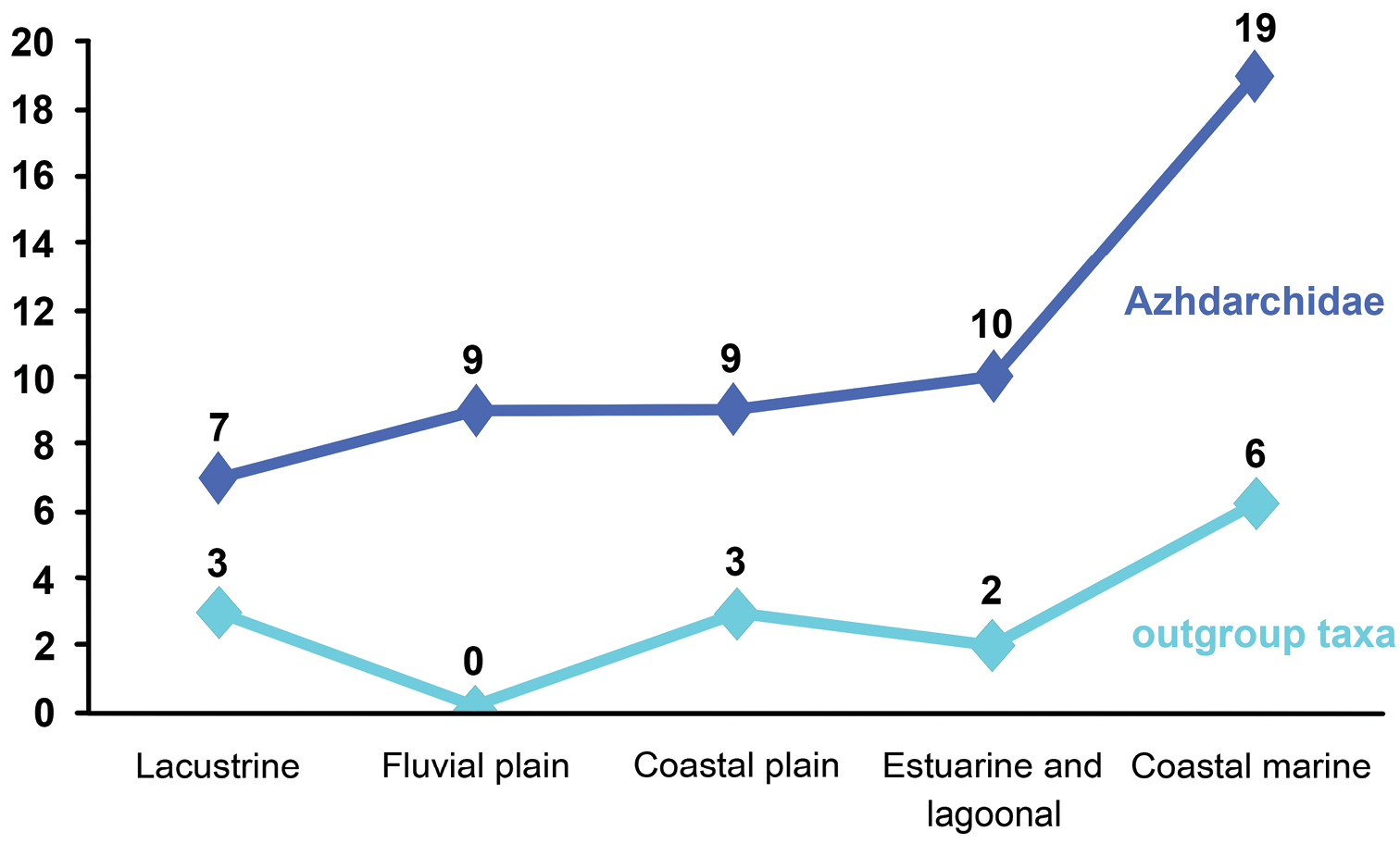
ReferencesAbed AM (1989) On the genesis of the phosphorite-chert association in the Amman Formation in Tel es Sur area, Ruseifa, Jordan. Sciences Géologiques Bulletin 42: 141–153.
Abed AM, Amireh BS (1999) Sedimentology, geochemistry, economic potential and palaeogeography of an Upper Cretaceous phosphorite belt in the southeastern desert of Jordan. Cretaceous Research 20: 119–133. doi:
10.1006/cres.1999.0147Agnolin FL, Novas FE (2012) A carpometacarpus from the Upper Cretaceous of Patagonia sheds light on the ornithurine bird radiation. Paläontologische Zeitschrift 86: 85–89. doi:
10.1007/s12542-011-0112-2Agnolin FL, Powell JE, Novas FE, Kundrát M (2012) New alvarezsaurid (Dinosauria, Theropoda) from uppermost Cretaceous of north-western Patagonia with associated eggs. Cretaceous Research 35: 33–56. doi:
10.1016/j.cretres.2011.11.014Aires ASS, Kellner AWA, Müller RT, Da Silva LR, Pacheco CP, Dias-Da-Silva S (2014) New postcranial elements of the Thalassodrominae (Pterodactyloidea, Tapejaridae) from the Romualdo Formation (Aptian–Albian), Santana Group, Araripe Basin, Brazil. Palaeontology 57: 343–355. doi:
10.1111/pala.12069Alekseev AS, Kopaevich LF, Baraboshkin EYe, Gabdullin RR, Olferiev AG, Yakovishina EV (2005) [Late Cretaceous paleogeography of southern part of East-European Platform and adjacent folded belts. 1. Introduction and stratigraphical basis]. Byuleten' Moskovskogo Obschestva Ispytatelei Prirody, Otdel Geologicheskii 80: 80–92.
Alifanov VR, Averianov AO (2006) On the finding of ornithomimid dinosaurs (Saurischia, Ornithomimosauria) in the Upper Cretaceous beds of Tajikistan. Paleontological Journal 40: 103–108. doi:
10.1134/S0031030106010126Amard B (1996) Occurrence of
Libycoceras ismaeli (Zittel) in the Upper Maastrichtian of Eastern Tademait, Algerian Sahara. Journal of African Earth Sciences 22: 609–615. doi:
10.1016/0899-5362(96)00039-5Ando H, Tamura Y, Takamatsu D (2010) Fourth- to third-order cycles in the Hakobuchi Formation: Shallow-marine Campanian final deposition of the Yezo Group, Nakagawa area, northern Hokkaido, Japan. Island Arc 19: 567–589. doi:
10.1111/j.1440-1738.2010.00716.xAndo H, Tomosugi T, Kanakubo T (2001) Upper Cretaceous to Paleocene Hakobuchi Group, Nakatonbetsu area, northern Hokkaido. Lithostratigraphy and megafossil biostratigraphy. Journal of the Geological Society of Japan 107: 142–162. doi:
10.5575/geosoc.107.142Andres B, Myers TS (2013) Lone Star pterosaurs. Earth and Environmental Science Transactions of the Royal Society of Edinburgh 103: 383–398. doi:
10.1017/S1755691013000303Andres BB, Ji Q (2008) A new pterosaur from the Liaoning Province of China, the phylogeny of the Pterodactyloidea, and convergence in their cervical vertebrae. Palaeontology 51: 453–469. doi:
10.1111/j.1475-4983.2008.00761.xApesteguía S, Agnolin FL, Clalaeson K (2007) Review of Cretaceous dipnoans from Argentina (Sarcopterygii, Dipnoi) with descriptions of new species. Revista del Museo Argentino de Ciencias Naturales 9: 27–40.
Apesteguía S, Jones MEH (2012) A Late Cretaceous “tuatara” (Lepidosauria: Sphenodontinae) from South America. Cretaceous Research 34: 154–160. doi:
10.1016/j.cretres.2011.10.014Arambourg C (1952) Les vertébrés fossiles des gisements de phosphates (Maroc-Algérie-Tunisie). Notes et Mémoires du Service Géologique du Maroc 92: 1–372.
Arambourg C (1954) Sur la présence d'un Ptérosaurien gigantesque dans les Phosphates de Jordanie. Comptes rendus hebdomadaires des séances de l'Académie des sciences 238: 133–134.
Arambourg C (1959)
Titanopteryx philadelphiae nov. gen., nov. sp. Ptérosaurien géant. Notes et Mémoires sur le Moyen-Orient 7: 229–234.
Arambourg C, Dubertret L, Signeux J, Sornay J (1959) Contributions à la stratigraphie et à la paléontologie du Crétacé et du Nummulitique de la marge NW de la Péninsule arabique. Notes et Mémoires sur le Moyen-Orient 7: 193–251.
Arbour VM, Burns ME, Sissons RL (2009) A redescription of the ankylosaurid dinosaur
Dyoplosaurus acutosquameus Parks, 1924 (Ornithischia: Ankylosauria) and a revision of the genus. Journal of Vertebrate Paleontology 29: 1117–1135. doi:
10.1671/039.029.0405Archibald JD, Averianov AO (2005) Mammalian faunal succession in the Cretaceous of the Kyzylkum Desert. Journal of Mammalian Evolution 12: 9–22. doi:
10.1007/s10914-005-4867-3Archibald JD, Averianov AO (2012) Phylogenetic analysis, taxonomic revision, and dental ontogeny of the Cretaceous Zhelestidae (Mammalia: Eutheria). Zoological Journal of the Linnean Society 164: 361–426. doi:
10.1111/j.1096-3642.2011.00771.xArchibald JD, Sues H-D, Averianov AO, King C, Ward DJ, Tsaruk OI, Danilov IG, Rezvyi AS, Veretennikov BG, Khodjaev A (1998) Précis of the Cretaceous paleontology, biostratigraphy and sedimentology at Dzharakuduk (Turonian?-Santonian), Kyzylkum Desert, Uzbekistan. In: Lucas SG, Kirkland JI, Estep JW (Eds) Lower to Middle Cretaceous Terrestrial Ecosystems. Bulletin of the New Mexico Museum of Natural History and Science, 21–28.
Arkhangelsky MS (1999) [On the findings of turtle remains in the Upper Cretaceous of Lower Volga region]. Trudy nauchno-issledovatel'skogo Instituta geologii Saratovskogo gosudarstvennogo universiteta, Novaya seriya 1: 142–144.
Arkhangelsky MS, Averianov AO, Pervushov EM (2007) Short-necked plesiosaurs of the family Polycotylidae from the Campanian of Saratov Region. Paleontological Journal 41: 656–660. doi:
10.1134/S0031030107060093Astibia H, Buffetaut E, Buscalioni AD, Cappetta H, Corral JC, Estes RD, Garcia-Garmilla F, Jaeger J-J, Jimenez-Fuentes E, Le Loeuff J, Mazin J-M, Orue-Etxebarria X, Pereda Suberbiola X, Powell JE, Rage J-C, Rodriguez-Lazaro J, Sanz JL, Tong H (1991) The fossil vertebrates from Laño (Basque Country, Spain); new evidence on the composition and affinities of the Late Cretaceous of continental Europe. Terra Nova 2: 460–466. doi:
10.1111/j.1365-3121.1990.tb00103.xAstibia H, Corral JC, Murelaga X, Orue-Etxebarria X, Pereda Suberbiola X (1999) Geology and paleontology of the Upper Cretaceous vertebrate-bearing beds of the Laño quarry (Basque-Cantabrian Region, Iberian Peninsula). Estudios del Museo de Ciencias Naturales de Alava 14: 1–380.
Averianov AO (2000) Mammals from the Mesozoic of Kirgizstan, Uzbekistan, Kazakhstan and Tadzhikistan. In: Benton MJ, Shishkin MA, Unwin DM, Kurochkin EN (Eds) The Age of Dinosaurs in Russia and Mongolia. Cambridge University Press, 627–652.
Averianov AO (2001) Systematics of the Cretaceous–Paleogene chimaeroid fish of the genus
Elasmodus (Chondrichthyes, Holocephali). Paleontological Journal 35: 289–300.
Averianov AO (2002a) An ankylosaurid (Ornithischia: Ankylosauria) braincase from the Upper Cretaceous Bissekty Formation of Uzbekistan. Bulletin de l'Institut Royal des Sciences Naturelles de Belgique, Sciences de la Terre 72: 97–110.
Averianov AO (2002b) Review of Mesozoic and Cenozoic sea turtles from the former USSR. Russian Journal of Herpetology 9: 137–154.
Averianov AO (2004) New data on Cretaceous flying reptiles (Pterosauria) from Russia, Kazakhstan, and Kyrgyzstan. Paleontological Journal 38: 426–436.
Averianov AO (2007a) Mid-Cretaceous ornithocheirids (Pterosauria, Ornithocheiridae) from Russia and Uzbekistan. Paleontological Journal 41: 79–86. doi:
10.1134/S003103010701008XAverianov AO (2007b) New records of azhdarchids (Pterosauria, Azhdarchidae) from the Late Cretaceous of Russia, Kazakhstan, and Central Asia. Paleontological Journal 41: 189–197. doi:
10.1134/S0031030107020098Averianov AO (2007c) Theropod dinosaurs from the Late Cretaceous of North-East Aral Sea area, Kazakhstan. Cretaceous Research 28: 532–544. doi:
10.1016/j.cretres.2006.08.008Averianov AO (2008) [Superorder Pterosauria]. In: Ivakhnenko MF, Kurochkin EN (Eds) [Fossil Vertebrates of Russia and Adjacent Territories Fossil Reptiles and Birds Part 1. ] GEOS, Moscow, 319–342.
Averianov AO (2009) [Ankylosaur (Ornithischia, Ankylosauridae) humerus from the Late Cretaceous of Kazakhstan]. In: Shishkin MA, Tverdokhlebov VP (Eds) [Researches on paleontology and biostratigraphy of ancient continental deposits (Memories of Professor Vitalii G Ochev)]. «Nauchnaya Kniga» Publishers, Saratov, 78–80.
Averianov AO (2010) The osteology of
Azhdarcho lancicollis Nesov, 1984 (Pterosauria, Azhdarchidae) from the Late Cretaceous of Uzbekistan. Proceedings of the Zoological Institute RAS 314: 264–317.
Averianov AO (2012)
Ornithostoma sedgwicki--valid taxon of azhdarchoid pterosaurs. Proceedings of the Zoological Institute of the Russian Academy of Sciences 316: 40–49. doi:
10.1134/S0031030112050036Averianov AO, Alifanov VR (2012) New data on duck-billed dinosaurs (Ornithischia, Hadrosauridae) from the Upper Cretaceous of Tajikistan. Paleontological Journal 46: 512–519.
Averianov AO, Archibald JD (2005) Mammals from the mid-Cretaceous Khodzhakul Formation, Kyzylkum Desert, Uzbekistan. Cretaceous Research 26: 593–608. doi:
10.1016/j.cretres.2005.03.007Averianov AO, Archibald JD (2013a) New material and reinterpretation of the Late Cretaceous eutherian mammal
Paranyctoides from Uzbekistan. Acta Palaeontologica Polonica 58: 17–23.
Averianov AO, Archibald JD (2013b) Variation and taxonomy of Asiamerican eutherian mammal
Paranyctoides. Canadian Journal of Earth Sciences 50: 895–903. doi:
10.1139/cjes-2013-0051Averianov AO, Archibald JD, Ekdale EG (2010) New material of the Late Cretaceous deltatheroidan mammal
Sulestes from Uzbekistan and phylogenetic reassessment of the metatherian-eutherian dichotomy. Journal of Systematic Palaeontology 8: 301–330. doi:
10.1080/14772011003603499Averianov AO, Archibald JD, Dyke GJ (in press) A new eutherian mammal from the Late Cretaceous of Kazakhstan. Acta Palaeontologica Polonica:
Averianov AO, Arkhangelsky MS, Pervushov EM (2008) A new Late Cretaceous azhdarchid (Pterosauria, Azhdarchidae) from the Volga Region. Paleontological Journal 42: 634–642.
Averianov AO, Arkhangelsky MS, Pervushov EM, Ivanov AV (2005) A new record of an azhdarchid (Pterosauria: Azhdarchidae) from the Upper Cretaceous of the Volga Region. Paleontological Journal 39: 433–439.
Averianov AO, Atabekyan AA (2005) The first discovery of a flying reptile (Pterosauria) in Armenia. Paleontological Journal 39: 210–212.
Averianov AO, Leshchinskiy SV, Skutschas PP, Rezvyi AS (2003) [Pterosaur teeth from the Lower Cretaceous of Russia and Uzbekistan]. Sovremennaya Gerpetologiya 2: 5–11.
Averianov AO, Panteleyev AV (2009) [Coracoid fragments of a large azhdarchid (Pterosauria, Azhdarchidae) from the Campanian of Saratov Province]. In: Shishkin MA, Tverdokhlebov VP (Eds) [Researches on paleontology and biostratigraphy of ancient continental deposits (Memories of Professor Vitalii G Ochev)]. «Nauchnaya Kniga» Publishers, Saratov, 74–77.
Averianov AO, Popov EV (1995) A new species of chimaeroid fish from the Upper Cretaceous of the Saratov Region, Russia. Palaeontology 38: 659–664.
Averianov AO, Popov EV (2014) A pterosaurian vertebra from the Upper Cretaceous of the Saratov Region. Paleontological Journal 48: 326–329. doi:
10.1134/S0031030114030034Averianov AO, Sues H-D (2007) A new troodontid (Dinosauria: Theropoda) from the Cenomanian of Uzbekistan, with a review of troodontid records from the territories of the former Soviet Union. Journal of Vertebrate Paleontology 27: 87–98. doi:
10.1671/0272-4634(2007)27[87:ANTDTF]2.0.CO;2Averianov AO, Sues H-D (2009) First record of a basal neoceratopsian dinosaur from the Late Cretaceous of Kazakhstan. Acta Palaeontologica Polonica 54: 553–556. doi:
10.4202/app.2008.0079Averianov AO, Sues H-D (2012a) Correlation of Late Cretaceous continental vertebrate assemblages in Middle and Central Asia. Journal of Stratigraphy 36: 462–485.
Averianov AO, Sues H-D (2012b) Skeletal remains of Tyrannosauroidea (Dinosauria: Theropoda) from the Bissekty Formation (Upper Cretaceous: Turonian) of Uzbekistan. Cretaceous Research 34: 284–297. doi:
10.1016/j.cretres.2011.11.009Averianov AO, Yarkov AA (2000) Some turtle remains from the Cretaceous and Paleogene of Volgograd Region, Russia. Russian Journal of Herpetology 7: 161–166.
Averianov AO, Yarkov AA (2004a) New turtle remains from the Late Cretaceous and Paleogene of Volgograd Region, Russia. Russian Journal of Herpetology 11: 41–50.
Averianov AO, Yarkov AA (2004b) On the occurrence of a giant flying reptile (Pterosauria) in the terminal Late Cretaceous of the Lower Volga Region. Paleontological Journal 38: 669–671.
Avnimelech MA (1949) On vertebrate remains in Senonian phosphate beds in Transjordan. Eclogae geologicae Helvetiae 42: 486–490.
Báez AM, Moura GJB, Gómez RO (2009) Anurans from the Lower Cretaceous Crato Formation of northeastern Brazil: implications for the early divergence of neobatrachians. Cretaceous Research 30: 829–846. doi:
10.1016/j.cretres.2009.01.002Baird D, Galton PM (1981) Pterosaur bones from the Upper Cretaceous of Delaware. Journal of Vertebrate Paleontology 1: 67–71. doi:
10.1080/02724634.1981.10011880Bakhurina NN, Unwin DM (1995) A survey of pterosaurs from the Jurassic and Cretaceous of the former Soviet Union and Mongolia. Historical Biology 10: 197–245. doi:
10.1080/10292389509380522Barck A (1992) Paleontology of the Glen Rose Formation (Lower Cretaceous), Hood County, Texas. Texas Journal of Science 44: 3–24.
Bardet N (1994) Extinction events among Mesozoic marine reptiles. Historical Biology 7: 313–324. doi:
10.1080/10292389409380462Bardet N, Pereda Suberbiola X (2002) Marine reptiles from the Late Cretaceous Phosphates of Jordan: palaeobiogeographical implications. Geodiversitas 24: 831–839.
Bardet N, Pereda Suberbiola X, Iarochene M, Bouyahyaoui F, Bouya B, Amaghzaz M (2004)
Mosasaurus beaugei Arambourg, 1952 (Squamata, Mosasauridae) from the Late Cretaceous phosphates of Morocco. Geobios 37: 315–324. doi:
10.1016/j.geobios.2003.02.006Barrett PM, Butler RJ, Edwards NP, Milner AR (2008) Pterosaur distribution in time and space: an atlas. Zitteliana B 28: 61–107.
Barrett PM, Evans SE (2002) A reassessment of the Early Cretaceous reptile '
Patricosaurus merocratus' Seeley from the Cambridge Greensand, Cambridgeshire, UK. Cretaceous Research 23: 231–240. doi:
10.1006/cres.2002.0312Barsbold R (1981) [Toothless carnivorous dinosaurs of Mongolia]. Trudy Sovmestnoi Sovetsko-Mongol'skoi Paleontologicheskoi Ekspeditsii 15: 28–39.
Barsbold R (1983) [Carnivorous dinosaurs from the Cretaceous of Mongolia]. Trudy Sovmestnoi Sovetsko-Mongol'skoi Paleontologicheskoi Ekspeditsii 19: 1–120.
Barsbold R, Perle A (1980) Segnosauria, a new infraorder of carnivorous dinosaurs. Acta Palaeontologica Polonica 25: 187–195.
Batten DJ (2007) Spores and pollen from the Crato Formation: biostratigraphic and palaeoenvironmental implications. In: Martill DM, Bechly G, Loveridge RF (Eds) Fossil Beds of Brazil: Window into an Ancient World. Cambridge University Press, New Yourk, 566–573.
Bazhanov VS (1972) [First Mesozoic Mammalia (
Beleutinus orlovi Bashanov) from the USSR]. Teriologiya. 74–80.
Beavan NR, Russell AP (1999) An elasmobranch assemblage from the terrestrial-marine transitional Lethbridge Coal Zone (Dinosaur Park Formation: Upper Campanian), Alberta, Canada. Journal of Paleontology 73: 494–503.
Bennett SC (1989) A pteranodontid pterosaur from the Early Cretaceous of Peru, with comments on the relationships of Cretaceous pterosaurs. Journal of Paleontology 63: 669–677.
Bennett SC (1994) Taxonomy and systematics of the Late Cretaceous pterosaur
Pteranodon (Pterosauria, Pterodactyloidea). Occasional Papers of the Museum of Natural History, the University of Kansas 169: 1–70.
Bennett SC (2001) The osteology and functional morphology of the Late Cretaceous pterosaur
Pteranodon. Part I. General description and osteology. Palaeontographica, Abteilung A 260: 1–112.
Bennett SC, Long JA (1991) A large pterodactyloid pterosaur from the Late Cretaceous (Late Maastrichtian) of Western Australia. Records of the Western Australian Museum 15: 435–443.
Benson RBJ, Brusatte SL, Hutt S, Naish D (2009) A new large basal tetanuran (Dinosauria: Theropoda) from the Wessex Formation (Barremian) of the Isle of Wight, England. Journal of Vertebrate Paleontology 29: 612–615. doi:
10.1671/039.029.0202Benson RBJ, Mannion PD, Butler RJ, Upchurch P, Goswami A, Evans SE (2013) Vertebrate palaeobiodiversity patterns and the impact of sampling bias. Palaeogeography, Palaeoclimatology, Palaeoecology 372: 88–107. doi:
10.1016/j.palaeo.2012.10.028Benton MJ (1989) Patterns of evolution and extinction in vertebrates. In: Allen KC, Briggs DEG (Eds) Evolution and the Fossil Record. Belhaven Press, London, 218–241.
Benton MJ, Cook E, Grigorescu D, Popa E, Tallodi E (1997) Dinosaurs and other tetrapods in an Early Cretaceous bauxite filled fissure, northwestern Romania. Palaeogeography, Palaeoclimatology, Palaeoecology 130: 275–292. doi:
10.1016/S0031-0182(96)00151-4Benton MJ, Spencer PS (1995) Fossil Reptiles of Great Britain. Chapman and Hall, London, 386 pp. doi:
10.1007/978-94-011-0519-4Bilotte M, Ségura F (1991) Stratigraphie séquentielle des séries du sénonien supérieur dans la zone sous-pyrénéenne (Petites-Pyrénées et Dômes annexes, SW France). Comptes rendus de l'Académie des sciences Série 2, Mécanique, Physique, Chimie, Sciences de l'univers, Sciences de la Terre 312: 393–398.
Blows WT (1995) The Early Cretaceous brachiosaurid dinosaurs
Ornithopsis and
Eucamerotus from the Isle of Wight, England. Palaeontology 38: 187–197.
Bock WJ, Buehler P (1996) Nomenclature of Cretaceous birds from Romania. Cretaceous Research 17: 509–514. doi:
10.1006/cres.1996.0031Bogan S, Taverne L, Agnolin FL (2010) First fossil record of an amiid fish (Halecomorphi, Amiidae) from the Latest Cretaceous of Patagonia, Argentina, and comments on the status of
Pappichthys patagonica Ameghino, 1906 (Teleostei, Osteoglossidae). Bulletin de L'Institut Royal des Sciences Naturelles de Belgique, Sciences de la Tere 80:
Bogolyubov NN (1911) [From the history of plesiosaurs in Russia]. Uchenye zapiski Imperatorskogo Moskovskogo Universiteta, Otdel estesvennoistoricheskii: 1–412.
Bogolyubov NN (1914) [On the pterodactyl vertebra from the Upper Cretaceous deposits of Saratov Province]. Ezhegodnik po Geologii i Mineralogii Rossii 16: 1–7.
Breithaupt BH (1982) Paleontology and paleoecology of the Lance Formation (Maastrichtian), east flank of Rock Springs uplift, Sweetwater County, Wyoming. Contributions to Geology, University of Wyoming 21: 123–151.
Brinkman DB (1990) Paleoecology of the Judith River Formation (Campanian of Dinosaur Provincial Park, Alberta, Canada: evidence from vertebrate microfossil localities. Palaeogeography, Palaeoclimatology, Palaeoecology 78: 37–54. doi:
10.1016/0031-0182(90)90203-JBrinkman DB, Braman DR, Neuman AG, Ralrick PE, Sato T (2005a) A vertebrate assemblage from the marine shales of the Letherbridge Coal Zone. In: Currie PJ, Koppelhus EB (Eds) Dinosaur Provincial Park: A Spectacular Ancient Ecosystem Revealed. Bloomington, Indianapolis, Indiana University Press, 486–500.
Brinkman DB, Nesov LA, Peng J-H (1994)
Khunnuchelys gen. nov., a new trionychid (Testudines: Trionychidae) from the Late Cretaceous of Inner Mongolia and Uzbekistan. Canadian Journal of Earth Sciences 30: 2214–2223. doi:
10.1139/e93-192Brinkman DB, Russell AP, Peng J-H (2005b) Vertebrate microfossil sites and their contribution to studies of paleoecology. In: Currie PJ, Koppelhus EB (Eds) Dinosaur Provincial Park: A Spectacular Ancient Ecosystem Revealed. Indiana University Press, Bloomington, Indianapolis, 88–98.
Brito PM (1997) Révision des Aspidorhynchidae (Pisces-Actinopterygii) du Mésozoïque: ostéologie et relations phylogénétiques. Geodiversitas 19: 681–772.
Brodkorb P (1963) Birds from the Upper Cretaceous of Wyoming. In: Sibley CG, Hickey JJ, Hickey MB (Eds) Proceedings of the XIIIth International Ornithological Congress, Ithaca, 17–24 June 1962 Volume 1. American Ornithologists Union, Baton Rouge, 55–70.
Brusatte S, Vremir M, Csiki-Sava Z, Turner AH, Watanabe A, Erickson GM, Norell MA (2013) The osteology of
Balaur bondoc, an island-dwelling dromaeosaurid (Dinosauria, Theropoda) from the late Cretaceous of Romania. Bulletin of the American Museum of Natural History 374: 1–100. doi:
10.1206/798.1Buffetaut E (1979) Revision der Crocodylia (Reptilia) aus den Gosau-Schichten (Ober-Kreide) von Österreich. Beiträge zur Paläontologie von Österreich 6: 89–105.
Buffetaut E (1989) Erster Nachweis von Choristodera (Reptilia, Diapsida) in der Oberkreide Europas: Champsosaurierwirbel aus den Gosau-Schichten (Campan) Niederösterreichs. Sitzungberichte Österreichische Akademie der Wissenschaften, Mathematisch-naturwissenschaftliche Klasse 197: 389–394.
Buffetaut E (1998) First evidence of enantiornithine birds from the Upper Cretaceous of Europe: postcranial bones from Cruzy (Hérault, southern France). Oryctos 1: 131–136.
Buffetaut E (1999) Pterosauria from the Upper Cretaceous of Laño (Iberian Peninsula): a preliminary comparative study. In: Astibia H, Corral JC, Murelaga X, Orue-Etxebarria X, Pereda Suberbiola X (Eds) Geology and Palaeontology of the Upper Cretaceous Vertebrate-Bearing Beds of the Laño Quarry (Basque-Cantabrian Region, Iberian Peninsula). Esudios del Museo Ciencias Naturales de Alava 14, Número especial 1, 289–284.
Buffetaut E (2001) An azhdarchid pterosaur from the Upper Cretaceous of Cruzy (Herault, France). Comptes Rendus de l'Académie des Sciences, Series II, Fascicule A: Sciences de la Terre et des planetes 333: 357–361.
Buffetaut E (2004) A giant pterosaur from the Lower Cretaceous of the eastern Paris Basin. Bulletin de la Société géologique de France 175: 573–577. doi:
10.2113/175.6.573Buffetaut E (2005) Late Cretaceous vertebrates from the Saint-Chinian area (southern France): a review of previous research and an update on recent finds Acta Palaeontologica Romaniae 5: 39–48.
Buffetaut E (2008) Late Cretaceous pterosaurs from France: a review. Zitteliana B 28: 249–255.
Buffetaut E (2010) A basal bird from the Campanian (Late Cretaceous) of Dinosaur Provincial Park (Alberta, Canada). Geological Magazine 147: 469–472. doi:
10.1017/S0016756810000129Buffetaut E (2011)
Samrukia nessovi, from the Late Cretaceous of Kazakhstan: A large pterosaur, not a giant bird. Annales de Paleontologie 97: 133–138. doi:
10.1016/j.annpal.2011.10.001Buffetaut E (2012) An early azhdarchid pterosaur from the Lower Cretaceous of the eastern Paris basin. Bulletin de la Société géologique de France 183: 525–528. doi:
10.2113/gssgfbull.183.6.525Buffetaut E, Clarke JB, Le Loeuff J (1996) A terminal Cretaceous pterosaur from the Corbieres (southern France) and the problem of pterosaur extinction. Bulletin de la Société géologique de France 167: 753–759Buffetaut E, Grigorescu D, Csiki Z (2002) A new giant pterosaur with a robust skull from the latest Cretaceous of Romania. Naturwissenschaften 89: 180–184.
Buffetaut E, Grigorescu D, Csiki Z (2003) Giant azhdarchid pterosaurs from the terminal Cretaceous of Transylvania (western Romania). In: Buffetaut E, Mazin J-M (Eds) Evolution and Palaeobiology of Pterosaurs. Geological Society Special Publication, n. 217, 91–104.
Buffetaut E, Laurent Y, Le Loeuff J, Bilotte M (1997) A terminal Cretaceous giant pterosaur from the French Pyrenees. Geological Magazine 134: 553–556. doi:
10.1017/S0016756897007449Buffetaut E, Le Lœuff J, Mechin P, Mechin-Salessy A (1995) A large French Cretaceous bird. Nature 377: 110. doi:
10.1038/377110a0Buffetaut E, Le Lœuff J, Tong H, Duffaud S, Cavin L, Garcia G, Ward DJ, Cruzy lAcaepd (1999) Un nouveau gisement de vertébrés du Crétacé supérieur à Cruzy (Hérault, sud de la France). Comptes Rendus de l'Académie des Sciences, Series II, Fascicule A: Sciences de la Terre et des plantes 328: 203–208.
Buffetaut E, Mechin P, Mechin-Salessy A (2000) An archaic bird (Enantiornithes) from the Upper Cretaceous of Provence (southern France). Comptes Rendus de l'Académie des Sciences, Series II, Fascicule A: Sciences de la Terre et des planètes 331: 557–561.
Buffetaut E, Mechin P, Mechin-Salessy A (2006) An azhdarchid pterosaur from the Upper Cretaceous of Provence (southern France). In: Csiki Z (Ed) Mesozoic and Cenozoic vertebrates and paleoenvironments, tributes the career of Professor Dan Grigorescu. Editura Ars Docendi, Bucharest, 95–100.
Buffetaut E, Osi A, Prondvai E (2011) The pterosaurian remains from the Grünbach Formation (Campanian, Gosau Group) of Austria: a reappraisal of ‘
Ornithocheirus buenzeli’’. Geological Magazine 148: 334–339. doi:
10.1017/S0016756810000981Buffetaut E, Ouaja M (2002) A new specimen of
Spinosaurus (Dinosauria, Theropoda) from the Lower Cretaceous of Tunisia, with remarks on the evolutionary history of the Spinosauridae. Bulletin de la Société géologique de France 173: 415–421. doi:
10.2113/173.5.415Buisonje PH, de (1980)
Santanadactylus brasilensis nov. gen., nov. sp., a long necked, large pterosaurier from the Aptian of Brazil. Proceedings of the Koninklijke Nederlandse Akademie van Wetenschappen, Series B 83: 145–172.
Bunzel E (1871) Die Reptilienfauna der Gosauformation in der Neuen Welt bei Wiener-Neustadt. Abhandlungen der kaiserlich-koeniglichen geologischen Reichsanstalt 5: 1–18.
Butler RJ, Benson RBJ, Barrett PM (2013) Pterosaur diversity: Untangling the influence of sampling biases, Lagerstätten, and genuine biodiversity signals. Palaeogeography, Palaeoclimatology, Palaeoecology 372: 78–87. doi:
10.1016/j.palaeo.2012.08.012Cai Z, Wei F (1994) On a new pterosaur (
Zhejiangopterus linhaiensis gen. et sp. nov.) from Upper Cretaceous in Linhai, Zhejiang, China. Vertebrata PalAsiatica 32: 181–194.
Caldwell MW (2005) The squamates: origins, phylogeny, and paleoecology. In: Currie PJ, Koppelhus EB (Eds) Dinosaur Provincial Park: A Spectacular Ancient Ecosystem Revealed. Indiana University Press, Bloomington, Indianapolis, 235–248.
Cappetta H (1987) Mesozoic and Cenozoic Elasmobranchii, Chondrichthyes II. Hanbook of Paleoichthyology Volume 3B. Gustav Fisher-Verlag, Stuttgart, 193 pp.
Cappetta H, Pfeil FH, Schmidt-Kittler N, Martini E (2000) New biostratigraphical data on the marine Upper Cretaceous and Palaeogene of Jordan. Newsletters on Stratigraphy 38: 81–95.
Carpenter K (1982) Baby dinosaurs from the Late Cretaceous Lance and Hell Creek Formations and a description of a new species of theropod. Contributions to Geology, University of Wyoming 20: 123–134.
Carr TD, Williamson TE (2010)
Bistahieversor sealeyi, gen. et sp. nov., a new tyrannosauroid from New Mexico and the origin of deep snouts in Tyrannosauroidea. Journal of Vertebrate Paleontology 30: 1–16. doi:
10.1080/02724630903413032Case GR (1979) Cretaceous selachians from the Peedee Formation (late Maastrichtian) of Duplin County, North Carolina. Brimleyana 2: 77–89.
Case JA, Goin FJ, Woodburne JM (2005) "South American” marsupials from the Late Cretaceous of North America and the origin of marsupial cohorts. Journal of Mammalian Evolution 12: 461–494. doi:
10.1007/s10914-005-7329-3Cavin L, Brito PM (2001) A new Lepisosteidae (Actinopterygii, Ginglymodi) from the Cretaceous of the Kem Kem beds, Southern Morocco. Bulletin de la Société géologique de France 172: 141–150. doi:
10.2113/172.5.661Cavin L, Forey PL (2001) Osteology and systematic affinities of
Palaeonotopterus greenwoodi Forey 1997 (Teleostei: Osteoglossomorpha). Zoological Journal of the Linnean Society 133: 25–52.
Cavin L, Forey PL (2004) New mawsoniid coelacanth (Sarcopterygii, Actinistia) remains from the Cretaceous of the Kem Kem beds, SE Morocco. In: Tintori A, Arratia G (Eds) Mesozoic Fishes III - Systematics, Paleoenvironments and Biodiversity. Verlag Dr. Friedrich Pfeil, Munich 493–506.
Cavin L, Forey PL (2008) A new tselfatiiform teleost from the mid-Cretaceous (Cenomanian) of the Kem Kem beds, Southern Morocco. In: Arratia G, Schultze H-P, Wilson MVH (Eds) Mesozoic Fishes IV - Homology and Phylogeny. Verlag Dr. Friedrich Pfeil, Munich, 199–216.
Cavin L, Forey PL, Buffetaut E, Tong H (2005) Latest European coelacanth shows Gondwanan affinities. Biology Letters 1: 176–177. doi:
10.1098/rsbl.2004.0287Cavin L, Tong H, Boudad L, Meister C, Piuz A, Tabouelle J, Aarab M, Amiot R, Buffetaut E, Dyke GJ, Hua S, Le Loeuff J (2010) Vertebrate assemblages from the early Late Cretaceous of southeastern Morocco: an overview. Journal of African Earth Sciences 57: 391–412. doi:
10.1016/j.jafrearsci.2009.12.007Cazau LB, Uliana MA (1972) El Cretacico superior continental de la Cuenca Neuquina. Adas V Congreso Geoldgico Argentino, Buenos Aires. 131–163.
Chang M-M, Chen P, Wang Y, Wang Y, Miao D (2008) The Jehol Fossils. The Emergence of Feathered Dinosaurs, Beaked Birds and Flowering Plants. Academic Press, Beijing, 208 pp.
Chen P (2000) Paleoenvironmental changes during the Cretaceous in eastern China. In: Okada H, Mateer NJ (Eds) Cretaceous Environments of Asia. Elsevier, Amsterdam, 81–90.
Chen P, Chang Z (1994) Nonmarine Cretaceous stratigraphy of eastern China. Cretaceous Research 15: 245–257. doi:
10.1006/cres.1994.1015Chester SGB, Sargis EJ, Szalay FS, Archibald JD, Averianov AO (2010) Mammalian distal humeri from the Late Cretaceous of Uzbekistan. Acta Palaeontologica Polonica 55: 199–211. doi:
10.4202/app.2009.0023Chester SGB, Sargis EJ, Szalay FS, Archibald JD, Averianov AO (2012) Therian femora from the Late Cretaceous of Uzbekistan. Acta Palaeontologica Polonica 57: 53–64.
Chitoku T (1996) Pterosaur bone from the Upper Cretaceous of Enbetsu, Hokkaido. Bulletin of the Hobetsu Museum 12: 17–24.
Chkhikvadze VM (1976) [New data on fossil turtles of Mongolia, China and eastern Kazakhstan]. Soobshcheniya Akademii Nauk Gruzinskoi SSR 82: 745–748.
Christopher RA, Prowell DC (2002) A palynological biozonation for the Maastrichtian Stage (Upper Cretaceous) of South Carolina, USA. Cretaceous Research 23: 639–669. doi:
10.1006/cres.2002.1029Chure DJ, Manabe M, Tanimoto M, Tomida Y (1999) An unusual theropod tooth from the Mifune Group (late Cenomanian to early Turonian), Kumamoto, Japan. In: Tomida Y, Rich THV, Vickers-Rich P (Eds) Proceedings of the Second Gondwanan Dinosaur Symposium. National Science Museum Monographs 15, Tokyo, 291–296.
Clark JM, Perle A, Norell MA (1994) The skull of
Erlicosaurus andrewsi, a Late Cretaceous "segnosaur" (Theropoda: Therizinosauridae) from Mongolia. American Museum Novitates: 1–39.
Clemens WA (1963) Fossil mammals of the type Lance Formation, Wyoming. Part I. Introduction and Multituberculata. University of California Publications in Geological Sciences 48: 1–105.
Clemens WA (1966) Fossil mammals of the type Lance Formation Wyoming. Part II. Marsupialia. University of California Publications in Geological Sciences 62: 1–122.
Clemens WA (1973) Fossil mammals of the type Lance Formation, Wyoming. Part III. Eutheria and summary. University of California Publications in Geological Sciences 94: 1–102.
Collins JI (1970) The chelonian
Rhinochelys Seeley from the Upper Cretaceous of England and France. Palaeontology 13: 355–378.
Company J, Galobart À, Gaete R (1998) First data on the hadrosauroid dinosaurs (Ornithischia, Dinosauria) from the Upper Cretaceous of Valencia, Spain. Oryctos 10: 121–126.
Company J, Murelaga X, Pereda Suberbiola X, Ruiz-Omeñaca JI (1999a) The vertebrate fauna from the new Late Cretaceous Chera locality (Valencia Province, Spain). In: Canudo JI, Cuenca-Bescós G (Eds) IV European Workshop on Vertebrate Palaeontology Programme and Abstracts. Universidad de Zaragoza, Zaragoza, 47–48.
Company J, Pereda Suberbiola X, Ruiz-Omeñaca JI (2005) A new species of
Doratodon (Crocodyliformes: Ziphosuchia) from the Late Cretaceous of Spain. Journal of Vertebrate Paleontology 25: 343–353. doi:
10.1671/0272-4634(2005)025[0343:ANSODC]2.0.CO;2Company J, Ruiz-Omeñaca JI, Pereda Suberbiola X (1999b) A long-necked pterosaur (Pterodactyloidea, Azhdarchidae) from the Upper Cretaceous of Valencia, Spain. Geologie en Mijnbouw 78: 319–333. doi:
10.1023/A:1003851316054Company J, Szentesi Z (2012) Amphibians from the Late Cretaceous Sierra Perenchiza Formation of the Chera Basin, Valencia Province, Spain. Cretaceous Research 37: 240–245. doi:
10.1016/j.cretres.2012.04.003Company J, Szentesi Z, Makádi L (2009a) Amphibians and lizards from the Upper Cretaceous (late Campanian – early Maastrichtian) Sierra Perenchiza Formation (Valencia Province, Spain). In: Schwarz-Wings D, Wings O, Sattler F (Eds) 7th Annual Meeting of the European Association of Vertebrate Palaeontologists, Abstract Volume. Museum für Naturkunde, Berlin, 20.
Company J, Szentesi Z, Makádi L (2009b) Amphibians from the Latest Maastrichtian La Solana locality, Valencia Province, Spain. In: Schwarz-Wings D, Wings O, Sattler F (Eds) 7th Annual Meeting of the European Association of Vertebrate Palaeontologists Abstract Volume. Museum für Naturkunde, Berlin, 20.
Company J, Torices A, Pereda Suberbiola X, Ruiz-Omeñaca JI (2009c) Theropod teeth from the Late Cretaceous of Chera (Valencia, eastern Spain). Journal of Vertebrate Paleontology 29: 81A
Company J, Unwin DM, Ruiz-Omeñaca JI, Pereda Suberbiola X (2001) A giant azhdarchid pterosaur from the latest Cretaceous of Valencia, Spain - The largest flying creature ever? Journal of Vertebrate Paleontology 21: 41A-42A
Connor CW (1992) The Lance Formation-Petrography and stratigraphy, Powder River Basin and nearby basins, Wyoming and Montana. United States Geological Survey Bulletin 1917-I: 1–17.
Coria RA (2001) New theropod from the Late Cretaceous of Patagonia. In: Tanke DH, Carpenter K (Eds) Mesozoic Vertebrate Life. Indiana University Press, Bloomington, 3–9.
Coria RA, Riga BG, Casadio S (2012) Un nuevo hadrosaurido (Dinosauria, Ornithopoda) de la Formation Allen, Provincia De La Pampa, Argentina. Ameghiniana 49: 552–572. doi:
10.5710/AMGH.9.4.2012.487Csiki-Sava Z, Balc R, Brusatte S, Dyke GJ, Naish D, Norell MA, Vremir M (2012) Petreşti-Arini (Transylvanian Basin, Romania) - a very important but ephemeral Late Cretaceous (Early Maastrichtian) vertebrate site. In: Royo-Torres R, Gascó F, Alcalá L (Eds) 10th Annual Meeting of the European Association of Vertebrate Palaeontologists. ¡Fundamental!, Teurel, 53–55.
Csiki Z, Vremir M, Brusatte SL, Norell MA (2010) An aberrant island-dwelling theropod dinosaur from the Late Cretaceous of Romania. Proceedings of the National Academy of Sciences USA 107: 15357–15361. doi:
10.1073/pnas.1006970107Cuny G, Martin JE, Sarr R (2012) A neoselachian shark fauna from the Late Cretaceous of Senegal. Cretaceous Research 34: 107–115. doi:
10.1016/j.cretres.2011.10.008Currie PJ, Carabajal AP (2012) A new specimen of
Austroraptor cabazai Novas, Pol, Canale, Porfiri and Calvo, 2008 (Dinosauria, Theropoda, Unenlagiidae) from the latest Cretaceous (Maastrichtian) of Rio Negro, Argentina. Ameghiniana 49: 662–667. doi:
10.5710/AMGH.30.8.2012.574Currie PJ, Godfrey SJ, Nesov LA (1994) New caenagnathid (Dinosauria: Theropoda) specimens from the Upper Cretaceous of North America and Asia. Canadian Journal of Earth Sciences 30: 2255–2272. doi:
10.1139/e93-196Currie PJ, Jacobsen AR (1995) An azhdarchid pterosaur eaten by a velociraptorine theropod. Canadian Journal of Earth Sciences 32: 922–925. doi:
10.1139/e95-077Currie PJ, Koppelhus EB (2005) Dinosaur Provincial Park. A Spectacular Ancient Ecosystem Revealed. Indiana University Press, Bloomington, 648 pp.
Currie PJ, Padian K (1983) A new pterosaur record from the Judith River (Oldman) Formation of Alberta. Journal of Paleontology 57: 599–600.
Currie PJ, Russell DA (1982) A giant pterosaur (Reptilia: Archosauria) from the Judith River (Oldman) Formation, Alberta. Canadian Journal of Earth Sciences 19: 894–897. doi:
10.1139/e82-074Dal Sasso C, Maganuco S, Buffetaut E, Mendez MA (2005) New information on the skull of the enigmatic theropod
Spinosaurus, with remarks on its size and affinities. Journal of Vertebrate Paleontology 25: 888–896. doi:
10.1671/0272-4634(2005)025[0888:NIOTSO]2.0.CO;2Danilov IG (1999) A new linholmemydid genus (Testudines: Linholmemydidae) from the mid-Cretaceous of Uzbekistan. Russian Journal of Herpetology 6: 63–71.
Danilov IG (2007) New data on soft-shelled turtles (Trionychidae) from the Bissekty Formation (late Turonian) of Dzharakuduk, Uzbekistan. Journal of Vertebrate Paleontology 27: 66A.
Danilov IG, Parham JF (2005) A reassessment of the referral of an isolated skull from the Late Cretaceous of Uzbekistan to the stem-testudinoid turtle genus
Lindholmemys. Journal of Vertebrate Paleontology 25: 784–791. doi:
10.1671/0272-4634(2005)025[0784:AROTRO]2.0.CO;2Danilov IG, Sukhanov VB, Syromyatnikova EV (2011) New Asiatic materials on turtles of the family Adocidae with review of the adocid record in Asia. Proceedings of the Zoological Institute RAS 315: 101–132.
Danilov IG, Syromyatnikova EV (2008) New materials on turtles of the family Nanhsiungchelyidae from the Cretaceous of Uzbekistan and Mongolia, with a review of the nanhsiungchelyid record in Asia. Proceedings of the Zoological Institute of the Russian Academy of Sciences 312: 3–25.
Danilov IG, Syromyatnikova EV, Sukhanov VB (2007) [Turtles of the genus
Shachemys from the Upper Cretaceous of Asia]. In: Rozanov AY, Lopatin AV, Parkhaev PY (Eds) [Modern Paleontology: Classical and Newest Methods]. Paleontological Institute of the Russian Academy of Sciences, Moscow, 59–72.
Danilov IG, Vitek NS (2013) Soft-shelled turtles (Trionychidae) from the Bissekty Formation (Late Cretaceous: late Turonian) of Uzbekistan: Shell-based taxa. Cretaceous Research 41: 55–64. doi:
10.1016/j.cretres.2012.10.003Danilov IG, Vitek NS, Averianov AO, Glinskiy VN (in press) A new softshelled trionychid turtle of the genus
Khunnuchelys from the Upper Cretaceous Bostobe Formation of Kazakhstan. Acta Palaeontologica Polonica:
Davis KW (1974) Stratigraphy and depositional environments of the Glen Rose Formation, north central Texas. Baylor Geological Studies Bulletin 26: 1–24.
Davis SP, Martill DM (1999) The gonorynchiform fish
Dastilbe from the Lower Cretaceous of Brazil. Palaeontology 42: 715–740. doi:
10.1111/1475-4983.00094DeMar DG, Jr., Varricchio DD, Head JJ, Moore JR, Wilson GP (2012) A nearly complete fossil iguanian from the Upper Cretaceous (Campanian) Two Medicine Formation of western Montana. Society of Vertebrate Paleontology, 72nd Annual Meeting, 17–20 October, Raleigh, North Carolina, Program and Abstracts. 86.
Dietze K (2007) Redescription of
Dastilbe grandalli (Chanidae, Euteleostei) from the Early Cretaceous Crato Formation of North-Eastern Brazil. Journal of Vertebrate Paleontology 27: 8–16. doi:
10.1671/0272-4634(2007)27[8:RODCCE]2.0.CO;2Díez Díaz V, Tortosa T, Le Loeuff J (2013) Sauropod diversity in the Late Cretaceous of southwestern Europe: The lessons of odontology. Annales de Paleontologie 99: 119–129. doi:
10.1016/j.annpal.2012.12.002Dong Z-M, Sun Y-W, Wu S-Y (2003) On a new pterosaur from the Lower Cretaceous of Chaoyang Basin, Western Liaoning, China. Global Geology 22: 1–7.
Dong Z (1979) [Cretaceous dinosaurs of Hunan, China]. [Mesozoic and Cenozoic Red Beds of South China: Selected Papers from the “Cretaceous-Tertiary Workshop”]. Science Press, Nanxiong, 342–350.
Dorsey RJ, Lenegan RJ (2007) Structural controls on middle Cretaceous sedimentation in the Toney Butte area of the Mitchell inlier, Ochoco basin, central Oregon. Geological Society of America, Special Paper 419: 97–115.
Dutheil DB (1999) An overview of the freshwater fish fauna from the Kern Kem beds (Late Cretaceous: Cenomanian) of southeastern Morocco. In: Arratia G, Schultze H-P (Eds) Mesozoic Fishes 2: Systematics and Fossil Record Proceedings of the International Meeting, Buckow, 1997. Verlag Dr. Friedrich Pfeil, Muenchen, 553–563.
Dyke GJ, Benton MJ, Posmosanu E, Naish D (2011) Early Cretaceous (Berriasian) birds and pterosaurs from the Cornet bauxite mine, Romania. Palaeontology 54: 79–95. doi:
10.1111/j.1475-4983.2010.00997.xDyke GJ, Malakhov DV (2004) Abundance and taphonomy of dinosaur teeth and other vertebrate remains from the Bostobynskaya Formation, north-east Aral Sea region, Republic of Kazakhstan. Cretaceous Research 25: 669–674. doi:
10.1016/j.cretres.2004.06.004Dyke GJ, Ősi A (2010) A review of Late Cretaceous fossil birds from Hungary. Geological Journal 45: 434–444.
Eaton JG, Kirkland JI, Hutchison JH, Denton R, O'Neill RC, Parrish JM (1997) Nonmarine extinction across the Cenomanian-Turonian boundary, southwestern Utah, with a comparison to the Cretaceous-Tertiary extinction event. Geological Society of America Bulletin 109: 560–567. doi:
10.1130/0016-7606(1997)109<0560:NEATCT>2.3.CO;2Eberth DA (2005) The geology. In: Currie PJ, Koppelhus EB (Eds) Dinosaur Provincial Park: A Spectacular Ancient Ecosystem Revealed. Indiana University Press, Bloomington, 54–82.
Eberth DA, Brinkman DB (1997) Paleoecology of an estuarine, incised-valley fill in the Dinosaur Park Formation (Judith River Group, Upper Cretaceous) of Southern Alberta, Canada. Palaios 12: 43–58. doi:
10.2307/3515293Eberth DA, Delgado-de Jesús CR, Lerbekmo JF, Brinkman DB, Rodríguez-de la Rosa RA, Sampson SD (2004) Cerro del Pueblo Formation (Difunta Group, Upper Cretaceous), Parras Basin, southern Coahuila, Mexico: reference sections, age, and correlation. Revista Mexicana de Ciencias Geologicas 21: 335–352.
Eberth DA, Hamblin AP (1993) Tectonic, stratigraphic, and sedimentologic significance of a regional discontinuity in the upper Judith River Group (Belly River wedge) of southern Alberta, Saskatchewan, and northern Montana. Canadian Journal of Earth Sciences 30: 174–200. doi:
10.1139/e93-016Efimov MB (1983) [A review of fossil crocodiles of Mongolia]. Trudy Sovmestnoi Sovetsko-Mongol'skoi Paleontologicheskoi Ekspeditsii 24: 76–96.
Efimov MB (1988a) [Fossil crocodiles and champsosaurs of Mongolia and USSR]. Trudy Sovmestnoi Sovetsko-Mongol'skoi Paleontologicheskoi Ekspeditsii 36: 1–108.
Efimov MB (1988b) [On the fossil crocodiles of Mongolia and the USSR]. Trudy Sovmestnoi Sovetsko-Mongol'skoi Paleontologicheskoi Ekspeditsii 34: 81–90.
Ekrt B, Košt'ák M, Mazuch M, Voigt S, Wiese F (2008) New records of teleosts from the late Turonian (Late Cretaceous) of the Bohemian Cretaceous Basin (Czech Republic). Cretaceous Research 29: 659–673. doi:
10.1016/j.cretres.2008.01.013Elgin RA, Campos HBN (2012) A new specimen of the azhdarchoid pterosaur
Tapejara wellnhoferi. Historical Biology 24: 586–591. doi:
10.1080/08912963.2011.613467Elgin RA, Frey E (2011) A new azhdarchoid pterosaur from the Cenomanian (Late Cretaceous) of Lebanon. Swiss Journal of Geosciences 104: 21–33. doi:
10.1007/s00015-011-0081-1Elgin RA, Hone DWE, Frey E (2011) The extent of the pterosaur flight membrane. Acta Palaeontologica Polonica 56: 99–111. doi:
10.4202/app.2009.0145Elzanowski A, Galton PM (1991) Braincase of
Enaliornis, an Early Cretaceous bird from England. Journal of Vertebrate Paleontology 11: 90–107. doi:
10.1080/02724634.1991.10011377Elzanowski A, Stidham TA (2011) A galloanserine quadrate from the Late Cretaceous Lance Formation of Wyoming. Auk 128: 138–145. doi:
10.1525/auk.2011.10113Estes RD (1964) Fossil vertebrates from the Late Cretaceous Lance Formation, Eastern Wyoming. University of California Publications in Geological Sciences 49: 1–180.
Estes RD (1965) A new fossil salamander from Montana and Wyoming. Copeia: 90–95. doi:
10.2307/1441245Estes RD (1969) A new fossil discoglossid frog from Montana and Wyoming. Breviora 328: 1–7.
Estes RD, Berberian P, Meszoely CAM (1969) Lower vertebrates from the Late Cretaceous Hell Creek Formation, McCone County, Montana. Breviora: 1–32.
Estes RD, Sanchíz B (1982) New discoglossid and palaeobatrachid frogs from the Late Cretaceous of Wyoming and Montana, and a review of other frogs from the Lance and Hell Creek formations. Journal of Vertebrate Paleontology 2: 9–20. doi:
10.1080/02724634.1982.10011914Ettachfini EM, Andreu B (2004) Le Cénomanien et le Turonien de la Plate-forme Préafricaine du Maroc. Cretaceous Research 25: 277–302. doi:
10.1016/j.cretres.2004.01.001Evans DC, Bavington R, Campione NE (2009) An unusual hadrosaurid braincase from the Dinosaur Park Formation and the biostratigraphy of Parasaurolophus (Ornithischia: Lambeosaurinae) from southern Alberta. Canadian Journal of Earth Sciences 46: 791–800. doi:
10.1139/E09-050Evans DC, Reisz RR (2007) Anatomy and relationships of
Lambeosaurus magnicristatus, a crested hadrosaurid dinosaur (Ornithischia) from the Dinosaur Park Formation, Alberta. Journal of Vertebrate Paleontology 27: 373–393. doi:
10.1671/0272-4634(2007)27[373:AAROLM]2.0.CO;2Evans DC, Reisz RR, Dupuis K (2007) A juvenile
Parasaurolophus (Ornithischia: Hadrosauridae) braincase from Dinosaur Provincial Park, Alberta, with comments on crest ontogeny in the genus. Journal of Vertebrate Paleontology 27: 642–650. doi:
10.1671/0272-4634(2007)27[642:AJPOHB]2.0.CO;2Evans SE, Barrett PM, Ward DJ (2004) The first record of lizards and amphibians from the Wessex Formation (Lower Cretaceous: Barremian) of the Isle of Wight, England. Proceedings of the Geologists' Association 115: 239–247.
Evans SE, Yabumoto Y (1998) A lizard from Early Cretaceous Crato Formation, Araripe Basin, Brazil. Neues Jahrbuch für Geologie und Paläontologie, Monatshefte: 349–364.
Faupl P, Pober E, Wagreich M (1987) Facies development of the Gosau Group of the eastern parts of the Northern Calcareous Alps during the Cretaceous and the Paleogene. In: Flügel H-W, Faupl P (Eds) Geodynamics of the Eastern Alps. Wien, 142–155.
Fielding S, Martill DM, Naish D (2005) Solnhofen-style soft-tissue preservation in a new species of turtle from the Crato Formation (Early Cretaceous, Aptian) of North-East Brazil. Palaeontology 48: 1301–1310. doi:
10.1111/j.1475-4983.2005.00508.xFielitz C, Stewart JD, Wiffen J (1999) +
Aethocephalichthys hyainarhinos gen. et sp. nov., a new and enigmatic Late Cretaceous actinopterygian from North America and New Zealand. In: Arratia G, Schultze H-P (Eds) Mesozoic Fishes 2: Systematics and Fossil Record Proceedings of the International Meeting, Buckow, 1997. Verlag Dr. Friedrich Pfeil, Muenchen, 95–106.
Figueiredo RG, Kellner AWA (2009) A new crocodylomorph specimen from the Araripe Basin (Crato Member, Santana Formation), northeastern Brazil. Paläontologische Zeitschrift 83: 323–331. doi:
10.1007/s12542-009-0016-6Fischer V, Bardet N, Guiomar M, Godefroit P (2014) High diversity in Cretaceous ichthyosaurs from Europe prior to their extinction. PLoS One 9: e84709. doi:
10.1371/journal.pone.0084709Forey PL (1997) A Cretaceous notopterid (Pisces, Osteoglossomorpha) from Morocco. South African Journal of Science 93: 564–569.
Forey PL, Cavin L (2007) A new species of †
Cladocyclus (Teleostei: Ichthyodectiformes) from the Cenomanian of Morocco. Palaeontologia Electronica 10: 12A:10p, 702KB
Forey PL, Grande L (1998) An African twin to the Brazilian
Calamopleurus (Actinopterygii, Amiidae). Zoological Journal of the Linnean Society 123: 179–195. doi:
10.1111/j.1096-3642.1998.tb01299.xForey PL, López-Arbarello A, MacLeod N (2011) A new species of
Lepidotes (Actinopterygii: Semiontiformes) from the Cenomanian (Upper Cretaceous) of Morocco. Palaeontologia Electronica 14: 7A:12p.
Fortier DC, Schultz CL (2009) A new neosuchian crocodylomorph (Crocodyliformes, Mesoeucrocodylia) from the Early Cretaceous of north-east Brazil. Palaeontology 52: 991–1007. doi:
10.1111/j.1475-4983.2009.00894.xFox RC, Naylor BG (2006) Stagodontid marsupials from the Late Cretaceous of Canada and their systematic and functional implications. Acta Palaeontologica Polonica 51: 13–36.
Frey E, Buchy M-C, Martill DM (2003) Middle- and bottom-decker Cretaceous pterosaurs: unique designs in active flying vertebrates. In: Buffetaut E, Mazin J-M (Eds) Evolution and Palaeobiology of Pterosaurs. Geological Society Special Publication, n. 217, 267–274.
Frey E, Martill DM (1996) A reappraisal of
Arambourgiania (Pterosauria, Pterodactyloidea): One of the world's largest flying animals. Neues Jahrbuch für Geologie und Palaentologie Abhandlungen 199: 221–247.
Fritsch A (1881a) O zkamenĕlém ptáku Chocĕnském. Vesmír: 232–233.
Fritsch A (1881b) Ueber die Entdeckung von Vogelresten in der böhmischen Kreideformation. Sitzungsberichte der königlich-bömischen Gesellschaft der Wissenschaften in Prag: 275–276.
Fritsch A (1883) Studien im Gebiet der böhmisches Kreideformation. Paläontologische Untersuchung der einzelnen Schichten. III Die Iserschichten. Archiv der naturwissenschaftenlichen Landesdurchforschung von Böhmen, Geologische Abtheilung 5: 1–140.
Gaffney ES (1972) The systematics of the North American family Baenidae (Reptilia, Cryptodira). Bulletin of the American Museum of Natural History 147: 241–320.
Gaffney ES, Tong H, Meylan PA (2002)
Galianemys, a new side-necked turtle (Pelomedusoides: Bothremydidae) from the Late Cretaceous of Morocco. American Museum Novitates: 1–20.
Gaffney ES, Tong H, Meylan PA (2006) Evolution of the side-necked turtles, the families Bothremydidae, Euraxemydidae, and Araripemydidae. Bulletin of the American Museum of Natural History 300: 1–698. doi:
10.1206/0003-0090(2006)300[1:EOTSTT]2.0.CO;2Gaffney ES, Zangerl R (1968) A revision of the chelonian genus
Bothremys (Pleurodira: Pelomedusidae). Fieldiana: Geology 16: 193–239.
Galton PM (2009) Notes on Neocomian (Lower Cretaceous) ornithopod dinosaurs from England -
Hypsilophodon,
Valdosaurus, "
Camptosaurus", "
Iguanodon" - and referred specimens from Romania and elsewhere. Revue de Paléobiologie 28: 211–273.
Galton PM, Martin LD (2002)
Enaliornis, an Early Cretaceous hesperornithiform bird from England, with comments on other Hesperornithiformes. In: Chiappe LM, Witmer LM (Eds) Mesozoic Birds: Above the Heads of Dinosaurs. University of California Press, Berkeley, 121–125.
Gao K, Fox RC (1996) Taxonomy and evolution of Late Cretaceous lizards (Reptilia: Squamata) from Western Canada. Bulletin of the Carnegie Museum of Natural History 33: 1–107.
Gao K, Nesov LA (1998) Early Cretaceous squamates from the Kyzylkum Desert, Uzbekistan. Neues Jahrbuch für Geologie und Paläontologie Abhandlungen 207: 289–309.
Gardner JD (2000) Albanerpetontid amphibians from the Upper Cretaceous (Campanian and Maastrichtian) of North America. Geodiversitas 22: 349–388.
Gardner JD, Averianov AO (1998) Albanerpetontid amphibians from the Upper Cretaceous of Middle Asia. Acta Palaeontologica Polonica 43: 453–467.
Gardner JD, DeMar DG, Jr. (2013) Mesozoic and Palaeocene lissamphibian assemblages of North America: a comprehensive review. Palaeobiodiversity and Palaeoenvironments 93: 459–515. doi:
10.1007/s12549-013-0130-zGasparini ZB, de, Salgado L, Casadio S (2003) Maastrichtian plesiosaurs from northern Patagonia. Cretaceous Research 24: 157–170. doi:
10.1016/S0195-6671(03)00036-3Gates TA, Sampson SD, Delgado-de Jesús CR, Zanno LE, Eberth DA, Hernandez-Rivera R, Aguillón Martínez M, Kirkland JI (2007)
Velafrons coahuilensis, a new lambeosaurine hadrosaurid (Dinosauria: Ornithopoda) from the late Campanian Cerro del Pueblo Formation, Coahuila, Mexico. Journal of Vertebrate Paleontology 27: 917–930. doi:
10.1671/0272-4634(2007)27[917:VCANLH]2.0.CO;2Geist NR, Hillenius WJ, Frey E, Jones TD, Elgin RA (in press) Breathing in a box: Constraints on lung ventilation in giant pterosaurs. Anatomical Record
Gheerbrant E, Astibia H (1994) Un nouveau mammifère du Maastrichtien de Laño (Pays Basque espagnol). Comptes rendus de l'Académie des sciences Série 2 Sciences de la terre et des planètes 318: 1125–1131.
Gheerbrant E, Astibia H (2012) Addition to the Late Cretaceous Laño mammal faunule (Spain) and to the knowledge of European ‘Zhelestidae’ (Lainodontinae nov.). Bulletin de la Société géologique de France 183: 537–546. doi:
10.2113/gssgfbull.183.6.537Gilmore CW (1928) A new pterosaurian reptile from the marine Cretaceous of Oregon. Proceedings of the United States National Museum 73: 1–5. doi:
10.5479/si.00963801.73-2728.1Glazunova AE (1972) [Paleontological Justification of Stratigraphic Patrition of the Cretaceous Deposits of Volga Region. Upper Cretaceous]. Nedra, Moscow, 204 pp.
Glickman LS, Mertinene RA, Nesov LA, Rozhdestvensky AK, Khosatzky LI, Yakovlev VN (1987) [Vertebrates]. In: Moskvin MM (Ed.) [Stratigraphy of the USSR Cretaceous System Semivolume 2]. 255–262.
Gmira S (1995) Étude des chéloniens fossiles du Maroc (antomie - systématique - phylogénie). CNRS, Paris, 140 pp.
Godefroit P, Alifanov VR, Bolotsky YL (2004) A re-appraisal of
Aralosaurus tuberiferus (Dinosauria, Hadrosauria) from the Late Cretaceous of Kazakhstan. Bulletin de l'Institut Royal des Sciences Naturelles de Belgique, Sciences de la Terre 74, supplement: 139–154.
Godefroit P, Escuillié F, Bolotsky YL, Lauters P (2012) A new basal hadrosauroid dinosaur from the Upper Cretaceous of Kazakhstan. In: Godefroit P (Ed) Bernissart Dinosaurs and Early Cretaceous Terrestrial Ecosystems. Indiana University Press, Bloomington, 335–358.
Godfrey SJ, Currie PJ (2005) Pterosaurs. In: Currie PJ, Koppelhus EB (Eds) Dinosaur Provincial Park: A Spectacular Ancient Ecosystem Revealed. Indiana University Press, Bloomington, 292–311.
Gómez RO, Báez AM, Rougier GW (2008) An anilioid snake from the Upper Cretaceous of northern Patagonia. Cretaceous Research 29: 481–488. doi:
10.1016/j.cretres.2008.01.002González Riga BJ, Calvo JO, Porfiri JD (2008) An articulated titanosaur from Patagonia (Argentina): New evidence of neosauropod pedal evolution. Palaeoworld 17: 33–40. doi:
10.1016/j.palwor.2007.08.003Gradstein FM, Ogg JG, Smith Al (2004) A Geologic Time Scale 2004. Cambridge University Press, Cambridge, New York, Melbourne, 589 pp.
Grigorescu D (1992) Nonmarine Cretaceous Formations of Romania. In: Mateer NJ, Chen P (Eds) Aspects of nonmarine Cretaceous geology. China Ocean Press, Beijing, 142–164.
Grigorescu D, Seclamen M, Norman DB, Weishampel DB (1990) Dinosaur eggs from Romania. Nature 346: 417. doi:
10.1038/346417a0Grigorescu D, Venczel M, Csiki Z, Limberea R (1999) New latest Cretaceous microvertebrate fossil assemblages from the Hateg Basin (Romania). Geologie en Mijnbouw 78: 301–314. doi:
10.1023/A:1003890913328Grigorescu D, Weishampel DB, Norman DB, Seclamen M, Rusu M, Baltres A, Teodorescu V (1994) Late Maastrichtian dinosaur eggs from the Haţeg Basin (Romania). In: Carpenter K, Hirsch KF, Horner JR (Eds) Dinosaur Eggs and Babies. Cambridge University Press, Cambridge, 75–87.
Habib M (2007) Structural characteristics of the humerus of
Bennettazhia oregonensis and their implications for specimen diagnosis and azhdarchoid biomechanics. In: Hone D (Ed.) Flugsaurier The Wellnhofer Pterosaur Meeting. Bavarian State Collection for Palaeontology, Munich, 16.
Harries PJ, Little CTS (1999) The early Toarcian (Early Jurassic) and the Cenomanian–Turonian (Late Cretaceous) mass extinctions: similarities and contrasts. Palaeogeography, Palaeoclimatology, Palaeoecology 154.
Harris WB, Self-Trail JM (2006) Late Cretaceous base level lowering in Campanian and Maastrichtian depositional sequences, Kure Beach, North Carolina. Stratigraphy 3: 195–216. doi:
10.1671/0272-4634(2006)26[192:AAPCVF]2.0.CO;2Henderson MD, Peterson JE (2006) An azhdarchid pterosaur cervical vertebra from the Hell Creek Formation (Maastrichtian) of southeastern Montana. Journal of Vertebrate Paleontology 26: 192–195.
Henderson RA, McNamara KJ (1985) Maastrichtian non-heteromorph ammonites from the Miria Formation, Western Australia. Palaeontology 28: 35–88.
Herman AB, Kvaček J (2007) Early Campanian Grünbach flora of Austria: Systematic composition and paleoclimatic interpretations. Acta Paleobotanica 47: 37–55.
Hicks JF, Brinkman DL, Nichols DL, Watabe M (1999) Paleomagnetic and palynologic analyses of Albian to Santonian strata at Bayn Shireh, Burkhant, and Khuren Dukh, eastern Gobi Desert, Mongolia. Cretaceous Research 20: 829–850. doi:
10.1006/cres.1999.0188Hilton EJ, Grande L (2006) Review of the fossil record of sturgeons, family Acipenseridae (Actinopterygii: Acipenseriformes), from North America. Journal of Paleontology 80: 672–683. doi:
10.1666/0022-3360(2006)80[672:ROTFRO]2.0.CO;2Hirayama R (1998) [Fossil turtles from the Mifune Group (Late Cretaceous) of Kumamoto Prefecture, Western Japan]. Report of the research on the distribution of important fossils in Kumamoto Prefecture, "Dinosaurs from the Mufune Group, Kumamoto Prefecture, Japan". 85–99.
Hirayama R, Chitoku T (1996) Family Dermochelyidae (Superfamily Chelonioidea) from the Upper Cretaceous of North Japan. Transactions and Proceedings of the Palaeontological Society of Japan, New Series: 597–622.
Holroyd PA, Hutchison JH (2002) Patterns of geographic variation in latest Cretaceous vertebrates: evidence from the turtle component. In: Hartman JH, Johnson KR, Nichols DJ (Eds) The Hell Creek Formation and the Cretaceous-Tertiary Boundary in the Northern Great Plains, an Integrated Continental Record of the End of the Cretaceous. Geological Society of America Special Paper, 177–190.
Hone D, Tsuihiji T, Watabe M, Tsogtbaatr K (2012) Pterosaurs as a food source for small dromaeosaurs. Palaeogeography, Palaeoclimatology, Palaeoecology 331–332: 27–30. doi:
10.1016/j.palaeo.2012.02.021Hooley RW (1914) On the ornithosaurian genus
Ornithocheirus, with a review of the specimens of the Cambridge Greensand in the Sedgwick Museum. Annals and Magazine of Natural History, Series 8 13: 529–557.
Hope S (2002) The Mesozoic radiation of Neornithes. In: Chiappe LM, Witmer LM (Eds) Mesozoic Birds: Above the Heads of Dinosaurs. University of California Press, Berkeley, 339–388.
Horner JR (1982) Evidence for colonial nesting and "site fidelity" among ornithischian dinosaurs. Nature 297: 675–676. doi:
10.1038/297675a0Horner JR, Makela R (1979) Nest of juveniles provides evidence of family structure among dinosaurs. Nature 282: 296–298. doi:
10.1038/282296a0Horner JR, Varricchio DJ, Goodwin MB (1992) Marine transgressions and the evolution of Cretaceous dinosaurs. Nature 358: 59–61. doi:
10.1038/358059a0Howse SCB (1986) On the cervical vertebrae of the Pterodactyloidea (Reptilia: Archosauria). Zoological Journal of the Linnean Society 88: 307–328. doi:
10.1111/j.1096-3642.1986.tb02249.xHowse SCB, Milner AR (1995) The pterodactyloids from the Purbeck Limestone Formation of Dorset. Bulletin of the Natural History Museum(Geology) 51: 73–88.
Hunt RK, Lehman TM (2008) Attributes of the ceratopsian dinosaur
Torosaurus, and new material from the Javelina Formation (Maastrichtian) of Texas. Journal of Paleontology 82: 1127–1138. doi:
10.1666/06-107.1Hutt S, Naish D, Martill DM, Barker MJ, Newbery P (2001) A preliminary account of a new tyrannosauroid theropod from the Wessex Formation (Early Cretaceous) of southern England. Cretaceous Research 22: 227–242. doi:
10.1006/cres.2001.0252Hwang KG, Huh M, Lockley MG, Unwin DM, Wright JL (2002) New pterosaur tracks (Pteraichnidae) from the Late Cretaceous Uhangri Formation, southwestern Korea. Geological Magazine 139: 421–435. doi:
10.1017/S0016756802006647Ibrahim N, Unwin DM, Martill DM, Baidder L, Zouhri S (2010) A new pterosaur (Pterodactyloidea: Azhdarchidae) from the Upper Cretaceous of Morocco. PLoS One 5: e10875. doi:
10.1371/journal.pone.0010875ICZN (1999) International Code of Zoological Nomenclature, Fourth Edition. International Trust for Zoological Nomenclature, c/o The Natural History Museum, London, 306 pp.
Ikegami N (1997) An azhdarchid pterosaur from the Mifune Group, Kumamoto Prefecture, Japan. Abstracts of the 104th Annual Meeting of the Geological Society of Japan: 350.
Ikegami N, Kellner AWA, Tomida Y (2000) The presence of an azhdarchid pterosaur in the Cretaceous of Japan. Paleontological Research 4: 165–170.
Ikegami N, Tamura M (1996) New dinosaurs and a pterosaur from the Mifune Group. Proceedings of the Nishinihon Branch Geological Society of Japan: 9–10.
Ikegami N, Tomida Y (2003) Early Late Cretaceous vertebrate fauna of the Mifune Group in Kumamoto Prefecture, Japan. Journal of Vertebrate Paleontology 23: 64A-65A
Insole AN, Hutt S (1994) The palaeoecology of the dinosaurs of the Wessex Formation (Wealden Group, Early Cretaceous), Isle of Wight, Southern England. Zoological Journal of the Linnean Society 112: 197–215. doi:
10.1111/j.1096-3642.1994.tb00318.xIsaac MJ, Moore PR, Joass YJ (1991) Tahora Formation: The basal facies of a Late Cretaceous transgressive sequence, northeastern New Zealand. New Zealand Journal of Geology and Geophysics 34: 227–236. doi:
10.1080/00288306.1991.9514460Jacobs LL (1995) Lone Star Dinosaurs. Texas A&M University Press, College Station, 160 pp.
Jacobs LL, Winkler DA (1998) Mammals, archosaurs, and the Early to Late Cretaceous transition in north-central Texas. In: Tomida Y, Flynn LJ, Jacobs LL (Eds) Advances in Vertebrate Paleontology and Geochronology. National Science Museum Monographs 14, Tokyo, 253–280.
Jianu C-M, Weishampel DB, Stiuca E (1997) Old and new pterosaur material from the Hateg basin (Late Cretaceous) of western Romania, and comments about pterosaur deversity in the Late Cretaceous of Europe. Abstracts of the Second European Workshop on Vertebrate Palaeontology. One page not paginated.
Jin X, Azuma Y, Jackson FD, Varricchio DD (2007) Giant dinosaur eggs from the Tiantai basin, Zhejiang Province, China. Canadian Journal of Earth Sciences 44: 81–88. doi:
10.1139/e06-077Juárez Valieri RD, Haro JA, Fiorelli LE, Calvo JO (2010) A new hadrosauroid (Dinosauria: Ornithopoda) from the Allen Formation (Late Cretaceous) of Patagonia, Argentina. Revista del Museo Argentino de Ciencias Naturales 12: 217–231.
Kauffman EG (1995) Global change leading to biodiversity crisis in a greenhouse world: the Cenomanian–Turonian (Cretaceous) mass extinction. In: Stanley SM, Usselmann T (Eds) The Effects of Past Global Change on Life Studies in Geophysics. National Academy Press, Washington, 47–71.
Kear BP, Ekrt B, Prokop J, Georgalis GL (2014) Turonian marine amniotes from the Bohemian Cretaceous Basin, Czech Republic. Geological Magazine 151: 183–198. doi:
10.1017/S0016756813000502Kear BP, Long JA, Martin JE (2005) A review of Australian mosasaur occurrences. Netherlands Journal of Geosciences 84: 307–313.
Kellner AWA (1995) The relationships of the Tapejaridae (Pterodactyloidea) with comments on pterosaur phylogeny. In: Sun A, Wang Y (Eds) Sixth Symposium on Mesozoic Terrestrial Ecosystems and Biota Short Papers. China Ocean Press, Beijing, 73–77.
Kellner AWA (2002) A review of Avian Mesozoic fossil feathers. In: Chiappe LM, Witmer LM (Eds) Mesozoic Birds, Above the Heads of Dinosaurs. University of California Press, 389–404.
Kellner AWA (2003) Pterosaur phylogeny and comments on the evolutionary history of the group. In: Buffetaut E, Mazin J-M (Eds) Evolution and Palaeobiology of Pterosaurs. Geological Society Special Publication, n. 217, 105–137.
Kellner AWA (2004) New information on the Tapejaridae (Pterosauria, Pterodactyloidea) and discussion of the relationships of this clade. Ameghiniana 41: 521–534.
Kellner AWA (2010) Comments on the Pteranodontidae (Pterosauria, Pterodactyloidea) with the description of two new species. Anais da Academia Brasileira de Ciências 82: 1063–1084. doi:
10.1590/S0001-37652010000400025Kellner AWA, Calvo JO, Sayão JM, Porfiri JD (2006) Pterosaur bones from the Portezuelo Formation (Cretaceous), Neuquén Group, Patagonia, Argentinia. Arquivos do Museu Nacional, Rio de Janeiro 64: 369–375.
Kellner AWA, Campos DdA (1994) A new species of
Tupuxuara (Pterosauria, Tapejaridae) from the Early Cretaceous of Brazil. Anais da Academia Brasileira de Ciências 66: 467–473.
Kellner AWA, Campos DdA (2002) The function of the cranial crest and jaws of a unique pterosaur from the Early Cretaceous of Brazil. Science 297: 389–392. doi:
10.1126/science.1073186Kellner AWA, Campos DdA (2007) Short note on the ingroup relationships of the Tapejaridae (Pterosauria, Pterodactyloidea). Boletim do Museu Nacional, Nova Série 75: 1–14.
Kellner AWA, Hasegawa Y (1993) Postcranial skeleton of
Tupuxuara (Pterosauria, Pterodactyloidea, Tapejaridae) from the Lower Cretaceous of Brazil. Journal of Vertebrate Paleontology 13: 44A.
Kellner AWA, Langston W Jr. (1994) The cranial anatomy of an azhdarchid pterosaur from the Late Cretaceous of Texas. Journal of Vertebrate Paleontology 14: 32A
Kellner AWA, Langston W Jr. (1996) Cranial remains of
Quetzalcoatlus (Pterosauria, Azhdarchidae) from Late Cretaceous sediments of Big Bend National Park, Texas. Journal of Vertebrate Paleontology 16: 222–231. doi:
10.1080/02724634.1996.10011310Kellner AWA, Mader BJ (1996) First record of Pterosauria (Pterodactyloidea, Azhdarchidae) from Cretaceous rocks of Morocco. Journal of Vertebrate Paleontology 16: 45A
Kellner AWA, Mello AMS, Ford TL (2007) A survey of pterosaurs from Africa with the description of a new specimen from Morocco. In: Carvalho IdS, Cassab RdCT, Schwanke C, Carvalho MdA, Fernandes ACS, Rodrigues MAdC, Carvalho MSSd, Arai M, Oliveria MEQ (Eds) Paleontologia: Cenários da Vida, Vol 1. Interciência, 257–268.
Kennedy WJ, Cobban WA (1993) Lower Campanian (Upper Cretaceous) ammonites from the Merchantville Formation of New Jersey, Maryland, and Delaware. Journal of Paleontology 67: 828–849.
Keyes IW (1977) Records of the northern hemisphere Cretaceous sawfish genus Onchopristis (order Batoidea) from New Zealand. New Zealand Journal of Geology and Geophysics 20: 263–272. doi:
10.1080/00288306.1977.10420706Khozatsky LI, Yur'ev KB (1964) [Superoder Pterosauria. Flying reptiles]. In: Rozhdestvensky AK, Tatarinov LP (Eds) [Fundamentals of Paleontology Amphibians, Reptiles and Birds]. Nauka, Moscow, 589–603.
Kielan-Jaworowska Z, Cifelli RL, Luo Z-X (2004) Mammals from the Age of Dinosaurs: Origins, Evolution, and Structure. Columbia University Press, New York, 630 pp.
Kielan-Jaworowska Z, Nesov LA (1990) On the metatherian nature of the Deltatheroida, a sister group of the Marsupialia. Lethaia 23: 1–10. doi:
10.1111/j.1502-3931.1990.tb01776.xKielan-Jaworowska Z, Nesov LA (1992) Multituberculate mammals from the Cretaceous of Uzbekistan. Acta Palaeontologica Polonica 37: 1–17.
Kim C-B, Huh M, Cheong C-S, Lockley MG, Chang H-W (2003) Age of the pterosaur and web-footed bird tracks associated with dinosaur footprints from South Korea. The Island Arc 12: 125–131. doi:
10.1046/j.1440-1738.2003.00385.xKim JY, Kim KS, Lockley MG, Yang SY, Seo SJ, Choi HI, Lim J-D (2008) New didactyl dinosaur footprints (
Dromaeosauripus hamanensis ichnogen. et ichnosp. nov.) from the Early Cretaceous Haman Formation, south coast of Korea. Palaeogeography, Palaeoclimatology, Palaeoecology 220: 72–78. doi:
10.1016/j.palaeo.2008.02.003Kim JY, Kim SH, Kim KS, Lockley MG (2006) The oldest record of webbed bird and pterosaur tracks from South Korea (Cretaceous Haman Formation, Changseon and Sinsu Islands): More evidence of high avian diversity in East Asia. Cretaceous Research 27: 56–69. doi:
10.1016/j.cretres.2005.10.005Kim JY, Lockley MG, Kim KS, Seo SJ, Lim J-D (2012) Enigmatic giant pterosaur tracks and associated ichnofauna from the Cretaceous of Korea: Implication for the bipedal locomotion of pterosaurs. Ichnos 19: 50–65. doi:
10.1080/10420940.2011.625779Kitamura N (2013) “
Carcharias”
amonensis (Chondrichthyes, Odontaspididae) from the Upper Cretaceous Mifune Group in Kumamoto, Japan. Paleontological Research 17: 230–235. doi:
10.2517/1342-8144-17.3.230Kocsis L, Ősi A, Vennemann T, Trueman CN, Palmer MR (2009) Geochemical study of vertebrate fossils from the Upper Cretaceous (Santonian) Csehbánya Formation (Hungary): Evidence for a freshwater habitat of mosasaurs and pycnodont fish. Palaeogeography, Palaeoclimatology, Palaeoecology 280: 532–542. doi:
10.1016/j.palaeo.2009.07.009Kolodny Y (1996) Dinosaur bones: fossils or pseudomorphs ? The pitfalls of physiology reconstruction from apatitic fossils. Palaeogeography, Palaeoclimatology, Palaeoecology 126: 161–171. doi:
10.1016/S0031-0182(96)00112-5Kordikova EG, Polly DP, Alifanov VR, Rocek Z, Gunnell GF, Averianov AO (2001) Small vertebrates from the Late Cretaceous and Early Tertiary of the northeastern Aral Sea Region, Kazakhstan. Journal of Paleontology 75: 390–400. doi:
10.1666/0022-3360(2001)075<0390:SVFTLC>2.0.CO;2Krause DW (1992)
Clemensodon megaloba, a new genus and species of Multituberculata (Mammalia) from the Upper Cretaceous Type Lance Formation, Powder River Basin, Wyoming. PaleoBios 14: 1–8.
Kurzanov SM (1976) [Structure of the braincase of carnosaur
Itemirus gen. nov. and some questions of dinosaur cranial anatomy]. Paleontologicheskii Zhurnal: 127–137.
Kuznetsov VV (1976) [A fresh-water turtle from the Senonian deposits of North-East Aral Sea area]. Paleontologicheskii Zhurnal 4: 125–127.
Kuznetsov VV, Chkhikvadze VM (1987) [The Late Cretaceous trionychids from Shakh-Shakh locality in Kazakhstan]. Materialy po Istorii Fauny i Flory Kazakhstana 9: 33–39.
Kuznetsov VV, Shilin PV (1983) [Late Cretaceous turtle from Baibishe (North-East Aral Sea region)]. Izvestiya Akademii Nauk Kazakhskoi SSR, Seriya biologicheskaya 6: 41–44.
Kvaček J, Herman A (2004) The Campanian Grünbach flora of Lower Austria: Paleoecological interpretations. Annalen des Naturhistorischen Museums in Wien 106A: 91–101.
Langston W Jr. (1974) Nonmammalian Comanchean tetrapods. Geoscience and Man 8: 77–102.
Lappartient JR, Monteillet J (1980) Le gisement fossilifère sénonien supérieur des carrières de Paki (Sénégal). Bulletin de l'Institut fondamental d'Afrique noire Série A: Sciences naturelles 42: 431–439. doi:
10.1371/journal.pone.0054329Larson DW, Currie PJ (2013) Multivariate analyses of small theropod dinosaur teeth and implications for paleoecological turnover through time. PLoS One 8: e54329.
Larsson HCE, Sidor CA (1999) Unusual crocodyliform teeth from the Late Cretaceous (Cenomanian) of southeastern Morocco. Journal of Vertebrate Paleontology 19: 398–401. doi:
10.1080/02724634.1999.10011152Larsson HCE, Sues H-D (2007) Cranial osteology and phylogenetic relationships of
Hamadasuchus rebouli (Crocodyliformes, Mesoeucrocodylia) from the Cretaceous of Morocco. Zoological Journal of the Linnean Society 149: 533–567. doi:
10.1111/j.1096-3642.2007.00271.xLauginiger EM (1984) An Upper Campanian vertebrate fauna from the Chesapeake and Delaware canal, Delaware. The Mosasaur 2: 141–149.
Lauginiger EM (1988) Cretaceous fossils from the Chesapeake and Delaware Canal: a guide for students and collectors. Delaware Geological Society Special Publication 18: 1–57.
Lavocat R (1949) Les gisements de vertébrés crétacés du sudMarocain. Comptes Rendus de l'Academie des Sciences 7: 125–126.
Lavocat R (1954) Sur les dinosauriens du Continental Intercalaire des Kem-Kem de la Daoura. Comptes Rendus 19th International Geological Congress, 1952, 1. 65–68.
Lawson DA (1975b) A pterosaur from the latest Cretaceous of West Texas: discovery of the largest flying creature. Science 187: 947–948. doi:
10.1126/science.187.4180.947Lawson DA (1976) Tyrannosaurus and Torosaurus, Maestrichtian dinosaurs from Trans-Pecos, Texas. Journal of Paleontology 50: 158–164.
Le Loeuff J, Buffetaut E (1998) A new dromaeosaurid theropod from the Upper Cretaceous of southern France. Oryctos 1: 105–112.
Le Loeuff J, Buffetaut E, Martin M (1994) The last stages of dinosaur faunal history in Europe: a succession of Maastrichtian dinosaur assemblages from the Corbières (southern France). Geological Magazine 131: 625–630. doi:
10.1017/S0016756800012413Leal MEdC, Brito PM (2004) The ichthyodectiform
Cladocyclus gardneri (Actinopterygii: Teleostei) from the Crato and Santana Formations, Lower Cretaceous of Araripe Basin, North-Eastern Brazil. Annales de Paleontologie 90: 103–113. doi:
10.1016/j.annpal.2004.01.001Leanza HA, Apesteguia S, Novas FE, Fuente MS, de la (2004) Cretaceous terrestrial beds from the Neuquen Basin (Argentina) and their tetrapod assemblages. Cretaceous Research 25: 61–87. doi:
10.1016/j.cretres.2003.10.005Lee Y-N (1994) The Early Cretaceous pterodactyloid pterosaur
Coloborhynchus from North America. Palaeontology 37: 755–763.
Lehman TM (1987) Late Maastrichtian paleoenvironments and dinosaur biogeography in the Western Interior of North America. Palaeogeography, Palaeoclimatology, Palaeoecology 60: 189–217. doi:
10.1016/0031-0182(87)90032-0Lehman TM (2001) Late Cretaceous dinosaur provinciality. In: Tanke DH, Carpenter K (Eds) Mesozoic Vertebrate Life. Indiana University Press, Bloomingtion, 310–328.
Lehman TM, Coulson AB (2002) A juvenile specimen of the sauropod dinosaur
Alamosaurus sanjuanensis from the Upper Cretaceous of Big Bend National Park, Texas. Journal of Paleontology 76: 156–172. doi:
10.1666/0022-3360(2002)076<0156:AJSOTS>2.0.CO;2Lehman TM, Langston W Jr. (1996) Habitat and behaviour of
Quetzalcoatlus: paleoenvironmental reconstruction of the Javelina Formation (Upper Cretaceous), Big Bend National Park, Texas. Journal of Vertebrate Paleontology 16: 48A
Lehman TM, McDowell FW, Connelly JN (2006) First isotopic (U-Pb) age for the Late Cretaceous Alamosaurus vertebrate fauna of West Texas, and its significance as a link between two faunal provinces. Journal of Vertebrate Paleontology 26: 922–928. doi:
10.1671/0272-4634(2006)26[922:FIUAFT]2.0.CO;2Lewy ZA, Milner AC, Patterson C (1992) Remarkably preserved natural endocranial casts of pterosaur and fish from the Late Cretaceous of Israel. Geological Survey of Israel, Current Research 7: 31–35.
Lobacheva SV (1979) [Beleuty Formation]. In: Vereshchagin VN (Ed.) [Stratigraphic Dictionary of the USSR: Triassic, Jurassic, and Cretaceous]. Nedra, Leningrad, 58.
Lockley MG, Huh M, Lim S-K, Yang SY, Chun SS, Unwin DM (1997) First report of pterosaur tracks from Asia, Chollanam Province Korea. Journal of the Paleontological Society of Korea Special Publication no. 2: 17–32.
Loewen MA, Sampson SD, Lund EK, Farke AA, Aguillón-Martínez MC, De Leon CA, Rodríguez de la Rosa RA, Getty MA, Eberth DA (2010) Horned dinosaurs (Ornithischia: Ceratopsidae) from the Upper Cretaceous (Campanian) Cerro del Pueblo Formation, Coahuila, Mexico. In: Ryan MJ, Chinnery-Allgeier BJ, Eberth DA (Eds) New perspectives on horned dinosaurs. Indiana University Press, Bloomington, 99–116.
Long JA (1992) First dinosaur bones from Western Australia. Records of the Northern Territory Museum of Arts and Sciences 9: 21–28.
Longrich N (2006) An ornithurine bird from the Late Cretaceous of Alberta, Canada. Canadian Journal of Earth Sciences 43: 1–7. doi:
10.1139/e05-091Longrich NR (2008) A new, large ornithomimid from the Cretaceous Dinosaur Park Formation of Alberta, Canada: Implications for the study of dissociated dinosaur remains. Palaeontology 51: 983–997. doi:
10.1111/j.1475-4983.2008.00791.xLongrich NR (2009) An ornithurine-dominated avifauna from the Belly River Group (Campanian, Upper Cretaceous) of Alberta, Canada. Cretaceous Research 30: 161–177. doi:
10.1016/j.cretres.2008.06.007Longrich NR, Bhullar B-AS, Gauthier JA (2012a) Mass extinction of lizards and snakes at the Cretaceous–Paleogene boundary. Proceedings of the National Academy of Sciences USA 109: 21396–21401. doi:
10.1073/pnas.1211526110Longrich NR, Bhullar B-AS, Gauthier JA (2012b) A transitional snake from the Late Cretaceous period of North America. Nature 488: 205–208. doi:
10.1038/nature11227Longrich NR, Currie PJ (2009) A microraptorine (Dinosauria–Dromaeosauridae) from the Late Cretaceous of North America. Proceedings of the National Academy of Sciences USA 106: 5002–5007. doi:
10.1073/pnas.0811664106López-Martinez N, Canudo JI, Ardèvol Ls, Pereda Suberbiola X, Orue-Etxebarria X, Cuenca-Bescós G, Ruiz-Omeñaca JI, Murelaga X, Feist M (2001) New dinosaur sites correlated with Upper Maastrichtian pelagic deposits in the Spanish Pyrenees: implications for the dinosaur extinction pattern in Europe. Cretaceous Research 22: 41–61. doi:
10.1006/cres.2000.0236Lu C, Gu M, Yu Y, Liang H (2006) Age of the Tangshang Formation in different basins of eastern Zhejiang. Journal of Stratigraphy 30: 81–86.
Lü J, Jin X, Unwin DM, Zhao L, Azuma Y, Ji Q (2006) A new species of
Huaxiapterus (Pterosauria: Pterodactyloidea) from the Lower Cretaceous of Western Liaoning, China with comments on the systematics of tapejarid pterosaurs. Acta Geologica Sinica 80: 315–326.
Lü J, Unwin DM, Xu L, Zhang X (2008) A new azhdarchoid pterosaur from the Lower Cretaceous of China and its implications for pterosaur phylogeny and evolution. Naturwissenschaften 95: 891–897. doi:
10.1007/s00114-008-0397-5Lucas SG, Sullivan RM (2000) Stratigraphy and vertebrate biostratigraphy across the Cretaceous-Tertiary boundary, Betonnie Tsosie Wash, San Juan Basin, New Mexico. New Mexico Museum of Natural History and Science Bulletin 17: 95–104.
Lydekker R (1888) Catalogue of the Fossil Reptilia and Amphibia in the British Museum (Natural History). Part I. Ornithosauria, Crocodilia, Dinosauria, Squamata, Rhynchocephalia, and Proterosauria. Taylor and Francis, London, 309 pp.
Lyson TR, Joyce WG (2009) A revision of
Plesiobaena (Testudines: Baenidae) and an assessment of baenid ecology across the K/T boundary. Journal of Paleontology 83: 833–853. doi:
10.1666/09-035.1Mader BJ, Kellner AWA (1999) A new anhanguerid pterosaur from the Cretaceous of Morocco. Boletim do Museu Nacional, Nova Serie: Geologia: 1–11.
Maisey JG (1991) Santana Fossils: An Illustrated Guide. T.F.H. Publications, 459 pp.
Makádi L (2006)
Bicuspidon aff.
hatzegiensis (Squamata: Scincomorpha: Teiidae) from the Upper Cretaceous Csehbánya Formation (Hungary, Bakony Mts). Acta Geologica Hungarica 49: 373–385. doi:
10.1556/AGeol.49.2006.4.5Makádi L (2013) A new polyglyphanodontine lizard (Squamata: Borioteiioidea) from the Late Cretaceous Iharkút locality (Santonian, Hungary). Cretaceous Research 46: 166–176. doi:
10.1016/j.cretres.2013.08.001Makádi L, Caldwell MW, Ősi A (2012) The first freshwater mosasauroid (Upper Cretaceous, Hungary) and a new clade of basal mosasauroids. PLoS One 7: e51781. doi:
10.1371/journal.pone.0051781Mannion PD (2009) A rebbachisaurid sauropod from the Lower Cretaceous of the Isle of Wight, England. Cretaceous Research 30: 521–526. doi:
10.1016/j.cretres.2008.09.005Mannion PD, Upchurch P, Hutt S (2011) New rebbachisaurid (Dinosauria: Sauropoda) material from the Wessex Formation (Barremian, Early Cretaceous), Isle of Wight, United Kingdom. Cretaceous Research 32: 774–780. doi:
10.1016/j.cretres.2011.05.005Martill DM (1993) Fossils of the Santana and Crato Formations, Brazil. Palaeontological Association, London, 159 pp.
Martill DM, Bechly G, Loveridge RF (2007) The Crato Fossil Beds of Brazil: Window into an Ancient World Cambridge University Press, New York, 674 pp. doi:
10.1017/CBO9780511535512Martill DM, Frey E (1998a) A new pterosaur Lagerstatte in N. E. Brazil (Crato Formation; Aptian, Lower Cretaceous): preliminary observations. Oryctos 1: 79–85.
Martill DM, Frey E (1998b) A possible azhdarchid pterosaur from the Crato Formation (Early Cretaceous, Aptian) of Brazil. Abstract 3rd European Workshop on Vertebrate Palaeontology, Maastricht. 49.
Martill DM, Frey E (1999) A possible azhdarchid pterosaur from the Crato Formation (Early Cretaceous, Aptian) of northeast Brazil. Geologie en Mijnbouw 78: 315–318. doi:
10.1023/A:1003843115145Martill DM, Frey E, Sadaqah RM (1996) The first dinosaur from the Hashemite Kingdom of Jordan. Neues Jahrbuch für Geologie und Paläontologie Monatshefte: 147–154.
Martill DM, Frey E, Sadaqah RM, Khoury HN (1998) Discovery of the holotype of the giant pterosaur
Titanopteryx philadephiae Arambourg 1959, and the status of
Arambourgiania and
Quetzalcoatlus. Neues Jahrbuch für Geologie und PaläontologieAbhandlungen 207: 57–76.
Martill DM, Naish D (2001) Dinosaurs of the Isle of Wight. The Palaeontological Association, London, 433 pp.
Martill DM, Naish D (2006) Cranial crest development in the azhdarchoid pterosaur
Tupuxuara, with a review of the genus and tapejarid monophyly. Palaeontology 49: 925–941. doi:
10.1111/j.1475-4983.2006.00575.xMartill DM, O'Sullivan M, Newman C (2013) A possible azhdarchid pterosaur (Pterosauria, Azhdarchidae) in the Durlston Formation (Early Cretaceous, Berriasian) of southern England. Cretaceous Research 43: 26–39. doi:
10.1016/j.cretres.2013.02.007Martill DM, Unwin DM (2012) The world’s largest toothed pterosaur, NHMUK R481, an incomplete rostrum of
Coloborhynchus capito (Seeley, 1870) from the Cambridge Greensand of England. Cretaceous Research 34: 1–9. doi:
10.1016/j.cretres.2011.09.003Martill DM, Witton MP (2008) Catastrophic failure in a pterosaur skull from the Cretaceous Santana Formation of Brazil. Zitteliana B 28: 175–183.
Martill DM, Witton MP, Gale AS (2008) Possible azhdarchoid pterosaur remains from the Coniacian (Late Cretaceous) of England. Zitteliana B 28: 209–218.
Martin JE, Buffetaut E (2005) An overview of the Late Cretaceous crocodilian assemblage from Cruzy, southern France. Kaupia 14: 33–40.
Martin M (1984) Révision des Arganodontidés et des Néocératodontidés (Dipnoi, Ceratodontiformes) du Crétacé africain. Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen 169: 225–260.
Martinelli AG, Forasiepi AM (2004) Late Cretaceous vertebrates from Bajo de Santa Rosa (Allen Formation), Rio Negro province, Argentina, with the description of a new sauropod dinosaur (Titanosauridae). Revista del Museo Argentino de Ciencias Naturales Nueva Serie 6: 257–305.
Martinson GG (1965) [Biostratigraphy and fauna of continental Cretaceous of Fergana]. [Cretaceous Continental Deposits of Fergana]. Nauka, Moscow, Leningrad, 101–131.
Martinson GG (1982) [Mesozoic Lacustrine Basins of Mongolia. Paleogeography, Lithology, Paleobiogeochemistry, Paleontology]. Nauka, Leningrad, 212 pp.
Martinson GG (1990) [Non marine bivalvian mollusks from the Cretaceous deposits of Aral Sea area and Turgai flexure]. Paleontologicheskii Zhurnal: 17–24.
Martinson GG (1997) [Some new species of Cretaceous non-marine mollusks from the continental deposits of Aral Sea region]. Paleontologicheskii Zhurnal: 10–13.
Martinson GG, Nesov LA, Starobogatov YI (1986) [Unique finding of gill apparatus of bivalve molluscs Trigonioidoidea from the Cretaceous deposits of Kyzylkum]. Byuleten' Moskovskogo Obschestva Ispytatelei Prirody, Otdel Geologicheskii 61: 94–98.
Martinson GG, Nikitin VG (1978) [Stratigraphic position of mollusks from continental deposits of northern and eastern Aral Sea Area]. Izvestiya Akademii Nauk SSSR, Seriya Geologicheskaya: 139–142.
Martinson GG, Nikitin VG, Teplova LS, Vasiliev IV (1966) [Stratigraphy and correlation of Cretaceous continental deposits of Aral Sea area]. Sovetskaya Geologiya: 92–103.
Maryańska T (1977) Ankylosauridae (Dinosauria) from Mongolia. Palaeontologia Polonica 37: 85–151.
Mateer NJ (1989) Upper Cretaceous reptilian eggs from the Zhejiang Province, China. In: Gillette DD, Lockley MG (Eds) Dinosaur Tracks and Traces. Cambridge University Press, Cambridge, 115–118.
Matsukawa M, Obata I (1994) Dinosaurs and sedimentary environments in the Japanese Cretaceous: a contribution to dinosaur facies in Asia based on molluscan palaeontology and stratigraphy. Cretaceous Research 15: 101–125. doi:
10.1006/cres.1994.1005Matsukawa M, Shibata K, Kukihara R, Koarai K, Lockley M (2005) Review of Japanese dinosaur track localities: Implications for ichnotaxonomy, paleogeography and stratigraphic correlation. Ichnos 12: 201–222. doi:
10.1080/10420940591009231Matsumoto R, Evans SE (2010) Choristoderes and the freshwater assemblages of Laurasia. Journal of Iberian Geology 36: 253–274. doi:
10.5209/rev_JIGE.2010.v36.n2.11Matsumoto T (1939) [Geology of Mifune district, Kumamoto Prefecture, Kyusyu (with special reference to the Cretaceous system)]. Journal of the Geological Society of Japan 46: 1–12. doi:
10.5575/geosoc.46.1Matsumoto T, Noda M (1986) Some inoceramids (Bivalvia) from the Cenomanian (Cretaceous) of Japan. I. New or little known four species from Hokkaido and Kyushu. Transactions and Proceedings of the Palaeontological Society of Japan, New Series 143: 409–421.
McGowen MR, Padian K, De Sosa MA, Harmon RJ (2002) Description of
Montanazhdarcho minor, an azhdarchid pterosaur from the Two Medicine Formation (Campanian) of Montana. PaleoBios 22: 1–9.
McKenna MC (1961) On the shoulder girdle of the mammalian subclass Allotheria. American Museum Novitates 2066: 1–27.
Merriam JC, Gilmore CW (1928) An ichthyosaurian reptile from the marine Cretaceous of Oregon. Carnegie Institution of Washington Publication 393: 3–4.
Mertinene RA, Nesov LA (1985) [Hybodont sharks from the Cretaceous of Middle Asia]. Doklady Akademii Nauk Tadzhikskoi SSR 28: 588–592.
Mertinene RA, Nesov LA (1991) [Sclerophagous sharks from the group Archaoselachii of the Cretaceous of Middle asia and Mongolia]. Doklady Akademii Nauk Tadzhikskoi SSR 34: 54–58.
Miki A (1977) Late Cretaceous pollen and spore floras of northern Japan: Composition and interpretation. Journal of the Faculty of Science, Hokkaido University, Series 4, Geology and Mineralogy 17: 399–436]
Milner AC (2002) Reptiles. In: Smith AB, Batten DJ (Eds) Fossils of the Chalk Second Edition, Revised and Enlarged. The Palaeontological Association, London, 325–343.
Misaki A, Maeda H, Kumagae T, Ichida M (2014) Commensal anomiid bivalves on Late Cretaceous heteromorph ammonites from south-west Japan. Palaeontology 57: 77–95. doi:
10.1111/pala.12050Mkhitaryan TG, Averianov AO (2011) New material and phylogenetic position of
Aidachar paludalis Nesov, 1981 (Actinopterygii, Ichthyodectiformes) from the Late Cretaceous of Uzbekistan. Proceedings of the Zoological Institute RAS 315: 181–192.
Molnar RE, Wiffen J (1994) A Late Cretaceous polar dinosaur fauna from New Zealand. Cretaceous Research 15: 689–706. doi:
10.1006/cres.1994.1038Monteillet J, Lappartient JR (1981) Fruits et graines du Crétacé supérieur des carrières de Paki (Sénégal). Review of Palaeobotany and Palynology 34: 331–344. doi:
10.1016/0034-6667(81)90049-XMonteillet J, Lappartient JR, Taquet P (1982) Un Ptérosaurien géant dans le Crétacé supérieur de Paki (Sénégal). Comptes Rendus des Seances de l'Academie des Sciences, Serie II Mecanique-Physique, Chimie, Sciences de l'Univers, Sciences de la Terre 295: 409–414.
Montellano M (1988)
Alphadon halleyi (Didelphidae, Marsupialia) from the Two Medicine Formation (Late Cretaceous, Judithian) of Montana. Journal of Vertebrate Paleontology 8: 378–382. doi:
10.1080/02724634.1988.10011726Montellano M, Weil A, Clemens WA (2000) An exceptional specimen of
Cimexomys judithae (Mammalia: Multituberculata) from the Campanian Two Medicine Formation of Montana, and the phylogenetic status of
Cimexomys. Journal of Vertebrate Paleontology 20: 333–340. doi:
10.1671/0272-4634(2000)020[0333:AESOCJ]2.0.CO;2Morozumi Y (1985) Late Cretaceous (Campanian and Maastrichtian) ammonites from Awaji Island, southwest Japan. Bulletin of the Osaka Museum of Natural History 39: 1–58.
Mu Z, Cai Z (1992) The K-Ar age of the
Zhejiangopterus fossil layer and its geological significance. Acta Scientiarum Naturalium Universitatis Pekinensis 28: 250–256.
Murry PA, Winkler DA, Jacobs LL (1991) An azhdarchid pterosaur humerus from the Lower Cretaceous Glen Rose Formation of Texas. Journal of Paleontology 65: 167–170.
Mustafa HA, Zalmout IS (1999) Additional fish teeth fossils from the early Maastrichtian of Esh-Shidiya Mine, SE-Jordan. Abhath Al-Yarmouk 8: 111–126\
Naish D, Dyke GJ, Cau A, Escuillié F, Godefroit P (2012) A gigantic bird from the Upper Cretaceous of Central Asia. Biology Letters 8: 97–100. doi:
10.1098/rsbl.2011.0683Naish D, Martill DM (2002) A reappraisal of
Thecocoelurus daviesi (Dinosauria: Theropoda) from the Early Cretaceous of the Isle of Wight. Proceedings of the Geologists' Association 113: 23–30.
Naish D, Martill DM, Cooper D, Stevens KA (2004a) Europe's largest dinosaur? A giant brachiosaurid cervical vertebra from the Wessex Formation (Early Cretaceous) of southern England. Cretaceous Research 25: 787–795. doi:
10.1016/j.cretres.2004.07.002Naish D, Martill DM, Frey E (2004b) Ecology, systematics and biogeographical relationships of dinosaurs, including a new theropod, from the Santana Formation (?Albian, Early Cretaceous) of Brazil. Historical Biology 16: 57–70. doi:
10.1080/08912960410001674200Naish D, Simpson M, Dyke GJ (2013) A new small-bodied azhdarchoid pterosaur from the Lower Cretaceous of England and its implications for pterosaur anatomy, diversity and phylogeny. PLoS One 8: e58451. doi:
10.1371/journal.pone.0058451Nesov LA (1977a) [A new genus of two clawed turtle from the Upper Cretaceous of Karakalpakia]. Paleontologicheskii Zhurnal: 103–114.
Nesov LA (1977b) [On some cranial features of two late Cretaceous turtles]. Vestnik Leningradskogo Universiteta, Seriya Biologiya 21: 45–48.
Nesov LA (1978) [Archaic Late Cretaceous turtle from Western Uzbekistan]. Paleontologicheskii Zhurnal: 101–105.
Nesov LA (1981a) [Cretaceous salamanders and frogs of Kizylkum Desert]. Trudy Zoologicheskogo Instituta AN SSSR 101: 57–88.
Nesov LA (1981b) [Flying reptiles of Late Cretaceous of Kyzylkums]. Paleontologicheskii Zhurnal: 98–104.
Nesov LA (1982) [The most ancient mammals of the USSR]. Ezhegodnik Vsesoyuznogo Paleontologicheskogo Obshchestva 25: 228–242.
Nesov LA (1984a) Data on Late Cretaceous turtles from the USSR. Stvdia Geologica Salmanticensia 1 especial: 215–223.
Nesov LA (1984b) [On some remains of mammals in the Cretaceous deposits of the Middle Asia]. Vestnik Zoologii: 60–65.
Nesov LA (1984c) [Pterosaurs and birds from the Late Cretaceous of Middle Asia]. Paleontologicheskii Zhurnal: 47–57.
Nesov LA (1985a) [New mammals from the Cretaceous of Kyzylkum]. Vestnik Leningradskogo Universiteta, Seriya 7: 8–18.
Nesov LA (1985b) [Rare bony fishes, terrestrial lizards and mammals from the zone of estuaries and coastal plains of the Cretaceous of Kizylkum]. Ezhegodnik Vsesoyuznogo Paleontologicheskogo Obshchestva 28: 199–219.
Nesov LA (1986a) [The first finding of Late Cretaceous bird
Ichthyornis in Old World and some other bird bones from Cretaceous and Paleogene of Soviet Middle Asia]. Trudy Zoologicheskogo Instituta AN SSSR 147: 31–38.
Nesov LA (1986b) Some late Mesozoic and Paleocene turtles of Soviet Middle Asia. Stvdia Geologica Salamanticensia, Stvdia Palaecheloniologica: 7–22.
Nesov LA (1987a) On some Mesozoic turtles of the Soviet Union, Mongolia and China, with comments on systematics. Stvdia Geologica Salamanticensia, Stvdia Palaecheloniologica 2: 87–102.
Nesov LA (1987b) [Results of search and study of Cretaceous and early Paleogene mammals on the territory of the USSR]. Ezhegodnik Vsesoyuznogo Paleontologicheskogo Obshchestva 30: 199–218.
Nesov LA (1988a) [Assemblages of late Mesozoic and Paleocene vertebrates of Middle Asia]. In: Bogdanova TN, Oshurkova MV (Eds) [Formation and Evolution of the Continental Biotas]. Izdatelstvo Nauka, Leningrad, 93–101.
Nesov LA (1988b) Late Mesozoic amphibians and lizards of Soviet Middle Asia. Acta Zoologica Cracoviensa 31: 475–486.
Nesov LA (1988c) [New Cretaceous and Paleogene birds of Soviet Middle Asia and Kazakhstan and environments]. Trudy Zoologicheskogo Instituta AN SSSR 182: 116–123.
Nesov LA (1988d) [Traces of vital functions of organisms of the Late Mesozoic - Paleocene of Middle Asia and Kazakhstan as indicators of the ancient vertebrates biotops]. In: Bogdanova TN, Khozatsky LI, Ishchenko AA (Eds) [Fossil Traces of Vital Activity and Dynamics of the Environment in Ancient Biotopes]. Naukova Dumka, Kiev, 76–90.
Nesov LA (1989) [New findings of remains of dinosaurs, crocodiles, and flying reptiles of the Late Mesozoic of USSSR]. [Questions of Herpetology Abstracts of VII Herpetological Conference]. Kiev, 173–174.
Nesov LA (1990a) [Flying reptiles of the Jurassic and Cretaceous of the USSR and significance of their remains for the paleogeographic environmental reconstruction]. Vestnik Leningradskogo Universiteta, Seriya 7: Geologiya, Geografiya 4(28): 3–10.
Nesov LA (1990b) [Small ichthyornithiform bird and other bird remains from Bissekty Formation (Upper Cretaceous) of Central Kyzylkum Desert]. Trudy Zoologicheskogo Instituta AN SSSR 210: 59–62.
Nesov LA (1991) [Giant flying reptiles of the family Azhdarchidae. I. Morphology, systematics]. Vestnik Leningradskogo Universiteta, Seriya 7: Geologiya, Geografiya: 14–23.
Nesov LA (1992a) Mesozoic and Paleogene birds of the USSR and their paleoenvironments. In: Campbell KE, Jr. (Ed.) Papers in Avian Paleontology Honoring Pierce Brodkorb. Los Angeles County Museum, Los Angeles, 465–478.
Nesov LA (1992b) [Review of localities and remains of Mesozoic and Paleogene birds of the USSR and the description of new findings]. Russkii Ornitologicheskii Zhurnal 1: 7–50.
Nesov LA (1993) [New Mesozoic mammals of Middle Asia and Kazakhstan and comments about evolution of theriofaunas of Cretaceous coastal plains of Asia]. Trudy Zoologicheskogo Instituta RAN 249: 105–133.
Nesov LA (1995) [Dinosaurs of Northern Eurasia: New Data about Assemblages, Ecology and Paleobiogeography]. Izdatelstvo Sankt-Peterburgskogo Universiteta, Saint Petersburg, 156 pp.
Nesov LA (1997) [Cretaceous Nonmarine Vertebrates of Northern Eurasia] (Posthumous edition by L.B. Golovneva and A.O. Averianov). Izdatel'stvo Sankt-Peterburgskogo Universiteta, Saint Petersburg, 218 pp.
Nesov LA, Archibald JD, Kielan-Jaworowska Z (1998) Ungulate-like mammals from the Late Cretaceous of Uzbekistan and a phylogenetic analysis of Ungulatomorpha. In: Beard KC, Dawson MR (Eds) Dawn of the Age of Mammals in Asia. Bulletin of the Carnegie Museum of Natural History, 40–88.
Nesov LA, Borkin LY (1983) [New findings of birds' bones from the Cretaceous of Mongolia and Middle Asia]. Trudy Zoologicheskogo Instituta AN SSSR 116: 108–110.
Nesov LA, Golovneva LB (1983) [Changes of vertebrate complexes of Cenomanian - Santonian (Late Cretaceous) of Kyzylkums]. In: Oleinikov AN (Ed) [Paleontology and Evolution of Biosphera Transactions of XXV Session of the All-Union Paleontological Society]. Nauka, Leningrad, 126–134.
Nesov LA, Kaznyshkin MN (1983) [New sturgeons of the Cretaceous and Paleogene of the USSR]. [Problems of Modern Paleoichthyology]. Moscow, 68–76.
Nesov LA, Kaznyshkina LF, Cherepanov GO (1987) [Dinosaurs, crocodiles, and other archosaurs of Late Mesozoic of Middle Asia and their place in ecosystems]. In: Kruchinina NV, Modzalevskaya TL (Eds) [Theoretical and Applied Aspects of Modern Paleontology Abstracts of Reports of XXXIII Session of the All-Union Paleontological Society]. Vsesoyuznoe Paleontologicheskoe Obshchestvo, Leningrad, 46–47.
Nesov LA, Kaznyshkina LF, Cherepanov GO (1989) [Ceratopsian dinosaurs and crocodiles of the Mesozoic of Middle Asia]. In: Bogdanova TN, Khozatsky LI (Eds) [Theoretical and Applied Aspects of Modern Paleontology]. Nauka, Leningrad, 144–154.
Nesov LA, Khisarova GD (1988) [New data about Upper Cretaceous vertebrates from Shakh-Shakh and Baibolat localities (North-Eastern Aral area)]. Materialy po Istorii Fauny i Flory Kazakhstana 10: 5–14.
Nesov LA, Khosatzky LI (1980) [Turtles of the genus
Lindholmemys from the Late Cretaceous of the USSR]. Ezhegodnik Vsesoyuznogo Paleontologicheskogo Obshchestva 23: 250–264.
Nesov LA, Krassovskaya TB (1984) [Transformations in the composition of turtles assemblages of Late Cretaceous of Middle Asia]. Vestnik Leningradskogo Universiteta, Seriya Biologiya 1: 15–25.
Nesov LA, Mertinene RA (1982) [Teeth of cartilagenous fishes as source of data about age of estuarian deposits of the Cretaceous of Middle Asia]. In: Kruchinina NV (Ed) [Paleontology and Detailed Stratigraphic Correlation Abstracts of Reports of XXVIII Session of the All-Union Paleontological Society]. Vsesoyuznoe Paleontologicheskoe Obshchestvo, Tashkent, 56–57.
Nesov LA, Mertinene RA (1986) [Remains of chondrichthyan fishes from the Cretaceous of Middle Asia and Kazakhstan as a source of information on the age and origin of the deposits]. In: Krymgoltz GY, Belenkova VS (Eds) Paleontology and Detailed Stratigraphic Correlation. Izdatelstvo Nauka, Leningrad, 35–41.
Nesov LA, Panteleeva TM (1999) [Gar
Atractosteus turanensis sp. nov. (Lepisosteidae) from the Upper Cretaceous of Kyzylkum Desert]. Trudy Zoologicheskogo Instituta RAN 277: 104–118.
Nesov LA, Panteleyev AV (1993) [About similarities of the Late Cretaceous ornithofaunas of South America and Western Asia]. Trudy Zoologicheskogo Instituta RAN 252: 84–94.
Nesov LA, Sigogneau-Russell D, Russell DE (1994) A survey of Cretaceous tribosphenic mammals from Middle Asia (Uzbekistan, Kazakhstan and Tajikistan), of their geological setting, age and faunal environment. Palaeovertebrata 23: 51–92.
Nesov LA, Trofimov BA (1979) [The oldest insectivore of the Cretaceous of the Uzbek SSR]. Doklady Akademii Nauk SSSR 247: 952–954.
Nesov LA, Udovichenko NI (1986) [New findings of remains of Cretaceous and Paleogene vertebrates of Middle Asia]. Voprosy Paleontologii 9: 129–136.
Nesov LA, Verzilin NN (1983) [Remains of acipenserid fishes as evidence of vertical climatic zonation in Middle Asia during Cretaceous Period]. Vestnik Leningradskogo Universiteta, Seriya Geologiya 2: 5–10.
Nesov LA, Yarkov AA (1989) [New Cretaceous-Paleogene birds of the USSR and some remarks on the origin and evolution of the class Aves]. Trudy Zoologicheskogo Instituta AN SSSR 197: 78–97.
Neuman AG, Brinkman DB (2005) Fishes of the fluvial beds. In: Currie PJ, Koppelhus EB (Eds) Pterosaurs Dinosaur Provincial Park: A Spectacular Ancient Ecosystem Revealed. Indiana University Press, Bloomington, 167–185.
Newbrey MG, Wilson MVH, Ashworth AC (2007) Centrum growth patterns provide evidence for two small taxa of Hiodontidae in the Cretaceous Dinosaur Park Formation. Canadian Journal of Earth Sciences 44: 721–732. doi:
10.1139/e06-137Nikiforova KV (1960) [Cenozoic of Golodnaya Steppe of Central Kazakhstan]. Trudy Geologicheskogo Instituta AN SSSR 45: 1–254.
Nopcsa F (1926) Die Reptilien der Gosau in neuer Beleuchtung. Centralblatt für Mineralogie, Geologie und Paläontologie, Abteilung B: 520–523.
Norman DB, Kurzanov SM (1997) On Asian ornithopods (Dinosauria: Ornithischia). 2.
Arstanosaurus akkurganensis Shilin and Suslov, 1982. Proceedings of the Geologists' Association 108: 191–199.
Norman DB, Sues H-D (2000) Ornithopods from Kazakhstan, Mongolia and Siberia. In: Benton MJ, Shishkin MA, Unwin DM, Kurochkin EN (Eds) The Age of Dinosaurs in Russia and Mongolia. Cambridge University Press, Cambridge, 462–479.
Noubhani A, Cappetta H (1997) Les Orectolobiformes, Carcharhiniformes et Myliobatiformes (Elasmobranchii, Neoselachii) des Bassins à phosphate du Maroc (Maastrichtien-Lutétien basal). Systématique, biostratigraphie, évolution et dynamique des faunes. Palaeo Ichthyologica 8: 1–327.
Novacek MJ, Clemens WA (1977) Aspects of intrageneric variation and evolution of
Mesodma (Multituberculata, Mammalia). Journal of Paleontology 51: 701–717.
Novas FE, Kundrat M, Agnolín FL, Ezcurra MD, Ahlberg PE, Isasi MP, Arriagada A, Chafrat P (2012) A new large pterosaur from the Late Cretaceous of Patagonia. Journal of Vertebrate Paleontology 32: 1447–1452. doi:
10.1080/02724634.2012.703979Novas FE, Pol D, Canale JI, Porfiri JD, Calvo JO (2009) A bizarre Cretaceous theropod dinosaur from Patagonia and the evolution of Gondwanan dromaeosaurids. Proceedings of the Royal Society B: Biological Sciences 276: 1101–1107. doi:
10.1098/rspb.2008.1554O'Connor PM, Sertich JJW, Manthi FK (2011) A pterodactyloid pterosaur from the Upper Cretaceous Lapurr sandstone, West Turkana, Kenya. Anais da Academia Brasileira de Ciências 83: 309–315. doi:
10.1590/S0001-37652011000100019Obata I, Hasegawa Y, Otsuka H (1972) Preliminary report on the Cretaceous reptile fossils from Hokkaido. Memoirs of the National Science Museum 5: 213–222.
Obata I, Shibata K, Matsukawa M, Unwin DM (2007) New record of a pterosaur from the Late Cretaceous Izumi Group, Awaji Island, Hyogo Prefecture, Japan. Annual Report of the Fukada Geological Institute 8: 149–162.
Ochev VG (1976) [New pliosaur from the Upper Cretaceous of Penza Region]. Paleontologicheskii Zhurnal: 135–138.
Okazaki Y, Kitamura N (1996) The first discovery of a pterosaur from the Cretaceous Mifune Group, Kyushu, Japan. Bulletin of the Kitakyushu Museum of Natural History 15: 133–136.
Ősi A (2004a) Dinosaurs from the Late Cretaceous of Hungary - similarities and differences with other European Late Cretaceous faunas. Revue de Paléobiologie Volume spéciale 9: 51–54.
Ősi A (2004b) The first dinosaur remains from the Upper Cretaceous of Hungary (Csehbánya Formation, Bakony Mts). Geobios 37: 749–753. doi:
10.1016/j.geobios.2003.06.005Ősi A (2008) Enantiornithine bird remains from the Late Cretaceous of Hungary. Oryctos 7: 55–60.
Ősi A, Apesteguía S, Kowalewski M (2010a) Non-avian theropod dinosaurs from the early Late Cretaceous of central Europe. Cretaceous Research 31: 304–320. doi:
10.1016/j.cretres.2010.01.001Ősi A, Buffetaut E (2011) Additional non-avian theropod and bird remains from the early Late Cretaceous (Santonian) of Hungary and a review of the European abelisauroid record. Annales de Paleontologie 97: 35–49. doi:
10.1016/j.annpal.2011.07.001Ősi A, Buffetaut E, Prondvai E (2011) New pterosaurian remains from the Late Cretaceous (Santonian) of Hungary (Iharkút, Csehbánya Formation). Cretaceous Research 32: 456–463. doi:
10.1016/j.cretres.2011.01.011Ősi A, Butler RJ, Weishampel DB (2010b) A Late Cretaceous ceratopsian dinosaur from Europe with Asian affinities. Nature 465: 466–468. doi:
10.1038/nature09019Ősi A, Clark JM, Weishampel DB (2007) First report on a new basal eusuchian crocodyliform with multicusped teeth from the Upper Cretaceous (Santonian) of Hungary. Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen 243: 169–177. doi:
10.1127/0077-7749/2007/0243-0169Ősi A, Makádi L (2009) New remains of
Hungarosaurus tormai (Ankylosauria, Dinosauria) from the Upper Cretaceous of Hungary: skeletal reconstruction and body mass estimation. Paläontologische Zeitschrift 83: 227–245. doi:
10.1007/s12542-009-0017-5Ősi A, Pereda Suberbiola X, Földes T (2014) Partial skull and endocranial cast of the ankylosaurian dinosaur
Hungarosaurus from the Late Cretaceous of Hungary: implications for locomotion. Palaeontologia Electronica 17: 1A; 18p
Ősi A, Prondvai E, Butler RJ, Weishampel DB (2012a) Phylogeny, histology and inferred body size evolution in a new rhabdodontid dinosaur from the Late Cretaceous of Hungary. PLoS One 7: e44318. doi:
10.1371/journal.pone.0044318Ősi A, Rabi M, Makádi L, Szentesi Z, Botfalvai G, Gulyás P (2012b) The Late Cretaceous continental vertebrate fauna from Iharkút (Western Hungary): a review. In: Godefroit P (Ed) Bernissart Dinosaurs and Early Cretaceous Terrestrial Ecosystems. Indiana University Press, Bloomington, 532–569.
Ősi A, Weishampel DB, Jianu C-M (2005) First evidence of azhdarchid pterosaurs from the Late Cretaceous of Hungary. Acta Palaeontologica Polonica 50: 777–787.
Owens JP, Minard JP, Sohl NF, Mello JF (1970) Stratigraphy of the outcropping post-Magothy Upper Cretaceous formations in southern New Jersey and northern Delmarva Peninsula, Delaware and Maryland. United States Geological Survey Professional Paper 674: 1–60.
Padian K (1984) A large pterodactyloid pterosaur from the Two Medicine Formation (Campanian) of Montana. Journal of Vertebrate Paleontology 4: 516–524. doi:
10.1080/02724634.1984.10012027Padian K, Ricqles AJ, de, Horner JR (1995) Bone histology determines identification of a new fossil taxon of pterosaur (Reptilia, Archosauria). Comptes Rendus de L'Academie des Sciences, Serie II 320: 77–84.
Padian K, Smith MB (1992) New light on Late Cretaceous pterosaur material from Montana. Journal of Vertebrate Paleontology 12: 87–92. doi:
10.1080/02724634.1992.10011433Panteleyev AV (1999) The Late Cretaceous Enantiornithines of southwestern margin of ancient Asia. In: Leanza HA (Ed) VII International Symposium on Mesozoic Terrestrial Ecosystems Abstracts. Museo Argentino de Ciencias Naturales, Buenos Aires, 50–51.
Parris DC, Grandstaff BS, Clements D (2004) A pterosaur femur from the Upper Cretaceous of North Carolina. Southeastern Geology 43: 52–55.
Pereda Suberbiola X, Astibia H, Murelaga X, Elorza JJ, Gomez-Alday JJ (2000) Taphonomy of the Late Cretaceous dinosaur-bearing beds of the Laño Quarry (Iberian Peninsula). Palaeogeography, Palaeoclimatology, Palaeoecology 157: 247–275. doi:
10.1016/S0031-0182(99)00169-8Pereda Suberbiola X, Bardet N, Iarochene M, Bouya B, Amaghzaz M (2004) The first record of a sauropod dinosaur from the Late Cretaceous phosphates of Morocco. Journal of African Earth Sciences 40: 81–88. doi:
10.1016/j.jafrearsci.2004.07.002Pereda Suberbiola X, Bardet N, Jouve S, Iarochene M, Bouya B, Amaghzaz M (2003) A new azhdarchid pterosaur from the Late Cretaceous phosphates of Morocco. In: Buffetaut E, Mazin J-M (Eds) Evolution and Palaeobiology of Pterosaurs. Geological Society Special Publication, 217: 79–90.
Pereda Suberbiola X, Canudo JI, Company J, Cruzado-Caballero P, Ruiz-Omeñaca JI (2009) Hadrosauroid dinosaurs from the latest Cretaceous of the Iberian Peninsula. Journal of Vertebrate Paleontology 29: 946–951. doi:
10.1671/039.029.0317Pereda Suberbiola X, Company J, Ruiz-Omeñaca JI (2007) Azhdarchid pterosaurs from the Late Cretaceous (Campanian-Maastrichtian) of the Iberian Peninsula: an update. In: Hone D (Ed.) Flugsaurier The Wellnhofer Pterosaur Meeting. Bavarian State Collection for Palaeontology, Munich, One page not paginated.
Pereda Suberbiola X, Galton PM (2001) Reappraisal of the nodosaurid ankylosaur
Struthiosaurus austriacus Bunzel from the Upper Cretaceous Gosau Beds of Austria. In: Carpenter K (Ed) The Armored Dinosaurs. Indiana University Press, Bloomington, 173–210.
Perkins BF (1974) Paleoecology of a rudist reef complex in the Comanche Cretaceous Glen Rose Limestone of central Texas. Geoscience and Man 8: 131–173.
Perle A (1977) [On the first discovery of alectrosaur (Tyrannosauridae, Theropoda) from the Late Cretaceous of Mongolia]. Problemy Geologii Mongolii: 104–113.
Perle A (1979) [Segnosauridae--a new family of theropods from the Late Cretaceous of Mongolia]. Trudy Sovmestnoi Sovetsko-Mongol'skoi Paleontologicheskoi Ekspeditsii 8: 45–55.
Perle A (1981) [A new segnosaurid from the Upper Cretaceous of Mongolia]. Trudy Sovmestnoi Sovetsko-Mongol'skoi Paleontologicheskoi Ekspeditsii 15: 50–59.
Perle A, Norell MA, Clark JM (1999) A new maniraptoran theropod
Achillobator giganticus (Dromaeosauridae) from the Upper Cretaceous of Burkhant, Mongolia. National University of Mongolia, Ulan Bataar, 105 pp.
Pervushov EM, Arkhangelsky MS, Ivanov AV (1999) [Catalogue of Sea Reptiles Remains Localities in the Jurassic and Cretaceous Deposits of the Lower Volga Region]. College, Saratov, 230 pp.
Pinheiro FL, Fortier DC, Schultz CL, Andrade JAFG, De, Bantim RAM (2011) New information on the pterosaur
Tupandactylus imperator, with comments on the relationships of Tapejaridae. Acta Palaeontologica Polonica 56: 567–580. doi:
10.4202/app.2010.0057Pittet F, Cavin L, Poyato-Ariza FJ (2009) A new ostariophysan fish from the early Late Cretaceous (Cenomanian) of SE Morocco, with a discussion of its phylogenetic relationships. In: Grande T, Poyato-Ariza FJ, Diogo R (Eds) A Comprehensive Review of Gonorynchiformes and of Ostariophysan Relationships. Science Publishers, 332–355.
Popov EV, Machalski M (2014) Late Albian chimaeroid fishes (Holocephali, Chimaeroidei) from Annopol, Poland. Cretaceous Research 47: 1–18. doi:
10.1016/j.cretres.2013.09.011Posmosanu E, Cook E (2000) Vertebrate taphonomy and dinosaur palaeopathology from a Lower Cretaceous bauxite lens, North West Romania. Oryctos 3: 39–51.
Pufahl PK, Grimma KA, Abed AM, Sadaqah RM (2003) Upper Cretaceous (Campanian) phosphorites in Jordan: implications for the formation of a south Tethyan phosphorite giant. Sedimentary Geology 161: 175–205. doi:
10.1016/S0037-0738(03)00070-8Rabi M, Tong H, Botfalvai G (2012) A new species of the side-necked turtle
Foxemys (Pelomedusoides: Bothremydidae) from the Late Cretaceous of Hungary and the historical biogeography of the Bothremydini. Geological Magazine 149: 662–674. doi:
10.1017/S0016756811000756Rabi M, Vremir M, Tong H (2013) Preliminary overview of Late Cretaceous turtle diversity in Eastern Central Europe (Austria, Hungary, and Romania). In: Brinkman DB, Holroyd PA, Gardner JD (Eds) Morphology and Evolution of Turtles, Vertebrate Paleobiology and Paleoanthropology. Springer, Dordrecht. 307–336.
Rage J-C, Dutheil DB (2008) Amphibians and squamates from the Cretaceous (Cenomanian) of Morocco. A preliminary study, with description of a new genus of pipid frog. Palaeontographica, Abteilung A: Palaozoologie, Stratigraphie 285: 1–22.
Redman CM, Leighton LR (2009) Multivariate faunal analysis of the Turonian Bissekty Formation: Variation in the degree of marine influence in temporally and spatially averaged fossil assemblages. Palaios 24: 18–26. doi:
10.2110/palo.2007.p07-072rRezvyi AS (2007) Crocodyliforms from the Late Cretaceous (Turonian) Bissekty local fauna of Kyzylkum Desert, Uzbekistan. Journal of Vertebrate Paleontology 27: 134A
Riabinin AN (1931) [Dinosaur remains from the Upper Cretaceous of Amu-Darya River]. Zapiski Rossiiskogo Mineralogicheskogo Obshchestva 40: 114–118.
Riabinin AN (1935) [Remains of a turtle from the Upper Cretaceous deposits of Kyzylkum Desert]. Trudy Paleontologicheskogo Instituta AN SSSR 4: 69–77.
Rich THV, Gangloff RA, Hammer WR (1997) Polar dinosaurs. In: Currie PJ, Padian K (Eds) Encyclopedia of Dinosaurs. Academic Press, San Diego, London, Boston, New York, Sydney, Tokyo, Toronto, 562–573.
Roček Z, Nesov LA (1993) Cretaceous anurans from Central Asia. Palaeontographica, Abteilung A: Palaozoologie, Stratigraphie 226: 1–54.
Rodrigues T, Kellner AWA (2013) Taxonomic review of the
Ornithocheirus complex (Pterosauria) from the Cretaceous of England. ZooKeys 308: 1–112. doi:
10.3897/zookeys.308.5559Rodrigues T, Kellner AWA, Mader BJ, Russell DA (2006) Brief report on new pterosaur (Pterosauria, Pterodactyloidea) specimens from the Cretaceous of Morocco. Journal of Vertebrate Paleontology 26: 116A
Rodrigues T, Kellner AWA, Mader BJ, Russell DA (2011) New pterosaur specimens from the Kem Kem beds (Upper Cretaceous, Cenomanian) of Morocco. Rivista Italiana di Paleontologia e Stratigrafia 117: 149–160.
Rodriguez-De La Rosa RA (2003) Pterosaur tracks from the latest Campanian Cerro del Pueblo Formation of southeastern Coahuila, Mexico. In: Buffetaut E, Mazin J-M (Eds) Evolution and Palaeobiology of Pterosaurs. Geological Society Special Publication, 217, 275–282.
Rodriguez-De La Rosa RA, Cevallos-Ferriz SRS (1998) Vertebrates of the El Pelillal locality (Campanian, Cerro del Pueblo Formation), southeastern Coahuila, Mexico. Journal of Vertebrate Paleontology 18: 751–764. doi:
10.1080/02724634.1998.10011104Rogers JV (2003)
Pachycheilosuchus trinquei, a new procoelous crocodyliform from the Lower Cretaceous (Albian) Glen Rose Formation of Texas. Journal of Vertebrate Paleontology 23: 128–145. doi:
10.1671/0272-4634(2003)23[128:PTANPC]2.0.CO;2Rogers RR, Swisher CC, III, Horner JR (1993) 40Ar/39Ar age and correlation of the nonmarine Two Medicine Formation (Upper Cretaceous), northwestern Montana, U.S.A. Canadian Journal of Earth Sciences 30: 1066–1075. doi:
10.1139/e93-090Rougier GW, Chornogubsky L, Casadio S, Páez Arango N, Giallombardo A (2009) Mammals from the Allen Formation, Late Cretaceous, Argentina. Cretaceous Research 30: 223–238. doi:
10.1016/j.cretres.2008.07.006Rozhdestvensky AK (1964) [New data on the localities of dinosaurs on the territory of Kazakhstan and Middle Asia]. Nauchnye Trudy Tashkentskogo Gosudarstvennogo Universiteta Imeni VI Lenina, Seriya Geologiya: 227–241.
Rozhdestvensky AK (1968) [Hadrosaurs of Kazakhstan]. [Upper Paleozoic and Mesozoic Amphibians and Reptiles of the USSR]. Nauka, Moscow, 97–141.
Rozhdestvensky AK (1970) [About giant ungual phalanges of enigmatic Mesozoic vertebrates]. Paleontologicheskii Zhurnal: 131–141.
Rozhdestvensky AK (1973) [Study of Cretaceous reptiles in Russia]. Paleontologicheskii Zhurnal: 90–99.
Rozhdestvensky AK (1977) [Kansai locality of Cretaceous vertebrates in Fergana]. Ezhegodnik Vsesoyuznogo Paleontologicheskogo Obshchestva 20: 235–247.
Russell DA (1972) A pterosaur from the Oldman Formation (Cretaceous) of Alberta. Canadian Journal of Earth Sciences 9: 1338–1340. doi:
10.1139/e72-119Russell DA (1988) A check list of North American marine Cretaceous vertebrates including freshwater fishes. Occasional Papers of the Tyrrell Museum of Palaeontology 4: 1–57.
Russell DA (1996) Isolated dinosaur bones from the Middle Cretaceous of the Tafilat, Morocco. Bulletin du Muséum National d`Histoire Naturelle, Paris, 4e Série, Section C 18: 349–402.
Ryan MJ, Evans DC, Currie PJ, Brown CM, Brinkman DB (2012) New leptoceratopsids from the Upper Cretaceous of Alberta, Canada. Cretaceous Research 35: 69–80. doi:
10.1016/j.cretres.2011.11.018Sachs S, Hornung JJ (2006) Juvenile ornithopod (Dinosauria: Rhabdodontidae) remains from the Upper Cretaceous (Lower Campanian, Gosau Group) of Muthmannsdorf (Lower Austria). Geobios 39: 415–425. doi:
10.1016/j.geobios.2005.01.003Saegusa H, Furutani H (2004) [A cervical vertebra of an azhdarchid pterosaur from the Seidan Formation, Izumi Group]. Abstracts of the 111st Annual Meeting of the Geological Society of Japan. 252.
Sankey JT (2008) Diversity of latest Cretaceous (late Maastrichtian) small theropods and birds: teeth from Lance and Hell Creek formations, USA. In: Sankey JT, Baszio S (Eds) Vertebrate Microfossil Assemblages Their Role in Paleoecology and Paleobiogeography. Indiana University Press, Bloomington, 117–134.
Sato S (1961) Pollen analysis of carbonaceous matter from the Hakobuchi Group in the Enbetsu District, Northern Hokkaido, Japan. Palynological Study on Cretaceous Sediment (I). Journal of the Faculty of Science, Hokkaido University, Series 4, Geology and Mineralogy 11: 77–93.
Sato T, Konishi T, Hirayama R, Caldwell MW (2012) A review of the Upper Cretaceous marine reptiles from Japan. Cretaceous Research 37: 319–340. doi:
10.1016/j.cretres.2012.03.009Sayão JM, Kellner AWA (1998) Pterosaur wing with soft tissue from the Crato Member (Aptian-Albian), Santana Formation, Brazil. Journal of Vertebrate Paleontology 18: 75A
Sayão JM, Kellner AWA (2001) New data on the pterosaur fauna from Tendaguru (Tanzania), Upper Jurassic, Africa. Journal of Vertebrate Paleontology 21: 97A
Sayão JM, Kellner AWA (2007) Novo esqueleto parcial de pterossauro (Pterodactyloidea, Tapejaridae) do Membro Crato (Aptiano), Formacao Santana, Bacia do Araripe, nordeste do Brasil. Estudos Geologicos 16: 16–40.
Seeley HG (1870) The Ornithosauria: An Elementary Study of the Bones of Pterodactyles, Made from Fossil Remains Found in the Cambridge Upper Greensand. Deighton, Bell, and Co., Cambridge, 135 pp.
Seeley HG (1875) On an ornithosaurian (
Doratorhynchus validus) from the Purbeck Limestone of Langton near Swanage. Quarterly Journal of the Geological Society of London 31: 465–468. doi:
10.1144/GSL.JGS.1875.031.01-04.35Seeley HG (1881) The reptile fauna of the Gosau Formation preserved in the Geological Museum of the University of Vienna. Quarterly Journal of the Geological Society of London 37: 620–707. 10.1144/GSL.JGS.1881.037.01-04.49
Seeley HG (1901) Dragons of the Air. An Account of Extinct Flying Reptiles. Methuen, London, xiii+239 pp.
Selden P, Nudds JR (2012) Evolution of Fossil Ecosystems. Second Edition. University of Chicago Press, Chicago, 288 pp. doi:
10.1201/b15128Sepkoski JJ Jr. (1982) Mass extinctions in the Phanerozoic oceans: a review. Geological Society of America, Special Paper 190: 283–289.
Sepkoski JJ Jr. (1986) Phanerozoic overview of mass extinction. In: Raup DM, Jablonski D (Eds) Patterns and Processes in the History of Life. Springer Verlag, Berlin, 277–295. doi:
10.1007/978-3-642-70831-2_15Sereno PC, Beck AL, Dutheil DB, Gado B, Larsson HCE, Lyon GH, Marcot JD, Rauhut OWM, Sadleir RW, Sidor CA, Varricchio DD, Wilson GP, Wilson JA (1998) A long-snouted predatory dinosaur from Africa and the evolution of spinosaurids. Science 282: 1298–1302. doi:
10.1126/science.282.5392.1298Sereno PC, Dutheil DB, larochene M, Larsson HCE, Lyon GH, Magwene PM, Sidor CA, Varricchio DD, Wilson JA (1996) Predatory dinosaurs from the Sahara and Late Cretaceous faunal differentiation. Science 272: 986–991. doi:
10.1126/science.272.5264.986Sereno PC, Larsson HCE (2009) Cretaceous Crocodyliforms from the Sahara. ZooKeys 28: 1–143. doi:
10.3897/zookeys.28.325Setoguchi T, Tsubamoto T, Hanamura H, Hachiya K (1999) An early Late Cretaceous mammal from Japan, with reconsideration of the evolution of tribosphenic molars. Paleontological Research 3: 18–28.
Sha J (2007) Cretaceous trigonioidid (non-marine Bivalvia) assemblages and biostratigraphy in Asia with special remarks on the classification of Trigonioidacea. Journal of Asian Earth Sciences 29: 62–83. doi:
10.1016/j.jseaes.2006.01.003Shigeta Y, Tanabe K, Izukura M (2010)
Gaudryceras izumiense Matsumoto and Morozumi, a maastrichtian ammonoid from Hokkaido and Alaska and its biostratigraphic implications. Paleontological Research 14: 202–211. doi:
10.2517/1342-8144-14.3.202Shilin PV (1982) [Characteristic of Turonian and Senonian floras of Lower Syr-Dar'ya Uplift (Southern Kazakhstan)]. Materialy po Istorii Fauny i Flory Kazakhstana 8: 113–121.
Shilin PV (1986) [Late Cretaceous floras of Kazakhstan]. Nauka, Alma-Ata, 176 pp.
Shilin PV (1998) Cretaceous flora of Tyulikili, Northeastern Priaralye, Kazakstan. In: Lucas SG, Kirkland JI, Estep JW (Eds) Lower and Middle Cretaceous Terrestrial Ecosystems. New Mexico Museum of Natural History and Science Bulletin No. 14, 95–103.
Shilin PV, Suslov YV (1982) [Hadrosaur from North-Eastern Aral Sea area]. Paleontologicheskii Zhurnal: 131–135.
Shuvalov VF, Chkhikvadze VM (1975) [New data on Late Cretaceous turtles of South Mongolia]. Trudy Sovmestnoi Sovetsko-Mongol'skoi Paleontologicheskoi Ekspeditsii 2: 214–229.
Shuvalov VF, Trusova EK (1979) [Late Cretaceous conchostracans of Mongolia]. Trudy Sovmestnoi Sovetsko-Mongol'skoi Paleontologicheskoi Ekspeditsii 8: 83–93.
Simões TR (2012) Redescription of
Tijubina pontei, an Early Cretaceous lizard (Reptilia; Squamata) from the Crato Formation of Brazil. Anais da Academia Brasileira de Ciências 84: 79–93. doi:
10.1590/S0001-37652012005000001Simpson MI (1985) The stratigraphy of the Atherfield Clay Formation (Lower Aptian; Lower Cretaceous) at the type and other localities in southern England. Proceedings of the Geologists' Association 96: 23–45.
Sintsov IF (1872) [On Jurassic and Cretaceous fossils from Saratov Province]. Materialy dlya geologii Rossii 4: 1–127.
Skutschas PP (2007) New specimens of albanerpetontid amphibians from the Upper Cretaceous of Uzbekistan. Acta Palaeontologica Polonica 52: 819–821.
Skutschas PP (2009) Re-evaluation of
Mynbulakia Nesov, 1981 (Lissamphibia: Caudata) and description of a new salamander genus from the Late Cretaceous of Uzbekistan. Journal of Vertebrate Paleontology 29: 659–664. doi:
10.1671/039.029.0326Skutschas PP (2013) Mesozoic salamanders and albanerpetontids of Middle Asia, Kazakhstan, and Siberia. Palaeobiodiversity and Palaeoenvironments 93: 441–457. doi:
10.1007/s12549-013-0126-8Smith AB, Batten DJ (2002) Fossils of the Chalk. Second Edition. The Palaeontological Association, London, 374 pp.
Steel L, Martill DM, Kirk JRJ, Anders A, Loveridge RF, Frey E, Martin JG (1997)
Arambourgiania philadelphiae: giant wings in small halls. The Geological Curator 6: 305–313.
Storrs GW, Efimov MB (2000) Mesozoic crocodyliforms of north-central Eurasia. In: Benton MJ, Shishkin MA, Unwin DM, Kurochkin EN (Eds) The Age of Dinosaurs in Russia and Mongolia. Cambridge University Press, Cambridge, 402–419.
Sues H-D, Averianov AO (2009a) A new basal hadrosauroid dinosaur from the Late Cretaceous of Uzbekistan and the early radiation of duck-billed dinosaurs. Proceedings of the Royal Society B: Biological Sciences 276: 2549–2555. doi:
10.1098/rspb.2009.0229Sues H-D, Averianov AO (2009b)
Turanoceratops tardabilis —the first ceratopsid dinosaur from Asia. Naturwissenschaften 96: 645–652. doi:
10.1007/s00114-009-0518-9Sues H-D, Averianov AO (2013) Enigmatic teeth of small theropod dinosaurs from the Upper Cretaceous (Cenomanian–Turonian) of Uzbekistan. Canadian Journal of Earth Sciences 50: 306–314. doi:
10.1139/e2012-033Sukhanov VB (2000) Mesozoic turtles of Middle and Central Asia. In: Benton MJ, Shishkin MA, Unwin DM, Kurochkin EN (Eds) The Age of Dinosaurs in Russia and Mongolia. Cambridge University Press, Cambridge, 309–367.
Sukhanov VB, Narmandakh P (1975) [Turtles of the group
Basilemys (Chelonia, Dermatemydidae) in Asia]. Trudy Sovmestnoi Sovetsko-Mongol'skoi Paleontologicheskoi Ekspeditsii 2: 94–101.
Sullivan RM (2006) Ah-shi-sle-pah Wilderness Study Area San Juan Basin, New Mexico; a paleontological and historical treasure and resource. New Mexico Museum of Natural History and Science Bulletin 34: 169–174.
Sullivan RM, Fowler DW (2011)
Navajodactylus boerei, n. gen., n. sp., (Pterosauria, ?Azhdarchidae) from the Upper Cretaceous Kirtland Formation (upper Campanian) of New Mexico. New Mexico Museum of Natural History and Science, Bulletin 53: 393–404.
Sullivan RM, Jasinski SE, Lucas SG (2013) Re-Assessment of Late Campanian (Kirtlandian) turtles from the Upper Cretaceous Fruitland and Kirtland Formations, San Juan Basin, New Mexico, USA. In: Brinkman DB, Holroyd PA, Gardner JD (Eds) Morphology and Evolution of Turtles. Springer, Dordrecht, 337–387. doi:
10.1007/978-94-007-4309-0_20Sullivan RM, Lucas SG, Jasinski SE (2011a) Preliminary observations on a skull of the amiid fish
Melvius, from the Upper Cretaceous Kirtland Formation, San Juan Basin, New Mexico. New Mexico Museum of Natural History and Science Bulletin 53: 475–483.
Sullivan RM, Lucas SG, Jasinski SE, Tanke DH (2011b) An unusual sacral neural spine osteopathy of a chasmosaurine (Dinosauria: Ceratopsidae) from the Upper Cretaceous Kirtland Formation (Hunter Wash member), San Juan Basin, New Mexico. Bulletin of the New Mexico Museum of Natural History and Science 53: 484–488.
Summesberger H, Machalski M, Wagreich M (2007) First record of the Late Campanian heteromorph ammonite
Nostoceras hyatti from the Alpine Cretaceous (Grünbach, Gosau Group, Lower Austria). Acta Geologica Polonica 57: 443–451.
Suslov YV (1982) [Ungual phalanges of dromaeosaurid from the Late Cretaceous deposits of Kzyl-Orda Province]. Materialy po Istorii Fauny i Flory Kazakhstana 8: 5–16.
Suzuki S, Narmandakh P (2004) Change of the Cretaceous turtle faunas in Mongolia. Hayashibara Museum of Natural Sciences Research Bulletin 2: 7–14.
Sweetman SC (2004) The first record of velociraptorine dinosaurs (Saurischia, Theropoda) from the Wealden (Early Cretaceous, Barremian) of southern England. Cretaceous Research 25: 353–364. doi:
10.1016/j.cretres.2004.01.004Sweetman SC (2006) A gobiconodontid (Mammalia, Eutriconodonta) from the Early Cretaceous (Barremian) Wessex Formation of the Isle of Wight, southern Britain. Palaeontology 49: 889–897. doi:
10.1111/j.1475-4983.2006.00564.xSweetman SC (2008) A spalacolestine spalacotheriid (Mammalia, Trechnotheria) from the Early Cretaceous (Barremian) of Southern England and its bearing on spalacotheriid evolution. Palaeontology 51: 1367–1385. doi:
10.1111/j.1475-4983.2008.00816.xSweetman SC (2009) A new species of the plagiaulacoid multituberculate mammal
Eobaatar from the Early Cretaceous of southern Britain. Acta Palaeontologica Polonica 54: 373–384. doi:
10.4202/app.2008.0003Sweetman SC, Gardner JD (2013) A new albanerpetontid amphibian from the Barremian (Early Cretaceous) Wessex Formation of the Isle of Wight, southern England. Acta Palaeontologica Polonica 58: 295–324.
Sweetman SC, Martill DM (2010) Pterosaurs of the Wessex Formation (Early Cretaceous, Barremian) of the Isle of Wight, southern England: a review with new data. Journal of Iberian Geology 36: 225–242. doi:
10.5209/rev_JIGE.2010.v36.n2.9Sweetman SC, Underwood CJ (2006) A neoselachian shark from the non-marine Wessex Formation (Wealden Group: Early Cretaceous, Barremian) of the Isle ofWight, southern England. Palaeontology 49: 457–465. doi:
10.1111/j.1475-4983.2006.00549.xSyromyatnikova EV (2011) Turtles of the genus
Ferganemys Nesov et Khosatzky, 1977 (Adocidae): shell morphology and phylogenetic position. Proceedings of the Zoological Institute of the Russian Academy of Sciences 315: 38–52.
Syromyatnikova EV, Danilov IG (2009) New material and a revision of turtles of the genus
Adocus (Adocidae) from the Late Cretaceous of Middle Asia and Kazakhstan. Proceedings of the Zoological Institute of the Russian Academy of Sciences 313: 74–94.
Syromyatnikova EV, Danilov IG (2013) New material and phylogenetic position of
Adocus bostobensis, a poorly known adocid turtle from the Late Cretaceous of Kazakhstan. Proceedings of the Zoological Institute RAS 317: 195–201.
Szalay FS, Sargis EJ (2006) Cretaceous therian tarsals and the metatherian-eutherian dichotomy. Journal of Mammalian Evolution 13: 171–210. doi:
10.1007/s10914-006-9024-4Szentesi Z, Gardner JD, Venczel M (2013) Albanerpetontid amphibians from the Late Cretaceous (Santonian) of Iharkút, Hungary, with remarks on regional differences in Late Cretaceous Laurasian amphibian assemblages. Canadian Journal of Earth Sciences 50: 268–281. doi:
10.1139/e2012-024Szentesi Z, Venczel M (2010) An advanced anuran from the Late Cretaceous (Santonian) of Hungary. Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen 256: 291–302. doi:
10.1127/0077-7749/2010/0054Szentesi Z, Venczel M (2012) A new discoglossid frog from the Upper Cretaceous (Santonian) of Hungary. Cretaceous Research 34: 327–333. doi:
10.1016/j.cretres.2011.11.012Tabaste N (1963) Étude de restes de poissons du Crétacé saharien. Mélanges ichthyologiques dédiés à la mémoire d'Achille Valenciennes, 1794–1865, coauteur de l'historie naturelle des poissons, 1828–1849. Mémoires de l'Institut français d'Afrique noire, Dakar, 437–485.
Tamura M (1979) Cenomanian bivalves from the Mifune Group, Japan. Part 3. Memoirs of the Faculty of Education, Kumamoto University, Natural Science 28: 59–74.
Tamura M, Okazaki Y, lkegami N (1991) [Occurrence of carnosaurian and herbivorous dinosaurs from Upper Formation of Mifune Group, Japan]. Memoirs of the Faculty of Education, Kumamoto University, Natural Science 40: 31–45.
Tanimoto M (2005) Mosasaur remains from the Upper Cretaceous Izumi Group of southwest Japan. Netherlands Journal of Geosciences 84: 373–378.
Tanimoto M (2010) Mosasaurs and the associated vertebrate faunas of Southwest Japan. In: Bardet N (Ed.) Third Mosasaur Meeting Schedule, Abstracts, Field Trip. Muséum national d'Histoire naturelle, Paris, 22.
Taverne L (2000) Nouvelles données ostéologiques et phylogénétiques sur
Palaeonotopterus greenwoodi, notoptéridé (Teleostei, Osteoglossomorpha) du Cénomanien inférieur continental (Crétacé) du Maroc. Stuttgarter Beiträge zur Naturkunde, Serie B 293: 1–24.
Taverne L, Maisey JG (1999) A notopterid skull (Teleostei, Osteoglossomorpha) from the continental early Cretaceous of Southern Morocco. American Museum Novitates 3260: 1–12.
Tong H, Buffetaut E (1996) A new genus and species of pleurodiran turtle from the Cretaceous of southern Morocco. Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen 199: 133–150.
Tong H, Gaffney ES, Buffetaut E (1998)
Foxemys, a new side-necked turtle (Bothremydidae: Pelomedusoides) from the Late Cretaceous of France. American Museum Novitates 3251: 1–19.
Tsaregradskii VA (1926) [Mosasaurs remains from Saratov Province and Ural Region]. Izvestiya Geologicheskogo Komiteta 45: 563–572.
Tsybin YI, Kurzanov SM (1979) [New data on Upper Cretaceous vertebrate localities of Bayshin Tsav region]. Trudy Sovmestnoi Sovetsko-Mongol'skoi Paleontologicheskoi Ekspeditsii 8: 108–112.
Tumanova TA (1987) [The armored dinosaurs of Mongolia]. Trudy Sovmestnoi Sovetsko-Mongol'skoi Paleontologicheskoi Ekspeditsii 32: 1–80.
Uliana MA, Dellape DA (1981) Estratigrafia y evolucion paleoambiental de la sucesion Maestrichtiano-Eoterciaria del engolfamiento Neuquino (Patagonia septentrional). Actas del Congreso Geologico Argentino. 673–711.
Uličný D (2001) Depositional systems and sequence stratigraphy of coarse-grained deltas in a shallow-marine, strike-slip setting: the Bohemian Cretaceous Basin, Czech Republic. Sedimentology 48: 599–628. doi:
10.1046/j.1365-3091.2001.00381.xUnwin DM (2001) An overview of the pterosaur assemblage from the Cambridge Greensand (Cretaceous) of Eastern England. Mitteilungen aus dem Museum fuer Naturkunde in Berlin, Geowissenschaftliche Reihe 4: 189–221.
Unwin DM (2003) On the phylogeny and evolutionary history of pterosaurs. In: Buffetaut E, Mazin J-M (Eds) Evolution and Palaeobiology of Pterosaurs. Geological Society Special Publication, n. 217, 139–190.
Unwin DM, Bakhurina NN (2000) Pterosaurs from Russia, Middle Asia and Mongolia. In: Benton MJ, Shishkin MA, Unwin DM, Kurochkin EN (Eds) The Age of Dinosaurs in Russia and Mongolia. Cambridge University Press, Cambridge, 420–433.
Unwin DM, Bakhurina NN, Lockley MG, Manabe M, Lü J (1997) Pterosaurs from Asia. Paleontological Society of Korea Special Publication 2: 43–65. doi; 10.1080/08912969709386563
Unwin DM, Lü J (1997) On
Zhejiangopterus and the relationships of pterodactyloid pterosaurs. Historical Biology 12: 199–210.
Unwin DM, Lü J, Bakhurina NN (2000) On the systematic and stratigraphic significance of pterosaurs from the Lower Cretaceous Yixian Formation (Jehol Group) of Liaoning, China. Mitteilungen aus dem Museum fuer Naturkunde in Berlin, Geowissenschaftliche Reihe 3: 181–206.
Unwin DM, Martill DM (2007) Pterosaurs of the Crato Formation. In: Martill DM, Bechly G, Loveridge RF (Eds) The Crato Fossil Beds of Brazil: Window into an Ancient World. Cambridge University Press, Cambridge, 475–524. doi:
10.1017/CBO9780511535512.018Vajda V, Raine JI (2010) A palynological investigation of plesiosaur-bearing rocks from the Upper Cretaceous Tahora Formation, Mangahouanga, New Zealand. Alcheringa 34: 359–374. doi:
10.1080/03115518.2010.486642Varricchio DD (1993) Montana climatic changes associated with the Cretaceous Claggett and Bearpaw transgressions. In: Hunter LDV (Ed.) Montana Geological Society: Energy and Mineral Resources of Central Montana; 1993 Field Conference Guidebook Montana Geological Society, Billings, 97–104.
Vavrek MJ, Larsson HCE (2010) Low beta diversity of Maastrichtian dinosaurs of North America. Proceedings of the National Academy of Sciences USA 107: 8265–8268. doi:
10.1073/pnas.0913645107Vega FJ, Nyborg T, Kovalchuk G, Luque J, Rojas-Briceño A, Patarroyo P, Porras-Múzquiz H, Armstrong A, Bermúdez H, Garibay L (2010) On some Panamerican Cretaceous crabs (Decapoda: Raninoida). Boletín de la Sociedad Geológica Mexicana 62: 263–279.
Veldmeijer AJ, Signore M, Meijer HJM (2005) Description of two pterosaur (Pterodactyloidea) mandibles from the Lower Cretaceous Santana Formation, Brazil. Deinsea 11: 67–86.
Verzilin NN (1963) [Cretaceous deposits of North Fergana Depression and their oil and gas resources]. Trudy Leningradskogo Obshchestva Estestvoispytatelei 70: 1–220.
Vitek NS, Danilov IG (2010) New material and a reassessment of soft-shelled turtles (Trionychidae) from the Late Cretaceous of Middle Asia and Kazakhstan. Journal of Vertebrate Paleontology 30: 383–393. doi:
10.1080/02724631003617548Vitek NS, Danilov IG (2013) Soft-shelled turtles (Trionychidae) from the Bissekty Formation (Upper Cretaceous: Turonian) of Uzbekistan: Skull-based taxa and probable skull-shell associations. Cretaceous Research 43: 48–58. doI: 10.1016/j.cretres.2013.02.009
Vitek NS, Danilov IG (2014) Soft-shelled turtles (Trionychidae) from the Cenomanian of Uzbekistan. Cretaceous Research 49: 1–12.
Vremir M (2010) New faunal elements from the Late Cretaceous (Maastrichtian) continental deposits of Sebes¸ area (Transylvania). Terra Sebus, Acta Musei Sabesiensis 2: 635–684.
Vremir M, Dyke GJ, Csiki Z (2011) Late Cretaceous pterosaurian diversity in the Transylvanian and Hateg basins (Romania): new results. Eighth Romanian Symposium on Palaeontology, Abstract Volume. Bucharest, 131–132.
Vremir M, Kellner AWA, Naish D, Dyke GJ (2013) A new azhdarchid pterosaur from the Late Cretaceous of the Transylvanian Basin, Romania: Implications for azhdarchid diversity and distribution. PLoS One 8: e54268. doi:
10.1371/journal.pone.0054268Vyalov OS (1945a) [Scheme of division of Cretaceous deposits of Fergana]. Doklady Akademii Nauk SSSR 49: 127–131.
Vyalov OS (1945b) [Types of Cretaceous sections of Fergana]. Doklady Akademii Nauk SSSR 49: 285–288.
Wagreich M, Faupl P (1994) Palaeogeography and geodynamic evolution of the Gosau Group of the Northern Calcareous Alps (Late Cretaceous, Eastern Alps, Austria). Palaeogeography, Palaeoclimatology, Palaeoecology 110: 235–254. doi:
10.1016/0031-0182(94)90086-8Wagreich M, Marschalko R (1995) Late Cretaceous to early Tertiary palaeogeography of the Western Carpathians (Slovakia) and the Eastern Alps (Austria); implications from heavy mineral data. Geologische Rundschau 84: 187–199. doi:
10.1007/BF00192250Walker CA, Buffetaut E, Dyke GJ (2007) Large euenantiornithine birds from the Cretaceous of southern France, North America and Argentina. Geological Magazine 144: 977–986. doi:
10.1017/S0016756807003871Wang X, Kellner AWA, Jiang S, Meng X (2009) An unusual long-tailed pterosaur with elongated neck from western Liaoning of China. Anais da Academia Brasileira de Ciências 81: 793–812. doi:
10.1590/S0001-37652009000400016Wang X, Kellner AWA, Zhou Z, Campos DdA (2005) Pterosaur diversity and faunal turnover in Cretaceous terrestrial ecosystems in China. Nature 437: 875–879. doi:
10.1038/nature03982Wang X, Zhou Z (2003) Two new pterodactyloid pterosaurs from the Early Cretaceous Jiufotang Formation of western Liaoning, China. Vertebrata PalAsiatica 41: 34–41.
Watabe M, Tsuihiji T, Suzuki D, Tsogtbaatar K (2009) The first discovery of pterosaurs from the Upper Cretaceous of Mongolia. Acta Palaeontologica Polonica 54: 231–242. doi:
10.4202/app.2006.0068Weishampel DB, Barrett PM, Coria RA, Le Loeuff J, Xu X, Zhao X-J, Sahni A, Gomani EM, Noto CR (2004) Dinosaur distribution. In: Weishampel DB, Dodson P, Osmolska H (Eds) The Dinosauria Second Edition. University of California Press, Berkeley, Los Angeles, London, 517–606. doi:
10.1525/california/9780520242098.003.0027Weishampel DB, Norman DB, Grigorescu D (1991) The dinosaurs of Transylvania: island biogeography in the Late Cretaceous. National Geographic Research and Exploration 7: 196–215.
Wellnhofer P (1980) Flugsaurierreste aus der Gosau-Kreide von Muthmannsdorf (Niederosterreich) - ein Beitrag zur Kierfermechanik der Pterosaurier. Mitteilungen der Bayerischen Staatssammlung fuer Palaeontologie und historische Geologie 20: 95–112.
Wellnhofer P (1985) Neue Pterosaurier aus der Santana-Formation (Apt) der Chapada do Araripe, Brasilien. Palaeontographica, Abteilung A 187: 105–182.
Wellnhofer P (1991) The Illustrated Encyclopedia of Pterosaurs. Salamander Books, London, 192 pp.
Wellnhofer P, Buffetaut E (1999) Pterosaur remains from the Cretaceous of Morocco. Palaeontologische Zeitschrift 73: 133–142. doi:
10.1007/BF02987987Welton BJ, Farish RF (1993) The Collector’s Guide to Fossil Sharks and Rays from the Cretaceous of Texas. Before Time, Lewisville, Texas, 204 pp.
Wenz S (1981) Un coelacanthe géant,
Mawsonia lavocati Tabaste, de l`Albien–base du Cénomanien du Sud marocain. Annales de Paléontologie (Vertébrés) 67: 1–20.
Wick SL, Lehman TM (2013) A new ceratopsian dinosaur from the Javelina Formation (Maastrichtian) of West Texas and implications for chasmosaurine phylogeny. Naturwissenschaften 100: 667–682. doi:
10.1007/s00114-013-1063-0Wiese F, Cech S, Ekrt B, Kost'ak M, Mazuch M, Voigt S (2004) The Upper Turonian of the Bohemian Cretaceous Basin (Czech Republic) exemplified by the Upohlavy working quarry: integrated stratigraphy and palaeoceanography of a gateway to the Tethys. Cretaceous Research 25: 329–352. doi:
10.1016/j.cretres.2004.01.003Wiffen J (1980)
Moanasaurus, a new genus of marine reptile (Family Mosasauridae) from the Upper Cretaceous of North Island, New Zealand. New Zealand Journal of Geology and Geophysics 23: 507–528. doi:
10.1080/00288306.1980.10424122Wiffen J (1981) The first Late Cretaceous turtles from New Zealand. New Zealand Journal of Geology and Geophysics 24: 293–299. doi:
10.1080/00288306.1981.10422718Wiffen J (1983) The first record of
Pachyrhizodus caninus Cope (Order Clupeiformes) from the Late Cretaceous of New Zealand. New Zealand Journal of Geology and Geophysics 26: 109–119. doi:
10.1080/00288306.1983.10421527Wiffen J (1986) Flying reptiles from the Late Cretaceous. Geological Society of New Zealand Miscellaneous Publications 35A: 113. doi:
10.1080/00288306.1986.10427535Wiffen J, Moisley WL (1986) Late Cretaceous reptiles (Families Elasmosauridae and Pliosauridae) from the Mangahouanga Stream, North Island, New Zealand. New Zealand Journal of Geology and Geophysics 29: 205–252.
Wilson AE, Newbrey MG, Brinkman DB, Cook TD, Neuman AG (2013) Age and growth in
Myledaphus bipartitus, a Late Cretaceous freshwater guitarfish from Alberta, Canada. Canadian Journal of Earth Sciences 50: 930–944. doi:
10.1139/cjes-2013-0001Wilson JA, Zalmout IS (2006) Latest Cretaceous reptiles from the Hashemite Kingdom of Jordan. In: Barrett PM, Evans SE (Eds) 9th International Symposium on Mesozoic Terrestrial Ecosystems and Biota Abstracts and Proceedings Volume. Manchester, 183.
Wilson MVH, Brinkman DB, Neuman AG (1992) Cretaceous Esocoidei (Teleostei): early radiation of the pikes in North American fresh waters. Journal of Paleontology 66: 839–846.
Winkler DA, Murry PA, Jacobs LL (1990) Early Cretaceous (Comanchean) vertebrates of central Texas. Journal of Vertebrate Paleontology 10: 95–116. doi:
10.1080/02724634.1990.10011794Witmer LM, Chatterjee S, Franzosa J, Rowe TB (2003) Neuroanatomy of flying reptiles and implications for flight, posture and behaviour. Nature 425: 950–953. doi:
10.1038/nature02048Witton MP (2009) A new species of
Tupuxuara (Thalassodromidae, Azhdarchoidea) from the Lower Cretaceous Santana Formation of Brazil, with a note on the nomenclature of Thalassodromidae. Cretaceous Research 30: 1293–1300. doi:
10.1016/j.cretres.2009.07.006Witton MP (2013) Pterosaurs: Natural History, Evolution, Anatomy. Princeton University Press, Princeton and Oxford, pp.
Witton MP, Martill DM, Green M (2009) On pterodactyloid diversity in the British Wealden (Lower Cretaceous) and a reappraisal of “
Palaeornis”
cliftii Mantell, 1844. Cretaceous Research 30: 676–686. doi:
10.1016/j.cretres.2008.12.004Witton MP, Martill DM, Loveridge RF (2010) Clipping the wings of giant pterosaurs: Comments on wingspan estimations and diversity. Acta Geoscientica Sinica 31: 79–81.
Witton MP, Naish D (2008) A reappraisal of azhdarchid pterosaur functional morphology and paleoecology. PLoS One 3: e2271. doi:
10.1371/journal.pone.0002271Witton MP, Naish D (In press) Azhdarchid pterosaurs: water-trawling pelican mimics or "terrestrial stalkers"? Acta Palaeontologica Polonica
Wueringer BE, Squire L Jr., Collin SP (2009) The biology of extinct and extant sawfish (Batoidea: Sclerorhynchidae and Pristidae). Reviews in Fish Biology and Fisheries 19: 445–464. doi:
10.1007/s11160-009-9112-7Yang S-Y, Lockley MG, Lim S-K, Chun SS (1997) Cretaceous bird tracks of Korea. Journal of the Paleontological Society of Korea Special Publication 2: 33–42.
Zanno LE (2010) A taxonomic and phylogenetic re-evaluation of Therizinosauria (Dinosauria: Maniraptora). Journal of Systematic Palaeontology 8: 503–543. doi:
10.1080/14772019.2010.488045Zelenkov NV, Averianov AO (2011) Synsacrum of a primitive bird from the Upper Cretaceous of Uzbekistan. Paleontological Journal 45: 314–319. doi:
10.1134/S0031030111030166Zhou C-F (2010) New material of
Chaoyangopterus (Pterosauria: Pterodactyloidea) from the Early Cretaceous Jiufotang Formation of western Liaoning, China. Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen 257: 341–350. doi:
10.1127/0077-7749/2010/0081Zhou Z, Barrett PM, Hilton J (2003) An exceptionally preserved Lower Cretaceous ecosystem. Nature 421: 807–814. doi:
10.1038/nature01420