





Citation: Masek M, Bocak L (2014) The taxonomy and diversity of Platerodrilus (Coleoptera, Lycidae) inferred from molecular data and morphology of adults and larvae. ZooKeys 426: 29–63. doi: 10.3897/zookeys.426.7398
The Oriental neotenic net-winged beetles attracted attention of biologists due to conspicuous large-bodied females; nevertheless phylogenetic relationships remain contentious and only a few species are known in both the fully metamorphosed males and neotenic females. The phylogenetic analyses and morphology of larvae and adults provide data for investigation of relationships and species delineation. Platrilus Kazantsev, 2009, Platerodriloplesius Wittmer, 1944, and Falsocalochromus Pic, 1942 are synonymized to Platerodrilus Pic, 1921. Platrilus hirtus (Wittmer, 1938) and Pl. crassicornis (Pic, 1923) are transferred to Platerodrilus Pic, 1921. Platerodrilus hoiseni Wong, 1996 is proposed as a junior subjective synonym of Falsocalochromus ruficollis Pic, 1942. Platerodrilus is divided in three species-groups: P. paradoxus, P. major, and P. sinuatus groups defined based on the shape of genitalia and molecular phylogeny. The following species are described: Platerodrilus foliaceus sp. n., P. wongi sp. n. (P. paradoxus group); P. ngi sp. n., P. wittmeri (P. major group), P. ijenensis sp. n., P. luteus sp. n., P. maninjauensis sp. n., P. montanus sp. n., P. palawanensis sp. n., P. ranauensis sp. n., P. sibayakensis sp. n., P. sinabungensis sp. n., P. talamauensis sp. n., and P. tujuhensis sp. n. (P. sinuatus group). P. korinchiana robinsoni Blair, 1928 is elevated to the species rank as P. robinsoni Blair, 1928, stat. n. The conspecific semaphoronts are identified using molecular phylogeny for P. foliaceus sp. n., P. tujuhensis sp. n., P. montanus sp. n., P. maninjauensis sp. n.; additional female larvae are assigned to the species-groups. Diagnostic characters are illustrated and keys are provided for P. paradoxus and P. major groups.
Oriental Region, net-winged beetles, morphology, molecular phylogeny, taxonomy
The platerodriline net-winged beetles are one of elateroid lineages with modified female morphology (
The taxonomy of the neotenic lineages has quite short history despite the fact that the trilobite larvae were first time described in the 19th century (
The DNA data represent an independent source of information for species delineation (
Adult males and female larvae were used for morphological descriptions. A part of specimens used for morphological study was sequenced and labelled with the GenBank voucher numbers in the format UPOL + six-letter/number code and the status of all type specimens were designated with red labels (
Taxonomic coverage, locality data and GenBank accession numbers.
| Species | Voucher UPOL+ | Local. data | rrnL |
|---|---|---|---|
| Outgroup | |||
| Benibotarus nigripennis | 000572 | Japan | DQ181001 |
| Benibotarus spinicoxis | 000573 | Japan | DQ181002 |
| Dictyoptera elegans | 000570 | Japan | DQ181375 |
| Dictyoptera speciosa | 000571 | Japan | DQ181000 |
| Libnetis granicollis | 001012 | Japan | DQ181033 |
| Libnetis sp. | 001002 | Sumatra | DQ181030 |
| Libnetis sp. | 001008 | Malaysia | DQ181031 |
| Libnetis sp. | 000L02 | Sabah | DQ180964 |
| Lycoprogenthes sp. | 000801 | Sumatra | DQ181021 |
| Lycoprogenthes sp. | 000802 | Java | DQ181022 |
| Lycoprogenthes sp. | 000805 | Sumatra | DQ181023 |
| Lycoprogenthes sp. | 000358 | Java | DQ180996 |
| Pyropterus nigroruber | 000574 | Japan | DQ181003 |
| Lyropaeus sp. | VP0016 | India | KC736885 |
| Lyropaeus sp. | VP0017 | India | KC736886 |
| Lyropaeus sp. | VP2312 | India | KC736887 |
| Lyropaeus dominator | VP0003 | Malaysia | KC736882 |
| Lyropaeus optabilis | VP0004 | Malaysia | KC736883 |
| Lyropaeus optabilis | 000585 | Malaysia | DQ181014 |
| Lyropaeus ritsemae | VP0001 | Sumatra | KC736880 |
| Lyropaeus ritsemae | VP0006 | Sumatra | KC736884 |
| Lyropaeus rubrostriatus | 000L11 | Malaysia | DQ180968 |
| Lyropaeus waterhousei | VP0002 | Sumatra | KC736881 |
| Lyropaeus waterhousei | 000584 | Sumatra | DQ181013 |
| Ingroup | |||
| Horakiella emasensis | 001043 | Malaysia | DQ181036 |
| Macrolibnetis depressus | VP0050 | Malaysia | KF802467 |
| Macrolibnetis depressus | 000L21 | Malaysia | DQ180976 |
| Pendola sp. | 000M45 | Java | DQ180984 |
| Platerodrilini gen. sp. | VP0009 | Malaysia | KF802457 |
| Platerodrilini gen. sp. | VP0010 | Sumatra | KF802480 |
| Platerodrilini gen. sp. | VP0012 | Malaysia | KF802458 |
| Platerodrilini gen. sp. | VP0030 | India | KF802462 |
| Platerodrilini gen. sp. | VP0031 | India | KF802463 |
| Platerodrilini gen. sp. | VP0034 | India | KF802464 |
| Platerodrilus curtus | 001380 | Mindanao | KF625997 |
| Platerodrilus curtus | 001381 | Mindanao | KF626073 |
| Platerodrilus curtus | 001383 | Mindanao | KF626074 |
| Platerodrilus curtus | VP0014 | Mindanao | KF802459 |
| Platerodrilus curtus | VP2316 | Mindanao | KF802479 |
| Platerodrilus angustatus | 001388 | Sumatra | KF626001 |
| Platerodrilus corporaali | 001373 | Sumatra | KF625991 |
| Platerodrilus foliaceus | 000588 | Borneo | DQ181017 |
| Platerodrilus foliaceus | 000589 | Borneo | EF143214 |
| Platerodrilus ijenensis | 000586 | Java | DQ181015 |
| Platerodrilus luteus | 001379 | Sumatra | KF625996 |
| Platerodrilus major | 001387 | Sumatra | KF626000 |
| Platerodrilus maninjauensis | 001374 | Sumatra | KF625992 |
| Platerodrilus maninjauensis | 001377 | Sumatra | KF625994 |
| Platerodrilus maninjauensis | 001386 | Sumatra | KF626075 |
| Platerodrilus maninjauensis | VP2303 | Sumatra | KF802470 |
| Platerodrilus maninjauensis | VP2306 | Sumatra | KF802473 |
| Platerodrilus montanus | 001371 | Sumatra | KF625989 |
| Platerodrilus montanus | VP2308 | Sumatra | KF802475 |
| Platerodrilus ngi | VP0021 | Singapore | KF802461 |
| Platerodrilus ranauensis | 000587 | Sumatra | DQ181016 |
| Platerodrilus robinsoni | 001378 | Sumatra | KF625995 |
| Platerodrilus sibayakensis | 001372 | Sumatra | KF625990 |
| Platerodrilus sibayakensis | 001389 | Sumatra | KF802552 |
| Platerodrilus sp. | 000L01 | Sabah | DQ180963 |
| Platerodrilus sp. | VP0044 | Sabah | KF802465 |
| Platerodrilus sp. | VP2301 | Sabah | KF802468 |
| Platerodrilus sp. | VP0020 | Malaysia | KF802460 |
| Platerodrilus sp. | VP0047 | Sumatra | KF802466 |
| Platerodrilus sp. | VP2302 | Malaysia | KF802469 |
| Platerodrilus sp. | VP2304 | Thailand | KF802471 |
| Platerodrilus sp. | VP2307 | Sumatra | KF802474 |
| Platerodrilus sp. | VP2309 | Malaysia | KF802476 |
| Platerodrilus sp. | VP2310 | Malaysia | KF802477 |
| Platerodrilus sp. | VP2311 | Laos | KF802478 |
| Platerodrilus sp. | MB1382 | Palawan | EF625998 |
| Platerodrilus talamauensis | 001375 | Sumatra | KF626072 |
| Platerodrilus talamauensis | 001376 | Sumatra | KF625993 |
| Platerodrilus tujuhensis | 001385 | Sumatra | KF625999 |
| Platerodrilus tujuhensis | VP2305 | Sumatra | KF802472 |
Abbreviations and depositories. Descriptions: BL–length of body; WH–width at humeri; PL–length of pronotum; PW–width of pronotum; Ediam–maximum eye diameter; Edist–minimum interocular distance in frontal part of cranium. Depositories: LMBC–Voucher collection of the Laboratory of Molecular Systematics, Faculty of Science UP, Olomouc; BMNH–Natural History Museum, London; MNHP–Museum d'histoire naturelle, Paris; ZRCS–Zoological Reference Collection, Raffles Museum of Biodiversity Research, NUS; KMTC–Kiyoshi Matsuda Collection, Takarazuka.
Total DNA was extracted using Wizard SV96 kit (Promega Inc.) and primers 16a (5'-CGCCTGTTTAACAAAAACAT-3'), 16b (5'-CCGGTCTGAACTCAGATCATGT-3') and ND1A (5'-GGTCCCTTACGAATTTGAATATATCCT-3') were used for PCR amplification of the 530–810 base pairs of rrnL, which showed the best results in identification of immature stages (
Sequences were edited using the Sequencher 4.8 software package (Gene Codes Corp.). The rrnL mtDNA fragment was aligned using ClustalW 1.83 (
The ultrametric tree was produced from the tree depicted in Fig. 1 using r8s software (
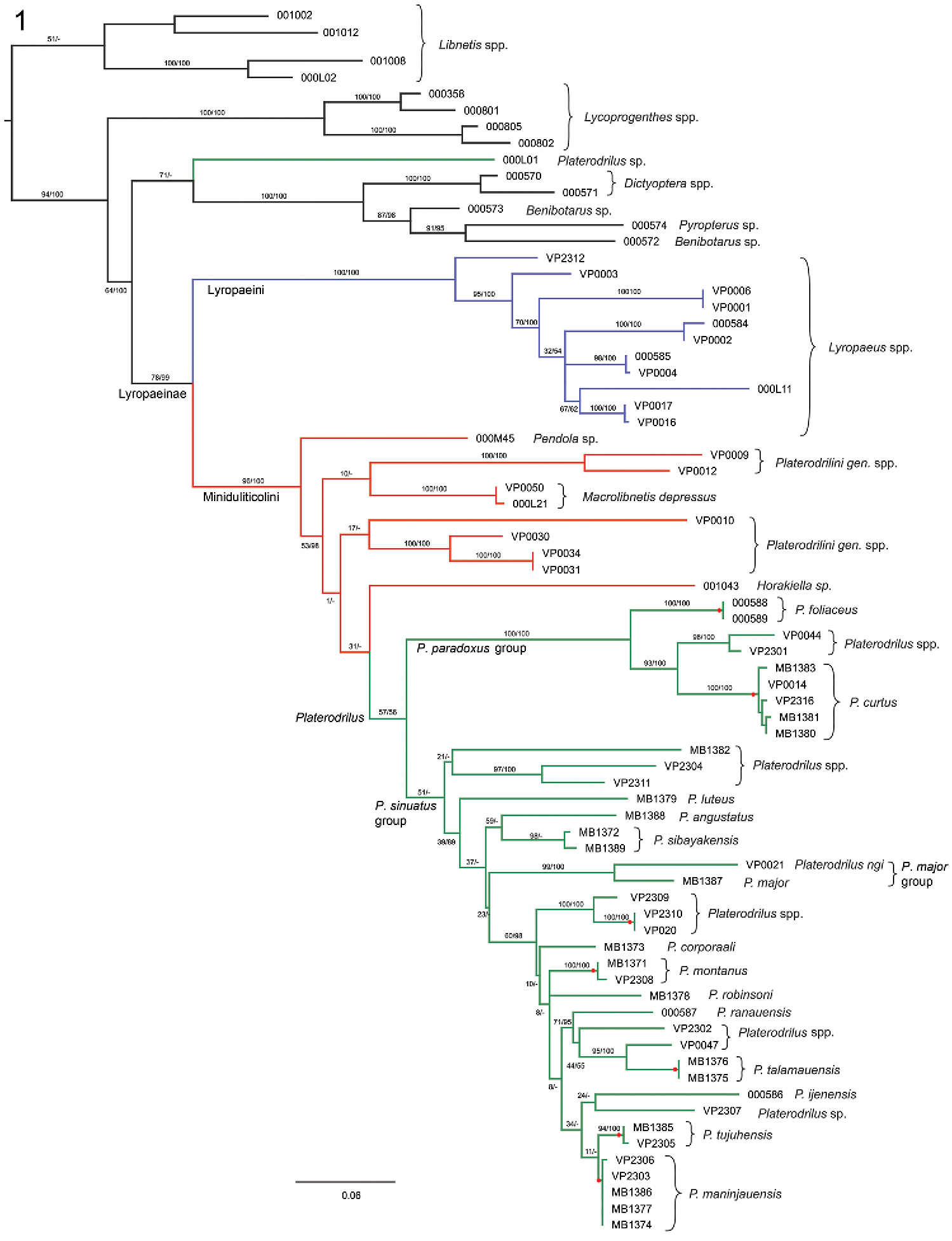
Phylogenetic hypothesis for Platerodrilus Pic, 1921 based on a maximum likelihood analysis of the Muscle alignment. Numbers at the branches are maximum likelihood bootstrap values and Bayesian posterior probabilities. The red dots designate GMYC species clusters.
The DNA sequences of rrnL were produced for 73 specimens. The dataset of aligned rrnL sequences contained 530–723 homologous positions depending on the applied alignment procedure; 253–267 characters were parsimony informative. The topologies produced from BlastAlign, Muscle and Clustal alignments analyzed under maximum likelihood method and Bayesian inference identified the same strongly supported principal clades of Lyropaeinae and although topologies differed somewhat with respect to the deeper nodes of Miniduliticolini, all analyses recovered Pendola, Macrolibnetis, Horakiella and related taxa as deeper splits of Miniduliticolini and Platerodrilus as a terminal lineage (Fig. 1). One terminal, Platerodrilus sp. 000L01 was recovered outside Lyropaeinae and we consider its position as an artefact of the single marker analysis. The species was found in relationships to another Platerodrilus in the six-gene analysis of Lycidae (
The Platerodrilus paradoxus and Platerodrilus major clades were well supported in all analyses except by analysis of the Clustal alignment (BS 100%, PP 52–100% and BS 97–99%, PP 52–100%, respectively). The Platerodrilus sinuatus group (including the nested Platerodrilus major group) obtained much lower support (BS 46–56%, PP < 89%). The species level clusters and relationships of closely related species were regularly well supported (Fig. 1).
The GMYC analysis of the normalized tree was used as an independent test for morphology based delineation of species. The analysis suggested the clusters designated as Platerodrilus tujuhensis and Platerodrilus maninjauensis (uncorrected genetic distance 1.4%) as separate species and these are well supported also by morphological differences (see Taxonomy). Similarly, two separate species were inferred for two specimens of Platerodrilus sibayakensis (uncorrected genetic distance 0.8%) and these do not differ in any morphological character and were collected in the same region.
The origin of large-bodied neotenic larvae was recovered in three unrelated lineages: Lyropaeus, Macrolibnetis depressus + unidentified species from India and Platerodrilus (Fig. 1). Males and female larvae of four species, i.e. Platerodrilus foliaceus, Platerodrilus maninjauensis, Platerodrilus montanus and Platerodrilus tujuhensis clustered with conspecific males in clades with very high bootstrap support. Additionally, a number of larvae was assigned to the species groups in relationships to the previously described species (Fig. 1). We found that the species of the Platerodrilus paradoxus clade share pronotum without glabrous prominent tubercles (Figs 1, 34, 40). Similarly, the species of the Platerodrilus sinuatus group from continental Asia (Fig. 1) have smooth terga (VP2304, VP2311). The glabrous tubercles in the discs of the thoracic terga are present only in the lineage of Sumatran and Malay species of the Platerodrilus sinuatus group (Fig. 1; terminals VP2308, VP2302, VP2307, VP0047 etc.). The robust, vermiform larva (Fig. 43) clustered with species of the Platerodrilus major clade (Fig. 1).
The results confirm that Lyropaeini (i.e., Lyropaeus Waterhouse, 1878 sensu lato) is an independent lineage with the large-bodied neotenic females and Platerodrilus belong to a sister-clade of Lyropaeini along with Macrolibnetis, Horakiella, and Pendola (Fig. 1). Most of these genera are also known only from males but no large bodied female has been assigned to them and their females are probably larviform but similar in body size to males. The only miniduliticoline taxon with the large-bodied female except Platerodrilus is Macrolibnetis (
The subfamiliar and tribal classification of Platerodrilus and related genera has been ambiguous.
Platerodrilus Pic, 1921, Duliticola Mjöberg, 1925, Macrolibnetis Pic, 1938, Platerodriloplesius Wittmer, 1941, Falsocalochromus Pic, 1942 and Platrilus Kazantsev, 2009 are available genus-group names referring to the platerodriline net-winged beetles with large-bodied neotenic females occurring in South East Asia (Fig. 2). The current results confirm that Macrolibnetis Pic, 1938 represents a distant lineage and does not belong to the Platerodrilus clade [considered as a synonym of Platerodrilus by

2 Female larvae of Platerodrilus. 3 Platerodrilus sp. from Gn. Sinabung, Sumatra, ditto from Gn. Apo, Mindanao.
The other Miniduliticolini with large-bodied neotenic females form a clade designated as Platerodrilus in Fig. 1. The deepest split of Platerodrilus consists of a clade of Platerodrilus curtus, Platerodrilus foliaceus and several unidentified larvae (Figs 37–38). Their male genitalia (Figs 44–47) resemble those of Platerodrilus paradoxus (see

Adult male, general appearance: 4 Platerodrilus foliaceus 5 Platerodrilus wongi 6 Platerodrilus robinsoni 7 Platerodrilus maninjauensis 8 Platerodrilus luteus 9 Platerodrilus ranauensis 10 Platerodrilus sibayakensis 11 Platerodrilus sinabungensis 12 Platerodrilus tujuhensis 13 Platerodrilus montanus 14 Platerodrilus ijenensis 15 Platerodrilus talamauensis 16 Platerodrilus palawanensis 17 Platerodrilus wittmeri. Scales 2 mm.
The sister clade to the Platerodrilus paradoxus clade contains Platerodrilus species with two types of male genitalia: (a) the phallus with short, densely pubescent parameres (Figs 48–51) and (b) the phallus slender, curved, parameres with long membranous apical process (Figs 52–72). We designate these groups as Platerodrilus major and Platerodrilus sinuatus groups (Fig. 1). The Platerodrilus major group (Figs 1, 17, 30, 48–51) contains species placed in Platrilus by
The Platerodrilus sinuatus group contains species from continental Asia, which form a deep split (terminals VP2304, VP2311) and further a group of species from the Sundaland and Palawan (Fig. 1). Larvae from continental Asia do not have any tubercles in the thoracic terga, similarly to the Platerodrilus paradoxus and Platerodrilus major group, only the species from Sumatra, Java and Malay Peninsula have the glabrous tubercles both in the disc and posterior margins of thoracic terga (Figs 32–33, 39). The males of these species can be assigned to continental or Sundaland lineages only with DNA data. Therefore, we propose to group them in the Platerodrilus sinuatus group despite paraphyly of the assemblage.

Male pronotum of Platerodrilus. 18 Platerodrilus ijenensis 19 Platerodrilus robinsoni 20 Platerodrilus maninjauensis 21 Platerodrilus montanus 22 Platerodrilus foliaceus 23 Platerodrilus luteus 24 Platerodrilus ranauensis 25 Platerodrilus sibayakensis 26 Platerodrilus sinabungensis 27 Platerodrilus tujuhensis 28 Platerodrilus wongi 29 Platerodrilus talamauensis 30 Platerodrilus wittmeri 31 Platerodrilus palawanensis. Larva, general appearance: 32 Platerodrilus maninjauensis 33 Platerodrilus montanus 34 Platerodrilus paradoxus. Scales 0.5 mm (Figs 18–31); Scales 5 mm (32–34).

Larvae of Platerodrilus and Macrolibnetis: 35–38 Platerodrilus spp. 39 Platerodrilus ruficollis 40 Platerodrilus foliaceus 41 Platerodrilus tujuhensis 42 Macrolibnetis depressus 43 Platerodrilus ngi. Scales 5 mm.
Further two genus-group names were proposed for species now placed in Platerodrilus. Platerodriloplesius was erected for taxa with flabellate male antennae (

Male genitalia of Platerodrilus: 44–45 Platerodrilus wongi 46–47 Platerodrilus foliaceus 48–49 Platerodrilus major 50–51 Platerodrilus wittmeri 52–53 Platerodrilus talamauensis 54–55 Platerodrilus ranauensis. Scales 0.25 mm.
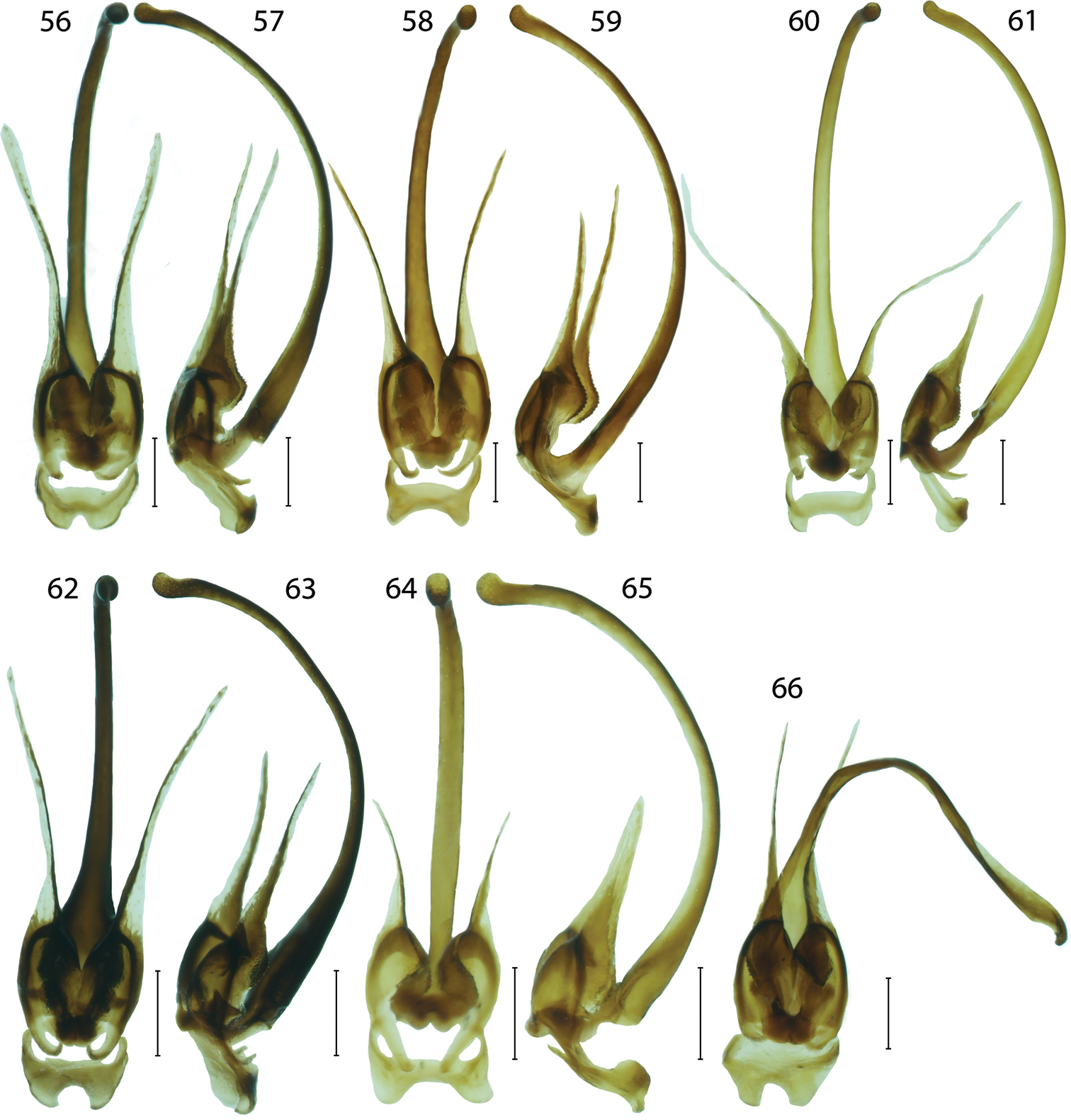
Male genitalia of Platerodrilus: 56–57 Platerodrilus tujuhensis 58–59 Platerodrilus sinabungensis 60–61 Platerodrilus maninjauensis 62–63 Platerodrilus robinsoni 64–65 Platerodrilus ijenensis 66 Platerodrilus montanus. Scales 0.25 mm.
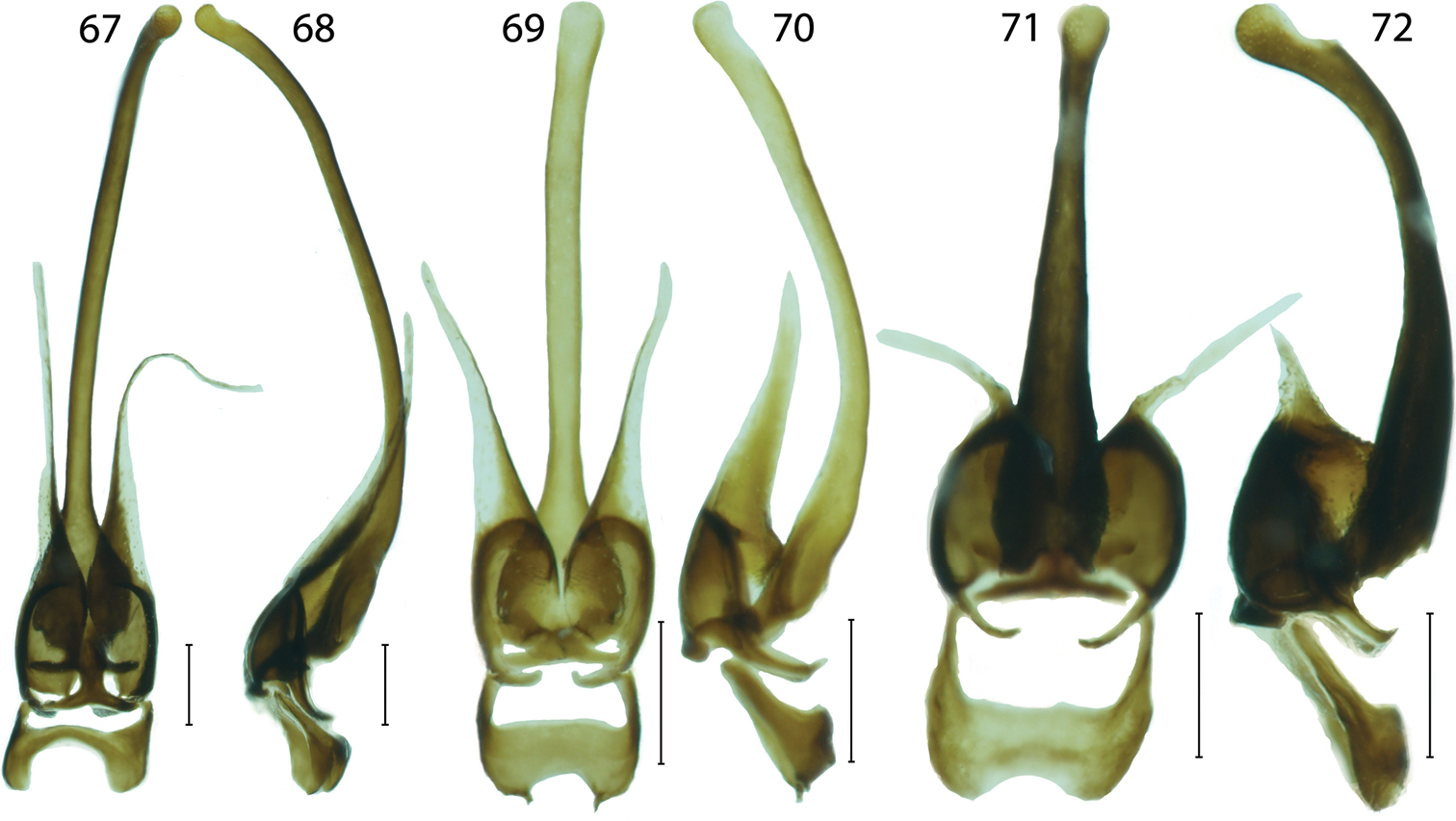
Male genitalia of Platerodrilus: 67–68 Platerodrilus luteus 69–70 Platerodrilus palawanensis 71–72 Platerodrilus sibayakensis. Scales 0.25 mm.
To sum up, we propose to consider Duliticola, Platerodriloplesius, Platrilus and Falsocalochromus to be junior synonyms of Platerodrilus. Platerodriloplesius represents a polyphyletic assemblage and Platrilus is a terminal branch supported by unique apomorphies rendering Platerodrilus in a paraphylum. As these taxa cannot be assigned to species groups without dissection of male genitalia, the proposed generic classification results in a definition of an easily recognisable monophyletic assemblage.
Rapid morphological divergence in male genitalia is widespread and results largely from sexual selection (
Diagnosis. Platerodrilus differs from most Miniduliticolini in the stout body (6–11 mm) and characteristic types of male genitalia (Figs 44–72). The morphologically similar Pendola has genitalia resembling those of Lyropaeus (
Description. Male. Body 5.8–10.8 mm, flat, slightly widened posteriorly, densely pubescent. Pronotum and elytra bicoloured, uniformly yellow or black (Figs 4–17).
Head small, prognathous to slightly hypognathous, partly retracted in pronotum. Eyes hemispherically prominent, frontal interocular distance longer than maximum eye diameter. Labrum sclerotized, transverse, separated from clypeus, mandibles slender, long, slightly curved, incisor margin simple, without teeth, maxilla tiny, with setose mala, stipes plate-like, cardo vestigial, palpifer short, maxillary palpi 4-segmented, palpomere 1 shortest, about twice longer than palpifer, palpomere 2 longest, slender, palpomere 3 slightly longer than wide, apical palpomere slender, drop-like, with slender apical part. Labium reduced, mentum plate-like, formed by single sclerite, ligula absent, palpi 3-segmented, basal palpomeres subequal, rectangular to slightly longer than wide, apical palpomere twice longer than wide at base, pointed to apex.
Antennal tubercles present, usually strongly prominent. Antennae 11-segmented, slightly surpassing middle of elytral length, dark coloured, never with apical antennomeres pale, antennae usually weakly serrate, a few species with flabellate antennae. Scapus pear-like, robust, pedicel and antennomere 3 subequal in length, antennomeres of serrate antennae flattened, from antennomere 4 gradually slenderer, apical antennomere long, parallel-sided. Flabellate antennae with lamellae of antennomeres 3–10 longer than body of antennomere; whole antennae with dense erected pubescence.
Pronotum transverse with prominent to obtuse anterior angles (Figs 18–31), without carinae. Lateral margins elevated, straight to widely rounded, frontal angles sometimes inconspicuous, posterior angles mostly sharp, posterior margin bisinuate. Pronotum with deep depression along lateral margins, with sparse, long, erected pubescence. Scutellum longer than wide, triangular, simply rounded at apex. Elytra flat, slightly widened posteriorly, with well marked humeri; elytral costae inconspicuous, only costa 2 and 4 traceable in whole elytra, costa 4 forming humeral edge, other costae apparent at humeri, undefined in rest of elytral length. Two rows of inconspicuous, irregular cells traceable between costae, giving appearance of secondary costa in some parts of elytra. Elytra with dense, long pubescence. Wings fully developed. Legs slender, coxae long, movable, trochanters very slender, femora flat, robust, tibiae slenderer than femora, tarsi slender, 5-segmented, tarsomeres 3–4 with small pads, tarsomere 5 long, slender, claws simple. Male abdomen slender, shorter and narrower than elytra, 8 segmented, tergum 8 simply rounded at apex, sternum 8 with strengthened lateral margins at base and membranous window basally. Male genitalia trilobate, variable in relative length of phallus and parameres, phallus stout, almost straight and sometimes laterally compressed or phallus long, very slender, sickle like (Figs 52–72); parameres reaching to half or four fifths of phallic length, with fine spines along internal margin, fully sclerotized (Figs 44–47) or considerably shorter, apically with membranous process, phallobase short, emarginate basally (Figs 48–51).
Diagnosis. Body shape characteristic ("trilobite larva" Figs 32–43). Two body types are present: the flat, wide (Figs 32–42) and robust, vermiform (Fig. 43). Although different in general appearance these larvae share common diagnostic characters: the fossa antennalis closed, separated from the mouth-parts by pleurostoma (the pleurostoma absent from other lycids); slender, longitudinal sclerite present ventrally of pleurostoma; mala sclerotized. The apical antennomere with several peg-like processes. The complex, oval meso- and metathoracic spiracles are cribriform, the sieve plate with multiple openings. They are situated in large depressions. Similar larvae of Lyropaeus differ in the shape of the apical antennomere and both Macrolibnetis and Lyropaeus do not have spiracular cavities in abdominal segments (
Description. Body wide, considerably flattened due to extensively projected lateral plates (Figs 32–41) or slender with postero-lateral processes (Fig. 43), usually dark brown, cryptically coloured, some species aposematically coloured with brightly coloured patches. Head prolonged, rounded anteriorly; epicranium consists of dorsal and pleural plates, membranous between plates. Complete fossa antennalis dorsally limited by epicranium, ventrally by sclerotized pleurostoma. Longitudinal sclerite situated ventrally of pleurostoma. Basal antennomere very short, apical antennomere with several peg-like processes ventrally and more extensive, sclerotized area dorsally. Mala sclerotized, with peg-like process. Cervical membrane extensive, with pigmented patches postero-ventrally. Pronotum trapezoid, terga with considerably widened lateral plates and sometimes with tubercles at posterior margin. Prosternum prolonged, episterna extensive, attached to prosternum. Extensive spiracular plates with spiracular openings at margin and bottom of extensive cavity in both, meso- and metathorax. Legs slender, relatively long, trochanters divided in two parts. Abdomen with large lateral processes, spiracular openings on margin and bottom of cavities in segments A1–A8. Sterna A1–A8 with slender postero-lateral processes, upper pleurites extensive, with similar process at outer posterior angle. Lower pleurites very small, with short process only in segments A3–A8. Segment A9 widest at apex, with short, fixed urogomphi (Figs 35–40, 43).
Sexually mature larviform females observed only by
The information on biology was given by
Diagnosis. The species group Platerodrilus paradoxus was recovered as a monophyletic assemblage representing one of principal Platerodrilus lineages. The species of this group share male genitalia with long, slender and completely sclerotized parameres (Figs 44–47). Known females do not have any glabrous tubercles in the middle of thoracic terga (Figs 34–38, 40), one species from Mt. Kinabalu has tubercles only at posterior margins of thoracic terga (Fig. 40). The following species are classified here: Platerodrilus bicolor Wittmer, 1941, Platerodrilus curtus Pic, 1931, Platerodrilus foliaceus sp. n., Platerodrilus paradoxus Mjöberg, 1925, Platerodrilus piceicollis Pic, 1943, Platerodrilus strbai Kazantsev, 2009, Platerodrilus svetae Kazantsev, 2009, Platerodrilus wongi sp. n.
Distribution. Most species are known from Borneo and the Philippines, only Platerodrilus wongi sp. n. occurs in Sumatra.
Remark. As only Platerodrilus curtus, Platerodrilus foliaceus and several unidentified female larvae were available for DNA isolation, the monophyly of this lineage needs further support before validity of the name Duliticola Mjöberg, 1925 can be reconsidered.
Syntype. Male (BMNH), Borneo. Syntype. Female (BMNH), Lundu, Sarawak, G. E. Bryant, 6. 1. 14.
Platerodrilus paradoxus belongs to a group of Bornean species with robust and long parameres. The male of Platerodrilus paradoxus resembles Platerodrilus foliaceus but differs in the slender apex of parameres. Additionally these species differ in larval morphology (Figs 34, 40).
Male. Body 7 mm, dark brown, only humeri and elytral suture slightly lighter. Head small, with hemispherically prominent eyes, head with eyes wider than frontal margin of pronotum, eye diameter 1.9 times frontal interocular distance. Antennae compressed, covered with long, erected, dense pubescence. Pronotum flat, without carinae, 1.8 times wider than long at midline, frontal margin widely rounded anteriorly, frontal angles obtuse, lateral margins almost straight, posterior margin bisinuate. Elytra flat, parallel-sided, elytral costae inconspicuous, elytra 2.6 times longer than width at humeri. Legs slender, compressed, densely pubescent. Male genitalia with robust parameres, phallus slightly curved with bulbous tip. Parameres stout with hooked tip, apical half of ventral edge serrate. Phallobase wide, deeply emarginate.
Female larva. Body flat and wide (Fig. 34), pronotum parallel-sided at base, then gradually tapering to front, triangular, without any glabrous tubercles in disc, only small tubercles in middle part of posterior margins of thoracic segments, in middle of anterior pronotal margin four subequal tubercles; mesothorax strongly transverse, with rounded lateral margins and straight posterior margin, posterior angles obtuse, metathorax similar in shape with more acutely projected posterior angles. Abdomen with short, robust lateral processes.
Male. BL 6.9 mm, PL 1.0 mm, PW 1.9 mm, HW 2.0 mm, Edist 0.85 mm, Ediam 0.44 mm. Larva. BL 53.0 mm, PL 8.7 mm, PW 13.4 mm.
Malaysia: Sarawak. Known only from the type locality.
http://zoobank.org/53AA2216-605A-4A64-A10B-C58AEF0B8E31
Holotype. Male (LMBC, UPOL 000589), Borneo, Central Kalimantan Prov., 60 km SE Muara Teweh, 1°20'25"S, 115°20'16"E, 24.–28. Jun. 2001, 150 m. Paratypes. 12 females, same locality data (LMBC, UPOL 000588).
Platerodrilus foliaceus belongs to the Platerodrilus paradoxus group and the male resembles Platerodrilus paradoxus in general appearance. Platerodrilus foliaceus differs in the rounded apex of parameres (Figs 46–47). The female larvae of Platerodrilus foliaceus are very flat and have much slenderer lateral processes of abdominal segments than Platerodrilus paradoxus (Figs 34, 40).
Male. Body small-sized, brown, head, antennae, legs except bases of femora and apical three fifths of elytra dark brown (Fig. 4). Head small, with hemispherically prominent eyes, head with eyes slightly wider than frontal margin of pronotum, eye diameter 1.7 times frontal interocular distance. Antennae compressed, covered with long, erected, dense pubescence, length of antennomere 3 0.7 times antennomere 2. Pronotum flat, without carinae, 1.7 times wider than long at midline, frontal margin slightly projected anteriorly, frontal angles obtuse, but apparent, lateral margins rounded, posterior margin slightly bisinuate (Fig. 22). Elytra flat, parallel-sided, elytral costae inconspicuous, elytra 2.9 times longer than width at humeri. Legs slender, compressed, densely pubescent. Male genitalia with robust parameres, phallus slightly curved with bulbous tip. Parameres stout with hooked tip, apical half of ventral edge serrate. Phallobase wide, deeply emarginate (Figs 46–47).
Female larva. Body extremely flat and wide (Fig. 40), pronotum triangular, without any glabrous tubercles, in middle of anterior margin four subequal tubercles; mesothorax strongly transverse, with rounded lateral margins and moderately projected posterior angles, metathorax similar in shape with more acutely projected posterior margins. Abdomen with very slender and long lateral processes.
Male. BL 6.0 mm, PL 0.9 mm, PW 1.5 mm, HW 1.7 mm, Edist 0.78 mm, Ediam 0.46 mm. Larva. BL 19.3 mm, PL 5.1 mm, PW 11.2 mm.
Indonesia: Kalimantan. Known only from the type locality.
The specific epithet refers to the flat body shape of the female larva.
The males and female larvae were identified as conspecific on the basis of highly similar sequences of rrnL (Fig. 1).
http://zoobank.org/67F26E3D-C5E3-4E35-A630-B6A636DB969D
Holotype. Male (LMBC), Sumatra Utara, Brastagi, Gn. Sibayak, 19–23. Feb. 1998, 700–2000 m.
Platerodrilus wongi is a single species of the Platerodrilus paradoxus group occurring in Sumatra. It resembles Platerodrilus curtus from the Philippines in the uniformly yellow elytra, but has relatively shorter parameres (Figs 44–45).
Body brown, head, pronotum, mesoscutellum and elytra yellow (Fig. 5). Head small, with eyes is slightly wider than frontal margin of pronotum, antennal tubercles deeply separated. Eyes hemispherically prominent, frontal interocular distance 2.4 times eye diameter. Antennae slender, compressed, reaching two thirds of elytra length, antennomere 3 1.1 times antennomere 2. Head and antennae covered with short dense pubescence. Pronotum transverse, 1.6 wider than length at midline. Anterior margin only slightly projected, anterior angles well marked, lateral margins almost straight, posterior margin shallowly bisinuate (Fig. 28). Elytra parallel-sided, with inconspicuous carinae. Elytra 3.0 times longer than width at humeri, elytra widest in apical sixth. Legs compressed, densely pubescent. Male genitalia with laterally compressed, slightly curved phallus with bulbous tip, parameres stout, long, with hooked tip, apical half of ventral edge serrated, phallobase wide, deeply emarginate (Figs 44–45).
BL 7.4 mm, PL 1.0 mm, PW 1.6 mm, HW 1.9 mm, Edist 0.91 mm, Ediam 0.38 mm.
Indonesia: Northern Sumatra.
The specific epithet is a patronym in honour of Alvin T. C. Wong.
| 1 | Parameres short, reaching slightly over half of phallic length | Platerodrilus strbai Kazantsev |
| – | Parameres reaching almost to the apex of the phallus | 2 |
| 2 | Male antennae flabellate | Platerodrilus bicolor Wittmer |
| – | Male antennae serrate | 3 |
| 3 | Whole elytra yellow (Fig. 5) | 4 |
| – | Elytra dark brown or light brown with dark coloured apical part | 5 |
| 4 | Parameres reaching to five sixths of the phallic length | Platerodrilus curtus Pic |
| – | Parameres reaching to three fourths of the phallic length | Platerodrilus wongi sp. n. |
| 5 | Only apical part of elytra dark coloured, pronotum black | Platerodrilus piceicollis Pic and Platerodrilus svetae Kazantsev* |
| – | Whole elytra dark brown, at most the narrow humeral part slightly lighter brown | 6 |
| 6 | Body slender, inner margin of parameres serrate only in its apical half | Platerodrilus foliaceus sp. n. |
| – | Body robust, inner margin of parameres serrate in three quarter of their length | Platerodrilus paradoxus (Mjöberg) |
* We failed to find any distinguishing character between Platerodrilus svetae and Platerodrilus piceicollis. The type of Platerodrilus svetae is deposited in a private collection and unavailable for study.
Diagnosis. The species group Platerodrilus major is a monophyletic assemblage representing a terminal branch, which includes the species placed in Platrilus Kazantsev, 2009 and it is represented in the current analysis by Platerodrilus major and Platerodrilus ngi (Fig. 1). The group is characterized by the short and setose parameres without long membranous apical processes (Figs 48–51). The following species are placed here: Platerodrilus atronotatus Pic, 1943, Platerodrilus crassicornis Pic, 1923, Platerodrilus hirtus Wittmer, 1938, Platerodrilus major Pic, 1921, Platerodrilus ngi sp. n. and Platerodrilus wittmeri sp. n. The species described by M. Pic were redescribed by
Holotype. Male (LMBC), Singapore, Bukit Timah and Central Water Catchment, 19.–22. May 2013, 50–100 m, E. Jendek and O. Šauša leg. Paratypes. Female larvae, 6 spec., Malaysia, Pahang, Tioman, Kg. Tekek–Juara trail, 50–300 m, 2°49'10"N, 104°10'21"E, 29. Mar.–2. Apr. 2013, same locality data, 4.–16. Mar. 1998, L. Dembicky and P. Pacholatko (LMBC); 1 spec., Singapore, Sime Road swamp, 30. Oct. 2008 (ZRCS);1 spec., Singapore, Bukit Timah Nature Reserve, A. T. C. Wong 1993 (ZRCS); 1 spec., Singapore, Sime Road, C. Lee (ZRCS, #6.20969, 1993.7277, 1993.7278).
Platerodrilus ngi is the only representative of the Platerodrilus major group known from Singapore and Tioman. It differs in dark red colouration of the pronotum and humeral two thirds of elytra from Platerodrilus atronotatus from the Malay Peninsula. Platerodrilus atronotatus has the black pronotum. Additionally, the phallus of Platerodrilus atronotatus is slender and antennomeres 3 and 4 short and much wider.
Male. Body small, dark brown to black, head, prothorax, mesoscutellum and basal two thirds of elytra dark red; whole body with dense, short, pubescence. Head small, including eyes slightly wider than frontal margin of pronotum. Eyes hemispherically prominent, frontal interocular distance 1.8 times eye diameter. Antennae robust at base, compressed, reaching two thirds of elytral length, antennomere 3 0.6 times antennomere 2, antennomere 3 as long as wide at apex. Pronotum flat, 1.7 times wider than long at midline. Anterior margin almost straight, anterior angles sharply marked, posterior margin bisinuate. Elytra slightly wider posteriorly, elytra 2.8 times longer than width at humeri; only slightly widened posteriorly, elytral costae conspicuous. Legs compressed, densely pubescent. Male genitalia with straight phallus and setose parameres, short phallobase slightly shorter than parameres.
Female larva. Body slender, parallel-sided (Fig. 43), pronotum triangular, with two basal, dark coloured, glabrous tubercles; mesothorax slightly transverse, without projected posterior angles, metathorax similar in shape. Abdomen with robust, short lateral processes and fixed urogomphi.
Male. BL 6.9 mm, PL 0.9 mm, PW 1.6 mm, HW 2.1 mm, Edist 0.86 mm, Ediam 0.48 mm. Larva. BL 30.2 mm, PL 5.7 mm, PW 5.6 mm.
Singapore, Malaysia: Pahang. Biology and female specimens collected in Singapore were reported by
The species name is a patronym in honour of Peter Ng.
http://zoobank.org/F646300B-C3B4-48BA-98B2-9BB9571C762B
Holotype. Male (LMBC), Java, K. O. Blawan, Ijen Plateau, Jul. 1940, 900–1500 m, H. Lucht coll.
Platerodrilus wittmeri is the only bicoloured species from the Platerodrilus major group in Java. It resembles Platerodrilus major from Northern Sumatra in colouration but differs in the shorter phallobase (Figs 48–49, 50–51).
Male. Body medium-sized, dark brown, head, prothorax, mesoscutellum and basal half of elytra testaceous; antennae, legs, apical half of elytra dark brown to black (Fig. 17). Head small, including eyes slightly wider than frontal margin of pronotum. Eyes hemispherically prominent, frontal interocular distance 1.9 times eye diameter. Antennae slender, compressed, reaching two thirds of elytral length, antennomere 3 0.7 times antennomere 2. Head and antennae with dense, short, pubescence. Pronotum flat, 1.9 times wider than long at midline. Anterior margin slightly projected anteriorly, anterior angles sharply marked, prominent, posterior margin bisinuate (Fig. 30). Elytra almost parallel-sided, 3.5 times longer than width at humeri; only slightly widened posteriorly, elytral costae inconspicuous. Legs compressed, densely pubescent. Male genitalia with straight phallus and setose parameres, short phallobase slightly shorter than parameres (Figs 50–51).
BL 10.1 mm, PL 1.1 mm, PW 2.0 mm, HW 2.6 mm, Edist 0.95 mm, Ediam 0.49 mm.
Indonesia: Java.
The specific epithet is a patronym in honour of the late W. Wittmer, who donated the specimen to the senior author in 1992.
| 1 | Metathorax orange brown | Platerodrilus crassicornis Pic |
| – | Metathorax dark brown to black | 2 |
| 2 | Phallobase shorter than parameres (≤ 0.95 times length of parameres) | Platerodrilus wittmeri sp. n. |
| – | Phallobase longer than parameres (≥ 1.05 times length of parameres | 3 |
| 3 | Phallus short and stout, about 0.90 the combined length of parameres and phallobase, pronotum similar in colour to humeral part of elytra | 4 |
| – | Phallus long and slenderer, about as long as the combined length of parameres and phallobase, pronotum blac | Platerodrilus atronotatus Pic |
| 4 | Antennomere 4 wide, about as long as wide at apex | Platerodrilus ngi sp. n. |
| – | Antennomere 4 at least 1.25 times longer than wide at apex | 5 |
| 5 | The expanded base of phallus in lateral view mostly hidden by parameres, phallobase deeply emarginate basally, elytra black apically | Platerodrilus major Pic |
| – | The expanded base of phallus in lateral view exposed, considerably widened, phallobase shallowly emarginate basally, elytra testaceous | Platerodrilus hirtus Wittmer |
Diagnosis. The group Platerodrilus sinuatus is a paraphyletic assemblage consisting of Platerodrilus luteus, the Indo-Burmese species (represented in the analyzed dataset by female larvae VP2304, VP2311), Platerodrilus palawanensis from Palawan, Platerodrilus indicus from Assam and Nepal and the terminal lineage of Platerodrilus from Sundaland (Figs 1–2). All species have a long, slender, considerably curved phallus and short parameres with the membranous apical process (Figs 52–72). The following species are placed in this species group: Platerodrilus angustatus Pic, 1921, Platerodrilus apicalis Pic, 1936, Platerodrilus atricolor Pic, 1938, Platerodrilus corporaali Pic, 1921, Platerodrilus grootaerti Kazantsev, 2009, Platerodrilus holynskae Kazantsev, 2009, Platerodrilus inapicalis Pic, 1937, Platerodrilus indicus Wittmer, 1966, Platerodrilus luteus sp. n., Platerodrilus maninjauensis sp. n., Platerodrilus montanus sp. n., Platerodrilus palawanensis sp. n., Platerodrilus ranauensis sp. n., Platerodrilus reductus Pic, 1926, Platerodrilus rotundicollis Wittmer, 1938, Platerodrilus ruficollis Pic, 1942, Platerodrilus rufus Pic, 1924, Platerodrilus sinuatus Pic, 1921, Platerodrilus talamauensis sp. n., Platerodrilus tujuhensis sp. n., Platerodrilus sibayakensis sp. n. and Platerodrilus sinabungensis sp. n. The female larvae of species occurring in continental Asia north of the Isthmus of Kra have terga without glabrous tubercles similarly to the species of the Platerodrilus paradoxus clade. The group of species from the Malay Peninsula, Sumatra and Java are characterized by larvae with glabrous tubercles in thoracic terga (Figs 1, 32–33, 39, 41). The male genitalia are similar in the shape of the basal part of the phallus and phallobase. The minute differences are difficult to describe in a form of the identification key and the DNA data were used for confirmation of the species delineation in several cases.
http://zoobank.org/21013FB1-3B9C-42F2-879D-A1CAC5AF4EED
Holotype. Male (LMBC, 000586), Java, Ijen N. P., 12 km W of Sodora, 3–5. May 2001, 1000 m.
Platerodrilus ijenensis is the only Javanese species of Platerodrilus sinuatus group with bicoloured elytra. It resembles in colour pattern the Sumatran species Platerodrilus corporaali, which differs in the reddish coloured head and basal antennomeres, and the very short antennomere 3.
Body and head dark brown to black, pronotum, mesoscutellum and elytra in humeral half orange, apical part of elytra dark brown to black (Fig. 14). Head small, including eyes narrower than pronotum at posterior angles, antennal tubercles slightly prominent. Eyes hemispherically prominent, eye diameter 1.8 times frontal interocular distance. Antennae slender, compressed, length of antennomere 3 1.2 times antennomere 2. Pronotum transverse, 1.8 wider than long at midline, anterior margin slightly projected forward, anterior angles marked, lateral margins almost straight, posterior margin bisinuate (Fig. 18). Elytra 3.3 times longer than width at humeri, elytra parallel-sided, elytral costae inconspicuous. Legs slender, compressed, densely pubescent. Male genitalia with slender, curved phallus and small rounded parameres bearing slender membranous process, phallobase wide, narrowly and deeply emarginate (Figs 64–65).
BL 7.2 mm, PL 0.9 mm, PW 1.6 mm, HW 1.8 mm Edist 0.70 mm, Ediam 0.40 mm.
Indonesia: Java.
The specific name refers to the type locality.
Lectotype (hereby designated). Male (BMNH), Sumatra, Korinchi, 4500 ft, N. 1914, K. G. Blair. Paralectotype. Female (BMNH), same locality data.
The male genitalia are missing. Therefore, only information on external morphology can be compared. The orange pronotum and humeral part of elytra resemble Platerodrilus corporaali or Platerodrilus maninjauensis, but no similar species is known from the Kerinci massif.
Male. Body black, head dark brown, pronotum and basal quarter of elytra orange; antennae, and legs dark brown to black. Head small, antennal tubercles weak, eyes hemispherically prominent, frontal interocular distance 2.3 times maximum eye diameter. Antennae slender, compressed, densely pubescent, antennomere 3 as long as antennomere 2. Pronotum transverse, 1.5 times wider than long, anterior margin almost straight, frontal angles conspicuous, lateral margins almost straight, posterior angles acute, surface mat at margins, slightly glabrous in middle. Elytra parallel-sided, elytral costae weak, covered with dense pubescence. Male genitalia missing.
PL 1.3 mm, PW 1.9 mm, HW 2.6 mm, Edist 0.84 mm, Ediam 0.36 mm.
Indonesia: Sumatra, Jambi, Kerinci massif.
The lectotype is damaged (the apical half of elytra and abdomen are missing). Platerodrilus korinchianus differs in the shape of pronotum and colouration of elytra from Platerodrilus tujuhensis and Platerodrilus robinsoni. The paralectotype, a female larva, is very similar to the female larva of Platerodrilus tujuhensis from the same locality and might not be conspecific with the male specimen. As larvae are generally difficult to identify without DNA data, we prefer to designate a lectotype to keep status and preserve the validity of the name. The species can be misidentified as high diversity of neotenic net-winged beetles in the region was documented (
http://zoobank.org/6BE29EC6-3F39-4775-83EC-7BAB16828345
Holotype. Male (LMBC, UPOL 001379), Sumatra, Jambi Kersik Tua, Gn. Kerinci, 19.–22. Jan. 2005, 1600–2200 m.
Platerodrilus luteus resembles in general appearance the syntopically occurring Platerodrilus robinsoni, but these species are distantly related according to the recovered molecular phylogeny (Fig. 1). The male of Platerodrilus luteus differs from similarly coloured Sumatran Platerodrilus in the very wide and broadly emarginate phallobase (Fig. 67).
Body black, head dark brown, pronotum and elytra orange; antennae, and legs dark brown to black (Fig. 8). Head small, antennal tubercles weak, eyes hemispherically prominent, frontal interocular distance 2.8 times maximum eye diameter. Antennae slender, compressed, densely pubescent, antennomere 3 1.5 times longer than antennomere 2. Pronotum transverse, 1.7 times wider than long, frontal angles inconspicuous, lateral margins slightly convex, posterior angles acute (Fig. 23). Elytra parallel-sided, 3.7 times longer than width at humeri, elytral costae weak, covered with dense pubescence. Male genitalia with curved phallus; parameres short, rounded, with slender membranous processes; phallobase wide, deeply emarginate (Fig. 67–68).
BL 10.1 mm, PL 1.1 mm, PW 1.8 mm, HW 2.3 mm, Edist 0.83 mm, Ediam 0.45 mm.
Indonesia: Sumatra.
The specific epithet refers to yellow colouration of the body.
http://zoobank.org/6F62649B-E974-4BF7-BD30-BF13FD3A8681
Holotype. Male (LMBC, UPOL 001386), Sumatra, Barat Lake Maninjau, E coast, 12.–23. Jan. 2005, 800 m. Paratype. Male (LMBC, UPOL 001374), Sumatra, Barat Lake Maninjau, E coast, 12.–23. Jan. 2005, 800 m. Paratypes. Male, 2 females (LMBC, UPOL 001377), Sumatra, Barat, Pasaman, Gn. Talamau, 14.–15. Jan. 2005, 1000 m; female larva (LMBC, UPOL VP2303), Sumatra, Barat Lake Maninjau, E coast, 12.–23. Jan. 2005, 800 m.
Platerodrilus maninjauensis is a sister species to Platerodrilus tujuhensis from Northern Sumatra. These species differ in the colouration (Figs 7, 12) and the shape of the phallus and phallobase (Figs 56–57, 60–61). The female larvae of both species are similar and differ only in the relative size of the mesonotal tubercles, which are smaller in Platerodrilus tujuhensis (Figs 32, 41).
Male. Body medium-sized, dark brown; head, prothorax, mesoscutellum and basal three fifths of elytra orange; antennae, legs, apical two fifths of elytra dark brown to black (Fig. 7). Head small, including eyes narrower than frontal margin of pronotum. Eyes hemispherically prominent, eye diameter 2.2 times frontal interocular distance. Antennae slender, compressed, reaching two thirds of elytral length, antennomere 3 0.9 times antennomere 2. Head and antennae with dense, short, pubescence. Pronotum flat, 1.1 times wider than long at midline. Anterior margin widely rounded, anterior angles inconspicuous, posterior margin bisinuate (Fig. 20). Elytra almost parallel-sided, 3.5 times longer than width at humeri; slightly widened posteriorly, widest at apical fourth. Elytral costae inconspicuous. Legs compressed, densely pubescent. Male genitalia with short rounded parameres bearing slender membranous process. Phallus curved, phallobase wide, widely emarginate (Figs 60–61).
Female larva. Body flat, wide (Fig. 32), pronotum triangular, with two glabrous rounded tubercles postero-laterally, another two tubercles in middle of anterior margin; mesothorax strongly transverse, with rounded lateral margins and weakly projected posterior angles, laterally with four tubercles, upper rounded, lower transverse, metathorax similar in shape with more acutely projected posterior margins. Abdominal segments with slender and long lateral processes.
BL 7.0 mm, PL 0.9 mm, PW 1.6 mm, HW 1.7 mm, Edist 0.84 mm, Ediam 0.38 mm. Larva. BL 24.1 mm, PL 5.4 mm, PW 10.3 mm.
Indonesia: Sumatra.
The specific epithet refers to the type locality of the holotype.
http://zoobank.org/A434CA37-4D6B-4055-92F6-A16A1D3CA9B5
Holotype. Male (LMBC, UPOL 001371), Sumatra Utara, Brastagi, Gn. Sibayak, 26. Jan.–1. Feb. 2005, 1600–2200 m. Paratype. Female larva (LMBC, UPOL VP2308), Sumatra Utara, Brastagi, Gn. Sinabung, 29.–30. Jan. 2005, 1400–2000 m.
Platerodrilus montanus and Platerodrilus sinabungensis are the only Sumatran species with the dark coloured pronotum. These species differ in the shape of the phallobase, when Platerodrilus montanus has the narrowly emarginate base (Figs 58–59, 66). The larva of Platerodrilus montanus has pronotum without any glabrous tubercles in the disc (Fig. 33).
Body medium-sized, dark brown to black, only basal half of elytra orange and pronotum with irregular light coloured patches in disc (Fig. 13). Head small, including eyes slightly narrower than frontal margin of pronotum. Eyes hemispherically prominent, eye diameter 2.4 times frontal interocular distance. Antennae compressed, length of antennomere 3 1.1 times antennomere 2. Head and antennae with dense, short, pubescence. Pronotum transverse, 1.9 times wider than long at midline; anterior margin slightly projected, lateral margins almost straight, anterior angles weakly marked, posterior margin of pronotum slightly bisinuate (Fig. 21). Elytra almost parallel-sided, elytra 3.5 times longer than width at humeri; slightly widened posteriorly, widest at apical fourth, elytral costae inconspicuous. Legs compressed, with dense pubescence. Male genitalia with short rounded parameres bearing slender membranous processes. Phallus curved, phallobase wide, deeply emarginate (Fig. 66).
Female larva. Body flat, wide (Fig. 33), pronotum triangular, without glabrous tubercles except two tubercles in middle of posterior margin; mesothorax strongly transverse, with rounded lateral margins and weakly projected posterior angles, laterally with four tubercles, upper rounded, lower only slightly transverse, metathorax similar in shape with more acutely projected posterior margins. Abdominal segments with slender and long lateral processes.
BL 8.2 mm, PL 0.9 mm, PW 1.8 mm, HW 2.1 mm, Edist 0.94 mm, Ediam 0.39 mm. Larva. BL 32.3 mm, PL 7.7 mm, PW 13.0 mm.
Indonesia: Sumatra, North Sumatra Province, Gn. Sibayak.
The specific epithet is derived from the Latin adjective montanus (mountainous) referring to the habitat of the species.
http://zoobank.org/F7FBBEE1-3910-4258-AEE4-E819747B834C
Holotype. Male (LMBC, UPOL 001382), Philippines, Palawan, Tanabak river, 150 m, 10°02'49"N, 118°58'31"E, 2.–5. Jan. 2007, Bolm lgt. Paratypes, 8 males (LMBC), same locality data, 22. Dec. 1991; 3 paratypes. Males (KMTC), Philippines, Palawan, Brooke's point, 8. Dec. 2002, leg. F. A. Dacasin. Paratype. Male (KMTC), Philippines, Palawan, Brooke's point, 18. May 2003, leg. F. A. Dacasin. Paratype. Male (KMTC), Philippines, Palawan, Brooke's point, 15. Jan. 2005, leg. F. A. Dacasin.
Platerodrilus palawanensis sp. n. resembles Platerodrilus borneensis in flabellate antennae and these species differ in colouration. Platerodrilus borneensis is dark brown and Platerodrilus palawanensis bicoloured (Fig. 16). Additionally Platerodrilus palawanensis has the very short pronotum and straight frontal pronotal margin (Fig. 31).
Body medium-sized, dark brown to black, only pronotum, mesoscutellum and humeral two thirds of elytra orange; apical third of elytra and two thirds of elytral suture dark brown to black. Head small, including eyes apparently narrower than frontal margin of pronotum, antennal tubercles robust, deeply separated. Eyes hemispherically prominent, eye diameter 1.9 times frontal interocular distance. Antennae flabellate, antennomere 3 with long process, pubescent, length of antennomere 3 1.6 times antennomere 2. Pronotum strongly transverse, 1.5 times wider than long at midline, anterior margin straight, with prominent anterior angles, lateral margins almost straight, posterior margin bisinuate, surface of disc mat, finely punctuate, with dense short pubescence. Elytra 2.9 times longer than width at humeri, elytra almost parallel-sided; slightly widened posteriorly, widest at apical fourth, elytral costae inconspicuous. Legs compressed, with dense pubescence. Male genitalia with curved phallus, short rounded parameres bearing slender membranous processes; phallobase wide, deeply emarginate (Figs 69–70).
BL 7.1 mm, PL 1.0 mm, PW 1.6 mm, HW 1.9 mm, Edist 0.66 mm, Ediam 0.34 mm.
Philippines: Palawan.
The specific epithet refers to the type locality of the holotype.
http://zoobank.org/F09DE9AF-6800-4A06-AF7D-B82F3207217E
Holotype. Male (LMBC, UPOL 000587), Sumatra, SW coast of Ranau Lake, 1–4. Jun. 2001, 1200 m.
Platerodrilus ranauensis was found as a sister species to Platerodrilus talamauensis (Fig. 1) and these species differ in the extent and shape of the orange part of elytra and in the shape of the posterior margin of phallobase (Figs 54–55).
Body medium-sized, dark brown to black, only pronotum testaceous to brown and basal quarter of elytra orange (Fig. 9). Head small, including eyes slightly narrower than frontal margin of pronotum, antennal tubercles small. Eyes hemispherically prominent, eye diameter 2.3 times frontal interocular distance. Antennae compressed, pubescent, length of antennomere 3 1.1 times antennomere 2. Pronotum transverse, 1.7 times wider than long at midline, anterior margin widely rounded, semicircular, without prominent anterior angles, posterior margin of pronotum bisinuate, surface of disc glabrous, with sparse long pubescence (Fig. 24). Elytra almost parallel-sided, 3.0 times longer than width at humeri; slightly widened posteriorly, widest at apical fourth. Elytral costae inconspicuous. Legs compressed, with dense pubescence. Male genitalia with short rounded parameres bearing slender membranous processes; phallus curved, phallobase wide, deeply emarginate (Figs 54–55).
BL 6.0 mm, PL 0.8 mm, PW 1.3 mm, HW 1.7 mm, Edist 0.79 mm, Ediam 0.35 mm.
Indonesia: Sumatra.
The specific epithet refers to the type locality of the holotype.
http://zoobank.org/3A4F0BBB-BA23-498D-8C1E-E39CA46F1D84
Holotype. Male (LMBC, UPOL 001389), Sumatra Utara, Brastagi, Gn. Sibayak, 26. Jan.–1. Feb. 2005, 1600–2200 m. Paratypes. 2 males (LBMC), Sumatra, SW of Brastagi, Gn. Sinabung, 22. Feb. 1991, 1300–1800 m. Paratype. Male (LBMC), Sumatra, SW of Brastagi, Gn. Sinabung, 19.–23. Feb. 1991, 1300–1800 m. Paratype. Male (LBMC, UPOL 001372), Sumatra, Utara, Brastagi, Gn. Sibayak, 26. Jan.–1. Feb. 2005, 1600–2200 m. Paratype. Male (LBMC), Sumatra, Sinabung, Mar. 1998.
Platerodrilus sibayakensis and Platerodrilus angustatus were recovered as sister species (Fig. 1) and they share the pronotum with acutely projected posterior angles. Platerodrilus sibayakensis differs in the 4.1 times longer antennomere 4 than its width in the middle and the more robust phallus tapering gradually from the base to apex (Figs 71–72).
Body medium-sized, dark brown to black, pronotum testaceous to brown and basal third of elytra orange (Fig. 10). Head small, including eyes slightly narrower than frontal margin of pronotum, antennal tubercles small. Eyes hemispherically prominent, eye diameter 1.7 times frontal interocular distance. Antennae compressed, pubescent, length of antennomere 3 0.7 times antennomere 2. Pronotum transverse, 1.9 times wider than long at midline, anterior margin almost straight, with marked anterior angles, lateral margins straight, posterior margin of pronotum bisinuate, posterior angles acutely projected, surface of disc weakly glabrous, finely punctuate, with long pubescence (Fig. 25). Elytra almost parallel-sided, elytra 3.5 times longer than width at humeri; slightly widened posteriorly, widest at apical fourth, elytral costae inconspicuous. Legs compressed, with dense pubescence. Male genitalia with short rounded parameres bearing slender membranous processes; phallus curved, phallobase wide, deeply emarginate (Figs 71–72).
BL 8.5 mm, PL 0.9 mm, PW 1.8 mm, HW 2.0 mm, Edist 0.73 mm, Ediam 0.44 mm.
Indonesia: Sumatra.
The specific epithet refers to the type locality of the holotype.
http://zoobank.org/4275B855-5901-45B6-BD6A-CF6EB0BFB430
Holotype. Male (LMBC), Sumatra, SW of Brastagi, Gn. Sinabung, 22. Feb. 1991, 1300–1500 m. Paratypes. 2 males (LBMC), Sumatra, SW of Brastagi, Gn. Sinabung, 22. Feb. 1991, 1400–1900 m.
Platerodrilus sinabungensis resembles Platerodrilus montanus in the dark coloured pronotum. These species differ in the shape of the phallobase (Figs 58–59, 66).
Body medium-sized, dark brown to black, only basal two fifths of elytra orange testaceous (Fig. 11). Head including eyes slightly wider than frontal margin of pronotum. Eye hemispherically prominent, their diameter 2.3 times frontal interocular distance. Antennae compressed, antennomere 3 as long as antennomere 2. Head and antennae with short dense pubescence. Pronotum transverse, 1.7 wider than long at midline, anterior and lateral margins weakly rounded, posterior margin of pronotum simply rounded to straight in middle, disc bare in middle, pubescent along lateral margins (Fig. 26). Elytra with inconspicuous carinae. Elytra 3.4 times longer than width at humeri, elytra widest posteriorly. Legs compressed with dense pubescence. Male genitalia with curved phallus and short rounded parameres bearing slender membranous processes with basal part serrate; phallobase wide, widely emarginate (Figs 58–59).
BL 8.9 mm, PL 1.0 mm, PW 1.7 mm, HW 2.1 mm, Edist 0.89 mm, Ediam 0.4 mm.
Indonesia: Sumatra.
The specific epithet refers to the type locality of the holotype.
http://zoobank.org/FB6FE156-5B2D-4232-897F-62EC10EFFB1F
Holotype. Male (LMBC, UPOL 001376), Sumatra Barat, Pasaman, Gn. Talamau, 14.–15. Jan. 2005, 1000 m. Paratype. Male (LMBC, UPOL 001375), Sumatra Barat, Pasaman, Gn. Talamau, 14.–15. Jan. 2005, 1000 m.
Platerodrilus talamauensis and Platerodrilus ranauensis are closely related (Fig. 1) and they differ in the extent and shape of the orange part of the elytra (Figs 9, 15) and in the shape of posterior margin of phallobase (Figs 52–55).
Male. Body medium-sized, dark brown to black, only pronotum, mesoscutellum and basal three fifths of elytra orange (Fig. 15). Head small, with eyes slightly wider than frontal margin of pronotum. Eyes hemispherically prominent, eye diameter 2.4 times frontal interocular distance. Antennae compressed, reaching two thirds of elytral length, antennomere 3 0.9 times antennomere 2. Head and antennae densely pubescent. Pronotum transverse, 1.2 wider than long at midline. Anterior margin of pronotum rounded, anterior angles inconspicuous, posterior margin bisinuate (Fig. 29). Elytra with inconspicuous carinae, parallel-sided, 2.9 times longer than width at humeri, widest posteriorly. Legs compressed with dense pubescence. Male genitalia with curved phallus, phallus twice longer than apical processes of parameres, phallobase widely emarginate. (Fig. 52–53).
BL 6.5 mm, PL 1.0 mm, PW 1.6 mm, HW 1.9 mm, Edist 0.88 mm, Ediam 0.36 mm.
Indonesia: Sumatra.
The specific epithet refers to the type locality of the holotype.
http://zoobank.org/52952DAE-6457-4DF0-B57C-E4A7F0989063
Holotype. Male (LMBC, UPOL 001385), Sumatra, Jambi Kersik Tua, Gn. Kerinci, 19–22. Jan. 2005, 1600–2200 m. Paratype. Female (LMBC, VP2305), Sumatra, Jambi prov. Kerinci Seblat N. P., 7 km E Kayuaro, Mt. Tujuh, 1°45'S, 101°25'E, 25. Feb.–2. Mar. 2003, 1750 ± 250 m.
Platerodrilus tujuhensis resembles in the uniformly light coloured head, pronotum and elytra Platerodrilus robinsoni, which differs in the more robust basal part of the phallus. The molecular phylogeny suggests the sister relationships of Platerodrilus maninjauensis and Platerodrilus tujuhensis. These species differ in colouration (Figs 7, 12) and the shape of the phallus and phallobase (Figs 56–57, 60–61). The female larvae of both species are similar and differ only in the relative size of mesonotal tubercles, which are smaller in Platerodrilus tujuhensis.
Body dark brown to black; head, pronotum and elytra yellow to orange, apical margins of elytra infuscate (Fig. 12). Head including eyes narrower than pronotum, antennal tubercles slightly prominent. Eyes hemispherically prominent, frontal interocular distance 2.6 times eye diameter. Antennae compressed, slender, length of antennomere 3 0.9 times antennomere 2. Pronotum transverse, 1.8 wider than long at midline, anterior margin widely rounded, anterior angles inconspicuous, posterior margin bisinuate (Fig. 27). Elytra 3.7 times longer than width at humeri, widest posteriorly, elytral costae inconspicuous. Legs slender, compressed with dense pubescence. Male genitalia with slender, curved phallus and small rounded parameres bearing slender membranous processes, phallobase wide, narrowly and deeply emarginate (Figs 56–57).
Female larva. Body flat, wide, dark brown, only margins of tergites lighter, pronotum triangular (Fig. 41), with two glabrous rounded tubercles postero-laterally, another two tubercles in middle of anterior margin; mesothorax strongly transverse, with rounded lateral margins and weakly projected posterior angles, laterally with four tubercles, upper rounded, lower transverse, metathorax similar in shape with more acutely projected posterior margins. Abdominal segments with slender and long lateral processes.
BL 8.1 mm, PL 0.9 mm, PW 1.6 mm, HW 1.9 mm, Edist 0.81 mm, Ediam 0.31 mm. Larva. BL 32.7 mm, PL 6.6 mm, PW 10.2 mm.
Indonesia: Sumatra.
The specific epithet refers to the locality of the paratype.
Holotype. Sumatra, Sungei Kumbang, Korinchi, 4500 ft, Apr. 1914 (BMNH). Other material examined. Male (LMBC, UPOL 001378), Sumatra, Jambi Kersik Tua, Gn. Kerinci, 19.–22. Jan. 2005, 1600–2200 m.
Platerodrilus robinsoni resembles Platerodrilus luteus in general appearance, but differs in the shape of the phallobase and phallus (Figs 62–63, 67–68).
Body black; head and mesoscutellum dark brown, pronotum and elytra orange yellow; antennae, and legs dark brown to black (Fig. 6). Head small, antennal tubercles separated by deep groove. Eyes hemispherically prominent, frontal interocular distance 2.1 times maximum eye diameter. Antennae slender, compressed, densely pubescent, antennomere 3 1.1 times longer than antennomere 2. Pronotum transverse, 1.6 times wider than long, anterior margin rounded, frontal angles obtuse, lateral margins slightly convex, posterior angles approximately rectangular (Fig. 19). Elytra parallel-sided, elytra 3.3 times longer than width at humeri, elytral costae weak, covered with dense pubescence. Male genitalia with curved phallus; parameres short, rounded, with slender membranous process; phallobase wide, deeply emarginate (Figs 62–63).
BL 7.8 mm, PL 1.0 mm, PW 1.6 mm, HW 2.0 mm, Edist 0.85 mm, Ediam 0.40 mm.
Indonesia: Sumatra.
Platerodrilus robinsoni differs from Platerodrilus korinchianus in the colouration; but both species are syntopic. Therefore, the species rank is proposed for Platerodrilus robinsoni stat. n.
Holotype, male, Presqu'ile Malaise (MNHP).
Platerodrilus testaceicollis was placed in Platerodrilus by Kazantsev, 2009, but the abdomen of the type is missing and the species cannot be placed in any species group unless further specimen is available. The redescription was given by
The type series contains just female larvae judging from the illustrations might include two species. The adult is unknown.
We are obliged to P. Ng and A. Wong for providing as with the manuscript of A. Wong's Master Thesis and for permission to publish all information. The authors are grateful to R. Bilkova for technical assistance, to M. Barclay for providing an access to the BMNH collection and to K. Matsuda for an access to additional material. The study was funded by the Grant Agency of the Czech Republic (P506/11/1757) and IGA Project from UP Olomouc. Additionally, this research received support from the Synthesys Project, which is financed by EC Research Infrastructure Action.