Monograph |
|
Corresponding author: Cornelis van Achterberg ( kees@vanachterberg.org ) Academic editor: Bernardo Santos
© 2020 Cornelis van Achterberg, Mark R. Shaw, Donald L. J. Quicke.
This is an open access article distributed under the terms of the Creative Commons Attribution License (CC BY 4.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Citation:
van Achterberg C, Shaw MR, Quicke DLJ (2020) Revision of the western Palaearctic species of Aleiodes Wesmael (Hymenoptera, Braconidae, Rogadinae). Part 2: Revision of the A. apicalis group. ZooKeys 919: 1-259. https://doi.org/10.3897/zookeys.919.39642
|
Abstract
The West Palaearctic species of the Aleiodes apicalis group (Braconidae: Rogadinae) as defined by van Achterberg & Shaw (2016) are revised. Six new species of the genus Aleiodes Wesmael, 1838, are described and illustrated: A. carbonaroides van Achterberg & Shaw, sp. nov., A. coriaceus van Achterberg & Shaw, sp. nov., A. improvisus van Achterberg & Shaw, sp. nov., A. nigrifemur van Achterberg & Shaw, sp. nov., A. turcicus van Achterberg & Shaw, sp. nov., and A. zwakhalsi van Achterberg & Shaw, sp. nov. An illustrated key to 42 species is included. Hyperstemma Shestakov, 1940, is retained as subgenus to accommodate A. chloroticus (Shestakov, 1940) and similar species. Fourteen new synonyms are proposed: Rogas bicolor Lucas, 1849 (not Spinola, 1808), Rogas rufo-ater Wollaston, 1858, Rhogas bicolorinus Fahringer, 1932, Rhogas reticulator var. atripes Costa, 1884, and Rhogas similis Szépligeti, 1903, of Aleiodes apicalis (Brullé, 1832); Rogas (Rogas) vicinus Papp, 1977, of Aleiodes aterrimus (Ratzeburg, 1852); Rogas affinis Herrich-Schäffer, 1838, of Aleiodes cruentus (Nees, 1834); Bracon dimidiatus Spinola, 1808, and Rhogas (Rhogas) dimidiatus var. turkestanicus Telenga, 1941, of Aleiodes gasterator (Jurine, 1807); Rogas alpinus Thomson, 1892, of Aleiodes grassator (Thunberg, 1822); Rhogas jaroslawensis Kokujev, 1898, of Aleiodes periscelis (Reinhard, 1863); Rhogas carbonarius var. giraudi Telenga, 1941, of Aleiodes ruficornis (Herrich-Schäffer, 1838); Ichneumon ductor Thunberg, 1822, of Aleiodes unipunctator (Thunberg, 1822); Rogas heterostigma Stelfox, 1953, of Aleiodes pallidistigmus (Telenga, 1941). Neotypes are designated for Rogas affinis Herrich-Schäffer, 1838; Rogas nobilis Haliday (in Curtis), 1834; Rogas pallidicornis Herrich-Schäffer, 1838; Rogas ruficornis Herrich-Schäffer, 1838. Lectotypes are designated for Rhogas (Rhogas) dimidiatus var. turkestanicus Telenga, 1941, and Rhogas hemipterus Marshall, 1897.
Keywords
Aleiodes apicalis group, key, new species, host range, biology, distribution, West Palaearctic, Europe, phenology
Introduction
In this 2nd part of a revision of western Palaearctic species of Aleiodes Wesmael we treat the group identified in Part 1 (
Specimens, methods, and presentation of records
The biological data from rearings of wild-collected hosts is in some cases supplemented by experimentation, and the protocols and means of scoring results are as outlined in
Overall, many of the species treated here have been widely misinterpreted in the literature and, as in Part 1 of our revision (
All available collections containing recently collected material of Aleiodes from the western Palaearctic region were used for our revision; collections with type material are separately listed under the description of the species. The following collections and acronyms are used:
AAC A.A. Allen Collection, Dawlish,
ALC A. Lozan Collection, Institute of Entomology, České Budĕjovice,
BZL Oberösterreichisches Landesmuseum, Biologiezentrum, Linz,
CC M. Čapek Collection, Moravian Museum, Brno,
CMIM C. Morley Collection, Ipswich Museum, Ipswich,
FC J.V. Falcó Collection, Valencia,
FRAH Forest Research, Alice Holt Lodge, Farnham,
HHC H. Haraldseide Collection,
HSC H. Schnee Collection,
IKC I. Kakko Collection,
KBIN Koninklijk Belgisch Instituut voor Natuurwetenschappen, Brussels,
JLC J. Lukáš Collection, Bratislava,
MRC M. Riedel Collection,
MSC M. Schwarz Collection, Linz,
MSNV Museo de Storia Naturale, Venice,
NNHM National Natural History Museum, Oslo,
RMNH Naturalis Biodiversity Center, Leiden,
SDEI Senkenberg Deutches Entomologisches Institut, Müncheberg,
SYKE Finnish Environment Institute, Friendship Park Research Centre, Kuhmo,
UNS Department of Biology and Ecology, University of Niš, Serbia,
WAE W.A. Ely Collection, Rotherham,
ZMC Zoological Museum, Copenhagen,
In addition, we have examined specimens from various smaller and private collections, which are cited in significant cases. Unless otherwise specified, reared material is in
The number of antennal (i.e., flagellar + 2) segments is frequently an important aid to species recognition and of interest also because in some species the female has more segments on average than the male (males have a greater number in other species, which is the normal condition seen in Braconidae). We give counts of antennal segments for the specimens we have examined, but for some species (especially when the segments did not need to be counted for determination) sometimes only for the first hundred or so of the specimens examined of each sex.
Attention has been paid to the apical tergites of males. The medial dorsal pores of A. fortipes (Reinhard), which are unique to this species within the A. apicalis group as treated here, are described and discussed in the entry for that species (note that the unknown male of A. caucasicus (Tobias) is likely to be similar). In the remainder of the species group there is either no evident modification, or a different development is evident to a greater or lesser extent. In some species specialised setae are present on tergites 4–6(7), presumably connected with pheromone dispersal from tergal glands. Broadly, two kinds of specialised setae can occur on these tergites. First, a fringe of short backwards-projecting setae (hereafter “fringe”), possibly associated with pores, originate from close to an apparent sulcus near the extreme base of the tergite (which is normally concealed). The presence and nature of the fringe varies between species, and even when present, it may not be visible in a given specimen owing to telescoping of the tergites. Second, there may be backwards-directed and more or less dense patches of longer setae (hereafter “setal patches”) on each side of the mid-line, the setae to some extent being adpressed in their anterior part but tending to be raised posteriorly (in extreme cases giving the tergites a concave appearance) and appearing different from the arrangement of setae on the more anterior tergites. A median glabrous area is left between the paired setal patches on each tergite, which collectively present as a glabrous and often shiny dorsal stripe along the length of these tergites (hereafter “glabrous stripe”). There is considerable variation in the extent to which these features are developed in the species keyed here, and indeed in some species they are scarcely present or wholly absent. In the species accounts given below we attempt to give a score from 1 to 4 for the development of the setal patches and glabrous stripe in males, with minimal elaboration (but including also mention of the setal fringe in cases for which we have been able to observe it). Type 1 = not at all developed, setae as on anterior tergites and evenly distributed. Type 2 = setal patches hardly developed, but glabrous stripe evident to some extent. Type 3 = setal patches clearly developed but relatively weak or sparse, glabrous stripe strong. Type 4 = setal patches strongly developed, making the tergites appear concave, glabrous stripe also strong. It should be borne in mind that there is some intraspecific variation, much of which may be artefactual (i.e., the condition of the specimen may make it hard to assess and score accurately).
For the recognition of braconid subfamilies, see

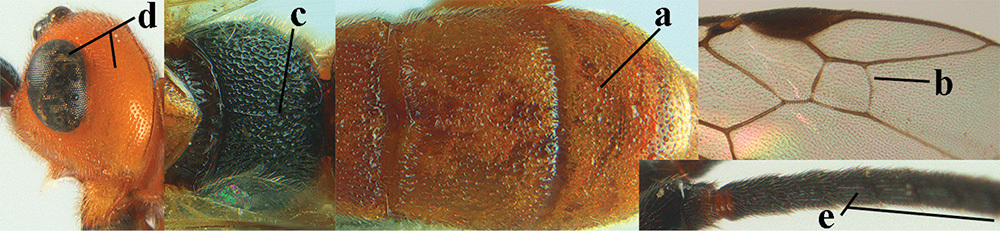
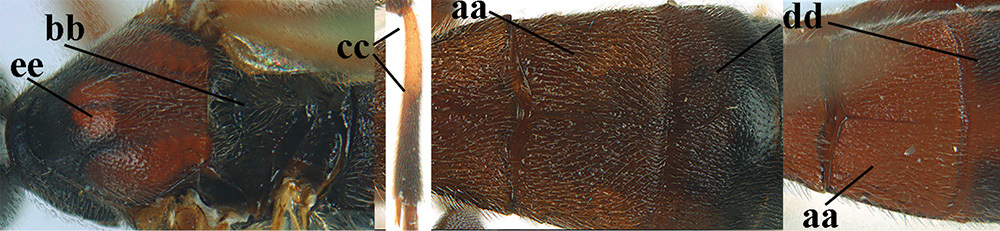
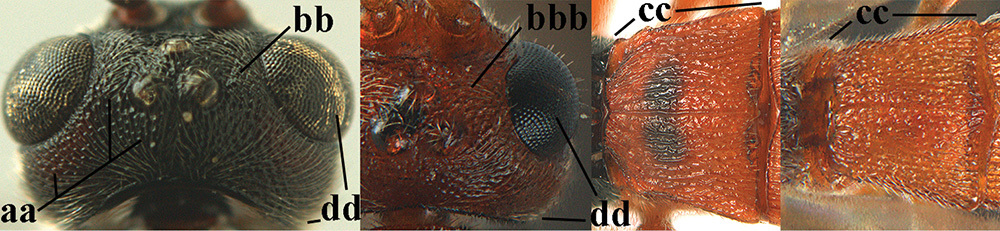
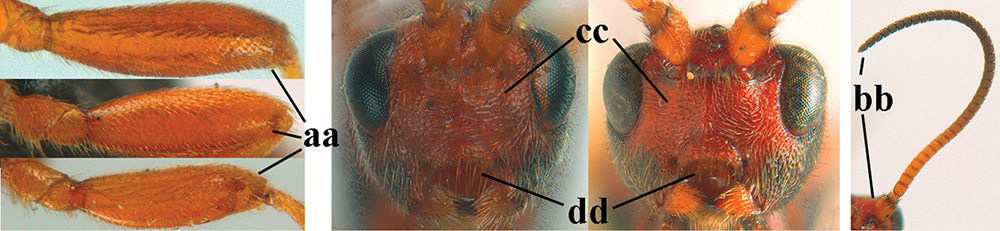
Terminology and measurements used in this paper 1 wing venation: pa = parastigma, pt = pterostigma, 1 = marginal cell, 2a, b, c = 1st, 2nd and 3rd submarginal cell, respectively, 3a, b = 1st and 2nd discal cell, respectively, 4a = 1st subdiscal cell, 5 = basal cell, 6 = subbasal cell 2 head, dorsal aspect: a = length of eye, b = length of temple 3 head, lateral aspect: c = width of temple, d = width of eye, e = height of eye, f = width of malar space (measured as actual true distance in its own plane) 4 head, anterior aspect: g = width of face, h = width of hypoclypeal depression 5 fore femur, lateral aspect: i = length, j = width 6 1st metasomal tergite, dorsal aspect: k = length of tergite (measured from adductor), l = apical width of tergite.
Molecular methods
A molecular dataset of the barcode region of cytochrome oxidase c subunit 1 (CO1) was compiled for a total of 141 Aleiodes specimens and three of Heterogamus (Fig.
GenBank accessions numbers are given in Appendix
Phylogeny
Three datasets were investigated with different levels of taxonomic and sequence inclusion.
Firstly, we conducted an overview analysis including representatives of a wide range of extra-limital species groups of Aleiodes, single representatives of the species treated in this paper for which molecular data were available (22 of the 42 species), and representatives of other West Palaearctic species groups, with three members of the genus Heterogamus used as outgroups (Fig.

Maximum likelihood tree based on DNA barcode sequence data for representatives of taxa included in this paper (‘Chelonorhogas’ group) together with data from additional West Palaearctic and extra-limital species showing broad picture of relationships. Terminal text show specimen voucher code and provenance (when known).
Secondly, we analysed a matrix comprising the most complete available sequence for each West Palaearctic species and using A. fortipes as the outgroup based on the results of the first analysis (Fig.

Thirdly, we constructed a tree for the available barcodes for the species treated in this paper (Fig.

Taxonomy
Aleiodes
Aleiodes
Wesmael, 1838: 194;
Petalodes
Wesmael, 1838: 123;
Schizoides Wesmael, 1838: 94. Unavailable name.
Nebartha
Walker, 1860: 310;
Tetrasphaeropyx
Ashmead, 1889: 634;
Neorhogas
Szépligeti, 1906: 605;
Chelonorhogas
Enderlein, [Sept. 1st] 1912a: 258;
Eucystomastax
Brues, [(end of?) Sept.] 1912: 223;
Leluthinus
Enderlein, 1912b: 96;
Aleirhogas
Baker, 1917b: 383, 411;
Hemigyroneuron
Baker, 1917a: 284, 322–327;
Heterogamoides
Fullaway, 1919: 43;
Cordylorhogas
Enderlein, 1920: 153;
Hyperstemma
Shestakov, 1940: 10;
Dimorphomastax
Shenefelt, 1979: 131–133;
Pholichora
van Achterberg, 1991: 48–53;
Arcaleiodes
Chen & He, 1997: 60–62;
Vietorogas
Long & van Achterberg, 2008: 313–314;
R (h)ogas auct; Tobias, 1971: 215–217 (transl. 1975: 83–86); Shenefelt, 1975: 1215–1256; Tobias, 1976: 81–89; Marsh, 1979: 179–181; Tobias, 1986: 74–84.
Notes
Hyperstemma Shestakov, 1940, is traditionally included in the genus Heterogamus Wesmael, 1838 (

Aleiodes apicalis
Diagnosis
Apical half of marginal cell of hind wing distinctly widened, its maximum width 1.6 × its width near hamuli or wider (Fig.
Biology
All species of the A. apicalis group for which host data exist are parasitoids of Noctuidae. However, the putatively more basal A. fortipes belonging to the Hemigyroneuron clade (see below) is a parasitoid of Geometridae. Also, only A. fortipes and A. sibiricus are known to parasitise hosts only in spring although these hosts would have been available in autumn of the previous year. Possibly others in the A. apicalis group will be found to do this too, and we consider the habit putatively as ancestral, in contrast with the more derived A. circumscriptus and A. bicolor groups in which species using hosts that overwinter as larvae invariably (as far as known) parasitise the host in the autumn and overwinter as a young larva inside it.
While we have no host data for a disappointingly large number of species of the A. apicalis group, the form of the clypeus may give important clues as to the site at which host mummification occurs, as those species in which mummification is known to take place in open situations (e.g., on a twig or in a leaf curl) invariably have a relatively small hypoclypeal depression and the clypeal margin blunt (A. apicalis, A. aterrimus, A. fortipes, A. nobilis, A. pulchripes, A. rugulosus) while species known to cause their hosts to mummify in concealed situations tend to have the hypoclypeal opening wider and the margin sharper (e.g., A. cruentus, A. dissector, A. ruficornis, A. sibiricus, A. unipunctator).
Notes
According to the 28S + COI analysis by
Key to West Palaearctic species of the Aleiodes apicalis group
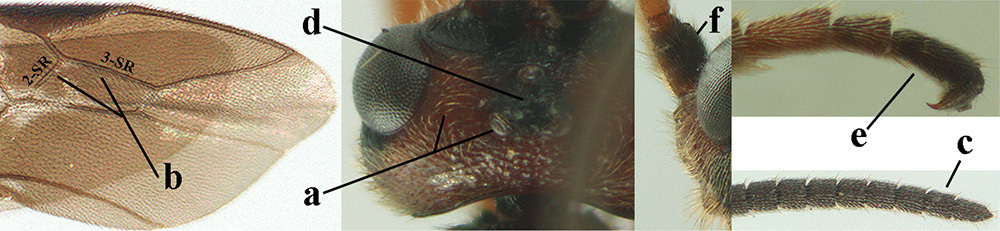
| 1 | Basal half of fore wing (except anteriorly) largely glabrous (a), or rather inconspicuously setose as remainder of wing; width of hypoclypeal depression 0.8–1.0 × minimum width of face (b) and anterior part of clypeus very narrow (c); vein r of fore wing 0.5–0.7 × vein 3-SR (d); mandibles massive triangular and coarsely punctate (e); [mandible with thick ventral lamella; antennal segments 47–63 and 4th segment of ♀ 1.1–1.3 × longer than wide; head (except clypeus and mandible) and mesosoma (except partly prothorax and mesoscutum) black; tarsal claws slender and only setose] | 2 |
 | ||
| – | Basal half of fore wing normally setose (except sometimes near veins) as remainder of wing (aa) or brachypterous (♀ A. hemipterus), if rarely with reduced setosity (A. venustulus) then width of hypoclypeal depression less than 0.7 × width of face (bb) and/or anterior part of clypeus moderately wide (cc), or vein r of fore wing 0.2–0.4 × vein 3-SR (dd); shape of mandible variable, often less massive and largely smooth (ee) | 3 |
 | ||
| 2 | Lateral lobes of mesoscutum whitish setose and with satin sheen (a); flagellum of antenna somewhat darker than scapus and pedicellus (b); middle lobe of mesoscutum distinctly punctate (c; more or less obscured by setosity); height of eye approx. 6 × length of malar space (d); basal half of metasoma dark brown, but laterally more or less yellowish (e) | A. agilis (Telenga, 1941) |
 | ||
| – | Lateral lobes of mesoscutum largely glabrous or sparsely setose and strongly shiny (aa); flagellum of antenna brownish yellow, similar to colour of scapus and pedicellus (bb); middle lobe of mesoscutum largely smooth (cc); height of eye nearly 7 × length of malar space (dd); basal half of metasoma brownish yellow, at most 1st tergite darker brown medio-basally (ee) | A. desertus (Telenga, 1941) |
 | ||
| 3 | Mesoscutum densely rugose or rugulose (a), with medio-longitudinal ridge or carina (b); mesopleuron mainly rugose (c); 3rd metasomal tergite densely sculptured (d) and convex posteriorly (e); propodeum angulate posteriorly (f); ♀ brachypterous and ♂ macropterous; N Africa | A. hemipterus (Marshall, 1897) |
 | ||
| – | Mesoscutum largely smooth and punctate or punctulate, mainly granulate or coriaceous (aa), usually without medio-longitudinal ridge or carina (bb); mesopleuron at most medially and antero-dorsally rugose (cc); 3rd metasomal tergite truncate posteriorly or nearly so (dd) and/or largely smooth posteriorly (ee); if mesopleuron largely sculptured (ccc) combined with 3rd tergite convex (ddd) and densely sculptured (eee) posteriorly, then propodeum rounded posteriorly (ff); both sexes macropterous | 4 |
 | ||
| 4 | Anterior part of clypeus short and subparallel-sided, near lower level of eyes (a) and hind femur slender (b); antenna with 65–72 segments and 5th–10th segments approx. as wide as long (c); tarsal claws slender (d) and without distinct pecten (e); marginal cell of hind wing slightly constricted subbasally (f) or subparallel-sided (fff below); [temple behind eye densely setose, convex and curved in dorsal view; 2nd–5th metasomal tergites more or less yellowish to reddish brown and head except mouthparts black] | A. sibiricus (Kokujev, 1903) |
 | ||
| – | Anterior part of clypeus medially distinctly wider than laterally (aa); if intermediate then partly above lower level of eyes or hind femur inflated (bb); antennal segments usually 62 or less, 4th–10th segments variable, often longer than wide (cc); tarsal claws often rather robust (dd), if slender (ddd) then either with distinct pecten (ee) and/or marginal cell of hind wing directly widened subbasally (ff) | 5 |
 | ||
| 5 | Temples extremely short (a), approx. 0.2 × as long as eye in dorsal view; basal half of marginal cell of hind wing parallel-sided (b) and pterostigma pale-yellowish or light brown (c); malar space slightly shorter than basal width of mandible (d); hind tibial spurs of ♂ obtuse apically (e); [OOL distinctly less than diameter of posterior ocellus; tarsal claws with coarse pecten; mesopleuron, mesosternum and scutellum brownish yellow] | A. pulchripes Wesmael, 1838 |
 | ||
| – | Temples medium-sized to long (aa), at least 0.3 × as long as eye in dorsal view; basal half of marginal cell of hind wing gradually widened (bb), if parallel-sided (bbb) then pterostigma dark brown (cc); malar space usually as long as basal width of mandible (dd) or longer; hind tibial spurs of ♂ usually acute apically (ee) | 6 |
 | ||
| 6 | Vein 2-SR+M of fore wing 0.8–1.0 × vein m-cu (a); pronotum and mesoscutum similarly coloured; clypeus width 0.3 × minimum width of face (b); length of fore wing 3.7–5.0 mm; length of hind femur 3.5–3.9 × its maximum width (c) and occipital carina reduced or anteriorly angled medio-dorsally (d); [4th–6th metasomal tergites of ♂ with setose round pits (but ♂ of A. caucasicus unknown); vein m-cu of fore wing more or less subvertical and relatively short; 3rd–10th antennal segments of ♀ pale yellowish, contrasting with entirely dark brown scapus] | 7 |
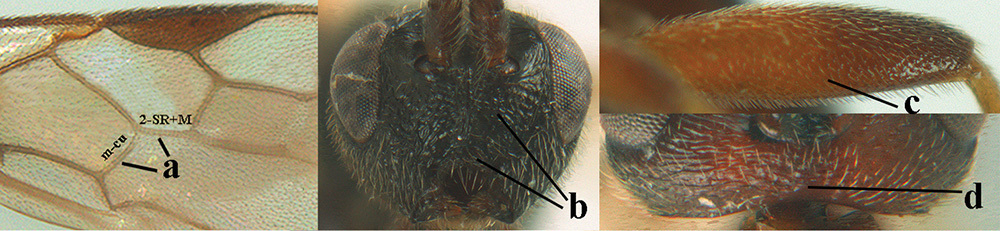 | ||
| – | Vein 2-SR+M of fore wing 0.2–0.6 × vein m-cu (aa), if more (some A. nobilis) then pronotum orange in contrast with blackish mesopleuron; clypeus width 0.4–0.8 × minimum width of face (bb); fore wing almost always longer than 4.9 mm; length of hind femur either more than 3.9 × its maximum width (cc) or occipital carina complete medio-dorsally (dd) | 8 |
 | ||
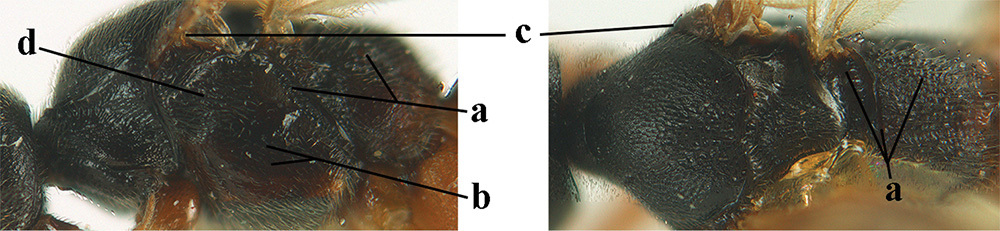
| 7 | Posterior half of mesosoma largely black or dark brown (a); precoxal area largely smooth, at most with some aciculae or punctures medially (b); tegulae brown (c); antero-dorsally mesopleuron coarsely rugose (d); [body of ♂ completely black and antenna completely blackish, dark brown or with some segments yellowish subbasally]; N & C Europe | A. fortipes (Reinhard, 1863) |
 | ||
| – | Posterior half of mesosoma largely yellowish brown (aa); precoxal area more or less vertically striate (bb); tegulae usually yellow (cc), but sometimes dark brown; mesopleuron antero-dorsally moderately rugose (dd); SE Europe [♂ unknown] | A. caucasicus (Tobias, 1976) |
 | ||
| 8 | Tarsal claws gradually narrowed submedially, slender and hardly curved (a) and 4th hind tarsal segment brownish yellow and 1.8–2.0 × as long as wide (c); clypeus yellowish brown, distinctly protruding anteriorly and ventrally thick (b); tarsal segments ventrally with long apical spiny bristles (d); [4th antennal segment of ♀ distinctly longer than wide; basal half of antenna and mesosoma anteriorly of ♀ largely yellowish brown, in ♂ more or less dark brown or infuscated; clypeus of ♂ yellowish and contrasting with black face] | A. schewyrewi (Kokujev, 1898) |
 | ||
| – | Tarsal claws more directly narrowed submedially, moderately robust and apically curved (aa); if slender (ccc) and hardly curved (aaa) then clypeus black, with thin ventral margin (bb) or 4th hind tarsal segment dark brown or infuscate and at most 1.5 × longer than its maximum width (cc) and tarsal segments ventrally with shorter apical bristles (dd) | 9 |
 | ||
| 9 | Vein 1-CU1 of fore wing 0.7–1.5 × as long as vein m-cu (a); if 0.7–0.9 × (A. aestuosus, A. zwakhalsi) then base of hind tibia yellowish dorsally (b), hind trochanter orange or yellowish (c) and pecten up to apical tooth of tarsal claw (d) | 10 |
 | ||
| – | Vein 1-CU1 of fore wing 0.3–0.8 × as long as vein m-cu (aa); if 0.7–0.8 × then base of hind tibia with dark brown patch dorsally (bb) or hind trochanter dark brown (cc) and in both cases pecten remaining removed from apical tooth of tarsal claw (dd) | 19 |
 | ||
| 10 | Pronotum orange (except antero-medially), distinctly contrasting with black posterior half of mesosoma in lateral view (a, rarely black) and 3rd–6th antennal segments of ♀ pale yellowish, contrasting with dorsally entirely dark brown scapus (b) and angle of vein m-cu of fore wing with vein 3-CU1 distinctly larger than 90° (c) and apex of hind femur black or dark brown (e); palpi yellow (d); [head black; hind basitarsus brownish yellow, contrasting with dark brown telotarsus] | 11 |
 | ||
| – | Pronotum black or reddish brown and less contrasting with posterior half of mesosoma in lateral view (aa); if pronotum orange brown and contrasting with dark posterior parts (A. venustulus) then 3rd–6th antennal segments of ♀ and scapus similarly dark brown (bb), angle of vein m-cu of fore wing with vein 3-CU1 closer to 90° (cc) and palpi dark brown (dd) or apex of hind femur yellowish or reddish brown (ee) | 12 |
 | ||
| 11 | Mesoscutum and scutellum black (a); temple rather mat and mainly granulate between punctulation (b); frons mat and strongly granulate (c); base of hind tibia pale yellowish (d); [precoxal area usually with some very superficial short rugulae or crenulae medially] | A. nobilis (Haliday, 1834) |
 | ||
| – | Mesoscutum and scutellum orange brown (aa); temple shiny and smooth between punctures, striae or rugae (bb); frons shiny and with distinct striae or rugae (cc); base of hind tibia more or less infuscate (dd) | A. schirjajewi (Kokujev, 1898) |
 | ||
| 12 | Tarsal claws without pecten near apical tooth (a); vein m-cu of hind wing absent (b); wing membrane subhyaline to slightly infuscate (c); [5th–10th antennal segments of ♀ distinctly longer than wide] | 13 |
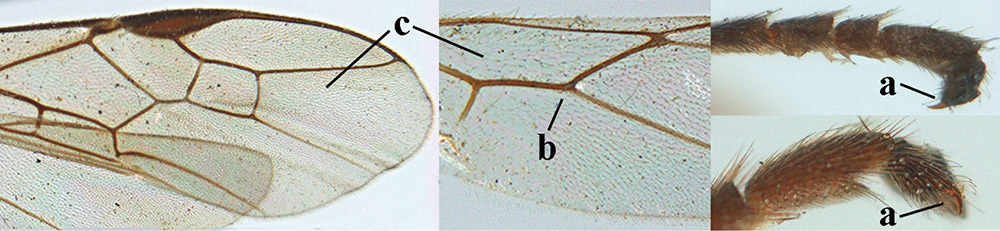 | ||
| – | Tarsal claws with pecten near apical tooth (aa); vein m-cu of hind wing at least weakly present (bb); wing membrane moderately infuscate or brownish (cc) | 14 |
 | ||
| 13 | Head of ♀ entirely yellowish brown or orange (a); ventral margin of clypeus thick and not protruding (b); vertex and OOL with smooth interspaces between punctures (c); mesopleuron remotely punctate and precoxal area coarsely punctate (d); 3rd tergite nearly flat in lateral view (e) and medio-posteriorly nearly truncate in dorsal view (f); [fore femur elongate and hind femur 4.3–4.7 × longer than wide; vertex at least partly densely punctate] | A. venustulus (Kokujev, 1905) |
 | ||
| – | Head of ♀ largely black (aa); ventral margin of clypeus thin and protruding anteriorly (bb); vertex and OOL without distinct smooth interspaces, rugose (cc); mesopleuron very densely and coarsely punctate and precoxal area rugose-punctate (dd); 3rd tergite convex in lateral view (ee) and medio-posteriorly convex in dorsal view (f); [3rd tergite coarsely punctate; fore and middle femora with dark patch] | A. krulikowskii (Kokujev, 1898) |
 | ||
| 14 | Head of ♀ entirely yellowish brown or orange (a); ventral margin of clypeus thin and protruding anteriorly (b); eye 0.8–1.2 × temple in dorsal view (c); apical third of metasoma of ♀ completely yellowish (d; but ♂ often with 1st tergite partly and 4th–6th tergites blackish); hind femur of ♀ distinctly inflated (e), but sometimes less so; [antenna of ♀ with 49–56 segments; hind tibia of ♀ ivory except dark brown apex] | A. aestuosus (Reinhard, 1863) |
 | ||
| – | Head of ♀ black (aa); ventral margin of clypeus thick and hardly protruding anteriorly (bb); eye 1.0–1.9 × temple in dorsal view (cc); apical third of metasoma of ♀ black (dd); hind femur of ♀ slender to moderately wide (ee) | 15 |
 | ||
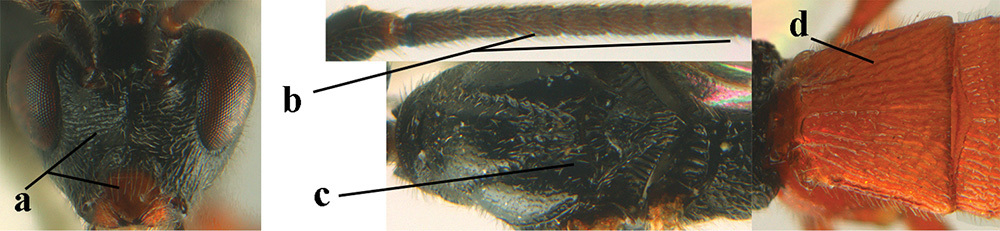
| 15 | Maximum width of hypoclypeal depression 0.3–0.4 × minimum width of face (a); 5th–10th antennal segments of ♀ distinctly longer than wide (b); posterior half of mesoscutum black (c); 1st metasomal tergite robust (d); [surroundings of veins M+CU1 and 1-+2-CU1 largely setose; vein M+CU of hind wing distinctly longer than vein 1-M; apical fifth of hind femur always blackish; 4th–6th metasomal tergites of ♂ appearing concave and with conspicuous setosity]; C Europe, Mediterranean area, Central Asia. Examined specimens from S England ( |
A. apicalis (Brullé, 1832) |
 | ||
| – | Maximum width of hypoclypeal depression 0.5–0.7 × minimum width of face (aa); 5th–10th antennal segments of ♀ approx. as long as wide (bb); posterior half of mesoscutum at least partly red (cc); 1st metasomal tergite rather slender (dd); [surroundings of veins M+CU1 and 1-+2-CU1 largely glabrous; vein r of fore wing 0.3–0.4 × vein 3-SR] | 16 |
 | ||
| 16 | Female: 2nd metasomal tergite of ♀ as long as wide basally (a; of ♂ 0.9 ×); 1st tergite only slightly widened posteriorly and 1.3–1.4 × as long as wide posteriorly (b; of ♂ 1.2 ×); 3rd tergite largely smooth basally, only sparsely punctulate (c; of ♂ rugose); OOL distinctly rugose anteriorly (d); [hind femur ca 4.5 × longer than wide; ♂ may be easily confused with A. cruentus] | A. quadrum (Tobias, 1976) |
 | ||
| – | Both sexes: 2nd tergite of ♀ 0.7–0.9 × as long as wide basally (aa); 1st tergite distinctly widened posteriorly and 1.0–1.1 × as long as wide posteriorly (bb); 3rd tergite distinctly punctate or punctate-rugulose medio-basally (cc); OOL usually densely and coarsely punctate anteriorly (dd), rarely striate or rugose (ddd), but less sculptured in males and in A. zwakhalsi | 17 |
 | ||
| 17 | Ocelli medium-sized to large (a), OOL of ♀ 0.5–0.8 × diameter of posterior ocellus, rarely up to 1.0 times; length of eye 1.5–1.9 × temple in dorsal view (b); 1st and 2nd metasomal tergites at least partly reddish or orange brown (c); [hypoclypeal depression usually 0.6–0.7 × width of face; hind femur 3.1–4.0 × longer than wide. If hind femur is 5 × longer than wide and hypoclypeal depression 0.5 × width of face, cf. A. parvicauda (Tobias, 1985) from Afghanistan] | A. cruentus (Nees, 1834) |
 | ||
| – | Ocelli smaller (aa), OOL of ♀ 0.9–1.2 × diameter of posterior ocellus; length of eye 1.0–1.3 × temple in dorsal view (bb); 1st and 2nd metasomal tergites entirely black or dark brown (cc) | 18 |
 | ||
| 18 | Vein 1-CU1 of fore wing distinctly shorter than vein m-cu (a); hind femur 4.0–4.2 × longer than wide (b); vein cu-a inclivous and parallel with vein 3-CU1 (c); 5th–10th antennal segments of ♀ as long as wide (d); vertex and OOL remotely punctate (e); width of hypoclypeal depression 0.7 × minimum width of face (f); [1st metasomal tergite slender and rounded latero-basally; 3rd tergite densely punctulate basally; metasoma of ♀ strongly compressed posteriorly; if body completely black, precoxal sulcus extensively rugose, pterostigma medially pale brown and OOL densely rugulose, cf. A. morio (Reinhard)] | A. zwakhalsi sp. nov. |
 | ||
| – | Vein 1-CU1 of fore wing approx. as long as vein m-cu (aa); hind femur 3.0–3.3 × longer than wide (bb); vein cu-a vertical and vein 3-CU1 diverging posteriorly (cc); 5th–10th antennal segments of ♀ shorter than wide (dd); vertex and OOL moderately to densely punctate (ee); width of hypoclypeal depression 0.5–0.7 × minimum width of face (ff); [metasoma of ♀ less compressed posteriorly; ovipositor sheath rather robust] | A. diversus (Szépligeti, 1903) |
 | ||
| 19 | Third metasomal tergite largely coarsely punctate and yellowish brown (a); 2nd submarginal cell of fore wing short and square (b); medio-longitudinal carina at least in middle part of propodeum absent, obsolescent or incomplete (c); eye much narrower than temple in lateral view (d) and 4th–10th antennal segments of ♀ distinctly longer than wide (e); [clypeus distinctly protruding in lateral view and ventrally thin (Fig. |
A. miniatus (Herrich-Schäffer, 1838) |
 | ||
| – | Third tergite rugose, striate, rugulose or smooth, if punctate then black (aa); 2nd submarginal cell longer than high (bb); medio-longitudinal carina of posterior half of propodeum complete or nearly so (cc); eye usually approx. as wide as temple in lateral view (dd), if distinctly narrower (ddd) then 4th–10th antennal segments of ♀ approx.as long as wide (ee) | 20 |
 | ||
| 20 | Pecten of hind tarsal claws of ♀ robust (a), close to apical tooth (b) and often dark brown or blackish (c); [pecten of ♂ sometimes less developed than in ♀ (e.g., of A. periscelis) but then with some robust teeth medially (aaa)] | 21 |
 | ||
| – | Pecten of hind tarsal claws absent or inconspicuous (aa), if present then remaining removed from apical tooth (bb) and often yellowish or brownish (cc), but dark brown in A. hirtus | 26 |
 | ||
| 21 | Ventral margin of [anterior part of] clypeus comparatively sharp (a), clypeus more or less protruding anteriorly (b); palpi yellowish (c); vein 1-M of fore wing dark brown (d); basal half of metasoma weakly sculptured (e); hind femur largely or completely reddish or brownish (f); width of hypoclypeal depression 0.6–0.7 × minimum width of face (g); [precoxal area completely smooth or nearly so; length of malar space 0.2 × length of eye in lateral view; outer side of posterior ocellus with deep groove; vertex flattened. If palpi black or dark brown, temple with long setae, width of hypoclypeal depression 0.5 × minimum width of face, OOL more than diameter of posterior ocellus, 1st tergite coarsely sculptured, and length of malar space 0.40–0.45 × length of eye in lateral view, cf. A. hirtus (Thomson)] | A. dissector (Nees, 1834) |
 | ||
| – | Ventral margin of clypeus (rather) obtuse apically (aa) and clypeus hardly protruding anteriorly (bb); palpi dark brown at least basally (cc) or vein 1-M of fore wing yellowish brown (dd; A. rugulosus); basal half of metasoma distinctly sculptured (ee); hind femur dark brown or black dorso-apically (ff; but yellowish in A. rugulosus); width of hypoclypeal depression usually 0.3–0.4 × minimum width of face (gg) | 22 |
 | ||
| 22 | Only apical two fifths of marginal cell of hind wing distinctly widened and remainder parallel-sided or nearly so (a), rarely hardly widened apically; vertex flattened behind ocelli (b) and apex of hind femur yellowish or reddish (c); first metasomal tergite with coarse sublongitudinal rugae (d); ovipositor sheath distinctly narrowed apically (e); [vein 1-M of fore wing yellowish brown; basal half of hind tibia pale yellowish/ivory or orange and its apical half black; mesopleuron nearly or completely smooth medio-ventrally; propodeum with pair of crest-like protuberances laterally] | A. rugulosus (Nees, 1811) |
 | ||
| – | At least apical half of marginal cell of hind wing gradually widened (aa); if less distinctly so, then vertex declivous behind ocelli (bb) and hind femur black or dark brown apically (cc); 1st tergite moderately striate, rugulose or vermiculate-rugose (dd); ovipositor sheath truncate apically or nearly so (ee) | 23 |
 | ||
| 23 | Second metasomal tergite of both sexes black (a); scutellum densely and finely coriaceous (b); hind tibia largely black (c), dorsally paler at extreme base; 3rd metasomal tergite of ♀ mainly punctate (d), but basal half more or less rugose in ♂; mesoscutum with satin sheen (e); [vein 2-SC+R of hind wing subquadrate or vertical; 4th–6th tergites of ♂ with medium-sized dense setosity and with narrow glabrous central stripe; mesosternal sulcus shallow, obsolescent or absent. If hind tibia completely dark brown basally and temple roundly narrowed in dorsal view, cf. A. sapporensis (Watanabe) from East Palaearctic region] | A. aterrimus (Ratzeburg, 1852) |
 | ||
| – | Second tergite of ♀ yellowish or dark reddish brown (aa; up to almost black in ♂ of A. periscelis); scutellum partly smooth and punctate (bb); basal half of hind tibia (largely) pale yellowish or ivory (cc), rarely brownish; 3rd tergite largely rugulose-striate basally (dd); mesoscutum rather shiny (ee) | 24 |
 | ||
| 24 | Antennal segments of ♀ 39–45 (of ♂ 50–56) and subbasal segments of ♀ yellowish (a; of ♂ darkened but basal half of hind tibia ivory); fore femur of ♀ more robust (b); antenna of ♀ robust (c), 0.8–1.0 × longer than fore wing (= 0.7–0.8 × body length); fore coxa dark brown (d); vertex of ♀ coarsely rugose laterally (e); [mandible blackish basally; fore femur 4.8 × as long as wide] | A. periscelis (Reinhard, 1863) |
 | ||
| – | Antennal segments of both sexes 52–62 and subbasal segments dark brown or blackish (aa); basal half of hind tibia of ♂ reddish to dark brown; fore femur of ♀ slenderer (bb); antenna of ♀ elongate (cc), 1.0–1.1 × longer than fore wing; fore coxa (brownish) orange (dd); vertex punctate-rugulose to coriaceous laterally (ee) | 25 |
 | ||
| 25 | Mesoscutum largely matt (a); base of fore femur, fore trochanter and trochantellus at least partly dark brown or infuscate (b); 2nd and 3rd metasomal tergites comparatively slender (maximum width of 2nd tergite ca 1.5 × its median length; c); OOL of ♀ 0.9–1.1 × longer posterior ocellus (d); posterior half of hind femur blackish dorsally (e) | A. coriaceus sp. nov. |
 | ||
| – | Mesoscutum rather shiny (aa); base of fore femur, fore trochanter and trochantellus yellowish brown (bb); 2nd and 3rd metasomal tergites robust (maximum width of 2nd tergite ca 1.6 × its median length; cc); OOL of ♀ 1.1–1.5 × longer posterior ocellus (dd); posterior half of hind femur partly yellowish brown dorsally (ee) | A. rufipes (Thomson, 1892) |
 | ||
| 26 | Length of malar space of ♀ 0.45–0.70 × height of eye (a) and clypeus below lower level of eye in lateral view (b), if intermediate (in A. ruficornis) then basal antennal segments of ♀ very short (4th segment approx. as long as wide; c); lateral lobes of mesoscutum mainly smooth, (rather) densely punctate, punctulate or rugose-punctate, interspaces (as far as present) between punctures usually largely smooth and shiny (d), but sometimes distinctly granulate; marginal cell of fore wing of ♀ usually robust and ending further removed from wing apex (e); [wing membrane more or less infuscate; precoxal area coarsely vermiculate-rugose medially; hind femur at least apico-dorsally dark brown or black; maximum width of hypoclypeal depression usually 0.3–0.4 × minimum width of face, if 0.5 × then ventral margin of clypeus thick; vein 1-R1 of fore wing 1.0–1.2 × length of pterostigma] | 27 |
 | ||
| – | Length of malar space of ♀ 0.20–0.45 × height of eye (aa) and clypeus near lower level of eye in lateral view (bb); basal antennal segments of ♀ usually moderately slender (with 4th segment distinctly longer than wide; cc); lateral lobes of mesoscutum finely granulate, punctulate or moderately punctate, and often with a satin sheen (dd), but sometimes shiny (A. hirtus); marginal cell of fore wing of ♀ slender and ending closer to wing apex (ee), except in A. morio (eee); [wing membrane usually subhyaline; basal half of hind tibia largely pale yellowish or reddish, but less so in A. morio; vein 1-R1 of fore wing usually at least 1.4 × length of pterostigma, but approx. equal in A. morio (eee)] | 35 |
 | ||
| 27 | Area between ocellus and eye, vertex and temple sparsely punctate (a); head of ♀ entirely brownish yellow (b; of ♂ variable, at least stemmaticum black); 1st metasomal tergite 1.5–1.7 × wider posteriorly than subbasally (c); length of eye 1.0–1.2 ×temple in dorsal view (d) | 28 |
 | ||
| – | Area between ocellus and eye, vertex and temple at least moderately densely punctate or rugose (aa); head of ♀ black (bb) or more or less dark red (bbb; of ♂ black); 1st tergite 1.3–1.4 × wider posteriorly than subbasally (cc); length of eye 1.2–1.4 × temple in dorsal view (dd) | 29 |
 | ||
| 28 | Eye small (a) and in lateral view maximum width of temple 1.5–1.6 × maximum width of eye (b); ventral margin of clypeus thin and protruding anteriorly (c); antennal segments of ♀ 45–47 (of ♂ 56–58); mesoscutum of ♀ comparatively convex (d); [apex of hind tibia and basal part of palpi of ♀ dark brown] | A. ruficeps (Telenga, 1941) |
 | ||
| – | Eye medium-sized (aa) and in lateral view temple hardly wider than eye (bb); ventral margin of clypeus thick and hardly protruding anteriorly (cc); antennal segments of ♀ 35–37; mesoscutum of ♀ less convex (dd); [apex of hind tibia and palpi of ♀ yellowish brown] | A. arnoldii (Tobias, 1976) |
 | ||
| 29 | Area between posterior ocellus and eye moderately punctate (a); posterior half of notauli shallow (b); head in anterior view rather trapezoid (c); [antenna of ♀ 1.1–1.2 × fore wing; 4th antennal segment of ♀ moderately robust; pterostigma blackish; antennal segments of ♀ approx. 47] | A. turcicus sp. nov. |
 | ||
| – | Area between posterior ocellus and eye densely (and finely) rugose (aa), sometimes superficially so and rugulose or with some punctures; posterior half of notauli deep (bb); head in anterior view less trapezoid (cc); [antenna of ♀ with 30–47 segments; if antenna of ♀ with 54–64 segments, cf. A. ferrugiteli (Shenefelt, 1975) from C. Asia] | 30 |
 | ||
| 30 | Fore femur of ♀ subparallel-sided and 3.9–4.0 × longer than wide (a; this character is less reliable for ♂); antenna of ♀ 0.8–0.9 × fore wing (b); hypoclypeal depression usually slightly wider, 0.45–0.50 × minimum width of face (c); head of ♀ largely black (d), rarely face partly reddish; antennal segments of ♂ 36–46(–51) (usually 39–44); [tegulae usually (partly) dark brown; antennal segments of ♀ 29–41] | 31 |
 | ||
| – | Fore femur of ♀ inflated and 3.0–3.6 × longer than wide (aa); antenna of ♀ 0.9–1.2 × fore wing (bb); hypoclypeal depression usually narrower, ca 0.40 × minimum width of face (cc); head of ♀ at least partly reddish brown (dd); antennal segments of ♂ 47–63 (usually 48–54); [pale males have whole frons and stemmaticum yellowish; palpi dark brown or blackish, rarely brown; OOL of ♂ 1.5–2.0 × diameter of ocellus] | 32 |
 | ||
| 31 | Antennal segments of ♀ ca 41; subbasal antennal segments of ♀ dark brown and robust, with 4th segment as long as wide (a); basal half of 3rd tergite entirely coarsely striate (b; of ♂ sometimes with curved striae posteriorly); hind trochanter and trochantellus largely dark brown (c); inner and dorsal side of hind tibia apically dark brown (d); parastigma mostly brown (e); vein 1-CU1 of fore wing slightly longer than vein cu-a (f); [palpi dark brown, if largely ivory, cf. A. periscelis; 3rd tergite only anteriorly reddish or yellowish; marginal cell of ♂ wide (Fig. |
A. improvisus sp. nov. |
 | ||
| – | Antennal segments of ♀ 29–39; subbasal antennal segments of ♀ yellow and comparatively slender, with 4th segment ca 1.2 × as long as wide (aa); 3rd tergite weakly sculptured, with (faint) curved or antero-medially transverse rugulae or striae (bb) or largely smooth (but sometimes with basal longitudinal striae laterally and often with distinct punctures laterally); hind trochanter and trochantellus yellowish or reddish brown (cc); inner and/or dorsal side of hind tibia (largely) yellowish or red apically (dd); parastigma mostly yellowish (ee); vein 1-CU1 distinctly longer than vein cu-a (ff); [palpi usually brownish or yellowish, but sometimes dark brown; pale males nearly always have frons medially and stemmaticum black; hind tibial spurs of male are usually blunt apically]; Mediterranean, C. Asia | A. gasterator (Jurine, 1807) |
 | ||
| 32 | OOL of ♀ approx. 2.6 × diameter of posterior ocellus (a); vein 3-SR of fore wing of ♀ 1.7–2.0 × vein 2-SR (b; of ♂ 1.2–1.5 ×); penultimate antennal segment of ♂ ca 1.2 × longer than wide (c); stemmaticum of ♀ usually black or dark brown (d), rarely reddish; telotarsi of ♀ dark brown (e); scapus of ♀ often black dorsally (f); [antenna of ♂ 0.9 × as long as body; inner side of hind tibia of ♀ dark brown apically] | A. carbonarius Giraud, 1857 |
 | ||
| – | OOL of ♀ 1.4–2.3 × diameter of posterior ocellus (aa); vein 3-SR of fore wing of ♀ 1.5–1.6 × vein 2-SR (bb; of ♂ 1.0–1.4 ×); penultimate antennal segment of ♂ approx. as long as wide (cc); stemmaticum of ♀ yellowish brown or reddish (dd); telotarsi of ♀ yellowish brown or reddish (ee); scapus of ♀ variable, brownish yellow dorsally (ff) to blackish | 33 |
 | ||
| 33 | Length of eye 1.5–2.0 × temple in dorsal view (a; if measured with posterior ocelli up to posterior level of eyes); OOL of ♀ 1.2–1.8 × diameter of posterior ocellus (b); subbasal antennal segments of ♀ slightly less moniliform (c); [inner side of hind tibia of ♀ usually dark brown or blackish apically; colour of legs of ♂ usually similar to legs of ♀ and usually partly yellowish; antenna of ♂ approx. as long as body and 1.2–1.4 × fore wing; antennal segments of ♀ 34–39(–47), of ♂ (44–)47–60] | A. ruficornis (Herrich-Schäffer, 1838) |
 | ||
| – | Length of eye 1.1–1.5 × temple in dorsal view (aa; if measured with posterior ocelli up to posterior level of eyes); OOL of ♀ usually 1.9–2.3 × diameter of posterior ocellus (bb); subbasal antennal segments of ♀ distinctly submoniliform (cc); [inner side of hind tibia of ♀ yellowish; hind femur and basitarsus of both sexes more robust (but in ♂ sometimes rather slender); legs of males of W. European specimens strongly infuscate, darker than legs of females, but legs of N. European specimens paler; antenna of ♂ 0.8–0.9 × body and 1.0–1.1 × fore wing; antennal segments of ♀ 35–45(–46), of ♂ (44–)47–61] | 34 |
 | ||
| 34 | Apical tooth of hind tarsal claws of ♀ robust (a); 2nd metasomal tergite of ♂ orange brown (b); hind femur (c), tibia and basitarsus (d) of ♂ more robust and femur basally yellowish brown (e), if rarely almost black then hind tibia basally yellowish brown (f); clypeus less protruding in front of face (g); boreal and highland species | A. grassator (Thunberg, 1822) |
 | ||
| – | Apical tooth of hind tarsal claws of ♀ slender (aa); 2nd metasomal tergite of ♂ black (bb); hind femur (cc), tibia and basitarsus (dd) of ♂ comparatively slender and femur basally black (ee); hind tibia basally black (ff); clypeus more protruding in lateral view (gg); lowland species | A. carbonaroides sp. nov. |
 | ||
| 35 | Head brownish yellow (a); ventral margin of clypeus thin and distinctly protruding anteriorly (b); vertex (c) and mesoscutum (d) shiny; maximum width of hypoclypeal depression 0.6–0.7 × minimum width of face (e); pterostigma brownish yellow (f); [tarsal claws medium-sized and yellowish pectinate; body laterally and dorsally (except more or less dark brown propodeum and 1st tergite) yellowish brown; occipital carina weakly indicated medio-dorsally] | A. fahringeri (Telenga, 1941) |
 | ||
| – | Head black (aa); ventral margin of clypeus thick and usually hardly protruding (bb); vertex (cc) and mesoscutum (dd) usually rather dull and with satin sheen (cc); if shiny (ccc) then maximum width of hypoclypeal depression 0.5–0.6 × minimum width of face (ee) and pterostigma dark brown (ff); [pterostigma yellowish (fff) in A. pallidistigmus] | 36 |
 | ||
| 36 | Vertex (a) and mesoscutum (b) distinctly shiny because of smooth interspaces between punctures or rugae; head conspicuously setose because of long setae (c; less distinctive in ♀), as are propodeum and first tergite laterally; trochanters (and often also trochantelli) nearly always at least somewhat infuscate, darker than orange part of femora (d); subbasal antennal segments of ♀ dark brown; [palpi blackish or dark brown; tarsal claws with small pecten; 3rd–6th antennal segments robust, hardly longer than wide; fore femur slender] | A. hirtus (Thomson, 1892) |
 | ||
| – | Vertex (aa) and mesoscutum (bb) rather dull and with satin sheen, interspaces finely coriaceous-granulate between punctures or rugulae; head usually less conspicuously setose (cc); if sculpture and setosity are intermediate then trochanters and trochantelli have same colour as basal part of femora (dd) and subbasal antennal segments of ♀ brownish yellow | 37 |
 | ||
| 37 | Vein cu-a of fore wing at least as long as vein 1-CU1 (a); 1st tergite of ♀ strongly widened apically (b); marginal cell of fore wing shorter (c); subbasal antennal segments of ♀ subquadrate (d); hind coxa (as femur) completely black (e); clypeus distinctly protruding anteriorly (f); fore femur largely dark brown (g); [OOL distinctly longer than diameter of posterior ocellus; 2nd tergite finely and densely sculptured; if fore and hind femora yellowish brown, clypeus hardly protruding and marginal cell of fore wing elongate, cf. A. sibiricus (Kokujev)] | 38 |
 | ||
| – | Vein cu-a of fore wing distinctly shorter than vein 1-CU1 (aa); 1st tergite moderately widened apically (bb); marginal cell of fore wing long (cc); subbasal antennal segments of ♀ longer than wide (dd); hind coxa orange or yellowish brown (ee); clypeus hardly or not protruding anteriorly (ff); fore femur brownish yellow (gg) | 39 |
 | ||
| 38 | Wings infuscate apically (a); 2nd submarginal cell of fore wing less widened posteriorly (b); pterostigma medially dark brown (c); basal 0.4 of hind tibia yellowish (d); 2nd–5th metasomal tergites orange brown (e); middle lobe of mesoscutum densely punctate, without distinct granulation in between punctures (f); [vein 2-1A of hind wing comparatively long; pecten of tarsal claws present and claws rather robust] | A. nigrifemur sp. nov. |
 | ||
| – | Wings subhyaline apically (aa); 2nd submarginal cell of fore wing widened posteriorly (bb); pterostigma medially pale or yellowish brown (cc); hind tibia mainly dark brown, only basally narrowly pale yellowish (dd); 2nd–5th tergites black (ee); middle lobe of mesoscutum coriaceous (ff); [basal half of 3rd tergite and OOL rugulose] | A. morio (Reinhard, 1863) |
 | ||
| 39 | Apex of hind tibia reddish or yellowish (a); pterostigma brownish yellow medially (b), rarely darkened; 3rd tergite dull (c); [antenna of ♀ with 54–64 segments] | A. pallidistigmus (Telenga, 1941) |
 | ||
| – | Apex of hind tibia dark brown or infuscated (aa); pterostigma more or less dark brown medially (bb); 3rd tergite usually shiny (cc), but sometimes rather dull (ccc) | 40 |
 | ||
| 40 | Tarsal claws with medium-sized pecten (a); precoxal area of mesopleuron smooth medially, but sometimes with sparse weak punctures or some rugae below it (b); temple shiny (c); basal half of antenna of ♀ largely yellowish brown (d); [tegula and humeral plate equally yellowish orange; hind tarsus partly yellowish or brownish; hind tibia dorsally dark brown at extreme base, then pale subbasally and infuscate apically] | A. pallidicornis (Herrich-Schäffer, 1838) |
 | ||
| – | Tarsal claws without pecten (aa) or with fine pale pecten; precoxal area of mesopleuron moderately rugose medially (bb); temple rather dull (cc); basal half of antenna blackish (dd) | 41 |
 | ||
| 41 | Area between posterior ocellus and eye mainly granulate or coriaceous, at most with some punctures or rugulae (a); eyes larger in lateral view (b) and less protuberant in dorsal view (c); 3rd metasomal tergite usually largely smooth, especially in ♀ (but basal half in ♂ sometimes extensively striate-rugulose) and as strongly glossy as following tergite (d); precoxal area comparatively narrow and posteriorly largely or completely smooth (e); antennal segments of ♀ 47–57(–58); hind femur largely reddish apically, with only slight infuscation (f); N & C Europe | A. unipunctator (Thunberg, 1822) |
 | ||
| – | Area between posterior ocellus and eye densely rugose, coarsely punctate or rugulose (aa); eyes smaller in lateral view (bb) and more protuberant in dorsal view (cc); basal half of 3rd tergite distinctly striate or densely rugulose and less shiny (dd), but intermediates occur; precoxal area comparatively wide and usually posteriorly rugose or distinctly punctate (ee); antennal segments of ♀ 54–62; hind femur apically more or less smudged dark brown or black (ff), but sometimes very indistinct; [if mesoscutum rather steep anteriorly and width of hypoclypeal depression 0.4–0.5 × minimum width of face, and long malar space, cf. A. gasterator (Jurine) and related spp.]; S & E Palaearctic | A. eurinus (Telenga, 1941) |
 | ||
Biology and descriptions
Aleiodes aestuosus
Rogas aestuosus
Reinhard, 1863: 265;
Rhogas aestuosus;
Aleiodes (Neorhogas) aestuosus;
Aleiodes (Chelonorhogas) aestuosus;
Aleiodes aestuosus;
Rhogas (Rhogas) aestuosus var. desertus Telenga, 1941: 152–153, 404 (not Rhogas (R.) desertus Telenga, 1941).
Type material
Holotype, ♀ (
Additional material
Albania, Bulgaria, Cyprus, Greece, Russia, Turkey, Tunisia, [Azerbaijan, Georgia, Iran, Iraq, Israel, Jordan, Syria, Turkmenistan, Uzbekistan]. Specimens in
Molecular data
MRS004 (Turkey).
Biology
Collected March–July, often at light, but it is not clear how many generations are represented nor how the winter is passed. Reared from Noctuidae: Heliothis peltigera (Denis & Schiffermüller) (4 [1
Diagnosis
Maximum width of hypoclypeal depression 0.6–0.7 × minimum width of face (Fig.
Description
Redescribed ♀ (RMNH) from Turkey (Icil). Length of fore wing 6.8 mm, of body 8.3 mm.
Head.
Antennal segments of ♀ 52, length of antenna 1.1 × fore wing, its subapical segments approx. as long as wide; frons with irregular curved rugae, shiny, and rugose behind antennal sockets; OOL 2.4 × diameter of posterior ocellus, and finely remotely punctate, interspaces much larger than diameter of punctures; vertex spaced punctate, shiny; clypeus short, coarsely and densely punctate; ventral margin of clypeus thick and rather protruding forwards (Fig.
Mesosoma. Mesoscutal lobes largely smooth, shiny, sparsely and finely punctate; prepectal carina medium-sized, reaching anterior border; precoxal area of mesopleuron and metapleuron remotely punctate, interspaces much wider than diameter of punctures, shiny; mesopleuron above precoxal area (except speculum) sparsely punctate; scutellum slightly convex, remotely punctate and evenly rounded laterally, no carina; propodeum evenly convex and coarsely rugose, medio-longitudinal carina complete, but irregular posteriorly, without tubercles.
Wings.
Fore wing: r 0.4 × 3-SR (Fig.
Legs.
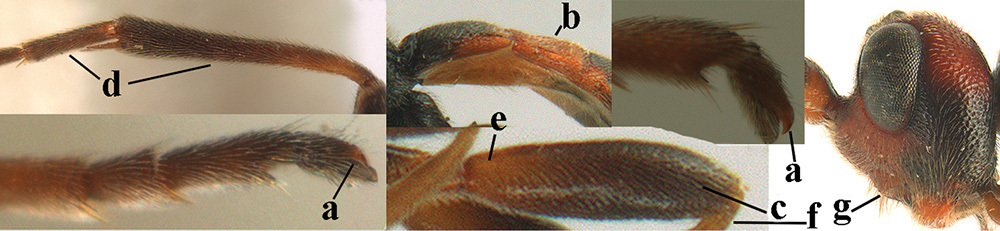
Tarsal claws subpectinate, with four brown medium-sized pectinal bristles (Fig.
Metasoma.
First tergite rather flattened, as long as wide apically; 1st and 2nd tergites coarsely and densely rugose, robust, with distinct median carina; medio-basal area of 2nd tergite wide and short; 2nd suture deep medially and shallow laterally; basal half of 3rd tergite finely rugose, remainder of metasoma largely smooth, punctulate; 4th and apical half of 3rd tergite without sharp lateral crease; ovipositor sheath with medium-sized setae and apically rounded (Fig.
Colour. Brownish yellow; antenna, mesosternum (except anteriorly) and mesopleuron (except anteriorly and dorsally), metapleuron, propodeum, ovipositor sheath and stemmaticum black; hind tibia (except apically) pale yellowish; apices of femora (dorsally) and tibiae, palpi, tarsi (except basally), veins and pterostigma dark brown; wing membrane rather infuscate.
Variation. Size of eyes and ocelli rather variable. Mesopleuron, mesosternum, metapleuron and propodeum brownish yellow or black; 1st tergite entirely brownish yellow or with dark brown patch basally; in desert areas body can be wholly orange. Antennal segments: ♀ 49 (1), 50 (3), 51 (9), 52 (13), 53 (10), 54 (3), 55 (5), 56 (2); ♂ 51 (10), 52 (11), 53 (5), 54 (4), 55 (3), 56 (1). The two sexes have comparable numbers of antennal segments. Apical tergites of ♂ type 3 and fringe moderately strong; inner hind tibial spur 0.50 × as long as hind basitarsus.
Distribution
Albania, Azerbaijan, Bulgaria, Cyprus, Georgia, *Greece, Iran, *Iraq, Israel, *Jordan, Russia, Syria, Turkey, Tunisia, *Turkmenistan, Uzbekistan.


Aleiodes agilis
Rhogas (Rhogas) agilis Telenga, 1941: 165–166, 417.
Rogas agilis;
Rogas (Rogas) agilis; Tobias, 1976: 83, 1986: 76 (transl. 122, 124) (lectotype designation).
Aleiodes agilis;
Aleiodes (Chelonorhogas) agilis;
Rhogas desertus var. armenica Telenga, 1959: 85; Tobias, 1976: 83 (as synonym of A. agilis (Telenga, 1941)), 1986: 76 (transl. 122, 124; id.).
Type material
Lectotype, ♀ (
Molecular data
None.
Biology
Unknown. It appears to fly very early in the year (March).
Diagnosis
Maximum width of hypoclypeal depression approx. 0.8 × minimum width of face; anterior part of clypeus very narrow (Fig.
Description
Paralectotype, ♀, length of fore wing 6.6 mm, of body 7.0 mm.
Head.
Antennal segments of ♀ 47, antenna as long as body and its subapical segments moderately slender; frons rugose, shiny; OOL 1.3 × diameter of posterior ocellus; OOL and vertex remotely punctate, with satin sheen, OOL also with some rugulae; anterior part of clypeus 9 × wider than high, coarsely punctate and rather convex; clypeus above lower level of eyes; ventral margin of clypeus thick and not protruding forwards (Fig.
Mesosoma. Lateral lobes of mesoscutum largely smooth, with satin sheen and whitish setose, middle lobe distinctly punctate and setose; prepectal carina complete and lamelliform; precoxal area of mesopleuron widely rugose, but posterior 0.2 narrowly striate; mesopleuron largely weakly and sparsely punctate, shiny, but anteriorly becoming densely punctate and somewhat rugulose; scutellum largely smooth, with some punctures; propodeum evenly convex, finely rugose and with medio-longitudinal carina, without tubercles.
Wings.
Fore wing: basal half largely glabrous; r 0.6 × 3-SR (Fig.
Legs.
Tarsal claws slender, slightly curved and only setose (Fig.
Metasoma.
First tergite robust, as long as wide apically, distinctly narrowed anteriorly and rather flat posteriorly; 1st and 2nd tergites finely longitudinally striate-rugulose; medio-longitudinal carina of 1st and 2nd tergites indistinct; 2nd tergite 0.7 × longer than its basal width; medio-basal area of 2nd tergite wide triangular, rather short; 2nd suture shallow and narrow; 3rd tergite mainly smooth and with satin sheen; 4th and apical half of 3rd tergite without sharp lateral crease; ovipositor sheath rather slender, with short setae and apically truncate (Fig.
Colour.
Black; pronotal side largely yellowish brown; mesoscutum medio-posteriorly and postero-laterally partly chestnut brown; tegulae, clypeus and antenna (except yellow scapus and pedicellus) yellowish brown; mandible, legs and palpi pale yellowish, but hind coxa and most of middle coxa dark brown; metasoma dark brown but with yellow patches (Fig.
Distribution
Armenia, Iran. Included in this revision, because it may occur in Turkey.


Aleiodes apicalis
Bracon apicalis Brullé, 1832: 381 [examined].
Rhogas apicalis;
Rogas apicalis;
Aleiodes apicalis;
Aleiodes (Chelonorhogas) apicalis; Falco et al. 1997: 60;
Rogas reticulator
Nees, 1834: 211;
Aleiodes reticulator; Papp, 1991a: 70 (as synonym of A. ductor).
Rogas bicolor
Lucas, 1849: 336–337 (not
Rogas rufo-ater
Wollaston, 1858: 24;
Rhogas rufoater;
Rhogas bicolorinus Fahringer, 1932: 318 (replacement name for Rogas bicolor Lucas). Syn. nov.
Rhogas reticulator var. atripes
Costa, 1884: 13;
Rhogas ductor var. atripes;
Aleiodes (Neorhogas) ductor var. atripes;
Rhogas similis
Szépligeti, 1903: 114 (not
Rhogas ductor var. similis;
Rogas ductor
auct. p.p.; Shenefelt, 1975: 1226–1227;
Aleiodes ductor
auct. p.p.;
Type material
Holotype of B. apicalis, ♂ (
Additional material
Albania, Austria, Bosnia & Herzegovina, Bulgaria, Croatia, Cyprus, Czech Republic, France (including Corsica), Germany, Greece (including Chios, Corfu, Crete, Lesbos, Rhodes), Hungary, Italy (including Sardinia, Sicily), Malta, Moldova, Montenegro, Morocco, North Macedonia, Portugal (including Madeira), Romania, Russia (including Dagestan), Serbia, Slovakia, Spain (including Mallorca and Canary Islands: Tenerife, Fuerteventura), Switzerland, Tunisia, Turkey, Ukraine, [Georgia, Kazakhstan, Oman, Iran, Iraq, Israel, Syria, Turkmenistan]. Specimens in
Molecular data
MRS008 (Turkey), MRS111 (Turkey), MRS112 (Turkey), MRS181 (Russia), MRS869 (Sweden).
Biology
Time of flight varies according to harshness of summer. In its more temperate sites plurivoltine April-September(October), overwintering in the mummy, but in Cyprus (and presumably other places with extremely hot dry summers) it appears to be most active from autumn to spring (October–May), with a prolonged summer diapause (June–October or later) in the mummy (reared series ex “Plusia” in
Diagnosis
Maximum width of hypoclypeal depression 0.3–0.4 × minimum width of face (Fig.
Description
Redescribed ♀ (RMNH) from Hungary (Budapest), length of fore wing 5.1 mm, of body 6.7 mm.
Head.
Antennal segments of ♀ more than 40, but apical segments missing (length of antenna of ♀ from Lesbos 1.4 × fore wing and its subapical segments robust); frons with coarse curved rugae, shiny; OOL 1.5 × diameter of posterior ocellus, and distinctly striate; vertex transversely striate, rather weak; clypeus normal, punctulate and convex; ventral margin of clypeus thick and not protruding forwards; width of hypoclypeal depression 0.3 × minimum width of face (Fig.
Mesosoma. Mesoscutal lobes largely smooth, punctulate, shiny; prepectal carina complete, rather strong; precoxal area of mesopleuron largely smooth; mesopleuron above precoxal area weakly and sparsely punctate, especially posteriorly; scutellum largely smooth, with striae laterally; propodeum evenly convex, coarsely vermiculate-rugose, only anteriorly with median carina, without tubercles.
Wings.
Fore wing: r 0.6 × 3-SR (Fig.
Legs.
Tarsal claws with rather slender and medium-sized brownish pecten (Fig.
Metasoma.
First tergite robust, evenly convex; 1st and 2nd tergites rather coarsely obliquely rugose; 1st tergite and basal half of 2nd tergite with median carina; 2nd tergite robust and with striae diverging posteriorly; medio-basal area of 2nd tergite wide triangular, rather short; 2nd suture rather deep medially; 3rd tergite largely smooth, except anteriorly with some striae; 4th and apical half of 3rd tergite without sharp lateral crease; ovipositor sheath with rather long setae and apically rounded (Fig.
Colour. Black; scapus, pedicellus, tegulae (but humeral plate brownish yellow), base of hind tibia narrowly, apical half of hind tibia, telotarsi, hind tarsus largely, ventral apical half of metasoma, pterostigma and veins (except C+SC+R of fore wing) dark brown; remainder of basal half of antenna and palpi yellowish brown; remainder of legs (but apical two-fifths of hind femur black), 1st and 2nd tergites, 3rd tergite basally and laterally orange brown; remainder of hind tibia pale yellowish; apex of middle femur and wing membrane somewhat infuscate.
Variation. A. apicalis is very variable in colour and the colour patterns are not restricted to certain areas, but in general southern Palaearctic specimens are darker than northern ones (or specimens from high altitudes). The tegula is dark brown or black, and the humeral plate usually paler than the tegula or equally black, but both usually yellowish in southern specimens; the hind tarsus is dark brown or black, but sometimes 3rd and 4th segments yellowish; the hind tibia variably reddish to black, but palest at extreme base; the pronotum is very occasionally reddish. The extent of black colouration of the legs is especially variable, and sometimes all legs are entirely black (var. rufoater (Wollaston, 1858)). Antenna, especially in females, can be more or less light reddish brown, especially basally, or dark brown/black throughout. Antennal segments: ♀ 44(1), 46(3), 47(11), 48(20), 49(31), 50(41), 51(19), 52(10), 54(3), 55(1), 57(1); ♂ 46(3), 47(7), 48(17), 49(29), 50(30), 51(32), 52(11), 53(5), 54(1). Males have on average approx. one antennal segment more than females. Apical tergites of ♂ type 4, setosity dense (making the tergites appear concave; Figs
Distribution
*Albania, Austria, *Bosnia & Herzegovina, *Bulgaria, *Croatia, Cyprus, *Czech Republic, *France (including Corsica), *Georgia, *Germany, Greece (including Chios, Corfu, Crete, Lesbos, Rhodes), *Hungary, Iran, *Iraq, *Israel, *Italy (including Sardinia, Sicily), *Kazakhstan, *Malta, *Moldova, *Montenegro, *Morocco, *North Macedonia, *Oman, *Portugal (including Madeira), *Romania, *Russia (including Dagestan), *Serbia, *Slovakia, Spain (including Mallorca and Canary Islands: Tenerife, Fuerteventura), *Syria, Switzerland, *Tunisia, Turkey, *Turkmenistan.
New synonymy
The synonymy of Rogas rufo-ater Wollaston, 1858, and Rhogas similis Szépligeti, 1903, are based on examination of the types listed above. The lectotype of Rogas bicolor Lucas, 1849 (not Spinola, 1808) and of Rhogas bicolorinus Fahringer, 1932, has been examined by Dr Jenö Papp and we agree with his opinion that it is a synonym of A. ductor auct. (= A. apicalis). The types of Rogas reticulator Nees, 1834, and Rhogas reticulator var. atripes Costa, 1884, are lost or unavailable and their synonymy is based on the original description and the interpretation by later authors.


Aleiodes apicalis (Brullé), ♀, Bulgaria, Rodopi 54 wings 55 mesosoma lateral 56 mesosoma dorsal 57 1st –3rd metasomal tergites dorsal 58 fore femur lateral 59 hind femur lateral 60 head anterior 61 head dorsal 62 head lateral 63 outer hind tarsal claw 64 base of antenna 65 apex of antenna.

Aleiodes arnoldii
Rogas (Rogas) arnoldii Tobias, 1976: 84, 222, 1986: 78 (transl.: 128).
Aleiodes (Neorhogas) arnoldi
[sic!];
Aleiodes (Neorhogas) arnoldii;
Type material
Holotype, ♀ (
Additional material
1 ♂ (RMNH), “Turkey, Hakkâri, Tanin Tanin Pass, 25.vi.1985, 2200 m, C.J. Zwakhals”. Male is provisionally associated with this species; it may belong to a related species.
Molecular data
None.
Biology
Unknown. The holotype was collected in May.
Diagnosis
Maximum width of hypoclypeal depression 0.4–0.5 × minimum width of face (Fig.
Description
Holotype, ♀, length of fore wing 4.4 mm, of body 5.7 mm.
Head.
Antennal segments of ♀ 37, length of antenna 0.85 × fore wing, its subapical segments quadrate; frons with rather coarse curved rugae, shiny, and rugose behind antennal sockets; OOL 2.0 × diameter of posterior ocellus, and finely remotely punctate, interspaces much larger than diameter of punctures; vertex spaced punctate, shiny; face transversely rugose; clypeus finely rugulose and with long setae; ventral margin of clypeus thick and not protruding forwards; width of hypoclypeal depression 0.45 × minimum width of face; length of eye 1.1 × temple in dorsal view (Fig.
Mesosoma. Mesoscutal lobes largely smooth, shiny, sparsely and finely punctate; precoxal area of mesopleuron coarsely rugose, but absent posteriorly; metapleuron remotely punctate, interspaces much wider than diameter of punctures, shiny; mesopleuron above precoxal area (except speculum) punctate and dorsally rugose; scutellum sparsely punctate or punctulate, medio-posteriorly rugulose and with some striae laterally, no carina; propodeum evenly convex and coarsely vermiculate-rugose, medio-longitudinal carina strong in basal 0.6, and without tubercles.
Wings.
Fore wing: just reaching apex of metasoma; r 0.35 × 3-SR (Fig.
Legs. Tarsal claws subpectinate, with six yellowish medium-sized pectinal bristles; hind coxa obliquely striated dorsally, punctulate laterally; hind trochantellus robust; length of hind femur and basitarsus 3.6 and 4.6 × their width, respectively; length of inner hind spur 0.5 × hind basitarsus.
Metasoma.
First tergite rather flattened, as long as wide apically; 1st and 2nd tergites coarsely longitudinally and densely rugose, robust and posterior corners of 1st protruding outside base of 2nd tergite, with distinct median carina; medio-basal area of 2nd tergite wide and short; 2nd suture moderately deep and crenulate; basal half of 3rd tergite longitudinally striate, remainder of metasoma largely smooth, punctulate; 4th and apical half of 3rd tergite without sharp lateral crease; ovipositor sheath wide, setose and apically truncate (Fig.
Colour. Yellowish brown; mesosoma (except mesoscutum, scutellum medially, pronotum anteriorly and dorsally), ovipositor sheath, 3rd tergite (except antero-lateral corners) and following segments black; apical half of antenna, pedicellus, palpi, hind femur apico-dorsally, telotarsi, veins, parastigma basally and pterostigma dark brown; wing membrane rather brownish infuscate.
Variation.
Antennal segments of ♀ 37(1). Male is largely black, except for 2nd tergite and anterior half of 3rd tergite (Fig.
Distribution
Azerbaijan, *Turkey.
Notes
Easily confused with A. ruficornis (Herrich-Schäffer); the relative size of the clypeus (wider and somewhat shorter in A. arnoldii than in A. ruficornis) seems to be the main difference in both sexes. In addition, the female of A. arnoldii has the temple ventrally and the malar space yellowish brown (dark brown in A. ruficornis). The male has darker legs and 1st metasomal tergite than the female (the sexes more similar in A. ruficornis). Also reported from Uzbekistan (Yuldashev, 2006); the record from Poland (Huflejt, 1997) most likely concerns A. ruficornis (Herrich-Schäffer). Aleiodes arnoldii sensu




Aleiodes aterrimus
Bracon aterrimus
Ratzeburg, 1852: 35;
Aleiodes aterrimus;
Aleiodes grandis
Giraud, 1857: 178;
Aleiodes (Neorhogas) grandis;
Aleiodes (Chelonorhogas) aterrimus; Falco et al. 1997: 60.
Rogas grandis;
Rogas (Rogas) grandis;
Rhogas malaisei Shestakov, 1940: 7.
Rogas malaisei;
Aleiodes malaisei;
Rogas (Rogas) vicinus Papp, 1977a: 114, 115 [examined]. Syn. nov.
Aleiodes (Neorhogas) vicinis;
Type material
Lectotype of A. grandis, ♂ (
Additional material
Austria, Belgium, British Isles (England V.C.s 8, 9, 10, 11, 12, 14, 15, 20, 22, 28, 29, 39), Czech Republic, Finland, Germany, Hungary, Netherlands (GE: Brummen (Leuvenheim); LI: Epen; ZH: Schoonrewoerd), Poland, Romania, Russia, Slovakia, Spain, Switzerland. Specimens in
Molecular data
MRS024 (UK), MRS147 (UK).
Biology
Univoltine, spending ca ten months of the year in the exposed mummy on an aerial twig. Collected from April–June, among broadleaved trees (but see paragraph below). Reared from arboreal Amphipyra spp.: A. pyramidea (Linnaeus) (29; M.G. Bloxham, C. Bystrowski, J. Connell, A.P. Fowles, G.M. Haggett, B.T. Parsons, D.L.J. Quicke, M.R. Shaw); A. berbera (Rungs) (5:1 [5
Before becoming mummified the host moves to a narrow twig, to which the mummy will be very strongly glued. In the early stage of the mummification process (Fig.
Although the above is a consistent pattern for this species, it does not account for a small number of specimens (14 ♀, 4♂ in BZL, MRC,
Diagnosis
Maximum width of hypoclypeal depression 0.3–0.4 × minimum width of face (Fig.
Dr K. Samartsev (in litt.) kindly brought to the first author’s attention that the East Palaearctic A. sapporensis (Watanabe, 1937) occurs in southern European Russia (Middle and Lower Volga territories). Aleiodes aterrimus and A. sapporensis differ only slightly, mainly by the colour of the extreme base of the hind tibia (completely dark brown in A. sapporensis and usually narrowly pale yellowish in A. aterrimus) and by the shape of temple in dorsal view (roundly narrowed in A. sapporensis and rather linearly narrowed in A. aterrimus). There is also a slight difference in the proportions of the face (A. sapporensis has facial width 1.50–1.60 × medial height including clypeus and A. aterrimus 1.65–1.75 ×). A. sapporensis seems to have the lateral carinae of propodeum more protruding and has 58–66 antennal segments.
Description
Redescribed ♀ (RMNH) from England (Pamber Forest). Length of fore wing 7.3 mm, of body 8.6 mm.
Head.
Antennal segments of ♀ 59, length of antenna 1.1 × fore wing, its subapical segments rather robust; frons largely superficially granulate; OOL 1.8 × diameter of posterior ocellus, and superficially rugulose-granulate and shiny; vertex superficially rugulose-granulate, rather shiny; clypeus with some punctures; ventral margin of clypeus thick and not protruding forwards (Fig.
Mesosoma. Mesoscutal lobes densely and finely punctate-coriaceous, rather matt; precoxal area of mesopleuron largely smooth medially, densely punctate anteriorly and posteriorly; metapleuron densely punctate; metanotum with nearly complete median carina; scutellum punctate-coriaceous; propodeum rather convex and coarsely reticulate-rugose, medio-longitudinal carina nearly complete, and with slightly protruding carinae laterally.
Wings.
Fore wing: r 0.4 × 3-SR (Fig.
Legs.
Tarsal claws with conspicuous and robust blackish pecten (Fig.
Metasoma.
First tergite evenly convex, as long as wide apically; 1st and 2nd tergites with medio-longitudinal carina and coarsely longitudinally rugose, but posterior quarter of 2nd tergite irregularly rugose and no median carina; medio-basal area of 2nd tergite triangular and rather distinct (Fig.
Colour. Black; antenna (except scapus and pedicellus), palpi, tegulae, fore and middle telotarsi, veins and pterostigma dark brown; coxae, trochanters and trochantelli, apical third of hind femur (ventrally extended to its apical two-thirds), hind tibia (except pale yellowish basal ring) and hind tarsus black, remainder of legs yellowish brown; wing membrane subhyaline.
Variation.
Hind femur usually only apically dark brown, but sometimes entirely dark brown; coxae black or sometimes largely yellowish brown. Two females (both
Distribution
Austria, *Belgium, British Isles (England), Czech Republic, *Finland, Germany, Hungary, *Netherlands, Poland, *Romania, Russia, Serbia, Slovakia, Spain, *Switzerland.
New synonymy
The synonymy of Rogas vicinus Papp, 1977, with Aleiodes aterrimus (Ratzeburg, 1852) is based on the examination of the types listed above. The differences between R. vicinus and R. grandis (= A. aterrimus) listed in the original description (head less constricted posteriorly, apical antennal segments more robust, 1st metasomal tergite less robust and 2nd tergite somewhat longer) fall within the normal variation of A. aterrimus.




Aleiodes aterrimus (Ratzeburg), ♀, England, Pamber Forest, but 102 from Austria, Wien 102 fore wing 103 hind wing 104 mesosoma lateral 105 mesosoma dorsal 106 metasoma dorsal 107 fore femur lateral 108 hind femur lateral 109 head anterior 110 head dorsal 111 head lateral 112 hind tibia and tarsus lateral 113 outer hind tarsal claw 114 base of antenna 115 apex of antenna.
Aleiodes carbonarius
Aleiodes carbonarius Giraud, 1857: 177–178 [examined].
Aleiodes (Neorhogas) carbonarius;
Aleiodes carbonarius;
Rogas carbonarius;
Rhogas (Rhogas) carbonarius ab. giraudi
Fahringer, 1931: 236;
Type material
Lectotype of A. carbonarius, ♂ (
Additional material
3 ♀ (
Molecular data
MRS162 (Hungary), MRS163 (Hungary), MRS 164 (Hungary).
Biology
Adults of this lowland species have been collected from the very end of April to July (see also Papp, 1999), and it is found in grassland habitats. Reared from the noctuid Tholera decimalis (Poda) (3:1; M.R. Shaw/Hungary). The decidedly large mummy is very similar to that of A. grassator and forms underground (Fig.
Diagnosis
Maximum width of hypoclypeal depression 0.4–0.5 × minimum width of face (Fig.
Description
Redescribed ♀ (
Head.
Antennal segments of ♀ 46, 4th segment 0.9 × longer than wide (Fig.
Mesosoma. Mesoscutal lobes densely punctate, interspaces superficially granulate and with satin sheen; precoxal area of mesopleuron coarsely rugose medially and punctate posteriorly; remainder of mesopleuron mainly coarsely punctate; scutellum flat, sparsely finely punctate and with lateral carina; propodeum coarsely rugose, medio-longitudinal carina indistinct, rounded posteriorly and dorsal part rather short.
Wings.
Fore wing: r 0.4 × 3-SR; marginal cell ends near level of apex of 3-M (Fig.
Legs.
Tarsal claws robust and with only brownish bristly setae (Fig.
Metasoma.
First tergite rather flattened, 0.9 × as long as wide apically; 1st and 2nd tergites with medio-longitudinal carina and coarsely longitudinally rugose, but posterior quarter of 2nd tergite irregularly rugose and no median carina; medio-basal area of 2nd tergite triangular and short (Fig.
Colour. Dark orange brown; apical two-thirds of antenna, patch on hind femur dorso-apically, and telotarsi, dark brown; temple ventrally, malar space, mesosternum, mesopleuron, metapleuron, propodeum, pair of patches on 2nd tergite and most of apical 0.4 of tergite, and 3rd–7th tergites black; palpi (especially labial palp), veins and pterostigma dark brown, basal third of antenna (but scapus dorsally blackish) rather pale yellowish brown; tegulae and remainder of legs; yellowish brown; wings strongly infuscate.
Variation.
Antennal segments: ♀ 46(2), 49(1); ♂ 47(1), 50(1), 52(1), 54(1), 56(1), 57(1); length of fore wing of ♀ ca two-thirds of body length (0.8 × in ♂); males always darker than females; mainly black with legs mainly dark brown or blackish, but male from Austria has basal half of metasoma orange brown and legs partly yellowish brown. Males have 2nd submarginal cell distinctly shorter than in females (as in A. grassator), antenna 0.9 × length of body and slightly less robust subapically, temple and face long setose and malar space 0.5–0.7 × length of eye in lateral view; metasoma black or 1–2 basal tergites reddish and apical tergites type 1, fringe not observed (Fig.
Distribution
Austria, Czech Republic, Hungary, *Russia (Lake Baikal).
Notes
Very similar to A. grassator (Thunberg), and especially A. carbonaroides; males of A. carbonarius and carbonaroides are normally black but males with partly orange brown metasoma occur. The three species exhibit sexual dimorphism of the 2nd submarginal cell (less robust (and also longer in A. carbonarius) in female than in male).


Aleiodes carbonarius Giraud, ♀, Hungary, Veszprém 119 fore wing 120 hind wing 121 mesosoma lateral 122 mesosoma dorsal 123 metasoma dorsal 124 fore femur lateral 125 hind femur lateral 126 head anterior 127 head dorsal 128 head lateral 129 base of antenna 130 apex of antenna 131 inner hind tarsal claw.

Aleiodes carbonaroides , sp. nov.
Type material
Holotype, ♀ (
Molecular data
None.
Biology
Adults of this lowland species have been collected in April and May. The two paratypes from Suffolk were swept from Breck grassland (Morley, 1937, misidentified as A. carbonarius). Reared from the grass-feeding noctuid Cerapteryx graminis (Linnaeus) (6 [2 are RMNH]; K.P. Carl/Netherlands). If it is a specialist, it is presumably univoltine and overwinters in the mummy (the univoltine known host overwinters in the egg stage). Mummy similar to that of the closely related A. carbonarius and A. grassator, but slightly smaller.
Diagnosis
Maximum width of hypoclypeal depression 0.4–0.5 × minimum width of face (Fig.
Description
Holotype, ♀, length of fore wing 4.2 mm, of body 7.1 mm.
Head.
Antennal segments of ♀ 45, 4th segment 0.9 × longer than wide (Fig.
Mesosoma. Mesoscutal lobes moderately punctate, interspaces superficially granulate-coriaceous and with satin sheen; precoxal area of mesopleuron coarsely rugose medially, but largely smooth posteriorly; remainder of mesopleuron mainly punctate; scutellum flat, sparsely finely punctate and with irregular lateral carina; propodeum coarsely rugose, medio-longitudinal carina complete, rounded posteriorly and dorsal part approx. as long as posterior part.
Wings.
Fore wing: r 0.4 × 3-SR (Fig.
Legs.
Tarsal claws robust and with only brownish bristly setae (Fig.
Metasoma.
First tergite rather flattened, 0.7 × as long as wide apically; 1st and 2nd tergites with medio-longitudinal carina and coarsely longitudinally rugose, but posterior quarter of 2nd tergite without medio-longitudinal carina; medio-basal area of 2nd tergite triangular and short; 2nd suture deep and crenulate; basal half of 3rd tergite finely longitudinally rugose, remainder of metasoma superficially micro-sculptured; 4th and apical third of 3rd tergite without sharp lateral crease; ovipositor sheath wide, with long setae and apically truncate (Fig.
Colour. Dark orange brown; apical half of antenna, patch on hind femur dorso-apically, and telotarsi apically, dark brown; mesosternum, mesopleuron (except dorsally and postero-ventrally), metapleuron (except medio-dorsally), propodeum (except pair of posterior patches), 3rd–7th tergites (except antero-lateral corners of 3rd tergite) black; palpi, basal half of antenna, tegulae and remainder of legs rather pale yellowish brown; veins and pterostigma dark brown; wings strongly infuscate but hind wing less than fore wing.
Variation.
Basal third or half of antenna of ♀ pale yellowish brown; vein 3-SR 1.4–1.6 × as long as vein 2-SR; hind femur of ♀ 3.2–3.5 × longer than wide; 1st metasomal tergite 0.7–0.8 × its apical width; temple and occiput ventrally, and malar space ventrally orange brown or black. Antennal segments: ♀ 43(1), 45(1); ♂ 48(1), 49(2), 51(1), 50(1), 53(2); males clearly have many more antennal segments than females. Males are much darker than females; body black with palpi and legs mainly dark brown or blackish (Fig.
Distribution
Germany, Netherlands, U.K.
Etymology
The suffix “-oides” indicates similar to; in this case the high similarity to A. carbonarius Giraud.



Aleiodes caucasicus
Rogas (Rogas) caucasicus Tobias, 1976: 86, 222, 1986: 81 (transl.: 133) [examined].
Aleiodes (Neorhogas) caucasicus;
Aleiodes caucasicus;
Type material
Holotype, ♀ (
Additional material
Figured ♀ (
Molecular data
None.
Biology
Unknown. Specimens collected in April-May and flight time probably April–May. We have not seen reared material. Probably, like A. fortipes, it will be found to be univoltine, overwintering in the mummy, but direct evidence is lacking.
Diagnosis
Maximum width of hypoclypeal depression approx. 0.3 × minimum width of face (Fig.
Description
Holotype, ♀, length of fore wing 3.7 mm, of body 4.6 mm.
Head.
Antennal segments of ♀ 41, length of antenna 1.3 × fore wing, its subapical segments rather robust; frons largely finely rugulose medially; OOL 2.2 × diameter of posterior ocellus, and coarsely transversely striate; vertex transversely striate and rather shiny; clypeus rugulose, but ventrally depressed and smooth; ventral margin of clypeus thick and not protruding forwards (Fig.
Mesosoma.
Mesoscutal lobes largely rugulose-granulate, rather matt; precoxal area of mesopleuron transversely striate medially, distinctly rugose antero-dorsally and remainder largely punctulate; pleural sulcus moderately crenulate (Fig.
Wings.
Fore wing: r 0.6 × 3-SR; 1-CU1 horizontal, 0.5 × 2-CU1; r-m unsclerotized and 0.7 × 3-SR; 2nd submarginal cell medium-sized (Fig.
Legs.
Tarsal claws robust and with brownish bristles (Fig.
Metasoma.
First tergite evenly convex, 0.9 × longer than wide apically; 1st and 2nd tergites with indistinct medio-longitudinal carina and coarsely longitudinally rugose, but posterior quarter of 2nd tergite irregularly rugose and no median carina; medio-basal area of 2nd tergite triangular and rather distinct (Fig.
Colour. Orange brown; head, 3rd tergite (except antero-laterally) and subsequent tergites black; scapus, pedicellus basally, 11th and following antennal segments, palpi, veins, parastigma, pterostigma and femora apico-dorsally, tibia and tarsal segments apically, ventral half of metasoma and ovipositor sheath dark brown; tegulae, 3rd–10th antennal segments brownish yellow; wing membrane subhyaline.
Variation. Head black or mainly dark brown, specimen from Bulgaria also anterior half of mesosoma; antenna of ♀ with 38 or 41 segments according to the original description; 11th and 12th antennal segments of ♀ dark brown or brownish yellow; hind femur 3.6–3.8 × as long as wide. The male is unknown, or possibly has not been distinguished from that of A. fortipes.
Distribution
*Bulgaria, Russia (SW).
Notes
It remains unclear whether this predominantly rather yellowish orange species is distinct from A. fortipes, which in its more western localities is a much darker insect. Females intermediate in colour (and included in A. fortipes) seem to predominate in eastern Europe. More material (preferably with biological data) is needed to clarify the status of A. caucasicus.


Aleiodes caucasicus (Tobias), ♀, Russia, Sotchi 164 fore wing 165 hind wing 166 mesosoma lateral 167 mesosoma dorsal 168 metasoma dorsal 169 fore femur lateral 170 hind femur lateral 171 head anterior 172 head dorsal 173 head lateral 174 base of antenna 175 antenna 176 hind tibia and tarsus lateral 177 outer hind tarsal claw.
Aleiodes coriaceus , sp. nov.
Type material
Holotype, ♀ (
Molecular data
MRS311 (Sweden), MRS377 (Sweden).
Biology
Unknown. The available specimens were collected in July, and it is almost certainly univoltine, but we have not seen reared material.
Diagnosis
Maximum width of hypoclypeal depression approx. 0.4 × minimum width of face (Fig.
Description
Holotype, ♀, length of fore wing 6.1 mm, of body 6.7 mm.
Head.
Antennal segments of ♀ 54, antenna 1.1 × as long as fore wing, its basal segments robust, subapical segments medium-sized and apical segment with spine; frons largely smooth, except for some micro-sculpture; OOL 0.9 × diameter of posterior ocellus, rugulose-coriaceous and rather dull, groove beside posterior ocellus deep and smooth; vertex coriaceous with some rugulae, rather dull; face transversely rugose; clypeus densely rugulose; ventral margin of clypeus thin and not protruding forwards (Fig.
Mesosoma. Mesoscutal lobes largely coriaceous and matt; precoxal area of mesopleuron partly remotely punctulate and superficially micro-sculptured; medio-longitudinal carina of metanotum distinct posteriorly; scutellum punctate and with lateral carina; propodeum convex and rugose, medio-longitudinal carina absent posteriorly, and without protruding carinae laterally.
Wings.
Fore wing: r 0.35 × 3-SR (Fig.
Legs.
Tarsal claws with rather conspicuous and medium-sized dark brown pecten (Fig.
Metasoma.
First tergite convex and basally rather steep, as long as wide apically; 1st and 2nd tergites with medio-longitudinal carina and longitudinally rugose; maximum width of 2nd tergite 1.5 × its median length; medio-basal area of 2nd tergite medium-sized triangular and rather short (Fig.
Colour. Black; mesoscutum posteriorly, legs (but fore and middle telotarsi, fore and middle femora basally and apically, fore and middle trochanters and trochantelli, hind tarsus dark brown or infuscate, posterior half of hind femur dorsally and hind tibia largely blackish), propodeum and 1st –3rd metasomal tergites (but posterior half of 3rd tergite blackish posteriorly) reddish brown; tegulae brownish yellow, but humeral plate largely dark brown; palpi, pterostigma and veins dark brown; wing membrane slightly infuscate.
Variation.
Antennal segments: ♀ 52(1), 54(1); ♂ 53(1). Length of fore wing 5.3–6.1 mm. Male is very similar to female (Figs
Distribution
Sweden.
Etymology
Coriaceus is Latin for leathery, because of the coriaceous sculpture of vertex and mesoscutum.


Aleiodes coriaceus sp. nov., ♀, holotype 180 wings 181 mesosoma lateral 182 mesosoma dorsal 183 propodeum and 1st–3rd metasomal tergites dorsal 184 fore femur lateral 185 hind femur lateral 186 head anterior 187 head dorsal 188 head lateral 189 outer hind tarsal claw 190 base of antenna 191 apex of antenna.

Aleiodes cruentus
Rogas cruentus
Nees, 1834: 212;
Rogas (Rogas) cruentus;
Aleiodes (Neorhogas) cruentus;
Aleiodes (Chelonorhogas) cruentus;
Aleiodes cruentus;
Rhogas cruentus ab. nigricans
Fahringer, 1932: 238;
Rhogas cruentus ab. basalis Hellén, 1927: 22 (invalid name).
Rhogas cruentus ab. nigromaculata Hellén, 1927: 22 (invalid name).
Rhogas cruentus ab. rufofasciata Hellén, 1927: 22 (invalid name).
Rogas dorsalis
Herrich-Schäffer, 1838: 154;
Rogas affinis
Herrich-Schäffer, 1838: 124 (key only);
Aleiodes affinis;
Type material
Neotype of A. affinis here designated, ♀ (RMNH), “Museum Leiden, Nederland, Melissant (ZH), [at light], 10.viii.1980, K.J. Huisman”. It is important for nomenclatorial stability to fix our interpretation of A. affinis because the types of Braconidae described by Herrich-Schäffer are lost (Horn and Kahle 1935–37; the first author could not find any specimen in
Additional material
Austria, Bulgaria, Croatia, Czech Republic, Finland, France, Germany, Greece, Italy (including Sicily), Moldova, Netherlands (FR: Ried, GE: Beusichem; Heerde; Voorst (Twello), LI: Thorn, NB: Eindhoven; Tilburg (Kaaistoep), OV: Buurse; Hasselt, ZH: Lexmond; Melissant; Middelharnis; Oostvoorne, ZL: Oostkapelle), Norway, Romania, Slovakia, Slovenia, Spain, Sweden, Ukraine, [Mongolia]. Specimens in
Biology
Probably univoltine, certainly overwintering as a mummy. Collected June-August, often at light and including around Dianthus barbatus harbouring larvae of the noctuid Hadena confusa (Hufnagel) (H. Schnee/Germany). In Austria it has been collected up to 2000 m. Only one reared specimen seen, from H. confusa [
Molecular data
MRS558 (France), MRS624 (Germany), MRS625 (Germany).
Diagnosis
Maximum width of hypoclypeal depression (0.5–)0.6–0.7 × minimum width of face (Fig.
Description
Neotype of A. affinis, ♀, length of fore wing 7.3 mm, of body 10.2 mm.
Head.
Antennal segments of ♀ 61, length of antenna 1.2 × fore wing, its subapical segments rather robust; frons largely smooth and shiny, but rugulose near stemmaticum; OOL 0.6 × diameter of posterior ocellus, and coarsely punctate, interspaces approx. equal to diameter of punctures; vertex mainly densely punctate, shiny; clypeus coarsely punctate-rugose; ventral margin of clypeus thick and not protruding forwards (Fig.
Mesosoma. Mesoscutal lobes densely and finely punctate, with satin sheen; precoxal area of mesopleuron with some rugae medially, rather densely punctate anteriorly and posteriorly; metapleuron mainly sparsely punctate, shiny; scutellum rather weakly punctate and slightly convex; propodeum evenly convex and coarsely rugose, medio-longitudinal carina complete and straight.
Wings.
Fore wing: r 0.4 × 3-SR (Fig.
Legs.
Tarsal claws with conspicuous and robust dark brown pecten (Fig.
Metasoma.
First tergite rather flattened, as long as wide apically; 1st and 2nd tergites with medio-longitudinal carina and largely coarsely longitudinally rugose, but posterior quarter of 2nd tergite irregularly rugose and no median carina; medio-basal area of 2nd tergite triangular and rather distinct (Fig.
Colour. Black; posterior half of mesoscutum, scutellum largely, apical rim of 1st tergite and basal rim of 2nd tergite reddish brown; fore coxa, bases of middle and hind coxae blackish; apex of hind tibia, telotarsi, hind tarsus, palpi, veins and pterostigma dark brown; tegulae and remainder of hind tibia pale yellowish; remainder of legs reddish brown; wing membrane subhyaline.
Variation.
Vein 1-CU1 of fore wing 0.8–1.1 × as long as 2-CU1; mesoscutum, scutellum, metanotum, 1st and 2nd metasomal tergites are most often entirely reddish or orange brown but variably partly blackish, in particular 1st tergite sometimes with dark medial patch; pronotum and mesopleuron black or reddish dorsally; parastigma narrowly dark brown or yellowish brown; coxae entirely reddish to entirely dark brown. Antennal segments: ♀ 53(1), 55(1), 56(3), 57(5), 58(9), 59(9), 60(10), 61(9), 62(3), 63(1), 65(2), 67(1). ♂ 60(6), 61(7), 62(2), 63(5), 64(3), 65(1), 66(1), 67(5), 69(1). The males have on average approx. three more antennal segments than females. Males are very similar but often darker than females, 2nd tergite 0.9–1.0 × as long as basal width of tergite and apical tergites type 1 and (usually) type 2, with fringe present in the latter (Fig.
Distribution
*Austria, Bulgaria, Croatia, Czech Republic, Finland, France, Germany, *Greece, Italy, *Moldova, Mongolia, *Netherlands, Norway, *Romania, Slovakia, *Slovenia, Spain, Sweden, Ukraine.
Notes
An examined female (


Aleiodes cruentus (Nees), ♀, Germany, Markkleeberg 199 wings 200 mesosoma lateral 201 mesosoma dorsal 202 metasoma dorsal 203 fore femur lateral 204 hind femur lateral 205 outer hind tarsal claw 206 head anterior 207 head dorsal 208 head lateral 209 base of antenna 210 apex of antenna 211 hind tarsus lateral.

Aleiodes desertus
Rhogas (Rhogas) desertus Telenga, 1941: 184–185, 423 (not R. aestuosus var. desertus Telenga, 1941, from China) [examined].
Rogas desertus;
Rogas (Rogas) desertus;
Aleiodes desertus;
Type material
Lectotype, ♀ (
Molecular data
None.
Biology
Unknown. It seems to fly in spring (March–May) and may be univoltine.
Diagnosis
Maximum width of hypoclypeal depression 0.9–1.0 × minimum width of face; anterior part of clypeus very narrow, most of clypeus depressed (Fig.
Description
Lectotype, ♀, length of fore wing 7.5 mm, of body 8.2 mm.
Head.
Antennal segments of ♀ more than 45, but apical segments missing, length of antenna of paralectotype 1.1 × body and its subapical segments moderately slender; frons rugose, shiny; OOL 0.9 × diameter of posterior ocellus; OOL and vertex remotely punctate, shiny; anterior part of clypeus 9 × wider than high, coarsely punctate and rather convex; clypeus above lower level of eyes; ventral margin of clypeus thick and not protruding forwards; width of hypoclypeal depression 0.9 × minimum width of face (Fig.
Mesosoma. Lateral lobes of mesoscutum largely smooth, strongly shiny and glabrous, middle lobe remotely punctulate and with satin sheen; prepectal carina complete and lamelliform; precoxal area of mesopleuron widely rugose, but posterior 0.2 narrowly striate; mesopleuron above precoxal area anteriorly rugose and remainder weakly and sparsely punctate, shiny; axilla crenulate but posteriorly densely and coarsely rugose; scutellum largely smooth, with some punctures; propodeum evenly convex, finely rugose and with strong medio-longitudinal carina, without tubercles.
Wings.
Fore wing: basal half largely glabrous; r 0.7 × 3-SR (Fig.
Legs.
Tarsal claws slender, slightly curved and only setose (Fig.
Metasoma.
First tergite robust, 0.9 × longer than wide apically, strongly narrowed anteriorly (Fig.
Colour. Black; mesoscutum posteriorly partly chestnut brown; antenna, clypeus, malar space ventrally, mandible, pronotum and propleuron anteriorly and metasoma, brownish yellow; tegulae, legs and palpi pale yellowish; pterostigma and ovipositor sheath dark brown; veins of fore wing (but pale in basal 0.3 of fore wing) brown; wing membrane hyaline.
Variation. Length of body 7.0–8.2 mm, of fore wing 7.5–7.9 mm; temple punctate to smooth; precoxal sulcus area finely to rather coarsely rugose; pronotal side largely black (except ventrally) black or brownish yellow; lateral lobes of mesoscutum entirely dark chestnut brown or only posteriorly so, or mesoscutum largely yellowish brown posteriorly and prolonged to base of notauli; first tergite usually entirely brownish yellow, but sometimes dark brown and only posteriorly and laterally yellowish; pterostigma dark brown or brown. Antennal segments: ♀ 63(1).
Distribution
Turkmenistan, Uzbekistan.
Notes
We have included this extralimital species from Central Asia because we suspect it may occur in Turkey. It should not be confused with Rogas aestuosus var. desertus Telenga, 1941, described from China in the same paper. The latter is an unavailable name (a primary homonym) and most likely a colour variety of R. aestuosus.


Aleiodes desertus (Telenga), ♀, paralectotype, but 224 and 230 of lectotype 221 fore wing 222 hind wing 223 mesosoma lateral 224 mesosoma dorsal 225 metasoma dorsal 226 fore femur lateral 227 hind femur lateral 228 apex of antenna 229 head anterior 230 head dorsal 231 head lateral 232 outer hind tarsal claw 233 base of antenna.
Aleiodes dissector
Rogas dissector
Nees, 1834: 208;
Rogas (Rogas) dissector;
Aleiodes (Neorhogas) dissector; Papp, 1985a: 145, 1987b: 35, 1991a: 74, 1991d: 5, 1999: 550; Belokobylskij, 1996: 9;
Aleiodes (Chelonorhogas) dissector;
Aleiodes dissector;
Phylax aestivalis
Snellen van Vollenhoven, 1858: 282; Shenefelt, 1975: 1226 (as synonym of A. dissector);
Type material
Holotype of A. aestivalis, ♀ (RMNH), “[Netherlands], Haag [= near The Hague], 6 [= June], v.Voll.”. According to the original description the ♂ holotype of R. dissector from Germany should be in the Gravenhorst collection (Museum of Natural History, University of Wrocław, Wrocław), but so far it has not been found.
Additional material
Austria, British Isles (England: V.C.s 15, 17, 20, 22, 23, 24, 30, 31, 34, 37, 58; Scotland: V.C.s 73, 88, 89, 95, 96, 97, 107), Croatia, Czech Republic, Finland, France, Germany, Greece, Hungary, Montenegro, Italy, Netherlands (FL: Lelystad, GE: Barneveld, OV: Raalte (Heino), ZH: Wassenaar), Norway, Russia, Serbia, Slovakia, Switzerland, Ukraine, [Armenia]. Specimens in
Molecular data
MRS007 (Turkey), MRS025 (Turkey), MRS145 (UK), MRS146 (UK).
Biology
Univoltine, collected in May and June in deciduous scrub and woodland. In Britain it is widespread but particularly common in birch-dominated woodland in upland Scotland. Reared from the noctuids Orthosia incerta (Hufnagel) (17, M.R. Shaw), O. gothica (Linnaeus) (1, J.L. Yela) and Orthosia sp. (3), overwintering in the concealed mummy. An additional specimen, lacking a mummy but labelled as reared doubtfully from the sesiid Paranthrene tabaniformis (Rottemburg) (RMNH), which normally feeds under Populus bark at ground level or below, can be discounted as a probable substrate rearing in which the mummy of the true host was overlooked. Parasitised host larvae in their penultimate instar leave their feeding sites and enter the soil or other site of moderate concealment (including below loose bark), where they prepare a chamber as though to pupate. At this time the parasitoid larva within the strongly retarded host (Fig.
The moderately large hypoclypeal opening and protruding sharp-rimmed clypeus of A. dissector is seen in some other species (e.g., A. modestus (Reinhard), treated in part 1 of this work) whose hosts also pupate in shallow soil. In culture experiments A. dissector was found to prefer hosts in the early to middle part of the 3rd instar, although late 2nd instar host were often also acceptable. Oviposition into suitable hosts was rapid (1–2 seconds) and accomplished with a single insertion of the ovipositor, following only brief antennation and no use of the legs. There was no clear temporary paralysis. Experimental rearings from O. incerta (6:107\85\\75+10) and O. gothica (6:61\49\\34+15) were comparable (given that some insertions of less than a full second might have been scored as ovipositions incorrectly; and furthermore that some failures to oviposit into these hosts might be ascribed to temporary egg depletion, as the protocol of normally ceasing to offer hosts to a particular female after four apparent ovipositions on the day had not been developed until after the experiments were undertaken), and clearly demonstrated the suitability of both species as hosts. In contrast, no parasitoids developed (and indeed possibly no ovipositions occurred) in the other species of Orthosia tested, which were all found to be clearly outside the host range: O. cerasi (Fabricius) (3:32\?3\\0+3); O. cruda (Denis & Schiffermüller) (2:12\0\\-); O. munda (Denis & Schiffermüller) (3:10\0\\-); O. gracilis (Denis & Schiffermüller) (2:11\?1\\0+1). Of these four, only O. gracilis is not fully arboreal. There is no adverse venom effect on host development.
Diagnosis
Maximum width of hypoclypeal depression 0.6–0.7 × minimum width of face (Fig.
Description
Redescribed ♀ (RMNH) from Austria (Burgenland, Winden am See). Length of fore wing 8.5 mm, of body 9.0 mm.
Head.
Antennal segments of ♀ 60, antenna as long as fore wing, its subapical segments rather slender, slightly longer than wide; frons largely smooth; OOL 0.7× diameter of posterior ocellus, sparsely punctate, shiny and with deep groove near posterior ocellus (Fig.
Mesosoma. Mesoscutal lobes punctulate with interspaces superficially micro-sculptured and shiny; precoxal area of mesopleuron smooth except some punctulation, mesopleuron punctulate anteriorly and posteriorly; metapleuron densely punctate; metanotum with nearly complete median carina; scutellum flat (but with rugulose depression medio-posteriorly), remainder punctulate and with weak lateral carinae; propodeum evenly convex and coarsely rugose, and medio-longitudinal carina absent posteriorly.
Wings.
Fore wing: r 0.4 × 3-SR; 1-CU1 horizontal, 0.3 × 2-CU1; r-m 0.3 × 3-SR; 2nd submarginal cell medium-sized (Fig.
Legs.
Tarsal claws with conspicuous and robust dark brown pecten (Fig.
Metasoma.
First tergite flattened, basally narrowed, as long as wide apically; 1st and 2nd tergites with medio-longitudinal carina and largely finely punctate-rugose, but posterior quarter of 2nd tergite irregularly rugose and no median carina; medio-basal area of 2nd tergite wide and triangular, distinct (Fig.
Colour. Black; apical half of hind tibia and hind tarsus blackish; basal half of hind tibia pale yellowish; remainder of legs, palpi and tegulae yellowish brown; most veins and pterostigma dark brown; wing membrane slightly yellowish basally and remainder slightly infuscate.
Variation.
Interspaces between punctulation of mesoscutum smooth to superficially micro-sculptured; medio-longitudinal carina of propodeum complete or absent posteriorly; 3rd metasomal tergite largely finely sculptured (except posteriorly) to largely smooth; mesopleuron black or with brownish longitudinal stripe; hind tibia usually ivory or pale yellowish basally. Antennal segments: ♀ 51(2), 55(2), 56(7), 57(4), 58(7), 59(12), 60(14), 61(18), 62(6), 63(4); ♂ 51(1), 53(2), 54(2), 55(4), 56(8), 57(29), 58(29), 59(27), 60(15), 61(4), 62(4), 63(2), 64(2). Females have on average ca one to two more antennal segments than males. Males are very similar but hind femur more or less blackish and, in some males, hind tibia almost uniformly dark, OOL approx. as long as diameter of posterior ocellus (Fig.
Distribution
*Armenia, *Austria, British Isles (England, Scotland), Croatia, Czech Republic, Finland, France, Germany, *Greece, Hungary, *Montenegro, *Italy, Netherlands, Norway, Russia, *Serbia, Switzerland, Ukraine.

Aleiodes dissector (Nees), U.K., Scotland (in culture) parasitising Orthosia incerta (Hufnagel) 234 pre-mummy, removed from its hideaway, with unparasitised control from the same egg batch (below) 235 pre-mummy 236 early mummification 237 mummy with ventral ooze 238 three fully hard mummies 239 emerged mummy, cut open to expose silken lining.


Aleiodes dissector (Nees), ♀, Switzerland, Tessin 243 fore wing 244 hind wing 245 mesosoma lateral 246 mesosoma dorsal 247 propodeum and 1st –4th metasomal tergites dorsal 248 fore femur lateral 249 hind femur lateral 250 outer hind tarsal claw 251 head anterior 252 head dorsal 253 head lateral 254 base of antenna 255 apex of antenna.

Aleiodes diversus
Rhogas diversus
Szépligeti, 1903: 114;
Rogas dissector var. diversus;
Aleiodes (Neorhogas) diversus;
Aleiodes diversus;
Type material
Lectotype, ♀ (
Additional material
Austria, British Isles (England: V.C.s 8, 25, 70), Bulgaria, Croatia, Hungary, Italy (Sicily), Norway, Switzerland. Specimens in
Molecular data
None.
Biology
Unknown. Female specimens have been collected in (May–)June, and also September, suggesting that it may be plurivoltine. This is reinforced by the date of capture of the two available males (which would not have hibernated as an adult) in Sicily on 30.iv.1965 (
Diagnosis
Maximum width of hypoclypeal depression 0.5–0.6 × minimum width of face (Fig.
Description
Lectotype, ♀, length of fore wing 7.0 mm, of body 10.0 mm.
Head.
Antennal segments of ♀ 56, antenna as long as fore wing, its subapical segments robust; frons largely smooth behind antennal sockets; OOL 1.2 × diameter of posterior ocellus, and coarsely punctate, interspaces less than diameter of puncture; vertex coarsely punctate; clypeus rugose; ventral margin of clypeus thick and not protruding forwards (Fig.
Mesosoma. Mesoscutal lobes densely and finely punctate, interspaces largely smooth, shiny; precoxal area of mesopleuron coarsely punctate and without rugae medially, mesopleuron coarsely punctate anteriorly and posteriorly; metapleuron moderately punctate; scutellum remotely punctate; propodeum rather convex and coarsely rugose.
Wings.
Fore wing: r 0.5 × 3-SR (Fig.
Legs.
Tarsal claws with rather conspicuous, medium-sized dark brown pecten up to apical tooth (Fig.
Metasoma.
First tergite rather flattened, as long as wide apically; 1st and 2nd tergites with medio-longitudinal carina and coarsely vermiculate-rugose; medio-basal area of 2nd tergite triangular and distinct (Fig.
Colour. Black; mesoscutum (except anterior third), scutellum, clypeus ventrally, mandible, tegulae and legs largely brownish red; palpi, fore coxa largely, telotarsi, hind tarsus and apex of hind tibia (excluding spurs) dark brown; pterostigma blackish brown; veins dark brown, but near wing base yellowish; wing membrane slightly infuscate.
Variation.
OOL 1.0–1.2 × diameter of posterior ocellus; mesoscutum of ♀ entirely brownish red or yellowish brown, or anteriorly black; 1st tergite 1.0–1.1 × longer than wide apically; metasoma rarely partly obscurely reddish dark brown; mesopleuron may be just punctate or may have some rugae in lower half. Antennal segments ♀: 55(3), 56(3), 57(3), 58(1), 59(1); ♂ 58(1). Males have mesosoma black (Fig.
Distribution
*Austria, *British Isles (England; probably extinct), *Bulgaria, Croatia, Hungary, *Italy (Sicily), *Norway, *Switzerland.
Notes
Close to A. cruentus which, however, almost always has much or all of 1st and 2nd metasomal tergites orange-red (usually wholly black or dark brown in A. diversus). In addition to characters given in the key A. diversus is a more robust insect, and females have broader antennal segments (distinctly transverse near middle of flagellum) and on average they are fewer in number (although with overlap).


Aleiodes diversus (Szépligeti), ♀, Italy, Sicily 266 wings 267 mesosoma lateral 268 mesosoma dorsal 269 propodeum and metasoma dorsal 270 fore femur lateral 271 hind femur lateral 272 outer hind tarsal claw 273 head anterior 274 head dorsal 275 head lateral 276 base of antenna 277 apex of antenna.

Aleiodes eurinus
Rhogas (Rhogas) eurinus Telenga, 1941: 422.
Rogas eurinus;
Rogas (Rogas) eurinus;
Aleiodes (Neorhogas) eurinus;
Aleiodes (Chelonorhogas) eurinus; Chen and He, 1997: 39;
Aleiodes eurinus;
Rogas eurinus ab. nigratus Papp, 1967: 223 (invalid name).
Rogas eurinus ab. nigrimaculatus Papp, 1967: 223 (invalid name).
Rogas eurinus ab. nigripes Papp, 1967: 223 (invalid name).
Type material
None seen.
Additional material
Italy, Russia (Siberia and Far East), Spain, Turkey, [China, Mongolia]. Specimens in
Molecular data
None.
Biology
Specimens have been collected from April to August, and the presence of males in both April and July clearly demonstrates that it is plurivoltine and overwinters in the mummy. We have not seen reared material, but specimen labelling indicates that it occurs among Ammophila and Schoenus in the Venice Lido and Triticum (presumably cultivated wheat) in Turkey, suggesting that its hosts will occur in open grassland habitats.
Diagnosis
Maximum width of hypoclypeal depression 0.5–0.6 × minimum width of face (Fig.
Description
Redescribed ♀ (RMNH) from Turkey (Ankara). Length of fore wing 7.0 mm, of body 8.0 mm.
Head.
Antennal segments of ♀ 51 remaining, but apical segments missing, length of antenna 1.2 × fore wing; frons with coarse curved rugae and dorsally coarsely rugose; OOL equal to diameter of posterior ocellus, and densely rugose; vertex spaced rugose, rather dull; clypeus medium-sized and coarsely rugose (as face); ventral margin of clypeus rather thin and rather protruding forwards (Fig.
Mesosoma. Mesoscutal lobes distinctly punctate-granulate, and with satin sheen; precoxal area of mesopleuron coarsely rugose, rather wide medially and posteriorly coarsely punctate and some short rugae, densely punctate; remainder of mesopleuron mainly sparsely and finely punctate; metapleuron densely punctate; metanotum with nearly complete median carina; scutellum punctulate and weakly granulate; propodeum coarsely vermiculate-rugose, medio-longitudinal carina irregular.
Wings.
Fore wing: r 0.4 × 3-SR (Fig.
Legs.
Tarsal claws slender and brownish setose (Fig.
Metasoma.
First tergite rather flattened, 1.1 × longer than wide apically; 1st and 2nd tergites with medio-longitudinal carina and rather regularly longitudinally rugose; medio-basal area of 2nd tergite narrow triangular (Fig.
Colour. Black; palpi and basal half of antenna (except scapus and pedicellus) brown; scapus, pedicellus, clypeus largely, apex of hind femur (but ventrally reddish), apex of hind tibia, hind tarsus, all telotarsi, pterostigma (but basally narrowly pale) and veins (except yellowish veins of basal quarter of wings) dark brown; remainder of legs and 1st–3rd tergites orange brown; tegulae and hind tibia (except apically) pale yellowish; wing membrane subhyaline.
Variation.
Coxae and hind femur (except its basal third) largely dark brown, black or orange brown; apical half of hind tibia dark brown or only apically so; 1st tergite largely dark brown (except posteriorly), with pair of dark brown spots or entirely orange or reddish brown; apical half of 3rd tergite orange brown or largely black. Antennal segments: ♀ 54(1), 55(2), 57(2), 58(3), 59(1), 60(2); ♂ 52(1), 60(1). Male is very similar and has apical tergites type 1–2, setae moderately dense, glabrous stripe only rarely evident and fringe very short, negligible (Figs
Distribution
China, *Italy, Mongolia, Russia (Siberia and Far East), Spain, *Turkey.



Aleiodes fahringeri
Rhogas (Rhogas) fahringeri Telenga, 1941: 173.
Rogas fahringeri;
Rogas (Rogas) fahringeri;
Aleiodes (Chelonorhogas) fahringeri;
Aleiodes fahringeri; Chen and He, 1992: 125;
Rhogas (Rhogas) flavipennis Telenga, 1941: 174, 419.
Rogas flavipennis;
Aleiodes (Chelonorhogas) flavipennis;
Type material
None examined.
Additional material
3 ♀ (
Molecular data
None.
Biology
Unknown. Specimens have been collected in June–August. Presumed to be univoltine, but we have not seen reared material and the means of overwintering is unclear.
Diagnosis
Maximum width of hypoclypeal depression 0.6–0.7 × minimum width of face; OOL 0.9 × diameter of posterior ocellus, largely smooth with spaced punctures; ventral margin of clypeus thin, anterior part shiny and distinctly protruding anteriorly (Fig.
Description
Redescribed ♀ (RMNH) from Mongolia (Somon Bulgan). Length of fore wing 6.9 mm, of body 7.7 mm.
Head.
Antennal segments of ♀ 58, length of antenna 1.1 × fore wing, its basal and subapical segments slender (Figs
Mesosoma. Pronotum medio-dorsally flat, shiny and largely smooth; mesoscutal lobes largely smooth except for punctulation, shiny and densely setose; precoxal area of mesopleuron largely smooth medially, with only some superficial rugulae; remainder of mesopleuron finely punctate and antero-dorsally rugose; metapleuron remotely punctate and largely smooth medially; scutellum remotely punctulate; metanotum with fine complete median carina; propodeum weakly convex and densely rugose, its medio-longitudinal carina complete and fine.
Wings.
Fore wing: r 0.5 × 3-SR (Fig.
Legs.
Tarsal claws with rather inconspicuous and pale brownish pecten remaining far removed from apical tooth (Fig.
Metasoma.
First tergite rather flat, 1.1 × as long as wide apically; 1st and 2nd tergites with fine medio-longitudinal carina and finely longitudinally (1st) or irregularly (2nd) densely rugose; medio-basal area of 2nd tergite triangular and medium-sized (Fig.
Colour. Yellowish brown; antenna (except dark brown scapus and pedicellus), stemmaticum and ovipositor sheath black; tarsi, medio-posterior patch of propodeum, basal patch of 1st tergite and apex of hind tibia dark brown; veins rather dark brown at medial third of fore wing, remainder of veins pale brown or yellowish; pterostigma brownish yellow; wing membrane subhyaline.
Variation. Scapus entirely dark brown or largely yellowish brown; dark patches of propodeum and 1st tergite sometimes absent (♀ RMNH from Ningxia). Antennal segments: ♀ 56(2), 58(2), 59(1); ♂ 57(1), 59(1). Male is very similar with apical tergites type ?1–2, setae short, sparse and hard to see, with fringe very short and negligible.
Distribution
China (Ningxia), Mongolia.
Notes
This Asian species is included here because it was reported from Poland (Huflejt, 1997). The record needs confirmation to rule out confusion with a similar European species.


Aleiodes fahringeri (Telenga), ♀, Mongolia, Somon Bulgan 310 wings 311 mesosoma lateral 312 mesosoma dorsal 313 propodeum and 1st–3rd metasomal tergites dorsal 314 fore femur lateral 315 hind femur lateral 316 base of antenna 317 apex of antenna 318 head anterior 319 head dorsal 320 head lateral 321 inner hind tarsal claw
Aleiodes fortipes
Rogas fortipes
Reinhard, 1863: 272;
Aleiodes (Neorhogas) fortipes;
Aleiodes fortipes;
Rhogas freyi
Hellén, 1927: 25–26;
Rogas freyi;
Type material
Holotype of A. fortipes, ♂ (
Additional material
Austria, British Isles (England: V.C.s 16, 26, 28), Bulgaria, Czech Republic, Finland, France, Germany, Hungary, Netherlands (GE: ‘t Harde, Nunspeet), Poland, Spain, Sweden, Turkey. Specimens in
Molecular data
MRS650 (France), MRS807 (Poland).
Biology
The flight time of this univoltine species is (April)May–June, and ca 10 months of the year is spent as an exposed mummy. The only mummy seen (Fig.
There are two particularly significant aspects to the successful rearing. The first is that these Idaea species overwinter as quite well-grown larvae, so during the flight period of the parasitoid they are in late instars, and attacking hosts at this stage is an unusual strategy for Aleiodes (but see A. aterrimus and A. sibiricus). The second is that we know of no other Aleiodes species apart from A. sibiricus (q. v.) among those whose host overwinters as a larva that fails to take advantage of that to overwinter as an early instar larva within it. The apparently riskier strategy taken by A. fortipes, in both respects, may be plesiomorphic.
Aleiodes fortipes is the only known West Palaearctic species in which males have small, subapical setose pore (probably associated with pheromone release) situated mid-dorsally on each of the 4th–6th metasomal tergites (Fig.
Aleiodes fortipes is the only species among those treated in this part of our revision with known hosts outside the Noctuidae and, although no host is known for rather a lot of these species, the apparently basal position of A. fortipes in the group is noteworthy and using geometrid hosts may also be plesiomorphic. The rather slender ovipositor sheath (Fig.
Diagnosis
Maximum width of hypoclypeal depression approx. 0.3 × minimum width of face (Fig.
Description
Holotype of A. freyi, ♂, length of fore wing 4.5 mm, of body 5.3 mm.
Head.
Antenna incomplete, (length of antenna of ♀ from Santon Downham 1.2 × fore wing, its subapical segments rather robust: Fig.
Mesosoma. Mesoscutal lobes very densely coriaceous-granulate, with vague micro-reticulate sculpture, matt; precoxal area of mesopleuron largely smooth (except some micro-sculpture) medially, rather depressed; remainder of mesopleuron largely smooth, except some punctures and antero-dorsally coarsely rugose; scutellum superficially granulate and with some punctures; propodeum coarsely rugose-reticulate and medio-longitudinal carina nearly complete.
Wings.
Fore wing: r 0.6 × 3-SR (Fig.
Legs.
Tarsal claws small but robust and only yellowish setose (Fig.
Metasoma.
First tergite evenly convex, 1.3 × as long as wide apically; 1st and 2nd tergites with weak medio-longitudinal carina and together with basal half of 3rd tergite densely and finely longitudinally rugose; medio-basal area of 2nd tergite narrow but rather distinct (Fig.
Colour. Dark brown or blackish; palpi dark brown; mesopleuron with reddish brown streak; legs yellowish brown but tarsi, apex of hind femur (and indistinctly apices of fore and middle femora, and of tibiae) and base of hind coxa infuscate; tegulae and pterostigma brown; wing membrane slightly infuscate.
Variation.
Maximum width of marginal cell of hind wing 2.0–2.6 × its width near hamuli (Fig.
Distribution
*Austria, *British Isles (England), Bulgaria, Czech Republic, Finland, France, *Germany, Hungary, *Netherlands, *Poland, *Spain, *Sweden, *Turkey.


Aleiodes fortipes (Reinhard), ♀, England, Santon Downham 325 fore wing 326 hind wing 327 mesosoma lateral 328 mesosoma dorsal 329 1st –3rd metasomal tergites dorsal 330 fore femur lateral 331 hind femur lateral 332 head anterior 333 head dorsal 334 head lateral 335 base of antenna 336 apex of antenna 337 antenna 338 inner hind tarsal claw.

Aleiodes gasterator
Bracon gasterator Jurine, 1807: 118, pl. 8. [examined].
Rogas gasterator;
Rogas (Rogas) gasterator;
Aleiodes (Neorhogas) gasterator;
Aleiodes (Chelonorhogas) gasterator; Falco et al. 1997: 60;
Aleiodes gasterator;
Bracon dimidiatus
Aleiodes dimidiatus;
Rogas (Rogas) dimidiatus:
Rogas dimidiatus;
Aleiodes (Neorhogas) dimidiatus;
Aleiodes (Chelonorhogas) dimidiatus;
Rhogas (Rhogas) dimidiatus var. turkestanicus
Telenga, 1941: 184, 409;
Type material
Holotype of A. gasterator, ♀ (Museum Genève), “[? Switzerland], gasterator J.”, “Typus”, “Bracon gasterator Jur., Type”, “Type du g. Rogas [= incorrect]”, “vu par [R.D.] Shenefelt, U.S.A., 1967” (metasoma on separate card and pin). Lectotype of A. turkestanicus here designated, ♀ (
Additional material
Albania, Cyprus, France (including Corsica), Greece (including Crete), Italy (including Sardinia, Sicily), North Macedonia, Portugal (including Madeira), Spain (including Mallorca, Menorca, Tenerife), Tunisia, Turkey, [Iraq, Jordan, Syria]. Specimens in
Molecular data
MRS046 (France), MRS048 (France), MRS892 (Spain).
Biology
Collected chiefly in May–July and September–November, but specimens have occurred in every month of the year. Plurivoltine; there is no indication of a unique overwintering mode in the material seen. Reared from low-feeding Noctuidae: Agrotis segetum (Denis & Schiffermüller) (6 [6
Diagnosis
Maximum width of hypoclypeal depression 0.4–0.5 × minimum width of face (Fig.
Description
Redescribed ♀ (RMNH) from France (Isle sur le Sorque). Length of fore wing 4.9 mm, of body 6.1 mm.
Head.
Antennal segments of ♀ 35, length of antenna 0.9 × fore wing, its subapical segments robust (Fig.
Mesosoma. Mesoscutal lobes densely punctate, interspaces largely smooth with superficial granulation, shiny; precoxal area of mesopleuron evenly vermiculate-rugose medially, but only sparsely punctate posteriorly; metanotum without median carina; scutellum rather flat, sparsely punctate, but rugose laterally; propodeum coarsely vermiculate-rugose, medio-longitudinal carina nearly complete, and angulate latero-posteriorly.
Wings.
Fore wing: r 0.35 × 3-SR (Fig.
Legs.
Tarsal claws robust and with only bristly brownish setae (Fig.
Metasoma.
First tergite convex medially and 0.9 × as long as wide apically; 1st and 2nd tergites with medio-longitudinal carina and coarsely longitudinally rugose; medio-basal area of 2nd tergite short and rather distinct (Fig.
Colour. Black; face (except medio-dorsally), malar space, dorsal half of temple, frons largely laterally, notauli, mesoscutum laterally, scutellum, pronotum postero-dorsally, mesopleuron dorsally and posteriorly, metapleuron largely, 1st and 2nd metasomal tergites and base of 3rd tergite orange brown; palpi and humeral plate and veins of hind wing yellowish brown; tegula rather dark brownish; ventral half of temple largely, dorso-apical patch of hind femur, pterostigma and veins of fore wing dark brown; fore wing membrane slightly infuscate, of hind wing subhyaline.
Variation.
A very colour-variable species; head and mesoscutum of female may be largely black (nominate form) or reddish (= “A. dimidiatus / var. turkestanicus”, but especially the mesoscutum may be intermediate). Especially males may have the hind coxa black and most of hind tibia dark brown, sometimes the entire leg is nearly completely black or dark brown. Antennal segments: ♀ 29(1), 31(2), 32(3), 33(13), 34(13), 35(15), 36(9), 37(9), 38(3), 39(2); ♂ 36(2), 37(3), 38(4), 39(8), 40(13), 41(9), 42(5), 43(11), 44(7), 45(2), 46(2). Additionally, an exceptionally large male with 50 segmented antennae from Cyprus (
Distribution
*Albania, *Cyprus, France (including Corsica), Greece (including Crete), *Iraq, Italy (including Sardinia, Sicily), *Jordan, *North Macedonia, *Portugal (including Madeira), Spain (including Mallorca, Menorca and Tenerife), *Syria, *Tunisia, *Turkmenistan, Turkey.
New synonymy
The new synonymy of Rhogas dimidiatus var. turkestanicus Telenga, 1941, is based on direct comparison of the types of both taxa. The identity of Bracon dimidiatus Spinola, 1808, is problematic because the holotype from Italy (Genoa) is lost and the original description is far too incomplete for an easy identification. Its colour pattern (head completely yellowish, hind tibia and 3rd tergite reddish) does not fit with A. ruficornis (Herrich-Schäffer); if the head is largely reddish brown then the temple ventrally and malar space remain blackish. This pattern agrees better with that of pale specimens of A. gasterator (named as A. dimidiatus var. turkestanicus Telenga, 1941). Aleiodes ruficornis occurs also in Italy, but its females have the head partly black ventrally, the apex of the hind tibia dark brown and most of the 3rd metasomal tergite black. Therefore, we synonymise Bracon dimidiatus with A. gasterator (syn. nov.). The holotype of Bracon gasterator Jurine, 1807, has the 3rd metasomal tergite finely curved (nearly circular) aciculate or striate basally, palpi (as far as present) pale brownish, maximum with of hypoclypeal depression 0.45 × minimum width of face, vein 1-CU1 of fore wing half as long as vein 2-CU1 and 4th antennal segment 1.3 × as long as wide. Aleiodes arnoldii sensu


Aleiodes gasterator (Jurine), ♀, France, Les Constants 347 fore wing 348 hind wing 349 mesosoma lateral 350 mesosoma dorsal 351 metasoma dorsal 352 fore femur lateral 353 hind femur lateral 354 base of antenna 355 head anterior 356 head dorsal 357 head lateral 358 outer hind tarsal claw 359 apex of antenna.

Aleiodes grassator
Ichneumon grassator Thunberg, 1822: 256 [examined].
Rogas grassator;
Aleiodes (Neorhogas) grassator;
Aleiodes grassator;
Rhogas grassator ab. thoracicus
Hellén, 1927: 24;
Rogas (Rogas) flavipalpis Thomson, 1892: 1672 [examined].
Aleiodes flavipalpis;
Rogas alpinus
Thomson, 1892: 1671;
Aleiodes alpinus;
Type material
Holotype of A. grassator, ♀ (
Additional material
Austria, British Isles (England: V.C. 70; Scotland: V.C.s 83, 85, 88, 89, 97, 103; Finland, France (both Alps and Pyrenees), Italy, Germany, Norway, Romania, Sweden, Switzerland. Specimens in
Molecular data
MRS215 (UK), MRS721 (UK), MRS725 (UK).
Biology
Collected in (April)May–July. Univoltine, overwintering in the mummy. Reared from the noctuid Cerapteryx graminis (Linnaeus) (9: K.P. Bland, M.J.W. Cock, M.R. Shaw) and from mummies compatible with that (3), and it may be strictly monophagous. The known host overwinters in the egg stage, and feeds on Poaceae near ground level. The tough dark brown mummy is formed on or below the soil surface and seems spectacularly too large for the adult that will emerge from it (Fig.
Diagnosis
Maximum width of hypoclypeal depression 0.4–0.5 × minimum width of face (Fig.
Description
Redescribed ♀ (RMNH) from Finland (Sb: Leppävirta). Length of fore wing 4.6 mm, of body 5.7 mm.
Head.
Antennal segments of ♀ 39, 4th segment 0.8 × longer than wide (Fig.
Mesosoma. Mesoscutal lobes moderately punctate, laterally interspaces mainly smooth, medially superficially granulate and rather shiny; precoxal area of mesopleuron coarsely rugose medially and largely smooth posteriorly; remainder of mesopleuron mainly punctate, but dorsally coarsely rugose; scutellum flat, sparsely finely punctate and only anteriorly with lateral carina; propodeum coarsely rugose, medio-longitudinal carina present on anterior half, rounded posteriorly and dorsal part approx. as long as posterior part.
Wings.
Fore wing: r 0.3 × 3-SR (Fig.
Legs.
Tarsal claws robust and with only brownish bristly setae (Fig.
Metasoma.
First tergite rather flattened, 0.8 × as long as wide apically; 1st and 2nd tergites with medio-longitudinal carina and coarsely longitudinally rugose; medio-basal area of 2nd tergite wide triangular and short (Fig.
Colour. Orange brown; apical two thirds of antenna, labial palp, patch on hind femur dorso-apically, posterior patch of 2nd tergite and telotarsi, dark brown; head, mesosoma (except side of pronotum postero-dorsally and pair of latero-posterior patches of propodeum), 3rd–7th tergites (except antero-lateral corners of 3rd tergite) black; maxillary palp, basal third of antenna, tegulae and remainder of legs rather pale yellowish brown; veins and pterostigma dark brown; wings distinctly infuscate but hind wing less than fore wing.
Variation.
Basal third or half of antenna of ♀ pale yellowish brown; head partly and mesosoma anteriorly of ♀ dark orange brown or both entirely black; 3rd tergite longitudinally striate or rugulose basally (sometimes narrowly so), without curved sculptural elements (Fig.
Distribution
*Austria, British Isles (England, Scotland), Finland, *France, *Ireland, *Italy, *Germany, Norway, *Romania, Sweden, *Switzerland.
New synonymy
The synonymy of Rogas alpinus Thomson, 1892, with Aleiodes grassator (Thunberg, 1822) is based on direct comparison of the types listed above.
Notes
Although males of A. carbonaroides are generally easily distinguished from A. grassator through being black, it is possible that lighter forms occur which would be difficult to recognise. Also, females of A. carbonaroides are similar in colour to those of A. grassator. Therefore, specimens collected at low altitude away from northern areas that appear, on other characters, to be A. grassator might well really be A. carbonariodes. See also remarks under A. carbonarius and A. ruficornis.


Aleiodes grassator (Thunberg), ♀, Scotland, Beinn Ghlas 368 fore wing 369 hind wing 370 mesosoma lateral 371 mesosoma dorsal 372 metasoma dorsal 373 fore femur lateral 374 hind femur lateral 375 head anterior 376 head dorsal 377 head lateral 378 base of antenna 379 apex of antenna 380 outer hind tarsal claw.

Aleiodes hemipterus
Rhogas hemipterus Marshall, 1897: 137.
Rogas hemipterus;
Aleiodes hemipterus;
Aleiodes (Chelonorhogas) hemipterus;
Type material
Lectotype here designated, ♀ (
Additional material
1 ♀ + 1 ♂ (
Molecular data
None.
Biology. Unknown. The specimens seen do not have phenological data, and we have not seen reared material. As the female is brachypterous it is likely that the host will be found near the ground.
Diagnosis
Maximum width of hypoclypeal depression approx. 0.6 × minimum width of face (Fig.
Description
Lectotype, ♀, length of hind wing 1.7 mm (fore wing missing, but in other specimens ca one-third longer than hind wing and 2.2 mm, brachypterous), of body 7.8 mm.
Head.
Antenna incomplete, segments robust; frons largely striate-rugose (but transversely costate in figured ♀); OOL 1.2 × diameter of posterior ocellus, (as vertex) rather finely and densely reticulate-rugose and rather dull; clypeus rugose; ventral margin of clypeus rather thick ventrally and rather forward protruding (Fig.
Mesosoma.
Antescutal depression distinct; mesoscutal lobes coarsely rugose-punctate (but superficial in figured ♀) and rather matt, middle lobe of pair of submedian grooves (Fig.
Wings.
Fore wing brachypterous, hardly surpassing propodeum (Marshall, 1897): (of ♀ from Tunisia r 0.2 × 3-SR; 1-CU1 distinctly widened and oblique, 0.4 × 2-CU1; r-m 0.8 × 3-SR; 2nd submarginal cell medium-sized (Fig.
Legs.
Tarsal claws with rather conspicuous pale brown pecten, remaining far from apical tooth and much shorter (Fig.
Metasoma.
First tergite evenly convex, as long as wide apically; 1st–3rd tergites regularly finely and very densely longitudinally rugose, rather matt and medio-longitudinal carina rather weak; medio-basal area of 2nd tergite triangular and short (Fig.
Colour. Brown; stemmaticum and ovipositor sheath black; frons, vertex medially, occiput, femora, propodeum, 1st and 2nd tergites somewhat infuscate; wing membrane subhyaline.
Variation. Length of body 7.8–8.8 mm. Antennal segments: ♀ 46(1), 50(1); ♂ unknown. Male is normally winged (vein 3-SR of fore wing 1.5 × vein 2-SR, vein r 0.3 × 3-SR, vein cu-a oblique, vein 1-CU1 narrow and 0.3 × vein 2-CU1) and has marginal cell of hind wing 2.2 × wider than width at level of hamuli (with vein m-cu present anteriorly, 2-SC+R quadrate and M+CU:1-M:1r-m = 40:30:26). Apical metasomal segments of ♂ type 1 and sparsely setose.
Distribution
Morocco, Tunisia.
Notes



Aleiodes hirtus
Rogas hirtus
Thomson, 1892: 1672;
Aleiodes (Neorhogas) hirtus;
Aleiodes hirtus;
Rhogas hirtus ab. coloratus
Hellén, 1927: 23;
Type material
Lectotype of A. hirtus, ♂ (
Additional material
Austria, Belgium, British Isles (England: V.C.s 26, 29, 32, 62; Scotland: V.C. 78; Ireland: V.C. H12), Bulgaria, Czech Republic, Finland, France, Germany, Hungary, Netherlands (DR: Borger), Norway, Romania, Russia, Serbia, Slovakia, Switzerland, Ukraine, [? Mongolia]. Specimens in
Molecular data
MRS619 (UK), MRS882 (Romania), MRS883 (Romania).
Biology
Unknown. Collected in June–August, presumably univoltine but the mode of overwintering is unclear. Most British sites are more or less damp and calcareous grasslands, approaching fens. We have not seen reared material, but the clypeus suggests that the mummy will form in the soil.
Diagnosis
Maximum width of hypoclypeal depression 0.5–0.6 × minimum width of face (Fig.
Description
Redescribed ♂ (RMNH) from Germany (Graswang). Length of fore wing 6.2 mm, of body 8.0 mm. Entire body with long whitish setae.
Head.
Antennal segments of ♂ 60, length of antenna 1.3 × fore wing, its subapical segments somewhat longer than wide; frons medially largely smooth, laterally with some fine curved rugae; OOL 1.6 × diameter of posterior ocellus, and punctate-rugose, POL approx. half as long as diameter of ocellus; vertex spaced rugose, shiny; clypeus punctate; ventral margin of clypeus rather thick but distinctly protruding forwards (Fig.
Mesosoma. Pronotum rugose and anteriorly oblique, without antescutal depression; mesoscutal lobes large smooth and shiny, only indistinctly punctulate and densely setose; precoxal area of mesopleuron punctulate and medially with some superficial rugulae; remainder of mesopleuron sparsely punctate; scutellum sparsely punctate and largely smooth, posteriorly with lateral rugae; propodeum rather convex and coarsely rugose, its medio-longitudinal carina only in anterior half of propodeum.
Wings.
Fore wing: r 0.3 × 3-SR (Fig.
Legs.
Tarsal claws with rather conspicuous and medium-sized brownish pecten (Fig.
Metasoma.
First tergite evenly convex, approx. as long as wide apically; 1st and 2nd tergites with medio-longitudinal carina and densely longitudinally rugose, but posterior quarter of 2nd tergite irregularly rugose and no median carina; medio-basal area of 2nd tergite minute (Fig.
Colour. Black; legs (except black coxae, trochanters and trochantelli, 1st and 2nd metasomal tergites (but base of 1st tergite partly infuscate) and base of 3rd tergite orange brown; vaguely near base of femora, telotarsi, apex of hind femur, apical half of hind tibia, hind tarsus largely black or blackish; basal half of hind tibia pale yellow; palpi, tegulae, veins and pterostigma dark brown; wing membrane slightly infuscate.
Variation.
Hind femur varies from apically black to entirely orange. Propodeum can be partly orangish in posterior part. Usually both sexes have hind trochanter (often also trochantellus) more or less infuscate and darker than the orange part of the hind femur, but this is scarcely evident in a series from S. Russia (MRC,
Distribution
*Austria, *Belgium, *British Isles (England, Scotland, Ireland), *Bulgaria, *Czech Republic, Finland, *France, *Germany, *Hungary, *Netherlands, Norway, *Romania, *Russia, *Serbia, *Slovakia, *Switzerland, *Ukraine.


Aleiodes hirtus (Thomson), ♀, Scotland, Peebles 411 fore wing 412 hind wing 413 mesosoma lateral 414 mesosoma dorsal 415 metasoma dorsal 416 fore femur lateral 417 hind femur lateral 418 antenna 419 head anterior 420 head dorsal 421 head lateral 422 base of antenna 423 apex of antenna 424 inner hind tarsal claw.

Aleiodes improvisus , sp. nov.
Type material
Holotype, ♀ (
Molecular data
None.
Biology
Unknown. Collected above the tree line in the Alps in June–July, and presumably univoltine.
Diagnosis
Maximum width of hypoclypeal depression 0.4–0.5 × minimum width of face (Fig.
Description
Holotype, ♀, length of fore wing 5.5 mm, of body 7.7 mm.
Head.
Antennal segments of ♀ 41, antenna as long as fore wing, its subbasal and subapical segments rather robust (Fig.
Mesosoma. Mesoscutal lobes densely punctate, interspaces largely finely coriaceous and with satin sheen; precoxal area of mesopleuron coarsely vermiculate-rugose medially, but posteriorly rugose; mesopleuron remotely punctate and shiny medially; metapleuron densely rugose and rather dull; scutellum largely smooth (except for spaced punctures), shiny and nearly flat, with lateral carina; propodeum coarsely rugose but antero-laterally rugulose, laterally dorsal face longer than posterior one, somewhat angulate laterally but without tubercles, and with complete medio-longitudinal carina.
Wings.
Fore wing: r 0.4 × 3-SR; marginal cell fairly short (Fig.
Legs.
Tarsal claws mainly setose but submedially with four rather short and dark brown pectinal teeth (Fig.
Metasoma.
First tergite distinctly convex medially, its length 0.8 × apical width, robust and irregularly longitudinally rugose as 2nd tergite; both tergites with medio-longitudinal carina; medio-basal area of 2nd tergite triangular and small (Fig.
Colour. Black; antenna (but only scapus partly yellowish), right fore coxa, trochanter, trochantellus, and femur (but left all yellowish brown except dark base of coxa and infuscated apex of femur), middle femur dorso-apically, middle coxa basally, hind trochanter, trochantellus and femur (but dorso-basally yellowish and left femur also ventrally), apical third of hind tibia (but left tibia yellowish ventrally), tegulae, pterostigma, veins largely, and metasoma ventrally largely dark brown; dorsal part of scutellum, 1st tergite laterally and narrowly medially and posteriorly, 2nd tergite and antero-laterally 3rd tergite orange brown; right fore tibia (except basally and left one yellowish brown) and tarsi more or less infuscate (but left fore tarsus only telotarsus dark brown); fore wing membrane somewhat infuscate, but hind wing nearly subhyaline.
Variation.
Eye of ♀ as long as temple in dorsal view (of ♂ 1.0–1.4 ×); length of malar space 0.5–0.6 × length of eye in lateral view; palpi black or largely dark brown; 1-CU1 0.3–0.6 × 2-CU1; length of fore wing 4.0–6.5 mm. Antennal segments: ♀ 41(1); ♂ 44(2), 49(2), 51(2). Male often has much darker legs (largely dark brown with coxae black as right legs of holotype, but legs are more extensively orange, including basal half of hind femur, in two paratypes) than female and scutellum black; metasoma similarly sculptured and coloured or also basal half of 3rd tergite orange brown or 1st tergite only posteriorly orange or only 2nd and 3rd tergites (except posteriorly) dark reddish brown; in the largest male paratype (Winterstallen) traces of inwardly curved sculpture are discernible posteriorly on the almost completely longitudinally rugose 3rd tergite; marginal cell of fore wing similar to ♀, with apical tergites type 1 and fringe not observed (Figs
Distribution
Austria, Switzerland.
Etymology
Improvisus is Latin for unexpected, unforeseen, because at first sight the specimens were expected to belong to A. gasterator or A. ruficornis.
Notes
As suggested by its name this species can be easily confused with A. gasterator or A. ruficornis. It differs from A. gasterator mainly by being darker (subbasal antennal segments of ♀, hind trochanter and trochantellus, inner and dorsal side of hind tibia, parastigma) and somewhat higher number of antennal segments of ♀ (41 vs 29–39). Aleiodes ruficornis has an inflated fore femur (hardly or not inflated in A. improvisus), antenna of ♀ medium-sized (1.0–1.2 × fore wing vs 0.8–0.9 ×) and head of ♀ at least partly reddish brown.


Aleiodes improvisus sp. nov., ♀, holotype 432 fore wing 433 hind wing 434 mesosoma lateral 435 mesosoma dorsal 436 propodeum and 1st –3rd metasomal tergites dorsal 437 left fore femur lateral 438 left hind femur lateral 439 head anterior 440 head dorsal 441 head lateral 442 inner hind tarsal claw 443 base of antenna 444 apex of antenna.

Aleiodes krulikowskii
Rhogas (Rhogas) krulikowskii
Kokujev, 1898: 302;
Rogas krulikowskii;
Rogas (Rogas) krulikovskii;
Rogas (Rogas) krulikowskii;
Aleiodes (Neorhogas) krulikowskii;
Aleiodes (Chelonorhogas) krulikowskii;
Aleiodes krulikowskii;
Rhogas csikii Szépligeti, 1901 150.
Rogas csikii;
Aleiodes csikii; Papp 1991: 84 (as synonym of A. jaroslawensis); 2004: 216 (as synonym of A. krulikowskii).
Type material
Lectotype of A. krulikowskii, ♀ (
Additional material
1 ♀ (
Molecular data
None.
Biology
Unknown. The collection dates (June–August) suggest that it is univoltine, but there is nothing to suggest how it overwinters.
Diagnosis
Maximum width of hypoclypeal depression 0.7–0.8 × minimum width of face (Fig.
Description
Lectotype, ♀, length of fore wing 6.9 mm, of body 9.6 mm.
Head.
Antenna incomplete, 32 segments remaining; frons mainly with curved or oblique rugae; OOL 1.3 × diameter of posterior ocellus, coarsely rugose and rather matt; vertex densely and rather finely rugose, hardly shiny; anterior part of clypeus densely punctate and flat; ventral margin of clypeus thin and rather forward protruding (Fig.
Mesosoma. Lateral mesoscutal lobes densely and coarsely punctate, with interspaces narrower than punctures but interspaces becoming wider posteriorly, middle lobe coriaceous, but punctate near narrow and distinctly impressed notauli; precoxal area of mesopleuron and metapleuron coarsely and densely rugose punctate; remainder of mesopleuron densely punctate; metanotum with incomplete median carina; scutellum coarsely punctate; axilla largely densely rugose; propodeum rather flat and coarsely reticulate-rugose, medio-longitudinal carina on only anterior half.
Wings.
Fore wing: r 0.6 × 3-SR (Fig.
Legs.
Hind tarsal claws robust and with inconspicuous fine subbasal brownish pecten (Fig.
Metasoma.
First tergite rather convex and moderately widened (Fig.
Colour. Black; orbit near ocelli reddish brown; fore and middle legs (except blackish or dark brown coxae, trochanters and trochantelli), apex of hind trochantellus and basal third of hind tibia brownish yellow; tarsi darkened and remainder of legs dark brown; palpi (except basally) pale yellowish; mandible yellowish but basally largely dark brown; propleuron and tegula anteriorly dark brown and tegula posteriorly brown; 1st and 2nd metasomal tergites and metasoma ventrally (except apically) orange brown; pterostigma dark brown; veins brown; wing membrane subhyaline.
Variation. Orbit near hind ocellus sometimes only very slightly lighter in colour. Antennal segments: ♂ 68(1); according to the original descriptions of A. krulikowskii and A. csikii, the female types have 60 and 62 antennal segments, respectively. Apical tergites of ♂ type 2 and no fringe observed.
Distribution
*Hungary, Mongolia, *Romania, Russia (Central and Far East).


Aleiodes miniatus
Rogas miniatus
Herrich-Schäffer, 1838: 156;
Rogas (Rogas) miniatus;
Aleiodes (Neorhogas) miniatus;
Aleiodes (Chelonorhogas) miniatus;
Aleiodes miniatus;
Rogas bicoloratus
Boheman, 1853: 180;
Aleiodes formosus
Giraud, 1857: 177;
Type material
Lectotype of A. formosus, ♀ (
Additional material
Austria, Czech Republic, France, Finland, Germany, Hungary, Romania, Russia, Sweden, Ukraine, [Kazakhstan, Kyrgyzstan]. Specimens in
Molecular data
MRS950 (Sweden), MRS951 (Sweden).
Biology
Unknown, but it seems to inhabit herb-rich calcareous steppe grasslands. Collected in (May)June–August; presumably univoltine, but we have not examined reared material of this large and distinctive species and there is no indication of how it may overwinter. A series in BZL (one now in
Diagnosis
Maximum width of hypoclypeal depression approx. 0.5 × minimum width of face (Fig.
Description
Redescribed ♀ (RMNH) from Russia (Yaaseni). Length of fore wing 6.5 mm, of body 7.9 mm.
Head.
Antennal segments of ♀ 65, length of antenna 1.1 × fore wing, its subapical segments somewhat longer than wide; frons with coarse curved rugae; OOL 2.3 × diameter of posterior ocellus, and punctate; vertex densely punctate and shiny; clypeus densely punctate; ventral margin of clypeus thin and distinctly protruding forwards (Fig.
Mesosoma. Mesoscutal lobes densely punctate, with minute interspaces and rather shiny; precoxal area of mesopleuron wide and coarsely rugose medially, mesopleuron above it coarsely and densely punctate, even speculum with some punctures; scutellum convex and punctate; propodeum evenly convex and coarsely reticulate-rugose, medio-longitudinal carina incomplete.
Wings.
Fore wing: r 0.7 × 3-SR (Fig.
Legs.
Tarsal claws with only three conspicuous brownish and widened bristles basally (Fig.
Metasoma.
First tergite evenly convex, 0.9 × as long as wide apically; 1st tergite coarsely reticulate-rugose, 2nd tergite coarsely and densely rugose-punctate, without median carina; medio-basal area of 2nd tergite short triangular (Fig.
Colour. Brownish yellow; antenna, mesosternum, mesopleuron (except antero-dorsally), metapleuron, propodeum, 1st tergite, and ovipositor sheath black; propleuron, small patch on middle mesoscutal lobe anteriorly, apices of femora, fore and middle tibiae, tarsi, apical half of hind tibia, veins, and pterostigma dark brown; wing membrane subhyaline; basal half of hind tibia pale yellowish.
Variation. Second submarginal cell square or somewhat narrower; propleuron dark brown or yellowish; mesopleuron black or yellowish anteriorly and dorsally; medio-longitudinal carina of posterior half of propodeum absent, obsolescent or incomplete. Antennal segments: ♀ 64(5), 65(3), 66(2), 67(3), 68(2), 70(1); ♂ 61(1), 64(2), 66(1), 67(2), 68(1), 69(1), 70(1). On this limited evidence there seems to be little, if any, difference in the number of antennal segments between the sexes. Males are very similar but have the metasoma infuscated apically and the apical tergites are type 3, setae short and dense, glabrous stripe rather narrow and fringe not observed.
Distribution
Austria, Czech Republic, Finland, *France, Germany, Hungary, Kazakhstan, *Kyrgyzstan, *Romania, Russia, Sweden, *Ukraine.


Aleiodes miniatus (Herrich-Schäffer)), ♀, Russia, Stavropolskij kraj 469 fore wing 470 hind wing 471 mesosoma lateral 472 mesosoma dorsal 473 metasoma dorsal 474 fore femur lateral 475 hind femur lateral 476 head anterior 477 head dorsal 478 head lateral 479 base of antenna 480 apex of antenna 481 inner hind tarsal claw.
Aleiodes morio
Rogas morio
Reinhard, 1863: 255;
Rogas (Rogas) morio;
Aleiodes (Neorhogas) morio;
Aleiodes (Chelonorhogas) morio;
Aleiodes morio;
Type material
Lectotype ♀ from southern Germany most probably lost (
Additional material
1 ♀ (
Molecular data
None.
Biology
Unknown. Specimens of both sexes have been collected in April and May, from which from which it is safe to surmise that the winter is passed in the mummy as the male would not hibernate. But we have seen no reared material nor any indication of habitat for this central European species. Its early flight time might be one reason why it is seldom collected and apparently rare.
Diagnosis
Maximum width of hypoclypeal depression 0.5–0.6 × minimum width of face; OOL of ♀ 0.8 × (of ♂ 0.9 ×) diameter of posterior ocellus and rugose; ventral margin of anterior part of clypeus comparatively sharp and more or less protruding in lateral view (Fig.
Description
Redescribed ♀ (
Head.
Antenna incomplete, 47 segments remaining (54 in lectotype), length of complete antenna approx. 0.9 × fore wing, its subbasal and subapical segments short; frons largely rugose; OOL 0.8 × diameter of posterior ocellus, and mainly rugulose and with satin sheen; depression near posterior ocellus smooth; vertex densely rugulose and with satin sheen; clypeus with some punctures; ventral margin of clypeus rather thin and protruding forwards (Fig.
Mesosoma. Mesoscutal lobes densely and finely punctate, with satin sheen; precoxal area of mesopleuron widely and densely rugose, but densely punctate posteriorly; middle of mesopleuron densely rugulose and dorsally coarsely rugose; metapleuron largely rugose; scutellum punctate-coriaceous; propodeum rather flat and densely rugose or rugulose, medio-longitudinal carina complete, and without protruding carinae laterally.
Wings.
Fore wing: r 0.5 × 3-SR (Fig.
Legs.
Tarsal claws bristly setose, medium-sized, and with robust pecten basally (cf. Fig.
Metasoma.
First tergite evenly convex, 0.9 × as long as wide apically; 1st tergite with medio-longitudinal carina; 1st and 2nd tergites and basal half of 3rd tergite finely and densely longitudinally rugulose; medio-basal area of 2nd tergite short triangular (Fig.
Colour. Black; palpi brownish yellow, but basally dark brown; tegulae pale yellowish; legs (except pale base of tibiae), metasoma ventrally and veins dark brown; pterostigma brown and medially yellowish brown; wing membrane subhyaline.
Variation.
Clypeus distinctly to moderately protruding and ventrally rather thin to thick. Antennal segments of ♀ 51(1), 52(1), 54(1), of ♂ 55(1). Males are very similar to the redescribed female (including the wing venation: Fig.
Distribution
*Finland, Germany, Hungary.
Notes
The lost lectotype from Germany had hyaline wings (which separates it from the A. carbonarius/ grassator/ carbonaroides complex), the pterostigma paler medially than laterally (entirely dark brown), base of the hind tibia pale yellow (black in ♂) and the body of ♀ entirely black (more or less yellowish or reddish).


Aleiodes morio (Reinhard), ♀, Hungary, Budapest, but 490 of ♂ from Nadap 484 fore wing 485 hind wing 486 mesosoma lateral 487 mesosoma dorsal 488 1st–3rd metasomal tergites dorsal 489 fore femur lateral 490 hind femur lateral 491 head anterior 492 head dorsal 493 head lateral 494 outer middle tarsal claw 495 base of antenna.


Aleiodes nigrifemur , sp. nov.
Type material
Holotype, ♀, (RMNH), “Greece, Peloponn[esus], Chelmos, 1700 m, 29.v.1987, H. Teunissen”.
Molecular data
None.
Biology
Unknown; the only known specimen was collected at the end of May which gives no clue of voltinism or how the winter is passed.
Diagnosis
Maximum width of hypoclypeal depression 0.6 × minimum width of face (Fig.
Description
Holotype, ♀, length of fore wing 7.2 mm, of body 8.2 mm.
Head.
Antennal segments of ♀ 62, length of antenna 1.1 × fore wing, its subapical segments medium-sized (Fig.
Mesosoma. Mesoscutal lobes densely punctate, rather shiny and interspaces coriaceous; precoxal area of mesopleuron widely rugose but posteriorly punctate, and area above it densely punctate or rugulose; metapleuron densely punctate dorsally and rugose ventrally; metanotum with short median carina anteriorly; scutellum remotely punctate, with some lateral rugae; propodeum rather short and flat, coarsely reticulate-rugose, medio-longitudinal carina complete, and without protruding carinae laterally.
Wings.
Fore wing: r 0.5 × 3-SR (Fig.
Legs.
Tarsal claws rather robust, bristly setose and few small yellowish teeth (Fig.
Metasoma.
First tergite rather flat medially, 0.8 × as long as wide apically; 1st tergite and anterior half of 2nd tergite with medio-longitudinal carina; 1st–2nd tergites densely longitudinally rugose; 3rd tergite (except posterior third) mainly rugulose; medio-basal area of 2nd tergite triangular and rather distinct (Fig.
Colour.
Black; maxillary palp apically, basal 0.4 of hind tibia and tegulae pale yellowish; mandible (but with dark brown patch), side of pronotum dorso-posteriorly, fore and middle tibiae, hind basitarsus basally, 1st tergite apically, 2nd–5th tergites orange brown; remainder of legs dark brown; remainder of palp, veins and pterostigma dark brown; lateral lobes of mesoscutum (except anteriorly and medially) dark reddish brown; wing membrane subhyaline, but apically infuscated (Fig.
Distribution
Greece (main).
Etymology
The species is named after its black femur; niger is Latin for black, dark, dusky.


Aleiodes nigrifemur sp. nov. ♀, holotype 506 wings 507 mesosoma lateral 508 mesosoma dorsal 509 1st–3rd metasomal tergites dorsal 510 fore femur lateral 511 hind femur lateral 512 base of antenna 513 head anterior 514 head dorsal 515 head lateral 516 apex of antenna 517 outer hind tarsal claw 518 antennae.
Aleiodes nobilis
Rogas nobilis
Haliday [in Curtis], 1834: 512;
Rogas ductor var. nobilis;
Aleiodes (Neorhogas) nobilis;
Aleiodes (Chelonorhogas) nobilis;
Aleiodes nobilis;
Rogas medianus
Thomson, 1892: 1668;
Rogas (Rogas) medianus;
Aleiodes (Neorhogas) medianus;
Aleiodes medianus;
Aleiodes ductor; auct. p.p.
Type material
Neotype of A. nobilis here designated: ♀ (
Additional material
Austria, British Isles (Scotland: V.C.s 72, 88, 105; Ireland: V.C. H29), Bulgaria, Croatia, Czech Republic, Finland, Germany, Hungary, Italy, Moldova, Netherlands (LI: Gulpen; St. Pietersberg; Geulle (Bunderbos); NB: Udenhout (“de Brand”), OV: Voorst (Twello), ZH: Lexmond), Poland, Romania, Russia, Serbia, Slovakia, Sweden. Specimens in ALC,
Molecular data
MRS401 (Finland), MRS880 (Russia), MRS881 (UK).
Biology
Collected predominantly in grassy places, June–October. Reared from the noctuid Autographa gamma (Linnaeus) (4 [1
Diagnosis
Maximum width of hypoclypeal depression approx. 0.3 × minimum width of face (Fig.
Description
Redescribed ♀ (RMNH) from Slovakia (Kubrica). Length of fore wing 5.1 mm, of body 5.9 mm.
Head.
Antennal segments of ♀ 48, length of antenna 1.25 × fore wing, its subapical segments slender (Fig.
Mesosoma. Mesoscutal lobes densely and finely granulate, rather shiny near tegulae; precoxal area of mesopleuron smooth, surroundings sparsely punctulate; metapleuron mostly granulate; metanotum without median carina; scutellum granulate and with lateral carina; propodeum slightly convex, granulate with spaced rugosity, medio-longitudinal carina only anteriorly present, and no protruding carinae laterally.
Wings.
Fore wing: r 0.6 × 3-SR (Fig.
Legs.
Tarsal claws with conspicuous and robust dark brown pecten (Figs
Metasoma.
First tergite rather flattened, as long as wide apically; 1st and 2nd tergites rather regularly sublongitudinally striate, without medio-longitudinal carina on 2nd tergite; medio-basal area of 2nd tergite wide triangular and rather distinct (Fig.
Colour. Black; pterostigma (except yellowish extreme base and apex), veins (except brown vein C+SC+R), clypeus, apical third of hind tibia and telotarsus dark brown; palpi, tegulae, remainder of tibiae and tarsi, pale yellowish; apex of middle femur and apical half hind femur, black; remainder of legs, antenna (but apical segments and to some degree scapus infuscate) yellowish brown; 1st–3rd metasomal tergites (except black medial patch of 1st tergite), propleuron and pronotum orange; wing membrane subhyaline.
Variation.
1-CU1 0.7–1.2 × 2-CU1; striae of 2nd tergite regularly sublongitudinal or somewhat diverging posteriorly (Fig.
Distribution
*Austria, British Isles (Scotland, Ireland), Bulgaria, *Croatia, Czech Republic, Finland, Germany, Hungary, Italy, Moldova, Netherlands, *Poland, *Romania, Russia, *Serbia, *Slovakia, Sweden.


Aleiodes nobilis (Haliday), ♀, neotype 522 wings 523 mesosoma lateral 524 mesosoma dorsal 525 1st–4th metasomal tergites dorsal 526 fore femur lateral 527 hind femur lateral 528 hind tibia lateral 529 head anterior 530 head dorsal 531 head lateral 532 base of antenna 533 apex of antenna 534 outer hind tarsal claw 535 outer fore tarsal claw.


Aleiodes pallidicornis
Rogas pallidicornis
Herrich-Schäffer, 1838: 156;
Rhogas pallidicornis;
Rogas (Rogas) pallidicornis;
Aleiodes (Neorhogas) pallidicornis;
Aleiodes (Chelonorhogas) pallidicornis;
Aleiodes pallidicornis;
Rhogas pallidipennis Dalla Torre, 1898: 221 [invalid emendation].
Rogas ductor
auctt. p.p. [North & Central Europe, e.g.,
Type material
Neotype of A. pallidicornis here designated, ♀ (RMNH), “[Netherlands], [Zuid-]Holland, Asperen, 6.viii.1972, C.J. Zwakhals”. The neotype designation is necessary for nomenclatorial stability, because the types of Braconidae described by Herrich-Schäffer are lost (Horn and Kahle 1935–37; no specimens could be found by the first author in
Additional material
Austria, Belarus, British Isles (Scotland: V.C. ?92), Bulgaria, Germany, Hungary, Italy, Montenegro, Netherlands (ZH: Asperen; Schoonrewoerd; Waarder), Romania, Russia, Slovakia, Switzerland, Turkey [Iran, North Korea]. Specimens in
Molecular data
MRS885 (Russia).
Biology
Very little is known. Specimens collected in (May) June–August (September), the great majority in June-July strongly suggesting that it is at least largely univoltine. The Dutch specimens were collected in fairly humid coppice woods. The single British specimen (
Diagnosis
Maximum width of hypoclypeal depression approx. 0.3 × minimum width of face (Fig.
Description. Neotype, ♀, length of fore wing 5.9 mm, of body 6.6 mm.
Head.
Antennal segments of ♀ 54, length of antenna 1.3 × fore wing, its subapical segments rather robust (Fig.
Mesosoma. Mesoscutal lobes finely punctate with largely granulate interspaces, with satin sheen; precoxal area of mesopleuron distinctly remotely punctate, interspaces larger than punctures; metapleuron densely punctate-granulate; metanotum with median carina; scutellum punctate-granulate; propodeum evenly convex and coarsely rugose, its medio-longitudinal carina complete.
Wings.
Fore wing: r 0.4 × 3-SR (Fig.
Legs.
Tarsal claws with rather small dark brownish pecten, absent near apical tooth (Fig.
Metasoma.
First tergite rather flattened, as long as wide apically; 1st and 2nd tergites with medio-longitudinal carina and coarsely irregularly rugose, but posteriorly 2nd tergite largely smooth and no median carina; medio-basal area of 2nd tergite triangular and rather large (Fig.
Colour. Black; hind tarsus largely infuscate, but 3rd and 4th segments paler than other segments; apices of fore and middle tibiae slightly infuscate, base of middle and hind tibiae and telotarsi dark brown; apical two-fifths of hind femur and hind tibia (except a pale yellowish band subbasally) black; remainder of legs, 1st and 2nd tergites, and 3rd tergite antero-laterally orange brown; palpi and tegulae brownish yellow; most of veins and pterostigma dark brown; wing membrane subhyaline.
Variation. Antennal segments: ♀ 49(2), 50(2), 51(2), 52(6), 53(6), 54(1), 56(1); ♂ 50(1), 51(3), 52(1), 53(2), 54(2), 55(1), 56(3). On average males have ca two more antennal segments than females. Males are similar but have a large dark brown patch on 1st tergite, hind tarsus largely dark brown and apical tergites type 3, positioned rather posteriorly, setae long and fringe not observed.
Distribution
Austria, *Belarus, *British Isles (Scotland), Bulgaria, Hungary, *Iran, *Italy, *Montenegro, *Netherlands, North Korea, *Romania, Russia, *Slovakia, Switzerland, *Turkey.
Notes
The type of Rogas pallidicornis Herrich-Schäffer, 1838, has been lost. Traditionally, it has been considered to belong to Aleiodes ductor (Thunberg, 1822), but the latter species is a synonym (see under A. unipunctator). The inadequate original description indicates that the 2nd tergite has diverging rugae, which excludes part of A. ductor auctt. Female specimens with yellowish or brownish palpi, basal half of the antenna yellowish and blackish hind tibia (except its pale yellowish base) fit well the original description of A. pallidicornis.


Aleiodes pallidicornis (Herrich-Schäffer), ♀, neotype 545 fore wing 546 hind wing 547 mesosoma lateral 548 mesosoma dorsal 549 1st–3rd metasomal tergites dorsal 550 fore femur lateral 551 hind femur lateral 552 head anterior 553 head dorsal 554 head lateral 555 base of antenna 556 apex of antenna 557 inner hind tarsal claw.
Aleiodes pallidistigmus
Rhogas (Rhogas) pallidistigmus Telenga, 1941: xii, 143, 177 (but also as palidistigma (p. 409) and pallidistigma (p. 420)) [examined].
Rogas pallidistigmus;
Aleiodes (Neorhogas) pallidistigmus;
Aleiodes (Chelonorhogas) pallidistigmus;
Aleiodes pallidistigma;
Rogas heterostigma
Stelfox, 1953: 149;
Aleiodes (Neorhogas) heterostigma;
Aleiodes heterostigma;
Type material
Paratypes of A. heterostigma, 4 ♀ + 1 ♂ (
Additional material
1 ♀ (RMNH), “Belgium: Liège, Mt. Rigi, 650 m, 2.viii.1986, at light, C. v. Achterberg, RMNH”; 1 ♀ (
Molecular data
None.
Biology
Unknown for West Palaearctic populations. Specimens have been collected in (June)July-August in open boggy areas, certainly at least sometimes over limestone. Presumably univoltine, but we have not seen reared material from West Palaearctic and the overwintering mode is unclear.
Diagnosis
Maximum width of hypoclypeal depression 0.4–0.5 × minimum width of face (Fig.
Description
Redescribed ♀ paratype of A. heterostigma (RMNH) from Ireland (Rye Water). Length of fore wing 4.9 mm, of body 6.3 mm.
Head.
Antennal segments of ♀ 58, length of antenna 1.1 × fore wing, its subapical segments rather robust (Fig.
Mesosoma. Mesoscutal lobes densely and finely punctate-granulate, matt; precoxal area of mesopleuron coarsely rugose medially; remainder of mesopleuron coarsely punctate, with some rugae near speculum and interspaces superficially granulate; scutellum rather flat, punctulate-granulate and with weak lateral carinae; propodeum rather convex, shiny and coarsely rugose, medio-longitudinal carina distinct only on its anterior half.
Wings.
Fore wing: r 0.3 × 3-SR (Fig.
Legs.
Tarsal claws rather robust and only brownish setose (Fig.
Metasoma.
First tergite rather flattened, as long as wide apically; 1st and 2nd tergites and base of 3rd tergite finely and irregularly longitudinally rugose, with medio-longitudinal carina weak; medio-basal area of 2nd tergite triangular and short (Fig.
Colour. Black; telotarsi largely and basal quarter of antenna dark brown; palpi, tegulae and pterostigma pale yellow; remainder of legs, 1st and 2nd tergites, basal half of 3rd tergite largely and pronotum orange brown; veins brown; wing membrane subhyaline.
Variation. Pronotum anteriorly and basal half of antenna orange brown to dark brown, pterostigma is yellowish to largely (except base) rather dark brown; length of malar space 1.0–1.4 × basal width of mandible; OOL 0.7–1.8 × diameter of ocellus and metapleuron medially more or less punctate, rugulose-coriaceous or rugose. Antennal segments: ♀ 54(1), 58(1), 59(4), 60(2), 62(1), 64(1); ♂ 59(1), 60(1), 62(1). Apical tergites of ♂ type 1, fringe absent.
Distribution
*Belgium, British Isles (Ireland, Wales), China, *Denmark, *Finland, Russia (Far East).
New synonymy
We tried to separate the East Palaearctic A. pallidistigmus from the West Palaearctic A. heterostigma, but efforts were in vain. The differences such as the colour of the basal half of the antenna (dark brown in A. heterostigma and usually yellowish or brown in A. pallidistigmus), the eyes and ocelli often smaller, OOL 1.1–1.8 × diameter of ocellus (0.7–1.4 ×), malar space 1.2–1.3 × basal width of mandible (1.0–1.4 ×) and metapleuron with a shiny and more or less punctate area (less shiny and rugulose-coriaceous or rugose) are too variable to justify separation of A. heterostigma. Therefore, we synonymise A. heterostigma with A. pallidistigmus (syn. nov.).


Aleiodes pallidistigmus (Telenga), ♀, Denmark, Kragelund Mose 560 fore wing 561 hind wing 562 mesosoma lateral 563 mesosoma dorsal 564 1st–3rd metasomal tergites dorsal 565 fore femur lateral 566 hind femur lateral 567 apex of antenna 568 head anterior 569 head dorsal 570 head lateral 571 outer hind tarsal claw 572 base of antenna.
Aleiodes periscelis
Rogas periscelis
Reinhard, 1863: 254;
Rhogas (Rhogas) periscelis var. charkowensis Kokujev, 1898: 297.
Rhogas (Rhogas) periscelis var. charkoviensis
[sic!];
Rogas (Rogas) periscelis;
Aleiodes (Neorhogas) periscelis;
Aleiodes (Chelonorhogas) periscelis;
Aleiodes periscelis;
Rhogas jaroslawensis Kokujev, 1898: 302. Syn. nov.
Rhogas (Rhogas) jaroslavensis;
Rogas jaroslawensis;
Rogas (Rogas) jaroslavensis;
Rogas (Rogas) jaroslawensis;
Aleiodes (Neorhogas) jaroslawensis;
Aleiodes jaroslawensis;
Type material
Lectotype of R. periscelis, ♂ (
Additional material
Czech Republic, Germany, Hungary, Russia. Specimens in ALC, BZL,
Molecular data
None.
Biology
Unknown but presumably univoltine. Specimens of both sexes collected in April and May suggest that the winter is passed in the mummy. We have not seen reared material, but several Hungarian specimens appear to have been collected in Quercus-dominated woodland, but without indication of any association with Quercus as such.
Diagnosis
Maximum width of hypoclypeal depression approx. 0.4 × minimum width of face (Figs
Description
Redescribed ♀ (BZL), Czech Republic (Pisek); length of fore wing 5.8 mm, of body 8.1 mm.
Head.
Antennal segments 45 (holotype ♀ of A. jaroslawensis: 42), length of antenna approx. as long as fore wing, its subbasal and subapical segments robust (Figs
Mesosoma. Mesoscutum remotely punctulate and with satin sheen, interspaces of lateral lobes largely smooth, and of middle lobe superficially coriaceous; scutellum superficially punctate, laterally rugose; precoxal area of mesopleuron smooth and shiny; metapleuron largely densely punctate, but ventrally coarsely rugose; metanotum with distinct median carina anteriorly; propodeum rather flat and coarsely vermiculate rugose, medio-longitudinal carina complete, and slightly tuberculate laterally.
Wings.
Fore wing: r 0.3 × 3-SR (Fig.
Legs.
Tarsal claws with conspicuous and medium-sized brownish pecten, remaining removed from tarsal tooth (Fig.
Metasoma.
First tergite rather flat posteriorly, wide subbasally and 0.9 × longer than wide apically; 1st and 2nd tergites with coarse medio-longitudinal carina and coarsely longitudinally rugose, but posterior quarter of 2nd tergite rather finely rugose; medio-basal area of 2nd tergite triangular and wide (Fig.
Colour.
Black; palpi dark brown basally and remainder pale brown; antenna (except dark brown scapus and pedicellus), tegulae (but anteriorly dark brown), middle and hind trochanters and trochantelli brownish yellow; fore coxa, trochanter and femur dark brown; basal 0.4 of hind tibia ivory and remainder black; remainder of legs (but hind femur with a blackish patch dorso-apically), 1st and 2nd tergites and basal two thirds of 3rd tergite, largely dark reddish brown; pterostigma dark brown; veins mainly yellowish brown, but medially brown (Figs
Variation.
Holotype of A. jaroslawensis has apex of hind femur yellowish brown (Fig.
Distribution
Austria, Czech Republic, Germany, Hungary, Russia.
New synonymy
The ♀ holotype of Rhogas jaroslawensis lacks the antennae, but according to the original description the antenna was 42-segmented, distinctly shorter than the body, reddish brown, except for the darkened apex and the black scapus. This and the other characters still visible agree well with our interpretation of A. periscelis (except that the hind femur is yellowish brown apically); therefore, we synonymise R. jaroslawensis with A. periscelis (syn. nov.).


Aleiodes periscelis (Reinhard), ♀, Czech Republic, Pisek 577 fore wing 578 hind wing 579 mesosoma lateral 580 mesosoma dorsal 581 metasoma dorsal 582 fore femur lateral 583 hind femur lateral 584 head anterior 585 head dorsal 586 head lateral 587 base of antenna 588 apex of antenna 589 outer middle tarsal claw.


Aleiodes periscelis (Reinhard), ♂, Russia, Serpukhov 591 wings 592 mesosoma lateral 593 mesosoma dorsal 594 1st –5th metasomal tergites dorsal 595 fore femur lateral 596 hind femur lateral 597 hind tarsus lateral 598 head anterior 599 head dorsal 600 head lateral 601 base of antenna 602 apex of antenna 603 inner hind tarsal claw.
Aleiodes pulchripes
Aleiodes pulchripes
Rogas pulchripes;
Rogas (Rogas) pulchripes;
Aleiodes (Neorhogas) pulchripes;
Aleiodes pulchricornis
Kolubajiv, 1962: 27;
Type material
Holotype of A. pulchripes, ♂ (KBIN), “A. pulchripes ♂ mihi”, “A. pulchripes mihi, dét. C. Wesmael”, “Coll. Wesmael”, “Belgique, Charleroi/ teste Papp J., 1983”, “Holotypus”, “Aleiodes pulchripes Wesm., 1838, ♂, Papp, 1983”.
Additional material
Austria, British Isles (England: V.C. 59; Isle of Man: V.C. 71: Ireland: V.C. H21), Czech Republic, Finland, Germany, Hungary, Netherlands (GE: Vierhouten; ZH: Leiden; NH: Amsterdam; Sloten), Russia, Sweden. Specimens in
Molecular data
MRS847 (Sweden), MRS873 (Sweden).
Biology
Collected in (June)July and August. Univoltine, overwintering in an exposed mummy. Reared from the following arboreal acronictine Noctuidae: Acronicta aceris (Linnaeus) (2 [
Diagnosis
Maximum width of hypoclypeal depression 0.3–0.4 × minimum width of face (Fig.
Description
Redescribed ♀ (RMNH) from England (Chat Moss). Length of fore wing 5.3 mm, of body 6.5 mm.
Head.
Antennal segments of ♀ 56, length of antenna 1.3 × fore wing, its subapical segments slender (Fig.
Mesosoma. Mesoscutal lobes coriaceous, rather shiny; precoxal area of mesopleuron smooth as most of mesopleuron; metanotum with medio-longitudinal carina anteriorly; scutellum finely punctate, interspaces smooth, but posteriorly coriaceous; propodeum rather flat medially and rather remote rugose, medio-longitudinal carina nearly complete, and with slightly protruding carinae laterally.
Wings.
Fore wing: r 0.4 × 3-SR (Fig.
Legs.
Tarsal claws with conspicuous and robust dark brown pecten up to apical tooth of claw (Fig.
Metasoma.
First tergite evenly convex medially, 0.9 × longer than wide apically, wider than base of 2nd tergite; 1st and 2nd tergites with medio-longitudinal carina and coarsely irregularly sublongitudinally rugose; medio-basal area of 2nd tergite triangular and rather large (Fig.
Colour. Blackish or dark brown; telotarsi, apical 0.4 of hind tibia, hind tibial spurs and hind tarsus dark brown; remainder of hind tibia and palpi yellowish; remainder of legs, pterostigma and tegulae pale brownish yellow; veins brown; mesoscutum medio-dorsally, scutellum, metanotum, mesopleuron (except partly antero-dorsally), mesosternum and metapleuron orange yellow; wing membrane subhyaline.
Variation.
Scutellum largely finely punctate, coriaceous medio-posteriorly, but may be striate. Specimens from Sweden are appreciably darker than those from Britain. Vein m-cu of hind wing absent or faintly indicated. Antennal segments: ♀ 51(1), 52(2), 53(2), 54(3), 55(4), 56(6), 57(7), 58(7), 59(2), 60(1), 62(1); ♂ 49(1), 51(1), 53(2), 54(9), 55(6), 56(6), 57(2). Females have on average ca three more antennal segments than males. Males have obtuse hind tibial spurs and the tarsal pecten less developed than in females, propleuron and pronotum yellowish or blackish posteriorly; posterior half of mesoscutum largely yellowish or blackish; apical tergites type 2, somewhat sparse setose, glabrous stripe broad but with some setae directed into it and fringe rather weak (Figs
Distribution
*Austria, British Isles (England, Isle of Man, Ireland), Czech Republic, Finland, Germany, Hungary, *Netherlands, Russia, Sweden.


Aleiodes pulchripes Wesmael, ♀, England, Chat Moss 608 fore wing 609 hind wing 610 mesosoma lateral 611 mesosoma dorsal 612 1st–3rd metasomal tergites dorsal 613 fore femur lateral 614 hind femur lateral 615 hind tarsus lateral 616 head anterior 617 head dorsal 618 head lateral 619 base of antenna 620 apex of antenna 621 outer hind tarsal claw.

Aleiodes quadrum
Rogas (Rogas) quadrum Tobias, 1976: 83, 221, 1986: 76 (transl.: 125).
Aleiodes (Neorhogas) quadrum;
Aleiodes quadrum;
Rogas (Rogas) illustris Papp, 1977a: 112, 1985a: 162 (as synonym of A. quadrum), 1991a: 83 (id.), 2005: 176 (id.) [examined].
Type material
Holotype of A. illustris, ♀ (
Additional material
1 ♀ (
Molecular data
MRS796 (France), additionally MRS824 (Bulgaria) likely to be a male of this species.
Biology
Unknown. Collected in June–July, likely to be univoltine, but there is nothing to suggest how the winter is passed. We have not seen reared material, but the elongate and strongly apically compressed metasoma suggests that the host would be concealed, perhaps between spun leaves, in a leaf sheath, in a seed capsule or in some similar situation.
Diagnosis
Maximum width of hypoclypeal depression approx. 0.6 × minimum width of face (Fig.
Redescription
♀ (
Head.
Antennal segments of ♀ 53, antenna as long as fore wing, its subapical segments rather robust (Fig.
Mesosoma. Mesoscutal lobes densely and finely punctate, with interspaces approx. equal to diameter of punctures, smooth and shiny; scutellum finely and densely punctate; precoxal sulcus area of mesopleuron distinctly rugose but posteriorly only punctate, remainder of mesopleuron distinctly but remotely punctate; metapleuron remotely punctate, but ventrally rugose; propodeum evenly convex and coarsely vermiculate-rugose and medio-longitudinal carina nearly complete.
Wings.
Fore wing: r 0.4 × 3-SR (Fig.
Legs.
Tarsal claws with conspicuous and robust blackish pecten, close to level of apical tooth (Fig.
Metasoma.
First tergite evenly convex, 1.3 × longer than wide apically; 1st and 2nd tergites with medio-longitudinal carina and finely longitudinally rugose, but posterior quarter of 2nd tergite smooth and no median carina; medio-basal area of 2nd tergite triangular and wide (Fig.
Colour. Black; hind tibia dark brown apically and remainder pale yellowish; middle and hind tarsi, and fore telotarsus blackish or dark brown; mandible, remainder of legs, mesoscutum, pronotum postero-dorsally, mesopleuron dorsally, scutellum, metanotum, ovipositor sheath and 1st tergite (except pair of dark patches apically) orange; tegulae yellowish; palpi and pterostigma blackish; vein dark brown, but vein at base of wings yellowish; wing membrane largely slightly infuscate.
Variation.
OOL of ♀ 0.6–0.7 × diameter of posterior ocellus. The female from Turkey is very similar but has 1st tergite 1.4 × as long as wide apically and 2nd tergite 1.2 × longer than wide basally. Antennal segments: ♀ 53(1), 56(1), 57(1), 58(1). Apical tergites of male type 2 with fringe rather strong (Figs
Distribution
Azerbaijan, *Bulgaria, Croatia, *France, *Greece, Hungary, *North Macedonia, *Turkey.
Notes
The holotype of A. quadrum is a male and it is less reliable to identify this species from it than from the holotype female of A. illustris; nevertheless, we accept the synonymy proposed by


Aleiodes quadrum (Tobias), ♀, France, Accons 629 wings 630 mesosoma lateral 631 mesosoma dorsal 632 metasoma dorsal 633 fore femur lateral 634 hind femur lateral 635 outer hind tarsal claw 636 head anterior 637 head dorsal 638 head lateral 639 base of antenna 640 apex of antenna 641 antenna.


Aleiodes ruficeps
Rhogas (Rhogas) ruficeps Telenga, 1941: 179, 421, Fig. [examined].
Rogas ruficeps;
Rogas (Rogas) ruficeps;
Aleiodes ruficeps;
Rogas gasterator auctt. p.p.
Type material
Lectotype of A. ruficeps, ♀ (
Additional material
Bulgaria, Russia, Turkey, [Armenia, Iran]. Specimens in
Molecular data
None.
Biology
Unknown. Specimens collected in April-May; presumably univoltine, but there is nothing to suggest how it overwinters. We have not seen reared material.
Diagnosis
Maximum width of hypoclypeal depression 0.5–0.6 × minimum width of face (Fig.
Resembles A. grassator because of the robust antennal segments and dark wings, but A. ruficeps has frons, OOL, vertex, malar space, and third tergite less sculptured, apex of hind tibia and palpi dark brown, basal antennal segments of ♀ somewhat less robust, hypoclypeal depression wider, marginal cell of fore wing slenderer and vein r of fore wing shorter. Differs from the similar A. ruficornis by having hypoclypeal depression wider, clypeus wider and lower, apical antennal segments of ♀ slenderer, OOL less sculptured and more antennal segments (♀: 45–47 vs 35–39(–41) of A. ruficornis).
Description
Lectotype, ♀, length of fore wing 7.3 mm, of body 8.6 mm.
Head.
Antennal segments of ♀ 45, length of antenna 1.1 × fore wing, its subapical segments moderately robust (Fig.
Mesosoma. Mesoscutal lobes punctate and interspaces smooth, shiny; precoxal area of mesopleuron coarsely vermiculate-rugose, near precoxal area mesopleuron distinctly punctate; scutellum rather flattened, sparsely punctate; propodeum coarsely vermiculate-rugose, medio-longitudinal carina incomplete, absent posteriorly and propodeum rounded laterally.
Wings.
Fore wing: r 0.3 × 3-SR (Fig.
Legs.
Tarsal claws rather slender, hardly curved and with six brownish bristles (Fig.
Metasoma.
First tergite rather flattened, 0.9 × as long as wide apically; 1st and 2nd tergites with medio-longitudinal carina and regularly longitudinally rugose; medio-basal area of 2nd tergite triangular and short (Fig.
Colour. Orange brown; antenna, palpi, apices of femora, telotarsi, bases of fore and middle coxae, pterostigma, and veins (but 1-SR much paler than 1-M) dark brown; mesosoma (except for mesoscutum and scutellum), ovipositor sheath, 3rd tergite (except basally) and subsequent tergites black or blackish; apex of hind tibia only narrowly dark brown; wing membrane nearly entirely infuscate.
Variation. Vein 1-CU1 of fore wing 0.3–0.5 × vein 2-CU1. Females may have the flagellum extensively pale basally, or entirely dark. Antennal segments: ♀ 45(1), 46(1), 47(1); ♂ 56(1), 58(1). Males appear to have ca ten more antennal segments than females. Male has apical tergites type 1, setae rather long and sparse, and fringe not observed.
Distribution
*Armenia, Bulgaria, *Iran, Russia, *Turkey.


Aleiodes ruficeps (Telenga), ♀, Turkey, Konya 654 fore wing 655 hind wing 656 mesosoma lateral 657 mesosoma dorsal 658 1st –3rd metasomal tergites dorsal 659 fore femur lateral 660 hind femur lateral 661 head anterior 662 head dorsal 663 head lateral 664 base of antenna 665 apex of antenna 666 inner hind tarsal claw.
Aleiodes ruficornis
Rogas ruficornis
Herrich-Schäffer, 1838: 156, fig.;
Aleiodes (Neorhogas) ruficornis;
Aleiodes (Chelonorhogas) ruficornis;
Aleiodes ruficornis;
Aleiodes brevicornis
Wesmael, 1838: 98;
Aleiodes (Neorhogas) brevicornis;
Aleiodes nigripalpis
Wesmael, 1838: 97;
Rhogas dimidiatus ab. nigrobasalis Hellén, 1927: 24 (invalid name).
Rhogas dimidiatus ab. ruficollis Hellén, 1927: 24 (invalid name).
Rhogas carbonarius ab. giraudi Fahringer, 1931: 236 [unavailable name for melanistic males].
Rhogas carbonarius var. giraudi Telenga, 1941: 168. Syn. nov.
Rogas dimidiatus ab. infuscatus Hellén, 1957: 49 (invalid name).
Rogas dimidiatus ab. nigripes Hellén, 1957: 49 (invalid name).
Aleiodes (Aleiodes) arnoldii;
Rogas gasterator auctt. p.p.
Rogas dimidiatus auctt. p.p.
Type material
Neotype of A. ruficornis here designated, ♀ (
Additional material
Andorra, Austria, British Isles (England (V.C. 5, 11, 13, 15, 17, 19, 20, 22, 23, 24, 28, 29, 31, 33, 38, 39, 60, 63); Wales (V.C. 52) [no specimens seen from Scotland]), Bulgaria, Croatia, Czech Republic, Finland, France, Germany, Greece, Hungary, Italy (including Sicily), Moldova, Montenegro, Netherlands (DR: Borger; LI: Venlo, ZH: Oostkapelle), North Macedonia, Norway, Romania, Russia (including Far East), Serbia, Slovakia, Sweden, Switzerland, Turkey, Ukraine, [Afghanistan, Dagestan, Iran, Kazakhstan, Kyrgyzstan]. Specimens in ALC,
Molecular data
MRS140 (UK), MRS877 (Sweden), MRS888 (UK), MRS890 (UK), MRS891 (UK).
Biology
Specimens collected from April–September; probably plurivoltine in the southern part of its range but in Britain univoltine, flying from June–August with a varied means of overwintering (see below). Reared from the noctuids Agrotis clavis (Hufnagel) (4 [4
Diagnosis
Maximum width of hypoclypeal depression 0.4–0.5 × minimum width of face (Fig.
Description
Neotype, ♀, length of fore wing 3.9 mm, of body 6.5 mm.
Head.
Antennal segments of ♀ 35, antenna as long as fore wing, its subbasal and subapical segments robust (Figs
Mesosoma. Mesoscutal lobes distinctly punctate, interspaces of lateral lobes smooth and shiny; precoxal area of mesopleuron coarsely vermiculate-rugose medially, but posteriorly punctate; mesopleuron punctate medially; metapleuron distinctly rugose ventrally and dorsally punctate; scutellum largely smooth (except for punctulation), rather shiny and nearly flat, with lateral carina; propodeum coarsely reticulate-rugose, laterally dorsal face longer than posterior one, somewhat angulate laterally but without tubercles, and with complete medio-longitudinal carina.
Wings.
Fore wing: r 0.3 × 3-SR; marginal cell short (Fig.
Legs.
Tarsal claws mainly setose and medially with 4 yellowish rather short pectinal teeth (Fig.
Metasoma.
First tergite rather convex medially, 0.9 × longer than wide apically, robust and coarsely irregularly longitudinally rugose as 2nd tergite; 1st tergite and basal half of 2nd tergite with medio-longitudinal carina; medio-basal area of 2nd tergite triangular and rather distinct (Fig.
Colour.
Reddish or orange-brown; stemmaticum medially, malar space largely, temple and occiput ventrally, mesosternum, mesopleuron ventrally, 3rd tergite (except antero-laterally), 4th–7th tergites black; palpi (only labial palp darkened basally) brown; basal half of antenna, tegulae, parastigma, and base of pterostigma pale yellowish; apical half of antenna, pedicellus dorsally, propodeum dorsally and medially, middle and hind femora apico-dorsally, and telotarsi dark brown; remainder of pterostigma and veins dark brown or brown (Fig.
Variation.
Female: mesosoma occasionally wholly black. Male face and mesosoma usually black but can be variably marked with red; scape and pedicel usually (partly) reddish in central and southern populations but most often entirely black in more north-western ones (e.g., British Isles, Sweden); hind coxa varies from black to red. Length of malar space 0.5–0.6 × length of eye in lateral view; head black or largely reddish brown (except temple ventrally and malar space); interspaces of mesoscutal lobes smooth to micro-sculptured; 1-CU1 0.4–0.6 × 2-CU1; 3rd tergite longitudinally striate or rugulose basally (sometimes narrowly so), without curved sculptural elements (Fig.
Distribution
*Afghanistan, *Andorra, Austria, British Isles (England, Wales), *Bulgaria, *Croatia, *Czech Republic, *Finland, *France, Germany, *Iran, *Kazakhstan, *Kyrgyzstan, *Montenegro, *Netherlands, *North Macedonia, *Norway, *Romania, *Russia (including Dagestan and Far East), *Serbia, *Slovakia, *Sweden, Switzerland, *Turkey, *Ukraine.
Notes
An examined female (
Aleiodes ruficornis is the commonest and most widespread of a small group of related species parasitising grassland and “cutworm” hosts, exhibiting strong sexual dimorphism with unremarkable males but the more extensively orange females having a stronger build and much shorter antennae. The least extreme in these respects is A. gasterator, which largely (but not completely) replaces A. ruficornis in the Mediterranean region. Aleiodes grassator is similar to A. ruficornis, but it appears to be restricted to montane and northern habitats where it might be thought to replace A. ruficornis. However, some males that morphologically agree best with A. ruficornis have been collected at high altitude in the Alps (up to 2550 m), where A. improvisus also occurs, but whether these high-altitude A. ruficornis males are parts of breeding populations or have simply been carried up in thermals is impossible to say. The females in this group (excluding A. gasterator) are scarcer in collections than males, as they fly very little and rarely enter Malaise traps.


Aleiodes ruficornis (Herrich-Schäffer), ♀, neotype 670 wings 671 mesosoma lateral 672 mesosoma dorsal 673 metasoma dorsal 674 fore femur lateral 675 hind femur lateral 676 head anterior 677 head dorsal 678 head lateral 679 base of antenna 680 apex of antenna 681 outer hind tarsal claw.

Aleiodes rufipes
Rogas rufipes
Thomson, 1892: 1669;
Rogas (Rogas) rufipes;
Aleiodes (Neorhogas) rufipes;
Aleiodes (Chelonorhogas) rufipes;
Aleiodes rufipes;
Type material
Holotype, ♀ (
Additional material
Finland, Norway, Sweden. Specimens in
Molecular data
MRS294 (Sweden), MRS312 (Sweden), MRS314 (Sweden), MRS673 (Finland), MRS674 (Finland), MRS676 (Finland), MRS680 (Finland).
Biology
Unknown. Collected from July–August; presumably univoltine. We have not seen reared material and there is no indication of how the winter may be passed.
Diagnosis
Maximum width of hypoclypeal depression approx. 0.4 × minimum width of face (Fig.
Description
Redescribed ♀ (RMNH) from Finland (Enntekiö). Length of fore wing 5.3 mm, of body 5.8 mm.
Head.
Antennal segments of ♀ 51, antenna as long as fore wing, its subapical segments rather robust and apical segment with short spine (Fig.
Mesosoma. Mesoscutal lobes largely punctate-coriaceous, with satin sheen; precoxal area of mesopleuron partly remotely punctate as its surroundings; medio-longitudinal carina of metanotum distinct posteriorly; scutellum punctate; propodeum convex and coarsely rugose, medio-longitudinal carina absent posteriorly, and without protruding carinae laterally.
Wings.
Fore wing: r 0.3 × 3-SR (Fig.
Legs.
Tarsal claws with rather conspicuous and medium-sized brownish pecten (Fig.
Metasoma.
First tergite convex and basally rather steep, 0.9 × longer than wide apically; 1st and 2nd tergites with medio-longitudinal carina and longitudinally rugose; maximum width of 2nd tergite 1.6 × its median length; medio-basal area of 2nd tergite medium-sized triangular and rather short (Fig.
Colour. Black; mesoscutum largely, legs, and 1st–3rd metasomal tergites (but 3rd tergite narrowly infuscate posteriorly) reddish brown; tegulae brownish yellow; ovipositor sheath, palpi, pterostigma and veins dark brown; telotarsi partly infuscate; wing membrane subhyaline.
Variation. Legs usually largely reddish, but telotarsi, apices of hind femur and tibia frequently dark brown and sometimes most of hind tibia and apical half of hind femur black; clypeus blunt to rather acute ventrally; depression near posterior ocelli smooth or finely crenulate; mesoscutum of ♀ usually partly reddish brown, but sometimes largely or entirely black; ventral third of mesopleuron regularly and finely punctate. Antennal segments: ♀ 57(2), 59(1); ♂ 57(1), 59(1), 60(5), 61(1), 62(1). Male is very similar with mesoscutum black (rarely partly reddish) and at least 2nd tergite orange brown (sometimes with pair of dark brown patches), apical tergites type 1–2 with fringe rather strong when visible, and also often evident on tergites following the third.
Distribution
Finland, Norway, Sweden.


Aleiodes rufipes (Thomson), ♀, Sweden, Lillav 688 fore wing 689 hind wing 690 mesosoma lateral 691 mesosoma dorsal 692 1st–3rd metasomal tergites dorsal 693 fore femur lateral 694 hind femur lateral 695 head anterior 696 head dorsal 697 head lateral 698 outer hind tarsal claw 699 base of antenna 700 apex of antenna.
Aleiodes rugulosus
Bracon rugulosus
Nees, 1811: 32;
Rogas rugulosus;
Rogas (Rogas) rugulosus;
Aleiodes
(Neorhogas) rugulosus;
Aleiodes (Chelonorhogas) rugulosus;
Aleiodes rugulosus;
Rhogas rugulosus var. pictus
Kokujev, 1898: 296;
Type material
Neotype, ♀ (KBIN), “A. rugulosus”, “dét. C. Wesmael”, “Coll. Wesmael”, “Belgique, Bruxelles”/ teste Papp J., 1983”, “Neotypus, Bracon rugulosus Nees, 1812 [sic!], ♀,
Additional material
Albania, Austria, Belgium, Bulgaria, Czech Republic, British Isles (England: V.C.s 1, 3, 4, 11, 17, 25, 27, 29, 69; Wales: V.C.s 41, 48; Scotland: V.C.s 78, 81, 88, 98; Ireland: V.C. H30), Finland, France, Germany, Hungary, Moldova, Netherlands (DR: Borger; GE: ‘t Harde; Heerde; Otterlo, NB: Kampina; NH: Crailo; OV: Buurserzand), North Macedonia, Norway, Poland, Romania, Russia, Slovakia, Spain, Sweden. Specimens in ALC,
Molecular data
MRS191 (Hungary), MRS217 (UK), MRS398 (France), MRS884 (Poland).
Biology
Collected in (May)July–August(September), usually in open habitats such as mosses, heaths, herb-rich grasslands and fens. Partly plurivoltine, at least in the southern part of its range, but largely univoltine in the north (in a UK culture only one female out of 20 reared, and two males out of 69, emerged in the same year as mummy formation). Reared from Noctuidae, Acronictinae: Acronicta auricoma (Denis & Schiffermüller) (7 [1 BZL, 1 MSC, 1
Diagnosis
Maximum width of hypoclypeal depression 0.3–0.4 × minimum width of face (Fig.
Description
Redescribed ♀ (RMNH) from Netherlands (Buurserzand). Length of fore wing 6.3 mm, of body 7.7 mm.
Head.
Antennal segments of ♀ 65, length of antenna 1.3 × fore wing, its subapical segments rather slender (Fig.
Mesosoma. Mesoscutal lobes finely punctate with very finely granulate interspaces, rather matt; precoxal area of mesopleuron smooth, mesopleuron densely punctate posteriorly; scutellum coarsely punctate and rather flat; propodeum rather flattened medially, very coarsely reticulate-rugose, medio-longitudinal carina nearly complete, and with small crest-like protuberances laterally.
Wings.
Fore wing: r 0.6 × 3-SR (Fig.
Legs.
Tarsal claws with conspicuous and robust blackish pecten (Fig.
Metasoma.
First tergite moderately convex, 0.9 × longer than wide apically; 1st and 2nd tergites with coarse medio-longitudinal carina and very coarsely and irregularly longitudinally rugose; medio-basal area of 2nd tergite large and distinct (Fig.
Colour. Black (including fore and middle telotarsi, apical half of hind tibia and hind tarsus); basal half of hind tibia pale yellowish; palpi (but basally somewhat infuscate) and remainder of legs reddish brown; tegulae yellowish brown; hind tibial spurs and pterostigma dark brown; veins brown; mesopleuron with broad dark reddish longitudinal band; wing membrane slightly infuscate.
Variation.
Micro-sculpture of lateral lobes of mesoscutum very finely granulate or absent and resulting in a largely smooth surface; maximum width of marginal cell of hind wing 1.5–2.3 × its width near hamuli; body entirely black (both sexes) or 1st and 2nd metasomal tergites and mesosoma partly dark reddish (both sexes, but 2nd and 3rd tergites of males more or less darkened); hind tibial spurs dark brown or reddish brown. Antennal segments: ♀ 60(4), 61(7), 62(10), 63(18), 64(14), 65(2), 66(8), 67(4), 68(3), 69(1); ♂ 53(1), 56(2), 57(1), 58(7), 59(6), 60(15), 61(15), 62(19), 63(28), 64(12), 65(10), 66(4), 67(2), 68(1). Females have on average ca one more antennal segment than males. Male is similar to the dark female form, with acute hind tibial spurs (Fig.
Aleiodes rugulosus is a very colour-variable species; the 1st and 2nd metasomal tergites are quite frequently mostly or entirely dark red or orange brown (Fig.
Distribution
Albania, Austria, Belgium, Bulgaria, Czech Republic, British Isles (England, Wales, Scotland, Ireland), Finland, France, Germany, Hungary, *Netherlands, *North Macedonia, *Moldova. Norway, Poland, *Romania, Russia, *Slovakia, *Spain, Sweden.


Aleiodes rugulosus (Nees), ♀, U.K., culture 704 wings 705 mesosoma lateral 706 mesosoma dorsal 707 propodeum and 1st–3rd metasomal tergites dorsal 708 fore femur lateral 709 hind femur lateral 710 antenna 711 head anterior 712 head dorsal 713 head lateral 714 base of antenna 715 apex of antenna 716 outer hind tarsal claw.



Aleiodes schewyrewi
Rhogas (Rhogas) schewyrewi Kokujev, 1898: 304.
Rhogas (Rhogas) schevyrevi
[sic!];
Rogas schewyrewi;
Rogas (Rogas) schewyrewi;
Aleiodes (Chelonorhogas) schewyrewi;
Rhogas (Rhogas) schewyrewi var. zaydamensis
Kokujev, 1898: 305;
Rogas schewyrewi var. zaydamensis;
Rogas (Aleiodes) schewyrewi var. zaydamensis;
Type material
Holotype of A. schewyrewi zaydamensis, ♀ (
Additional material
1 ♀ (BZL), “S. Russia, [Volgograd obl.], Elton Lake env., 20.v.2001, J. Miatleuski”; 1 ♂ (
Molecular data
None.
Biology
Unknown. A female collected in May and a male in July may suggest that it is plurivoltine (or, less probably, that the female overwinters as an adult).
Diagnosis
Maximum width of hypoclypeal depression 0.6–0.7 × minimum width of face (Fig.
Description
Holotype, ♀, length of fore wing 5.8 mm, of body 7.5 mm.
Head.
Antennal segments of ♀ 48, length of antenna 1.1 × fore wing, its subapical segments moderately slender (Fig.
Mesosoma. Mesoscutal lobes largely superficially punctate, interspaces finely granulate and with satin sheen; precoxal area of mesopleuron largely smooth medially, densely punctate anteriorly and posteriorly densely rugose, but posterior 0.3 only finely punctate; metapleuron spaced coarsely punctate; metanotum with fine and nearly complete median carina; scutellum sparsely punctate, shiny; propodeum rather convex and rather coarsely rugose, medio-longitudinal carina on anterior 0.4 of propodeum.
Wings.
Fore wing: r curved and 0.4 × 3-SR (Fig.
Legs.
Tarsal claws long and slender, nearly straight and only brownish bristly setose (Fig.
Metasoma.
First tergite evenly convex and strongly widened posteriorly, 0.9 × longer than wide apically; 1st and 2nd tergites with weak medio-longitudinal carina (absent posteriorly) and finely longitudinally rugose, but 2nd tergite smooth medio-posteriorly; medio-basal area of 2nd tergite wide triangular and rather distinct (Fig.
Colour. Yellowish brown (including basal half of antenna); apical half of antenna, frons largely, stemmaticum, occiput dorso-laterally, pronotal side medially, axilla, mesopleuron (except antero-dorsally), mesosternum, metapleuron, propodeum, hind femur and apical third of hind tibia, 5th–7th tergites, last two posterior sternites, ovipositor sheath and pterostigma dark brown or blackish brown; veins brown; wing membrane subhyaline.
Variation. Length of 4th hind tarsal segment 1.8–2.0 × longer than wide; malar space and temple ventrally largely dark brown or yellowish brown. Antennal segments: ♀ 45(1); ♂ 54(1); according to original description ♀ type has 58 segments. Male has clypeus yellowish and contrasting with black face, apical tergites type 1, and no fringe observed.
Distribution
*Iran, Mongolia, *Russia (European part).


Aleiodes schewyrewi var. zaydamensis (Kokujev) 730 fore wing 731 hind wing 732 mesosoma lateral 733 mesosoma dorsal 734 metasoma dorsal 735 fore femur lateral 736 hind femur lateral 737 head anterior 738 head dorsal 739 head lateral 740 outer hind tarsal claw 741 base of antenna 742 apex of antenna.
Aleiodes schirjajewi
Rhogas reticulator var. schirjajewi Kokujev, 1898: 299 [examined].
Rogas schirjajewi;
Rogas (Rogas) schirjaevi
[sic!];
Rogas (Rogas) schirjaewi
[sic!];
Aleiodes (Neorhogas) schirjajewi;
Aleiodes (Chelonorhogas) schirjajewi;
Aleiodes schirjajewi;
Type material
Holotype, ♂ (
Additional material
Bulgaria, Hungary, Italy, Moldova, Russia, Serbia, Ukraine [Dagestan, Kazakhstan]. Specimens in BZL,
Molecular data
None.
Biology
Unknown. Specimens collected throughout April–September, presumably plurivoltine. We have not seen reared material and it is unclear how the winter is passed.
Diagnosis. Maximum width of hypoclypeal depression 0.3–0.4 × minimum width of face (Fig.
Description
Holotype, ♂, length of fore wing 5.0 mm, of body 5.8 mm.
Head.
Antennal segments of ♂ 50, length of antenna 1.3 × fore wing, its subapical segments rather robust; 4th segment of maxillary palp slender and cylindrical; frons with rather coarse curved rugae and interspaces smooth; OOL 1.1 × diameter of posterior ocellus, coarsely rugose and shiny; coarsely transversely rugose and shiny; clypeus punctate-rugulose; ventral margin of clypeus thick and not protruding forwards (Fig.
Mesosoma. Mesoscutal lobes finely punctate, shiny, interspaces micro-sculptured; precoxal area of mesopleuron smooth medially except for some crenulations and punctures, its surroundings smooth; scutellum moderately punctate; propodeum rather convex and coarsely reticulate, coarse medio-longitudinal carina present anteriorly, carinae not protruding laterally.
Wings.
Fore wing: r 0.7 × 3-SR (Fig.
Legs.
Tarsal claws with medium-sized dark brown pecten (Fig.
Metasoma.
First tergite moderately flattened, 0.9 × longer than wide apically; 1st and 2nd tergites with medio-longitudinal carina and densely vermiculate-rugose; medio-basal area of 2nd tergite wide triangular and distinct (Fig.
Colour. Orange brownish; basal half of antenna, palpi largely and parastigma (except base) yellowish brown; head, mesosternum (except anteriorly), mesopleuron (except anteriorly and antero-dorsally), apical 0.4 of hind femur, 3rd tergite (except antero-laterally) and subsequent tergites black; scapus, pedicellus basally, apical half of antenna, apex of fore and middle femora, apex of middle and hind tibiae, hind basitarsus, 2nd hind tarsal segment apically, telotarsi, pterostigma, parastigma basally and veins dark brown; wing membrane subhyaline.
Variation.
Vein 1-CU1 of fore wing 1.0–2.3 × vein 2-CU1; maximum width of marginal cell of hind wing 1.6–2.6 × its width near hamuli (Fig.
Distribution
*Bulgaria, Hungary, *Italy, Kazakhstan, Moldova, Russia (including Dagestan and Far East), Serbia, Ukraine.


Aleiodes schirjajewi (Kokujev), ♀, Hungary, Budapest, but 756 ♀, Ukraine 744 fore wing 745 hind wing 746 mesosoma lateral 747 mesosoma dorsal 748 metasoma dorsal 749 fore femur lateral 750 hind femur lateral 751 head anterior 752 head dorsal 753 head lateral 754 base of antenna 755 apex of antenna 756 outer hind tarsal claw 757 antenna.
Aleiodes sibiricus
Rhogas sibiricus Kokujev, 1903: 286 [examined].
Rogas sibiricus;
Rogas (Rogas) sibiricus;
Aleiodes (Neorhogas) sibiricus;
Aleiodes sibiricus; Papp, 2005: 177.
Rhogas hungaricus
Szépligeti, 1906: 616;
Rogas hungaricus;
Rhogas reinhardi
Fahringer, 1931: 221 (description in key only), 1932: 275 (full description; as R. rheinhardi);
Type material
Paralectotype of A. sibiricus, ♀, (
Additional material
Albania, Austria, Bulgaria, France, Germany, Greece, Hungary, Italy, North Macedonia, Sweden, Turkey. Specimens in
Molecular data
MRS310 (Sweden), MRS313 (Sweden), MRS805 (France).
Biology
Collected in April and May, and presumably univoltine, but 2 ♀ from Sweden: Ångermanland, Lillavammasjon were collected apparently in July in window traps set on the trunks of Betula and Picea. We have examined four males collected in April which suggests spring emergence from the mummy rather than overwintering as an adult. This is corroborated by the data with the single reared specimen examined (
Diagnosis
Maximum width of hypoclypeal depression 0.5–0.6 × minimum width of face (Figs
Description
Paralectotype of A. sibiricus, ♀, length of fore wing 9.1 mm, of body 10.0 mm.
Head.
Antennal segments of ♀ 69, antenna as long as fore wing, its subapical segments medium-sized; frons smooth; OOL 1.1 × diameter of posterior ocellus, and finely coriaceous-rugulose; vertex rugulose and rather dull; clypeus coriaceous and strongly transverse (4–6 × wider than high; Figs
Mesosoma. Mesoscutal lobes densely punctate and interspaces smooth, rather matt; precoxal area of mesopleuron rugose medially and anteriorly, its surroundings moderately punctate; scutellum sparsely punctate and no lateral carina; propodeum rather convex and densely and finely rugose, medio-longitudinal carina complete and no protruding carinae laterally.
Wings.
Fore wing: r 0.3 × 3-SR (Fig.
Legs.
Tarsal claws with four inconspicuous brownish pecten-teeth (Fig.
Metasoma.
First tergite rather flattened, 0.8 × longer than wide apically; 1st and 2nd tergites with weak medio-longitudinal carina and densely finely rugose, but posterior quarter of 2nd tergite irregularly rugose and no median carina; medio-basal area of 2nd tergite wide and short (Fig.
Colour. Black; palpi and tegulae pale yellowish; legs (except black coxae; apex of hind femur dorsally, inner side of hind tibia apically (not outer side!) and telotarsi infuscated), apex of first tergite, 2nd–5th tergites and metasoma ventrally, yellowish brown; ovipositor sheath largely, pterostigma and most veins dark brown; vein 1-R1 of fore wing yellowish brown; wing membrane subhyaline.
Variation. Face, clypeus, mesoscutum, propleuron, upper part of mesopleuron, and first tergite partly, or rarely entirely, reddish brown. Usually in males and rarely in females mesoscutum wholly black; vein r of fore wing 0.3–0.5 × vein 3-SR; clypeus flattened and subparallel-sided or convex and ventrally concave; pterostigma medially dark brown or yellowish brown. Antennal segments ♀: 65(4), 66(8), 67(6), 68(4), 69(2), 70(5), 71(2), 72(2); ♂ 66(1), 71(1). Male apical tergites of type 1 and fringe not observed.
Distribution
*Albania, *Austria, *Bulgaria, *France, Germany, *Greece, Hungary, Italy (main), *North Macedonia, Russia (Siberia), *Sweden, *Turkey.
Notes
The holotype of A. reinhardi (Fahringer, 1931) from Bolzano (N Italy) was examined by



Aleiodes sibiricus (Kokujev), ♀, Hungary, Heves 763 fore wing 764 hind wing 765 mesosoma lateral 766 mesosoma dorsal 767 metasoma dorsal 768 fore femur lateral 769 hind femur lateral 770 head anterior 771 head dorsal 772 head lateral 773 base of antenna 774 apex of antenna 775 outer hind tarsal claw 776 antenna.
Aleiodes turcicus , sp. nov.
Type material
Holotype, ♀ (
Molecular data
MRS126 (Turkey).
Biology
Unknown. The material examined was collected in the period May–June. It is not clear how many generations occur, or how the winter is passed.
Diagnosis
Maximum width of hypoclypeal depression 0.4–0.5 × minimum width of face (Fig.
Description
Holotype, ♀, length of fore wing 5.3 mm, of body 7.9 mm.
Head.
Antennal segments of ♀ 47, length of antenna 1.15 × fore wing, length of 4th segment 1.1 × its width, and its subapical segments 1.2 × as long as wide (Figs
Mesosoma. Mesoscutal lobes smooth between rather remote punctures, strongly shiny, more densely punctate on middle lobe; notauli distinct but shallow, especially posterior half; mesoscutum short setose, widely and strongly rugose medio-posteriorly; scutellum slightly convex, remotely punctate and evenly rounded laterally, no carina; prepectal carina strong, reaching anterior border; precoxal area coarsely vermiculate rugose anteriorly and medially, posteriorly absent; mesopleuron above precoxal area (except speculum) remotely punctate, shiny, and antero-dorsally coarsely vermiculate-rugose; metapleuron densely rugose, but dorsally punctate, interspaces approx. equal to diameter of punctures; propodeum evenly convex and coarsely rugose, medio-longitudinal carina complete, but irregular, no tubercles.
Wings.
Fore wing: r 0.35 × 3-SR (Fig.
Legs.
Tarsal claws subpectinate, with four brown medium-sized pectinal bristles and some finer ones basally (Fig.
Metasoma.
First tergite rather flattened; 1st and 2nd tergites coarsely and densely rugose, robust, with distinct median carina; medio-basal area of 2nd tergite wide and short (Fig.
Colour. Black; palpi, base of middle coxa, apical 0.2 (dorsally)–0.3 (inner side) of hind tibia, apex of hind femur and telotarsi blackish; remainder of tarsi more or less darkened and base of hind tibia dark brown; basal seven segments of antenna (remainder more or less dark brown), 1st and 2nd tergites and antero-lateral corners of 3rd tergite, and remainder of legs orange brown; humeral plate pale yellowish but partly darkened; pterostigma blackish; veins dark brown; wing membrane blackish infuscate.
Variation. Apical metasomal tergites of ♂ type 2; inner hind tibial spur 0.50 × hind basitarsus; mesopleuron, metapleuron and propodeum may be largely yellowish.
Distribution
Turkey.


Aleiodes turcicus sp. nov., ♀, holotype 779 fore wing 780 hind wing 781 mesosoma lateral 782 mesosoma dorsal 783 metasoma dorsal 784 fore femur lateral 785 hind femur lateral 786 head anterior 787 head dorsal 788 head lateral 789 base of antenna 790 apex of antenna 791 outer hind tarsal claw.
Aleiodes unipunctator
Ichneumon unipunctator Thunberg, 1822: 267 [examined].
Rogas unipunctator;
Rogas (Rogas) unipunctator;
Aleiodes (Neorhogas) unipunctator;
Aleiodes (Chelonorhogas) unipunctator;
Aleiodes unipunctator;
Ichneumon ductor
Thunberg, 1822: 269;
Aleiodes irregularis
Wesmael, 1838: 101;
Rhogas unipunctator ab. nigrescens
Hellén, 1927: 23;
Type material
Holotype of A. unipunctator, ♂ (
Additional material
Austria, Belgium, British Isles (England: V.C.s 1, 3, 5, 6, 8, 15, 16, 17, 18, 20, 21, 23, 25, 26, 27, 28, 29, 32, 33, 40, 53, 55, 58, 59, 63, 64, 65, 66, 69; Wales: V.C.s 35, 52 ; Scotland: V.C.s 72, 75, 76, 77, 79, 80, 82, 83, 85, 86, 87, 88, 89, 90, 91, 95, 96, 97, 98, 99, 100, 101, 103, 105, 108, 109, 110, 111, 112; Ireland: V.C.s H5, H19, H20, H21, H22, H28, H30), Bulgaria, Czech Republic, Denmark, Finland, Germany, Greece (mainland, Corfu), Hungary, Italy, Montenegro, Netherlands (DR: Wijster; Borger; GE: Nunspeet; Kemperberg, NB: Bergen op Zoom, ZH: Arkel; Melissant; Oostvoorne; ZE: Oostkapelle), Norway, Romania, Russia, Sweden, [Kazakhstan, Tadzhikistan, W. Caucasus]. Specimens in AAC, ALC,
Molecular data
MRS211 (UK), MRS221 (Germany), MRS354 (UK), MRS893 (UK).
Biology
Univoltine, flying from May to August. Reared from the noctuids Apamea unanimis (Hübner) (23 [1
Diagnosis
Maximum width of hypoclypeal depression 0.5–0.6 × minimum width of face (Fig.
Description
Redescribed ♀ (RMNH) from Sweden (Storbacken). Length of fore wing 5.8 mm, of body 5.7 mm.
Head.
Antennal segments of ♀ 50, length of antenna 1.2 × fore wing, its subapical segments rather robust; frons largely smooth anteriorly (except some fine sculpture) and densely rugulose posteriorly; OOL 0.8 × diameter of posterior ocellus, granulate with some punctures and matt; vertex granulate with some rugulosity and rather dull; clypeus coarsely punctate; ventral margin of clypeus thick and not protruding forwards (Fig.
Mesosoma. Mesoscutal lobes moderately punctate and interspaces distinctly granulate, with satin sheen; precoxal area of mesopleuron moderately rugose medially, sparsely punctulate posteriorly as surroundings of precoxal area; scutellum rather sparsely punctate, but medio-posteriorly rugulose, shiny; propodeum rather convex and moderately rugose, medio-longitudinal carina complete, and no protruding carinae laterally.
Wings.
Fore wing: r 0.3 × 3-SR (Fig.
Legs.
Tarsal claws yellowish bristly setose, without distinct pecten (Fig.
Metasoma.
First tergite rather flattened, 1.2 × as long as wide apically; 1st and 2nd tergites with distinct medio-longitudinal carina and longitudinally striate; medio-basal area of 2nd tergite wide triangular and short (Fig.
Colour. Black; pronotum dorso-posteriorly, telotarsi largely, hind tarsus, apical half of hind tibia, pterostigma (except paler extreme base), most veins and apical fifth of 2nd tergite dark brown; palpi brown; tegulae and basal half of hind tibia pale yellowish; apical third of 1st tergite and 2nd tergite (except apically) and remainder of legs, orange brown; wing membrane subhyaline.
Variation.
Propodeum and pronotum sometimes weakly marked with orange. One male seen with vein r-m of fore wing absent. Antennal segments: ♀ 47(1), 48(7), 49(12), 50(21), 51(26), 52(40), 53(40), 54(15), 55(5), 56(3), 57(1); ♂ 47(1), 48(1), 49(1), 50(8), 51(7), 52(22), 53(27), 54(34), 55(35), 56(21), 57(6), 58(8), 59(1). On average males have ca one to two more antennal segments than females. Male is very similar with apical tergites type 2, setae rather sparse and fringe short (Figs
Distribution
Austria, British Isles (England, Wales, Scotland, Ireland), Bulgaria, Czech Republic, *Denmark, Finland, Germany, *Greece (mainland and Corfu), Hungary, Italy, *Kazakhstan, *Montenegro, Netherlands, Norway, *Romania, Russia (including W. Caucasus), Sweden, *Tadzhikistan.
New synonymy
Both male holotypes of I. unipunctator and I. ductor are preserved in the Thunberg collection, but are severely damaged. From the holotype of A. ductor only the head, fore coxa, mesoscutum and metasoma remain (Roman, 1912). The holotype of A. unipunctator has the head and the metasoma separately glued on a card and the mesosoma is still attached to the pin. Judging from these remnants (especially the mainly smooth and very shiny 3rd tergite, the wide hypoclypeal depression (0.6 × minimum width of face) and the coriaceous vertex), it is obvious that both belong to the same species. Consequently, A. ductor (Thunberg) is synonymised with A. unipunctator (Thunberg) (syn. nov.). Aleiodes ductor auctt. is divided among A. pallidicornis (Herrich-Schäffer, 1838) (N and C European populations) and A. apicalis (Brullé, 1832) (Mediterranean and southern C European populations).


Aleiodes unipunctator (Thunberg), ♀, England, Fletcher Moss 795 wings 796 mesosoma lateral 797 mesosoma dorsal 798 metasoma dorsal 799 fore femur lateral 800 hind femur lateral 801 antenna 802 head anterior 803 head dorsal 804 head lateral 805 base of antenna 806 apex of antenna 807 inner hind tarsal claw lateral.

Aleiodes venustulus
Rhogas venustulus Kokujev, 1905: 15.
Rogas venustulus;
Aleiodes venustulus;
Rhogas (Rhogas) robustus Telenga, 1941: 151, 415.
Rogas robustus;
Type material
Holotype of A. venustulus, ♀ (
Additional material
1 ♀ + 1 ♂ (
Molecular data
None.
Biology
Unknown.
Diagnosis
Maximum width of hypoclypeal depression 0.6–0.7 × minimum width of face (Fig.
Description
Holotype of A. venustulus, ♀, length of fore wing 6.7 mm, of body 8.6 mm.
Head.
Antenna incomplete, with eight segments remaining; frons rugose and shiny; OOL twice diameter of posterior ocellus, mainly rugose and shiny; stemmaticum densely punctate; vertex remotely punctate and shiny; clypeus punctate and slightly convex; ventral margin of clypeus thick and anterior part weakly protruding (Fig.
Mesosoma. Mesoscutum coarsely and remotely punctate, with interspaces smooth and wider than punctures; precoxal area coarsely and densely punctate, remainder of mesopleuron remotely punctate and antero-dorsally rugose; metapleuron densely and coarsely punctate; metanotum with coarse medio-longitudinal carina anteriorly; scutellum punctate; propodeum convex and coarsely rugose, its medio-longitudinal carina present only on anterior third of propodeum.
Wings.
Fore wing: r 0.4 × 3-SR (Fig.
Legs.
Tarsal claws rather robust and with some fine dark brown spiny bristles subbasally (Fig.
Metasoma.
First tergite evenly convex, as long as wide apically; 1st and 2nd tergites with medio-longitudinal carina and coarsely rugose-reticulate; medio-basal area of 2nd tergite triangular and rather distinct (Fig.
Colour. Black; antenna (except scapus and pedicellus), palpi, tegulae, fore and middle telotarsi, veins, and pterostigma dark brown; coxae, trochanters and trochantelli, apical third of hind femur (ventrally extended to its apical two-thirds), hind tibia (except pale yellowish basal ring), fore and middle femora apically, and hind tarsus black; remainder of legs yellowish brown; wing membrane subhyaline.
Variation. No specimens with intact antennae examined; 2nd tergite coarsely rugose-reticulate or coarsely longitudinally rugose. Male is very similar and with apical tergites type 3, setae quite dense, glabrous stripe narrow, and fringe very weak.
Distribution
Kazakhstan, Kyrgyzstan.
Notes
This Central Asian species bears a superficial resemblance to A. miniatus and A. aestuosus. It is included in this revision, because it has been reported twice from Turkey (


Aleiodes venustulus (Kokujev), ♀, holotype 815 fore wing 816 hind wing 817 mesosoma lateral 818 mesosoma dorsal 819 metasoma dorsal 820 fore femur lateral 821 hind femur lateral 822 head anterior 823 head dorsal 824 head lateral 825 inner hind tarsal claw lateral 826 base of antenna.
Aleiodes zwakhalsi , sp. nov.
Type material
Holotype, ♀ (RMNH), “Turkey, Ankara, Kizilcahaman, 1100 m, 17.vi.1985, C.[J.] Zwakhals”. Paratypes: 1 ♂ (RMNH), “Turkey, Agri, Mt. Ararat, 1800 m, 3.vii.1985, C.J. Zwakhals”; 1 ♀ (RMNH), “Turkiye, Gümüshane, 40–46 km E [of] Bagburt, 1500 m, 19.vii.1989, J.A.W. Lucas”; 1 ♀ (
Molecular data
None.
Biology
Unknown. Probably univoltine as all known adults were collected in the period June to August, but there is no indication of its means of overwintering.
Diagnosis
Maximum width of hypoclypeal depression approx. 0.7 × minimum width of face (Fig.
This new species is similar to A. cruentus (Nees), but it differs by having the surroundings of veins M+CU1 and 1-+2-CU1 partly setose (Fig.
Description
Holotype, ♀, length of fore wing 7.0 mm, of body 9.0 mm.
Head.
Antennal segments of ♀ 59, length of antenna 1.1 × fore wing, its subapical segments moderately slender; frons largely smooth; OOL equal to diameter of posterior ocellus, finely remotely punctate and shiny; vertex distinctly punctate and shiny; clypeus punctate-rugose, wide and short; ventral margin of clypeus thick and not protruding forwards (Fig.
Mesosoma. Mesoscutal lobes largely densely and finely punctate, shiny; precoxal area of mesopleuron densely punctate, medially with few rugae; surroundings of precoxal area densely punctate; scutellum sparsely and finely punctate, rather flat, shiny and laterally rugose-punctate; propodeum evenly convex and coarsely rugose, medio-longitudinal carina complete but irregular posteriorly, and no protruding carinae laterally.
Wings.
Fore wing: r 0.4 × 3-SR (Fig.
Legs.
Tarsal claws with rather conspicuous pale brownish pecten (Fig.
Metasoma.
First tergite rather flattened, as long as wide apically and distinctly narrowed basally (Fig.
Colour. Black (including fore coxa anteriorly and basally); apex of hind tibia, telotarsi, hind tarsus, palpi, pterostigma and veins, dark brown; hind tibia (except apex) brownish yellow; clypeus narrowly ventrally, remainder of legs, pronotum (except ventrally), mesopleuron dorsally, mesoscutum, scutellum, and metanotum, orange-brown; tegulae pale brownish yellow; wing membrane subhyaline.
Variation. Antennal segments of ♀ 58(1), 59(1), 60(1), of ♂ 62(1); vein r of fore wing 0.4–0.5 × vein 3-SR; vein 1-CU1 of fore wing 0.3–0.6 × vein 2-CU1; OOL of ♀ 0.8–1.0 × diameter of posterior ocellus; length of eye 1.1–1.6 × temple in dorsal view; clypeus ventrally orange brown or black; mesopleuron dorsally or largely orange brown; basal half of third tergite rugose, punctate-rugose or punctulate; entire 1st tergite orange brown, posterior half of 1st tergite brownish and rest of tergite blackish (as base of middle coxa) or entirely black. Male is very similar, apical tergites type 1 and no fringe observed.
Distribution
Turkey (Asian part: 1100–1800 m).


Erratum for Part 1
In the key given by
Acknowledgements
We thank Sergey Belokobylskij and Konstantin Samartsev (
References
- Abdolalizadeh F, Madjdzadeh SM, Farahani S, Askari Hesni M (2017) A survey of braconid wasps (Hymenoptera: Braconidae: Euphorinae, Homolobinae, Macrocentrinae, Rogadinae) in Kerman province, southeastern Iran. Journal of Insect Biodiversity and Systematics 3(1): 33–40.
- Ashmead WH (1889) Descriptions of new Braconidae in the collection of the US National Museum. Proceedings of the US National Museum 11(1888): 611–671. https://doi.org/10.5479/si.00963801.11-760.611
- Aydogdu M, Beyarslan A (2005) The first records of Aleiodes Wesmael 1838 (Hymenoptera: Braconidae: Rogadinae). The fauna of Thrace region of Turkey. Linzer biologische Beiträge 37/1: 185–193.
- Aydogdu M, Beyarslan A (2006) First records of Aleiodes Wesmael, 1838 species in East Marmara region of Turkey (Hymenoptera: Braconidae: Rogadinae). Acta Entomologica Slovenica 14(1): 81–88.
- Baker CF (1917a) Ichneumonoid parasites of the Philippines. I Rhogadinae (Braconidae), I. Philippine Journal of Science (D) 12: 281–327.
- Baker CF (1917b) Ichneumonoid parasites of the Philippines. II. Rhogadinae (Braconidae), II: The genus Rhogas. Philippine Journal of Science (D) 12: 383–422.
- Belokobylskij SA (1996) Contribution to the knowledge of braconid fauna of the subfamily Rogadinae (Hymenoptera, Braconidae) of Russian Far East and Eastern Siberia. Part 1. Far Eastern Entomologist 27–28: 1–12.
- Belokobylskij SA (2000) Rogadinae: 15–109. In: Ler PA (Ed.) Key to the insects of Russian Far East, vol. 4. Neuropteroidea, Mecoptera, Hymenoptera, pt 4: 1–651. Vladivostok.
- Belokobylskij SA, Taeger A (2001) Braconidae: 103–115. In: Dathe HH, Taeger A, Blank SM (Eds) Verzeichnis der Hautflügler Deutschlands (Entomofauna Germanica 4). Entomologische Nachrichten und Berichte (Dresden), Beiheft 7: 1–178.
- Belokobylskij SA, Taeger A, van Achterberg C, Haeselbarth E, Riedel M (2003) Checklist of the Braconidae (Hymenoptera) of Germany. Beiträge zur Entomologie 53(2): 341–435.
- Belokobylskij SA, Zaldívar-Riverón A, Maetô K, Saez AG (2008) Asian Betylobraconinae (Hymenoptera, Braconidae), with description of a new genus and phylogenetic affinities of the tribe Facitorini. Insect Systematics and Evolution 39(2): 133–154. https://doi.org/10.1163/187631208788784110
- Belokobylskij SA, Kula RR (2012) Review of the brachypterous, micropterous, and apterous Braconidae of the cyclostome lineage (Hymenoptera: Ichneumonoidea) from the Palearctic Region. Zootaxa 3240: 1–62. https://doi.org/10.11646/zootaxa.3240.1.1
- Bergamasco P, Pennacchio F, Scaramozzino PL, Tremblay E (1995) Hymenoptera Braconidae. Checklist della Specie della Fauna Italiana 95: 2–29.
- Beyarslan A, Gözüaçik C, Güllü M, Konuksal A (2017) Taxonomical investigation on Braconidae (Hymenoptera: Ichneumonoidea) fauna in northern Cyprus, with twenty six new records for the country. Journal of Insect Biodiversity and Systematics 3(4): 319–334.
- Boheman CH (1853) Entomologiska Anteckningar under en resa i Södra Sverige 1851. Kongliga Svenska Vetenskaps-Akademiens Handlingar 1851: 53–210. https://doi.org/10.5962/bhl.title.35818
- Brues CT (1912) Brazilian Ichneumonidae and Braconidae obtained by the Stanford expedition. Annals of the Entomological Society of America 5: 193–228. https://doi.org/10.1093/aesa/5.3.193
- Brullé MA (1832) Expédition scientifique de Morée. Section des sciences physiques. Tome III. 1re partie. Zoologie. Levrault, Paris, 400 pp.
- Butcher BA, Quicke DLJ (2011) Revision of Aleiodes (Hemigyroneuron) parasitic wasps (Hymenoptera: Braconidae: Rogadinae) with reappraisal of subgeneric limits, descriptions of new species and phylogenetic analysis. Journal of Natural History 45: 1403–1476. https://doi.org/10.1080/00222933.2011.557557
- Butcher BA, Smith MA, Sharkey MJ, Quicke DLJ (2012) A turbo-taxonomic study of Thai Aleiodes (Aleiodes) and Aleiodes (Arcaleiodes) (Hymenoptera: Braconidae: Rogadinae) based largely on COI barcoded specimens, with rapid descriptions of 179 new species. Zootaxa 3457: 1–232.
- Butcher BA, Zaldívar-Riverón A, van de Kamp T, Rolo TDS, Baumbach T, Quicke DLJ (2014) Extension of historical range of Betylobraconinae (Hymenoptera: Braconidae) into Palaearctic Region based on a Baltic amber fossil, and description of a new species of Mesocentrus Szepligeti from Papua New Guinea. Zootaxa 3860(5): 449–463. https://doi.org/10.11646/zootaxa.3860.2
- Butcher BA, Quicke DLJ (2015) First record of Aleiodes (Hemigyroneuron) (Hymenoptera: Braconidae: Rogadinae) from the Arabian Peninsula: description of new species with remarkable wing venation convergence to Gyroneuron and Gyroneuronella. Zootaxa 4033(2): 275–279. https://doi.org/10.11646/zootaxa.4033.2.7
- Butcher BA, Quicke DLJ (2016) First Australian record of Aleiodes (Hemigyroneuron) (Hymenoptera: Braconidae: Rogadinae) with description of a new species from Tasmania. Journal of Asia-Pacific Entomology 19: 977–980. https://doi.org/10.1016/j.aspen.2016.08.014
- Čapek M, Lukás J (1989) Apocrita Parasitica, Ichneumoidea, Braconidae. Acta Faunistica Entomologica Musei Nationalis Pragae 19: 27–44.
- Chen X-X, He J-H (1992) New records of Aleiodes Wesmael from China (Hymenoptera: Braconidae: Rogadinae). Acta Zootaxonomica Sinica 17(1): 125. [in Chinese]
- Chen X-X, He J-H, Ma Y (1992) New records of Aleiodes Wesmael from China 2 (Hymenoptera: Braconidae: Rogadinae). Acta Zootaxonomica Sinica 17(4): 495–496. [in Chinese with English summary]
- Chen X-X, He J-H (1997) Revision of the subfamily Rogadinae (Hymenoptera: Braconidae) from China. Zoologische Verhandelingen, Leiden 308: 1–187.
- Chen X-X, Piao M-H, Whitfield JB, He J-H (2003) A molecular phylogeny of the subfamily Rogadinae (Hymenoptera: Braconidae) based on the D2 variable region of 28S ribosomal RNA. Acta Entomologica Sinica 46(2): 209–217.
- Costa A (1884) Rendiconto dell’Accademia della Scienze Fisiche e Metametiche, Napoli 21: 13.
- Curtis J (1834) British Entomology; being illustrations and descriptions of the genera of insects found in Great Britain and Ireland 11: 512.
- Dalla Torre CG de (1898) Catalogus Hymenopterorum. IV. Braconidae. Engelmann, Lipsiae, 1–323.
- Dandy JE (1969) Watsonian Vice-counties of Great Britain. Ray Society, London 146: 1–36. [+ 2 maps]
- Delfin GH, Wharton RA (2000) Historical review of the genera Aleiodes and Rogas in Mexico, with redescription of Aleiodes cameronii (Hymenoptera: Braconidae). Pan-Pacific Entomologist 76: 58–70.
- Eiseman C, Charney N (2010) Tracks & signs of insects and other invertebrates: a guide to North American species. Stackpole Books, Mechanicsburg, PA, 592 pp.
- Enderlein G (1912a) H. Sauter’s Formosa-Ausbeute. Braconidae, Proctotrupidae und Evaniidae (Hym.). Entomologische Mitteilungen 1: 257–267.
- Enderlein G (1912b) Neue Gattungen und Arten von Braconiden. Archiv für Naturgeschichte 78(A): 94–100. https://doi.org/10.5962/bhl.part.27904
- Enderlein G (1920) Zur Kenntnis aussereuropäischer Braconiden. Archiv für Naturgeschichte 84(A) 11(1918): 51–224. https://doi.org/10.5962/bhl.part.13627
- Fahringer J (1931) Opuscula braconologica. Band 3. Palaearktischen Region. Opuscula braconologica 3: 161–240. Wagner, Wien.
- Fahringer J (1932) Opuscula braconologica. Band 3. Palaearktischen Region. Opuscula braconologica 4: 241–320. Wagner, Wien.
- Fahringer J (1934) Opuscula braconologica. Band 3. Palaearktischen Region. Opuscula braconologica 5–8: 321–594. Wagner, Wien.
- Falcó JV, Moreno J, Jiménez R (1997) Datos sobre ciclostominos espanoles [Some data about the Spanish cyclostomes] 3. Rogadinae s. str. (Hymenoptera: Braconidae). Boletin de la Asociacion Española de Entomologia 21(1–2): 55–64.
- Farahani S, Talebi AA, van Achterberg C, Rakhshani E (2015) A review of the subfamily Rogadinae (Hymenoptera: Braconidae) from Iran. Zootaxa 3973(2): 227–250. https://doi.org/10.11646/zootaxa.3973.2.2
- Folmer O, Black M, Hoeh W, Lutz R, Vrijenhoek R (1994) DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Molecular Marine Biology and Biotechnology 3: 294–299.
- Fortier JC, Shaw SR (1999) Cladistics of the Aleiodes lineage of the subfamily Rogadinae (Hymenoptera: Braconidae). Journal of Hymenoptera Research 8(2): 204–237.
- Fortier J, Sherman C (2008) A new Aleiodes (= Tetrasphaeropyx) (= A. pilosus group), with a host record from Missouri, U.S.A., biological remarks, and previous type depository corrections. Entomological News 119(5): 445–453. https://doi.org/10.3157/0013-872X-119.5.445
- Garro LS, Shimbori EM, Penteado-Dias AM, Shaw SR (2017) Four new species of Aleiodes (Hymenoptera: Braconidae: Rogadinae) from the Neotropical Region. Canadian Entomologist 149: 560–573. https://doi.org/10.4039/tce.2017.23
- Ghahari H, Fischer M, Papp J (2011) A study on the Braconidae (Hymenoptera: Ichneumonoidea) from Ilam province, Iran. Calodema 160: 1–5.
- Giraud J (1857) Description de quelques hyménoptères nouveaux ou rares. Verhandlungen der Zoologisch-Botanischen Gesellschaft in Wien 7: 163–184.
- Hajibabaei M, deWaard JR, Ivanova NV, Ratnasingham S, Dooh RT, Kirk SL, Mackie PM, Hebert PDN (2005) Critical factors for assembling a high volume of DNA barcodes. Philosophical Transactions of the Royal Society B: Biological Sciences 360: 1959–1967. https://doi.org/10.1098/rstb.2005.1727
- He J-H, Chen X-X, Ma Y (2000) Hymenoptera Braconidae. Fauna Sinica. Insecta 18: 1–757. Science Press, Beijing.
- Hellén W (1927) Zur Kenntnis der Braconiden (Hym.) Finnlands. I. Subfam. Braconinae (part.), Rhogadinae und Spathiinae. Acta Societatis pro Fauna et Flora Fennica 56(12): 1–59.
- Hellén W (1957) Zur Kenntnis der Braconidae: Cyclostomi Finnlands. Notulae Entomologicae 37(2): 33–52.
- Herrich-Schäffer GAW (1838) Faunae Insectorum Germaniae initiae oder Deutschlands Insecten. Hymenoptera. Heft 124 & 156. Regensburg.
- Horn W, Kahle I (1935/37) Über entomologische Sammlungen, Entomologen & Entomo-Museologie, ein Beitrag zur Geschichte der Entomologie. Entomologische Beihefte aus Berlin-Dahlem 2–4: i–vi, 1–536.
- Huflejt T (1997) Ichneumonoidea: 75–114. In: Razowski J (Ed.) Wykaz zwierzat Polski [Checklist of animals of Poland] Hymenoptera – poslowie 5(32/24): 1–260.
- Ivanova NV, DeWaard JR, Hebert PDN (2006) An inexpensive, automation-friendly protocol for recovering high-quality DNA. Molecular Ecology Notes 6: 998–1002. https://doi.org/10.1111/j.1471-8286.2006.01428.x
- Jakimavicius AB (1974) Species of braconids new for the fauna of the Lithuanian SSR (Hym., Braconidae). Trudy Akademii Nauk Litovskoi SSR B 2(66): 93–102.
- Jurine L (1807) 1. Hyménoptères. Ichneumonoidea p. 1–51 & 94–118. Nouvelle méthode de classer les Hyménoptères et les Diptères. Paschoud, Geneva, 319 pp. https://doi.org/10.5962/bhl.title.10282
- Kokujev NR (1898) Fragments Braconologiques. VI. Sur quelques especes du genre Vipio. Trudy Russkago Entomologicheskago Obshchestva. [Horae Societatis Entomologicae Rossicae] 32: 291–317.
- Kokujev NR (1903) Hymenoptera asiatica nova. Russkoe Entomologicheskoye Obozreniye 3: 285–288.
- Kokujev NR (1905) Hymenoptera asiatica nova VI. Russkoe Entomologicheskoye Obozreniye 5: 10–15.
- Kolubajiv S (1962) Die Ergebnisse der Zuchten von Entomophagen (der Parasiten und Räuber) der schädlichen Insekten (vorwiegend der Forstschädlinge) in der Zeitperiode von 1934 bis 1958. Rozpravy Ceske Akademie 72: 1–73.
- Kotenko AG (1992) A contribution to the fauna of Braconidae (Hymenoptera) of Dauria. In: Amirkhanov AM (Ed.) Insects of Dauria and adjacent territories: 94–107. Izdatelstvo Tsentralnoi Nauchno issledovatelskoi laboratorii okhotnichego khozyaistva i zapovednikov, Moskva, 141 pp.
- Ku DS, Belokobylskij SA, Cha JY (2001) Hymenoptera (Braconidae). Economic Insects of Korea 16. Insecta Koreana, Suppl. 23: 1–283.
- Long KD, van Achterberg C (2008) One new genus and seven new species of Rogadinae (Hymenoptera: Braconidae) from Vietnam. Zoologische Mededelingen Leiden 82(24–41): 313–329.
- Lozan A, Belokobylskij S, van Achterberg C, Monaghan M (2010) Diversity and distribution of Braconidae, a family of parasitoid wasps in the Central European peatbogs of South Bohemia, Czech Republic Journal of Insect Science 10: 16(1–21). https://doi.org/10.1673/031.010.1601
- Lucas H (1849) Exploration scientifique de l’Algérie pendant les années 1840, 1841, 1842. Sciences physiques, Zoologie, III. Insectes: 1–527. Paris.
- Lyle GT (1919) Contributions to our knowledge of British Braconidae. No. 4 – Rhogadinae. Entomologist 49: 134–136, 149–155, 178–181.
- Ma Y, Chen X-X, He J-H (2002) Newly recorded species of genus Aleiodes Wesmael (Hymenoptera: Braconidae) in China. Entomotaxonomia 24(2): 98. [in Chinese]
- Marsh PM (1979) Braconidae. Aphidiidae. Hybrizontidae. In: Krombein KV, Hurd Jr PD, Smith DR, Burks BD (Eds) Catalog of Hymenoptera in America north of Mexico. Smithsonian Institution Press Washington, 144–313.
- Marsh PM, Shaw SR (2001) Revision of North American Aleiodes Wesmael (Part 6): The gasterator (Jurine) and unipunctator (Thunberg) species-groups (Hymenoptera: Braconidae: Rogadinae). Proceedings of the Entomological Society of Washington 103(2): 291–307.
- Marshall TA (1897) Les Braconides (Supplément): 1–144. In: André E (Ed.) 1897–1900. Species des Hyménoptères d’Europe et d’Algérie 5 bis, 1–369.
- Merz B, Pasche A (2012) 8.31.10. Superfamily Ichneumonoidea (Braconidae): 241–244. In: Merz B (Ed.) Liste annotée des insectes (Insecta) du canton de Genève. Instrumenta Biodiversitatis 8: 1–532. Genève.
- Morley C (1916) Notes on Braconidae. XI. The tribe Rhogadides, with additions to the British list. Entomologist 49: 83–87; 109–112; 125–130.
- Morley C (1937) The Hymenoptera of Suffolk, portio tertio et ultimo. Transactions of the Suffolk Naturalists’ Society 3: 233–248.
- Nees von Esenbeck CG (1811) Ichneumonides Adsciti, in Genera et Familias Divisi. Magazin Gesellschaft Naturforschender Freunde zu Berlin 5: 1–37.
- Nees von Esenbeck CG (1834) Hymenopterorum Ichneumonibus affinium monographiae, genera Europaea et species illustrantes 1: 1–320. Stuttgartiae et Tubingae. https://doi.org/10.5962/bhl.title.26555
- O’Connor JP, Nash R, van Achterberg C (1999) A catalogue of the Irish Braconidae (Hymenoptera: Ichneumonoidea). Occasional Publication of the Irish Biogeographical Society 4: 1–123.
- Papp J (1967) Ergebnisse der zoologischen Forschungen von Dr. Z. Kaszab in der Mongolei Braconidae (Hymenoptera). Acta Zoologica Academiae Scientiarum Hungaricae 13: 191–226.
- Papp J (1971) Ergebnisse der zoologischen Forschungen von Dr. Z. Kaszab in der Mongolei. 215. Braconidae (Hym.) 3. Annales Historico-Naturales Musei Nationalis Hungarici 63: 307–363.
- Papp J (1977a) Contributions to the Braconid fauna of Yugoslavia. III. Folia Entomologica Hungarica 30: 105–117.
- Papp J (1977b) Braconidae (Hymenoptera) from Mongolia, VI. Folia Entomologica Hungarica 30(1): 109–118.
- Papp J (1983) A survey of the Braconid fauna of the Hortobágy National Park (Hymenoptera, Braconidae), II: 315–337. In: Mahunka S (Ed.) The Fauna of the Hortobágy National Park II. Budapest.
- Papp J (1985a) Contributions to the Braconid fauna of Hungary, VII. Rogadinae (Hymenoptera: Braconidae). Folia Entomologica Hungarica 46: 143–164.
- Papp J (1985b) Braconidae (Hymenoptera) from Korea. VII. Acta Zoologica Hungarica 31: 341–365.
- Papp J (1987a) First survey of the braconid fauna of the Kiskunság National Park, Hungary (Hymenoptera, Braconidae). In: Mahunka S (Ed.) The Fauna of the Kiskunság National Park II. Akademiai Kiado, Budapest, 314–334.
- Papp J (1987b) First outline of the braconid fauna of Southern Transdanubia, Hungary (Hymenoptera, Braconidae), 3. Helconinae, Homolobinae, Macrocentrinae, Blacinae, Doryctinae and Rogadinae. A Janus Pannonius Muzeum Evkonyve 30–31 (1985–1986): 33–36.
- Papp J (1990) Braconidae (Hymenoptera) from Tunisia, 3. Folia Entomologica Hungarica 51: 89–96.
- Papp J (1991a) Parasitic wasps of the superfamily Ichneumonoidea. Braconidae. 18. Doryctinae. 19. Rogadinae. Magyarorszag Allatvilaga [Fauna Hungariae] 167: 1–122.
- Papp J (1991b) Zur Insektenfauna von Gersau-Oberholz, Kanton Schwyz. X. Hymenoptera 1: Braconidae (Brackwespen). Entomologische Berichte, Luzern 25: 1–26.
- Papp J (1991c) Second survey of the braconid wasps in the Bátorliget Nature Conservation Areas, Hungary (Hymenoptera: Braconidae). In: Mahunka S (Ed.) The Bátorliget Nature Reserves after forty years. Magyar Természettudományi Múzeum, Budapest. Studia Naturalia. 1(2) (1990), 639–674.
- Papp J (1994) The dispersion of braconid wasps in an oak forest of Hungary (Hymenoptera: Braconidae). Folia Entomologica Hungarica 55: 305–320.
- Papp J, Rezbanyai-Reser L (1996) Contributions to the braconid fauna of Monte Generoso, Canton Ticino, southern Switzerland (Hymenoptera: Braconidae). Entomologische Berichte Luzern 35: 59–134.
- Papp J (1999) The Braconid wasps (Hymenoptera: Braconidae) of the Aggtelek National Park (N.E. Hungary). In: Mahunka S (Ed.) The Fauna of the Aggtelek National Park. Hungarian Natural History Museum, Budapest, 547–572.
- Papp J (2002) The Braconid wasps (Hymenoptera: Braconidae) of the Ferto-Hansag National Park (NW Hungary). In: Mahunka S (Ed.) The Fauna of the Ferto-Hansag National Park, Hungarian Natural History Museum, Budapest, 557–581.
- Papp J (2003) Braconid wasps from the Cape Verde Islands (Insecta: Hymenoptera: Braconidae) 2. Doryctinae, Braconinae, Hormiinae, Rogadinae, Gnamptodontinae, Homolobinae, Opiinae, Alysiinae, Cheloninae, Adeliinae and Microgastrinae. Faunistische Abhandlungen (Dresden) 24: 137–167.
- Papp J (2004) Type specimens of the braconid species by Gy. Szépligeti deposited in the Hungarian Natural History Museum (Hymenoptera: Braconidae). Annales Historico-Naturales Musei Nationalis Hungarici 96: 153–223.
- Papp J (2005) A checklist of the Braconidae of Hungary (Hymenoptera). Folia Entomologica Hungarica. 66: 137–194.
- Papp J (2009) Braconidae (Hymenoptera) from Mongolia, XVII. Eleven subfamilies. Acta Zoologica Academiae Scientiarum Hungaricae 55(2): 139–173.
- Papp J (2012) A contribution to the Braconid fauna of Israel (Hymenoptera: Braconidae), 3. Israel Journal of Entomology 41–42: 165–219.
- Papp J, Vas Z (2016) Contributions to the knowledge of Braconidae fauna of Sălaj County, Romania (Hymenoptera). Studia Universitatis “Vasile Goldiş”, Seria Ştiinţele Vieţii 26(1): 141–152.
- Perkins JF (1953) Note on British Ichneumonidae with descriptions of new species (Hym., Ichneumonidae). Bulletin of the British Museum (Natural History) (Entomology) 3: 103–176. https://doi.org/10.5962/bhl.part.1050
- Quicke DLJ, Shaw MR (2005) First host record for the rogadine genus Pholichora van Achterberg (Hymenoptera: Braconidae) with description of a new species and notes on convergent wing venation features. Journal of Natural History 39(7): 531–537. https://doi.org/10.1080/00222930410001708678
- Quicke DLJ, Shaw MR, van Achterberg C, Bland KP, Butcher BA, Lyszkowski R, Zhang YM (2014) A new Australian genus and five new species of Rogadinae (Hymenoptera: Braconidae), one reared as a gregarious endoparasitoid of an unidentified limacodid (Lepidoptera). Zootaxa 3881(3): 237–257. https://doi.org/10.11646/zootaxa.3881.3.3
- Rambaut A (2016) FigTree v.1.4.3. Available at: http://tree.bio.ed.ac.uk/software/figtree/
- Rastegar J, Sakenin H, Khodaparast S, Havaskary M (2012) On a collection of Braconidae (Hymenoptera) from East Azarbaijan and vicinity, Iran. Calodema 226: 1–4.
- Ratzeburg JTC (1852) Die Ichneumonen der Forstinsecten in forstlicher und entomologischer Beziehung. Dritter Band: 1–272. Berlin.
- Reinhard H (1863) Beiträge zur Kenntniss einiger Braconiden-Gattungen. Berliner Entomologische Zeitschrift 7: 248–274. https://doi.org/10.1002/mmnd.47918630304
- Riedel M, Hansen LO, Berg Ø (2002) Braconidae (Hymenoptera) of Norway, Part 1. Norwegian Journal of Entomology 49: 97–108.
- Samartsev KG, Belokobylskij SA (2013) On the fauna of the true cyclostome braconid wasps (Hymenoptera, Braconidae) of Astrakhan Province. Entomologicheskoe Obozrenie 92(2): 319–341. Entomological Review 93(6): 755–774.
- Shaw MR (1979) Rogas pulchripes (Wesmael) (Hymenoptera: Braconidae) and other parasites of arboreal Acronicta species (Lepidoptera: Noctuidae) at Chat Moss, Manchester. Entomologist’s Gazette 30: 291–294.
- Shaw MR (1981) Possible foodplant differences of Amphipyra pyramidea (L.) and A. berbera svenssoni Fletcher (Lepidoptera: Noctuidae), and a note on their parasitoids (Hymenoptera). Entomologist’s Gazette 32: 165–167.
- Shaw MR (1983) On[e] evolution of endoparasitism; the biology of some genera of Rogadinae (Braconidae). Contributions of the American Entomological Institute 20: 307–328.
- Shaw MR, Huddleston T (1991) Classification and biology of Braconid wasps (Hymenoptera: Braconidae). Handbooks for the Identification of British Insects 7(11): 1–126.
- Shaw MR (1994) Parasitoid host ranges. In: Hawkins BA, Sheehan W (Eds) Parasitoid community ecology. Oxford University Press, Oxford, 111–144.
- Shaw MR (1997) Rearing parasitic Hymenoptera. The Amateur Entomologist 25: 1–46. Amateur Entomologist’s Society, Orpington.
- Shaw MR (2003) Host ranges of Aleiodes species (Hymenoptera: Braconidae), and an evolutionary hypothesis. In: Melika G, Thuroczy C (Eds) Parasitic wasps: evolution, systematics, biodiversity and biological control (2002). Agroinform, Budapest, 321–327.
- Shaw MR (2017) Anatomy, reach and classification of the parasitoid complex of a common British moth, Anthophila fabriciana (L.) (Choreutidae). Journal of Natural History 51(19–20): 1119–1149. https://doi.org/10.1080/00222933.2017.1315837
- Shaw SR (1993) Systematic status of Eucystomastax Brues and characterization of the Neotropical species (Hymenoptera: Braconidae, Rogadinae). Journal of Hymenoptera Research 2: 1–11.
- Shaw SR, Marsh PM, Fortier JC (1997) Revision of North American Aleiodes (part 1): the pulchripes Wesmael species-group in the New World (Hymenoptera: Braconidae: Rogadinae). Journal of Hymenoptera Research 6: 10–35.
- Shaw SR, Marsh PM, Fortier JC (1998) Revision of North American Aleiodes Wesmael (Part 2): the apicalis (Brullé) species-group in the New World (Hymenoptera: Braconidae, Rogadinae) Journal of Hymenoptera Research 7: 62–73.
- Shenefelt RD (1975) Braconidae 8. Exothecinae, Rogadinae. Hymenopterorum Catalogus (nova editio) 12: 1115–1262.
- Shenefelt RD (1978) Braconidae 10. Braconinae, Gnathobraconinae, Mesostoinae, Pseudodicrogeniinae, Telengainae, Ypsistocerinae, plus Braconidae in general, major groups, unplaced genera and species. Hymenopterorum Catalogus (nova editio) 15: 1425–1872.
- Shestakov A (1940) Zur Kenntnis der Braconiden Ostsibiriens. Arkiv för Zoologi 32A (19): 1–21.
- Shimbori EM, Penteado-Dias AM (2011) Taxonomic contribution to the Aleiodes melanopterus (Erichson) species-group (Hymenoptera, Braconidae, Rogadinae) from Brazil. ZooKeys 142: 15–25. https://doi.org/10.3897/zookeys.142.1705
- Smith MA, Fisher BL, Hebert PDN (2005) DNA barcoding for effective biodiversity assessment of a hyperdiverse arthropod group: the ants of Madagascar. Philosophical Transactions of the Royal Society B Biological Sciences 360(1462): 1825–1834. https://doi.org/10.1098/rstb.2005.1714
- Snellen van Vollenhoven SC (1858) Naamlijst van Nederlandsche Vliesvleugelige insekten (Hymenoptera): 221–263. In: Herklots JA (Ed.) Bouwstoffen voor eene Fauna van Nederland. Tweede Deel. Brill, Leiden, 316 pp.
- Spinola M (1808) Insectorum Liguriae species novae aut rariores, quas in agro Ligustico nuper detexit, descripsit, et iconibus illustravit (Hymenoptera) 2: 1–262. Genua.
- Stamatakis A (2014) RAxML Version 8: a tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 30(9): 1312–1313. https://doi.org/10.1093/bioinformatics/btu033
- Stelfox AW (1953) A new Rogas (Hym., Braconidae) from Ireland. Entomologist’s Monthly Magazine 89: 146–147.
- Szépligeti G (1903) A palaeartikus Bracon-felek rendszere. 8 es 9. alcsalad: Rhogas es Rhyssalus-felek (Rhogadinae, Rhyssaloinae). Termeszet Tud. Közl. Állattani Közlem 2: 105–114.
- Szépligeti G (1906) Braconiden aus der Sammlung des ungarischen National-Museums, 1. Annales Historico-Naturales Musei Nationalis Hungarici 4: 547–618.
- Telenga NA (1941) Family Braconidae, subfamily Braconinae (continuation) and Sigalphinae. Fauna USSR. Hymenoptera 5(3): 1–466.
- Telenga NA (1959) Materials on the fauna of Braconidae and Ichneumonidae in Armenia. Izvestiya Akademii Nauk Armenski SSR 12(6): 85–88.
- Thomson CG (1892) XLIV. Bidrag till Braconidernas kannedom. Opuscula Entomologica 16: 1659–1751.
- Thunberg CP (1822) Ichneumonidea, Insecta Hymenoptera illustrata. Mémoires de l’Académie Imperiale des Sciences de Saint Petersbourg 8: 249–281.
- Tobias VI (1971) Review of the Braconidae (Hymenoptera) of the U.S.S.R. Trudy Vsesoyuznogo Entomologicheskogo Obshchestva 54: 156–268. [in Russian] [Translation (1975). 164 pp, New Delhi, India.]
- Tobias VI (1976) Braconids of the Caucasus (Hymenoptera, Braconidae). Opredeliteli po Faune SSSR 110: 1–286. [in Russian]
- Tobias VI (1986) Rogadinae: 72–85. In: Medvedev GS (Ed.) Opredelitel Nasekomych Evrospeiskoi Tsasti SSSR 3, Peredpontdatokrylye 4. Opredeliteli po Faune SSSR 145: 1–501. [Translation (1995): 512–588. Lebanon, U.S.A.]
- van Achterberg C (1988) Revision of the subfamily Blacinae Foerster (Hymenoptera, Braconidae), Zoolologische Verhandelingen, Leiden 249: 1–324.
- van Achterberg C (1990) Illustrated key to the subfamilies of the Holarctic Braconidae (Hymenoptera: Ichneumonoidea). Zoologische Mededelingen, Leiden 64: 1–20.
- van Achterberg C (1991) Revision of the genera of the Afrotropical and W. Palaearctic Rogadinae Foerster (Hymenoptera: Braconidae). Zoolologische Verhandelingen, Leiden 273: 1–102.
- van Achterberg C (1992) Revisionary notes on the subfamily Homolobinae (Hymenoptera: Braconidae). Zoologische Mededelingen, Leiden 66(25): 359–368.
- van Achterberg C (1993) Illustrated key to the subfamilies of the Braconidae (Hymenoptera: Ichneumonoidea). Zoolologische Verhandelingen, Leiden 283: 1–189.
- van Achterberg C (1997) Revision of the Haliday collection of Braconidae (Hymenoptera). Zoologische Verhandelingen, Leiden 314: 1–115.
- van Achterberg C (2014) Notes on the checklist of Braconidae (Hymenoptera) from Switzerland. Mitteilungen der Schweizerischen Entomologischen Gesellschaft 87: 191–213.
- van Achterberg C, Shaw MR (2016) Revision of the western Palaearctic species of Aleiodes Wesmael (Hymenoptera, Braconidae, Rogadinae). Part 1: Introduction, key to species groups, outlying distinctive species, and revisionary notes on some further species. ZooKeys 639: 1–164.
- Walker F (1860) Characters of some apparently undescribed Ceylon insects. Annals and Magazine of Natural History (3)5: 304–311. https://doi.org/10.1080/00222936008697221
- Wesmael C (1838) Monographie des Braconides de Belgique 4. Nouveaux Mémoires de l’Academie Royale des Sciences et Belles-lettres de Bruxelles 11: 1–166.
- Wollaston TV (1858) Brief diagnostic characters of undescribed Madeiran insects. Annals and Magazine of Natural History (3)1: 18–28. https://doi.org/10.1080/00222935808696865
- Yu DSK, van Achterberg C, Horstmann K (2016) Taxapad 2016, Ichneumonoidea 2015. Database on flash-drive. Ottawa, Ontario, Canada. www.taxapad.com
- Yuldashev EJ (2006) Braconid-wasps (Hymenoptera, Braconidae) in Uzbekistan. Uzbekskii Biologicheskii Zhurnal (2005) 5: 41–47. [in Russian with English summary]
- Zaldívar-Riverón A, Butcher BA, Shaw MR, Quicke DLJ (2004) Comparative morphology of the venom apparatus in the braconid wasp subfamily Rogadinae (Insecta, Hymenoptera, Braconidae) and related taxa. Zoologica Scripta 33(3): 223–237. https://doi.org/10.1111/j.0300-3256.2004.00144.x
- Zaldívar-Riverón A, Shaw MR, Sáez AG, Mori M, Belokobylskij SA, Shaw SR, Quicke DLJ (2008) Evolution of the parasitic wasp subfamily Rogadinae (Braconidae): phylogeny and evolution of lepidopteran host ranges and mummy characteristics. BMC Evolutionary Biology 8: 329. https://doi.org/10.1186/1471-2148-8-329
- Zaykov AN (1980a) Unknown genera and species of Braconidae for the fauna of Bulgaria. Travaux scientifiques d’Université de Plovdiv “Paissi Hilendarski”, Biologie 15(4) [1977]: 111–115. [in Bulgarian]
- Zaykov AN (1980b) New species of the family Braconidae (Hymenoptera) for the Bulgarian fauna. Acta Zoologica Bulgarica 16: 86–89. [in Bulgarian with English summary]
- Zaykov AN (1980c) Contribution to the braconid fauna of Bulgaria (Hymenoptera, Braconidae). Travaux scientifiques d’Université de Plovdiv “Paissi Hilendarski”, Biologie 16(4) [1978]: 227–234. [in Bulgarian with English summary]
- Žikić V, Brajković M, Tomanović Z (2002) Preliminary results of Braconid fauna research (Hymenoptera: Braconidae) found in Sicevo Gorge, Serbia. Acta Entomologica Serbica 5 (1–2) (2000): 95–110.
Appendix 1
| Genus | Species or species code | Voucher code/ Sample ID | Country | BOLD Process ID (if applicable) | GenBank accession number (s) |
|---|---|---|---|---|---|
| Aleiodes | abraxanae | MRS636 | UK | ASQSP978-10 | HQ551278/HQ551264 |
| adorabelleae | BCLDQ0252 | Thailand | ASQSP222-08 | JF963436 | |
| aestuosus | MRS004 | Turkey | GBAH4104-09 | EU979573 | |
| aff. wyomingensis | BIOUG01036-F12 | USA | JSHYN023-11 | KR791670 | |
| albitibia | MRS383 | Sweden | ASQSP075-08 | JF962835 | |
| alternator | MRS161 | UK | ASQBR258-09 | MK585882 | |
| angustipterus | MRS276 | UK | – | KU682232 | |
| antescutum | BCLDQ00210 | Thailand | ASQSP462-08 | JF962536 | |
| apicalis | MRS008 | Turkey | ASQBR099-09 | EU979575 | |
| MRS111 | Turkey | ASQBR099-09 | JF962839 | ||
| MRS112 | Turkey | ASQBR100-09 | MK585857 | ||
| MRS181 | Russia | ASQBR101-09 | MK585870 | ||
| MRS869 | Sweden | BCNCA217-18 | MK585872 | ||
| apiculatus | MRS079 | UK | – | KU682222 | |
| assimilis | MRS843 | France | BCNCA194-18 | MK585863 | |
| aterrimus | MRS024 | UK | ASQBR103-09 | JN000875 | |
| MRS147 | UK | GBAH4101-09 | EU979577 | ||
| bicolor | MRS197 | UK | ASQBR086-09 | MK585862 | |
| bobwhartoni | BCLDQ0730 | Thailand | ASQSQ011-09 | JF271188 | |
| bucculentus | BCLDQ00454 | USA | ASQSP644-08 | JF962486 | |
| buoculus | BCLDQ00927 | USA | ASQSQ223-09 | MH272394 | |
| buzuritriplus | BCLDQ01479 | Thailand | ASQSR027-11 | JN278254 | |
| cameronii Janz01 | DHJPAR0021064 | Costa Rica | – | JF792897 | |
| cantherius | MRS777 | Sweden | GBMIN74555-17 | KU682249 | |
| carbonarius | MRS162 | Hungary | ASQBR120-09 | MK585853 | |
| MRS163 | Hungary | ASQAS220-11 | JF962848 | ||
| MRS164 | Hungary | ASQBR121-09 | MK585851 | ||
| carminatus | MRS055 | France | ASQBR152-09 | JF962818 | |
| castaneus | BCLDQ0245 | Thailand | ASQSP215-08 | JQ388461 | |
| caudalis | MRS693 | France | ASQSP1001-10 | HQ551216 | |
| circumscriptus | MRS073 | UK | – | KU682256 | |
| compressor | MRS170 | UK | GBAH4098-09 | EU979580 | |
| concoronarius | BCLDQ01515 | Thailand | ASQSR079-11 | JN278271 | |
| coriaceus | MRS311 | Sweden | ASQBR140-09 | JF962853 | |
| MRS377 | Sweden | ASQBR141-09 | MK585885 | ||
| coronopus | BCLDQ00764 | Thailand | ASQSQ045-09 | JQ388389 | |
| corrusciput | BCLDQ01555 | Thailand | ASQSR119-11 | JN278306 | |
| coxalis | MRS606 | UK | ASQSP553-08 | MK585874 | |
| cruentus | MRS558 | France | ASQSP119-08 | MK585876 | |
| MRS624 | Germany | ASQSP989-10 | HQ551274 | ||
| MRS625 | Germany | ASQSP977-10 | HQ551263 | ||
| curticornis | MRS343 | Italy | ASQBR105-09 | JF962826/ KU682237 | |
| damus | BCLDQ00126 | Thailand | ASQSP378-08 | JQ388354 | |
| diarsianae | MRS030 | UK | ASQSP006-08 | JF962600 | |
| dissector | MRS007 | Turkey | ASQBR115-09 | JF957045 | |
| MRS025 | Turkey | ASQBR116-09 | MK585881 | ||
| MRS145 | UK | ASQBR117-09 | MK585849 | ||
| Aleiodes | dissector | MRS146 | UK | GBAH2876-07 | EF115471 |
| esenbeckii | MRS500 | Spain | ASQBR092-09 | JF962845/ KU682240 | |
| esenbeckii f. dendrolimi | MRS180 | Finland | GBAH4097-09 | EU979581 | |
| flavostriatus | BCLDQ0628 | Thailand | ASQSP849-08 | JF962636 | |
| fortipes | MRS650 | France | ASQSP966-10 | HQ551254 | |
| MRS807 | Poland | ASQBR957-18 | MH272207 | ||
| gasterator | MRS046 | France | ASQAS218-11 | EU979582 | |
| MRS048 | France | ASQBR118-09 | MK585879 | ||
| MRS892/ BF002924 | Spain | ASQBR119-09 | JF962819 | ||
| gastritor agg spG1 | MRS225 | UK | ASQBR273-09 | MK585878 | |
| gastritor agg spG2 | MRS017 | UK | ASQBR285-09 | MK585867 | |
| gastritor agg spG3 | MRS351 | UK | ASQSP101-08 | MK585847 | |
| gastritor agg spG4 | MRS153 | UK | ASQBR296-09 | MK585887 | |
| gastritor agg spG5 | MRS208/AL422 | UK | ASQBR959-19 | MK585859 | |
| grassator | MRS215 | UK | ASQBR122-09 | JF957046 | |
| MRS721 | UK | ASQBR960-19 | MK585855 | ||
| MRS725 | UK | ASQBR961-19 | MK585886 | ||
| hirtus | MRS619 | UK | ASQSP1000-10 | HQ551215 | |
| MRS882/ BCLDQ0505 | Romania | ASQSP695-08 | MK585888 | ||
| hirtus | MRS883 | Romania | ASQSP694-08 | MK585850 | |
| jakowlewi | MRS355 | Finland | ASQBR123-09 | JF962849 | |
| leptofemur | MRS156 | UK | ASQBR196-09 | JF962813 | |
| melanopterus | BCLDQ0799 | Brazil | ASQSQ080-09 | MH272348 | |
| mellificus | AL0037 | Thailand | ASQBR476-09 | JF962707 | |
| mexicanus | BMNHE897778 | Belize | ASQSQ642-10 | HQ551341 | |
| miniatus | MRS950 | Sweden | ALEIO030-19 | MN968689 | |
| MRS951 | Sweden | ALEIO031-19 | MN968690 | ||
| modestus | MRS282 | UK | ASQBR124-09 | JF962850 | |
| mubfsi | AL0323 | Uganda | GBAH2900-07 | EF115447 | |
| nigriceps | MRS613 | UK | ASQSP769-08 | KU682243 | |
| nigricornis | MRS216 | UK | GBAH4093-09 | EU979585 | |
| nobilis | MRS401 | Finland | ASQBR093-09 | MK585856 | |
| MRS880/ BCLDQ0277 | Russia | ASQSP247-08 | MK585871 | ||
| MRS881/ BCLDQ00123 | UK | ASQSP375-08 | JF962562 | ||
| nunbergi | MRS723 | Austria | ASQBR930-18 | MH272254 | |
| pallidator | MRS001 | Turkey | GBAH4092-09 | EU979586 | |
| pallidicornis | MRS885 | Russia | ASQSP742-08 | JF957043 | |
| pappi | CCDB-27844-E04 | Kenya | BBTH766-17 | MH272335 | |
| paulmarshi | CK0002 | Thailand | ASQSP942-10 | HQ551236 | |
| pictus | MRS556 | Austria | GBMIN74565-17 | KU682242 | |
| praetor | MRS654 | Bulgaria | ASQSP979-10 | HQ551265/KU682244 | |
| probuzurae | BCLDQ1202 | Thailand | ASQSQ410-09 | HM435162 | |
| pulchripes | MRS847 | Sweden | BCNCA197-18 | MK585848 | |
| MRS873/ BC-ZSM-HYM-27497-A05 | Sweden | BCHYM15694-17 | MK585880 | ||
| punctipes | MRS212 | UK | GBAH4091-09 | EU979587 | |
| quadrum | MRS824 | Bulgaria | AAHYM658-16 | MK585865 | |
| MRS796 | France | ASQBR939-18 | MH272232 | ||
| Aleiodes | reticulatus | MRS808 | Poland | GBAHB1497-18 | KU682262 |
| risaae | BCLDQ1268 | Thailand | ASQSQ476-09 | HM435190 | |
| rivulus | BCLDQ01646 | Thailand | ASQSR210-11 | JN278372 | |
| ruficornis | MRS140 | UK | ASQBR132-09 | MK585892 | |
| MRS877 | Sweden | BCNCA221-18 | MK585860 | ||
| MRS887/ AL0149 | UK | ASQBR131-09 | MK585846 | ||
| MRS888 | UK | GBAH2871-07 | EF115476 | ||
| MRS889 | ? | ASQBR127-09 | EF115475 | ||
| MRS890 | UK | ASQBR962-19 | MK585883 | ||
| MRS891 | UK | GBAH5262-09 | EF115477 | ||
| nr ruficornis | MRS886 | Hungary | ASQSP806-08 | JF962480 | |
| rufipes | MRS294 | Sweden | ASQBR134-09 | MK585884 | |
| MRS312 | Sweden | ASQSP069-08 | MH272380 | ||
| MRS314 | Sweden | ASQBR135-09 | MK585873 | ||
| MRS673/ SwedFin2 | Finland | ASQBR137-09 | MK585864 | ||
| MRS674/ SwedFin5 | Finland | ASQBR138-09 | MK585854 | ||
| MRS676/ SwedFin17 | Finland | ASQBR136-09 | JF962855 | ||
| MRS680/ SwedFin28 | Finland | ASQBR139-09 | JF962854 | ||
| rugulosus | MRS191 | Hungary | ASQBR142-09 | JF962857 | |
| MRS217 | UK | ASQBR143-09 | JF962856 | ||
| MRS398 | France | ASQSP011-08 | MH272286 | ||
| MRS884/ BCLDQ0509 | Poland | ASQSP699-08 | MK585875 | ||
| CollHH1599 | Norway | COLHH1790-18 | MK585877 | ||
| ryrholmi | MRS395 | Sweden | ASQBR322-09 | JF962792 | |
| seriatus agg | MRS616 | UK | ASQSP754-08 | MH272311 | |
| sibiricus | MRS310 | Sweden | ASQBR157-09 | JF962862 | |
| MRS313 | Sweden | ASQBR158-09 | MH272159 | ||
| MRS805 | France | BCNCA185-18 | MK585861 | ||
| signatus | MRS844 | Austria | BCNCA195-18 | MK585889 | |
| similis | MRS696 | Austria | ASQSQ727-10 | HQ551413 | |
| sophieae | BCLDQ01065 | Thailand | ASQSQ362-09 | JQ388368 | |
| sp M3 | MRS703/ MRSA 703 | Hungary | ASQSR240-11 | MK585852 | |
| spurivena | BCLDQ00003 | Vietnam | – | KY621612 | |
| terminalis | BCLDQ0692 | USA | ASQSP913-08 | JF962663 | |
| testaceus | MRS072 | UK | ASQBR963-19 | MK585866 | |
| trevelyanae | AL0226 | Uganda | – | EF115433 | |
| trianguliscleroma | CCDB27844-E03 | Malawi | BBTH765-17 | MH272236 | |
| trisphaeropyx | BCLDQ01643 | Thailand | ASQSR207-11 | JQ388329 | |
| turcicus | MRS126 | Turkey | ASQAS219-11 | JF962613 | |
| ungularis | MRS604 | France | ASQSP757-08 | JF962867 | |
| unipunctator | MRS211 | UK | ASQAS221-11 | JF962868 | |
| MRS221 | Germany | ASQBR171-09 | MK585858 | ||
| MRS354 | UK | ASQSP035-08 | MK585890 | ||
| MRS893 | UK | BCNCA223-18 | MK585891 | ||
| CollHH1603 | Norway | COLHH1794-18 | MK585893 | ||
| CollHH1604 | Norway | COLHH1795-18 | MK585868 | ||
| CollHH1605 | Norway | COLHH1796-18 | MK585869 | ||
| valinus | BCLDQ0267 | Thailand | ASQSP237-08 | JF963430 | |
| varius | MRS446 | Russia | – | HQ551275 | |
| Heterogamus | dispar | MRS066 | UK | ASQBR042-09 | JF963405 |
| excavatus | MRS717 | Sweden | ASQBR935-18 | MH272379 | |
| fasciatipennis | MRS669 | Sweden | ASQBR044-09 | MH272347 |