





Citation: Bu Y, Potapov MB, Yin WY (2014) Systematic and biogeographical study of Protura (Hexapoda) in Russian Far East: new data on high endemism of the group. ZooKeys 424: 19–57. doi: 10.3897/zookeys.424.7388
Proturan collections from Magadan Oblast, Khabarovsk Krai, Primorsky Krai, and Sakhalin Oblast are reported here. Twenty-five species are found of which 13 species are new records for Russian Far East which enrich the knowledge of Protura known for this area. Three new species Baculentulus krabbensis sp. n., Fjellbergella lazovskiensis sp. n. and Yichunentulus alpatovi sp. n. are illustrated and described. The new materials of Imadateiella sharovi (Martynova, 1977) are studied and described in details. Two new combinations, Yichunentulus borealis (Nakamura, 2004), comb. n. and Fjellbergella jilinensis (Wu & Yin, 2007), comb. n. are proposed as a result of morphological examination. Keys to species of the genera Fjellbergella and Yichunentulus are given. An annotated list of all species of Protura from Russian Far East is provided and discussed. Widely distributed species were not recorded in this area. This may be because of the high sensitivity of Protura to anthropogenic impact and low dispersal ability of the group.
Key, checklist, Baculentulus, Fjellbergella, Yichunentulus, biogeography
The Protura are minute soil-dwelling arthropods with more than 800 species known so far in the world (
The Russian Far East (abbreviated as RFE throughout the present paper) occupies an area of 3, 016 thousand sq. km. and extends from Wrangel Island southwards to Khasan Lake (Fig. 1). Forests occupy 39% of the territory and predominate in Primorsky Krai, Amur Oblast, Sakhalin Oblast, and Khabarovsk Lrai. The insect fauna is associated with mixed broadleaved-coniferous forests in the south of RFE. About 31, 500 species of insects have been recorded in the RFE so far where Eastern-Asiatic and local species make the most part of the fauna (
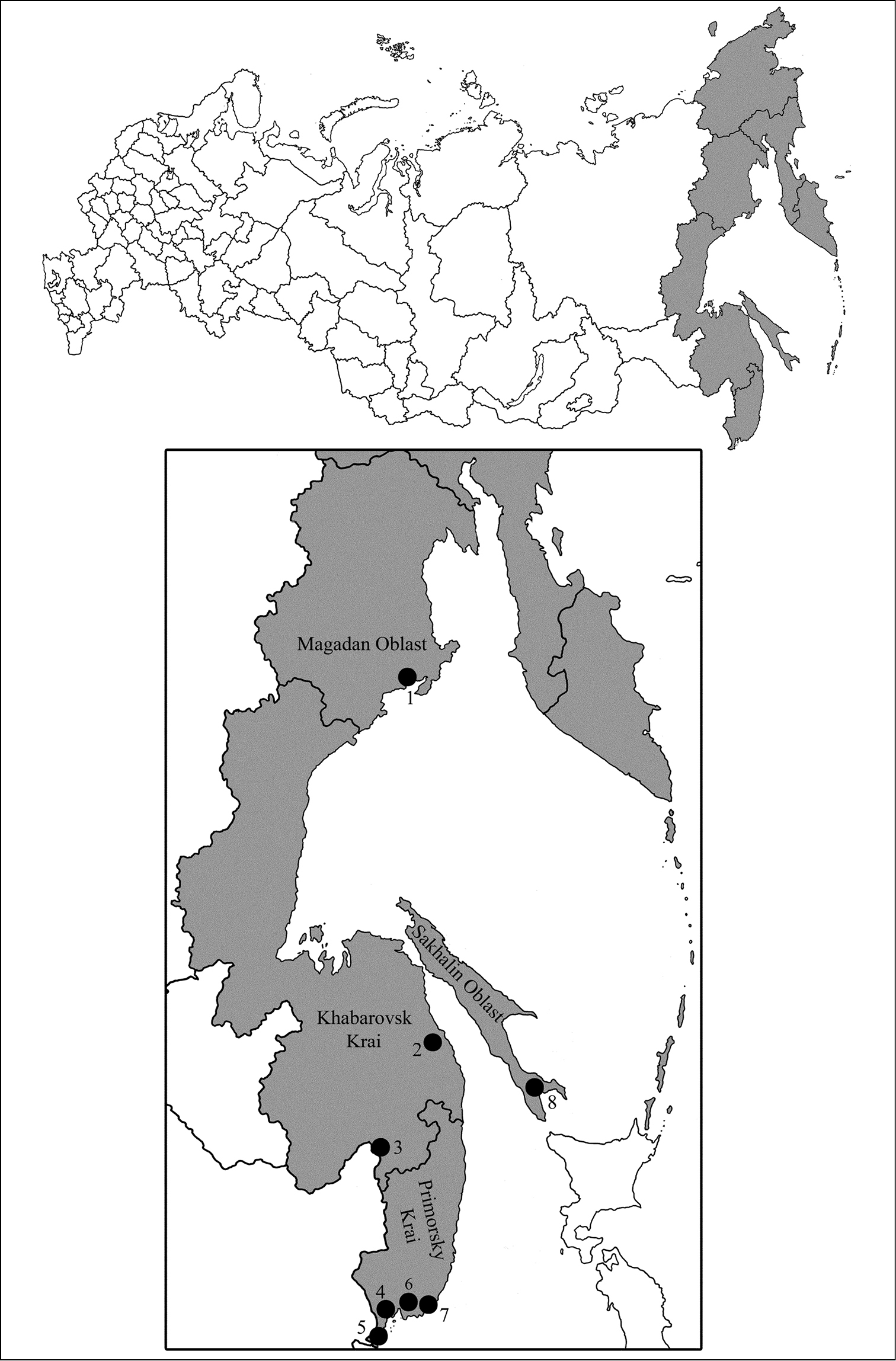
The sampling sites in RFE.
Much of the material was collected by us during a joint Chinese-Russian expedition in autumn of 2011 which was a part of study of the biodiversity of basal Hexapoda of Pacific coast of Asia (2011–2012). Several localities in southern RFE were searched: three locations of Primorsky Krai (Shkotovsky, Khasansky, and Lazovsky districts) and one of Khabarovsk Krai (Vaninsky district). Other materials collected by M. Potapov et al. (2009–2010) were also used. Around a thousand individuals are included in our study. Based on this material in all 25 species were identified, including 3 new species and 10 species newly recorded for the RFE. For another 12 species we give the new data on their distribution. The genera Fjellbergella and Yichunentulus are revised and rediagnosed, and the keys to species of the two genera are given.
The specimens were collected by Tullgren funnels using fast extraction with strong heating of samples during several hours. All specimens were mounted on slides in Hoyer’s medium and dried at 60 °C. Specimens were identified and drawn with the aid of a NIKON E600 phase contrast microscope. The photos were taken by digital camera Nikon DXM1200. Type specimens are deposited in the Shanghai Entomological Museum (SEM), Institute of Plant Physiology & Ecology, Shanghai Institutes for Biological Sciences, Chinese Academy of Sciences, and Moscow State Pedagogical University (MSPU).
Eight localities of RFE were sampled so far (Fig. 1) if including our collecting (denoted as 2–5 and 7); localities 1, 6 and 8 were studied in previous publications:
-
Magadan Oblast, Snezhnaja dolina, 59°32.92'N, 150°48.16'E.
-
Khabarovsk Krai, Vaninsky area, Valley of Mulinka, 49°49.42'N, 140°0.50'E.
-
Khabarovsk Krai, Khabarovsk district, Korfovsky, Khekhtsyr Range, 48°30.95'N, 135°6.06'E.
-
Primorsky Krai, Shkotovsky area, Anisimovka, Khualaza Mt. 43°10.33'N, 132°47.16'E.
-
Primorsky Krai, Khasansky area, 42°37.40'N, 130°52.35'E.
-
Primorsky Krai, Partizansky area, Chondolaz Range, 42°45.97'N, 133°3.75'E.
-
Primorsky Krai, Lazovsky area, nearby Preobrazheniye, 42°54.16'N, 133°53.33'E.
-
Sakhalin Oblast, Sakhalin Island, Yuzhno-Sakhalinsk, Susunaysky Range, Chenov Mt. 47°1.50'N, 142°52.35'E.
Abbreviations used in the text follow the paper of
http://zoobank.org/2AD017F3-F73F-4AF5-A0FE-C317387B9A77
Holotype, female (No. FE-2011022-2) (SEM), Russia, Far East, Primorsky Krai, Khasansky area, eastern part of Krabbe Peninsula, from mixed samples of soil and moss under a broad-leaved forest on a hill beside the coast, 42°37.40'N, 130°52.35'E, 16-IX-2011, coll. Y. Bu, C. W. Huang, M. Potapov & V. Alpatov. Paratypes, 2 females (Nos. FE-2011022-1, FE-2011023) (SEM; MSPU), 1 maturus junior (No. FE-2011022-3) (SEM), same data as holotype.
Adult body length 1000–1100 µm (n=3), pale yellow in color (Fig. 3A).
Head (Fig. 2A). Ovate, length 90–110 µm, width 70 µm. Setae d6 present, sd4 and sd5 short sensilliform. Setae d6 11 µm, d7 10 µm and sd7 19 µm in length. Clypeal pore cp and frontal pore fp present. Pseudoculus round, length 7–8 µm, with short posterior extension, PR=15 (Fig. 2B). Calyx of maxillary gland smooth, without any appendix, blind end split into two leaves, posterior filament 15 µm, CF=7 (Fig. 2C). Maxillary palpus with two tapering sensilla, subequal in length (Fig. 2D). Labial palpus reduced, with three setae and one lanceolate basal sensillum (Fig. 2E).
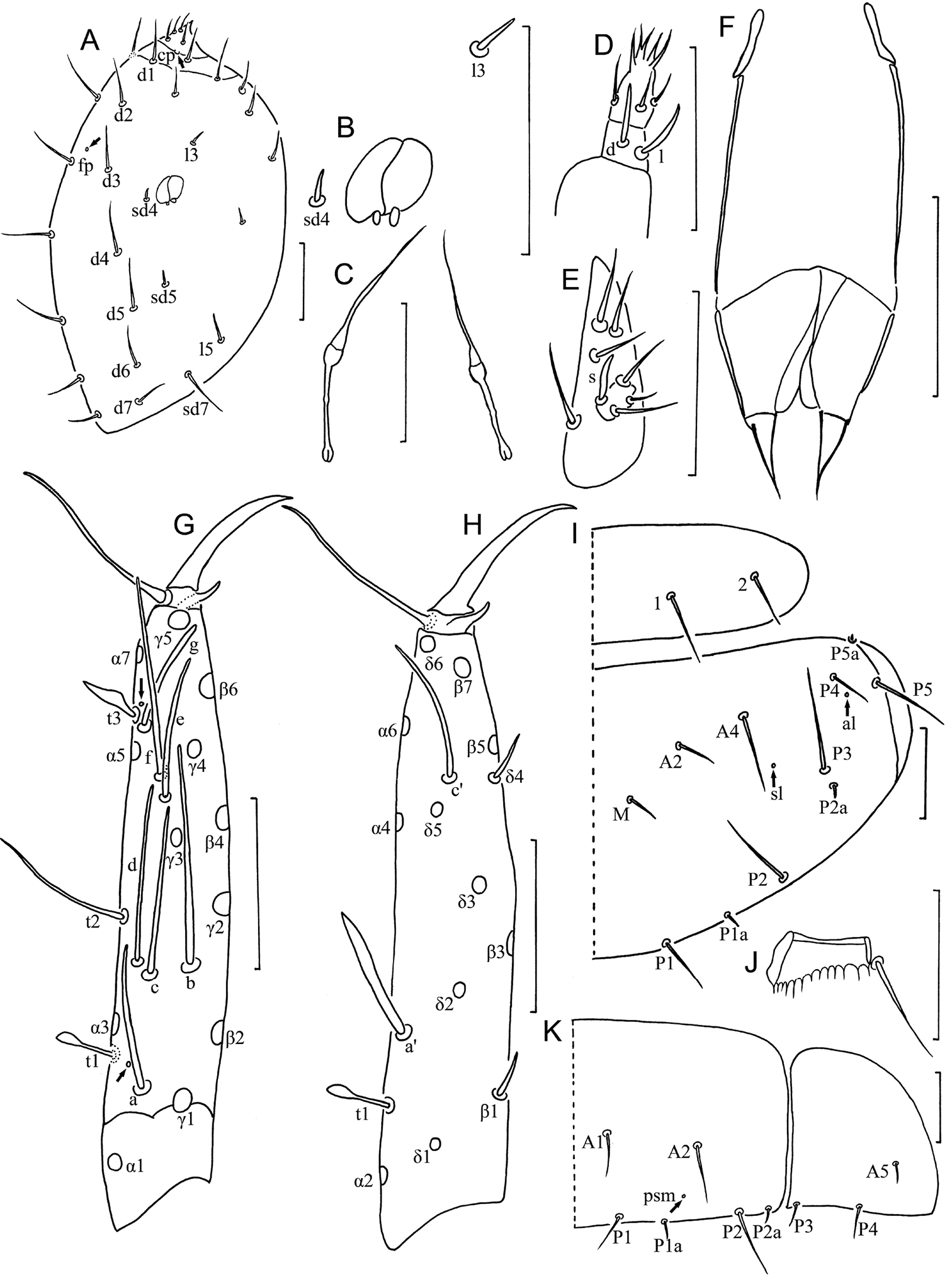
Baculentulus krabbensis sp. n. holotype. A Head, dorsal view (cp = clypeal pore, fp = frontal pore) B pseudoculus C canal of maxillary gland D maxillary palpus E labial palpus F female quama genitalis G foretarsus, exterior view H foretarsus, interior view I pronotum and mesonotum, right side (al = anterolateral pore, sl = sublateral pore) J comb K tergite I, right side. Arrows indicate pores. Scale bar: 20 μm.
Foretarsus (Fig. 2G, H). Length 80 µm, claw length 20 µm, without inner flap, TR=4; empodium length 3 µm, EU=0.15. Dorsal sensilla t-1 baculiform, t-2 slender and long (17 µm), BS=0.33, t-3 broad-leaf shape. All other exterior sensilla slender, except broadened sensillum g, a surpassing base of b and c, b broad at base and extremely long (29 µm), reaching base of γ4, c nearly reaching base of e and slightly lower than b and d, d located at same level to b, e short, f long, g broad and short. Interior sensilla a’ lanceolate and broad, b’ absent, c’ reaching base of claw. Relative length of sensilla: t1 = t3 < a’ < (g = t2 = c’) < a < (c = e) < d < f < b. Setae β1 and δ4 sensillum, 6 µm and 7 µm respectively. Pores close to base of sensilla a and t3 present. Length of middle tarsus 35 µm, claw length 15 µm. Length of hind tarsus 38 µm, claw length 18 µm.
Thorax. Thoracic chaetotaxy given in Table 1. Setae 1 and 2 on pronotum 16 µm and 12 µm length respectively, mesonotum and metanotum with eight pairs of P-setae, accessory setae short sensilliform; setae P1, P1a and P2 on mesonotum 15 µm, 3 µm and 20 µm respectively (Fig. 2I). Prosternum with two pairs of anterior seta, and setae A2 and M2 sensilliform (Fig. 3B). Mesosternum and metasternum each with 7 A-setae, and setae A2 sensilliform (Fig. 3C, D). Pronotum and prosternum without pores (Figs 2I, 3B). Mesonotum with pores sl and al, metanotum with pores sl only (Fig. 2I). Mesosternum and metasternum without pores (Fig. 3C, D). Single membranal pore present on membrane between each coxa and the body.
Baculentulus krabbensis sp. n. holotype. A Habitus B prosternum C mesosternum D metasternum E tergite III, left side F tergite VI, left side G tergite VIII H–J sternites V–VII. Arrows indicate pores. Scale bar: 100 μm in A, others, 20 μm.
Adult chaetotaxy of Baculentulus krabbensis sp. n.
| Segment | Dorsal | Ventral | |||
|---|---|---|---|---|---|
| Formula | Setae | Formula | Setae | ||
| Th. | I | 4 | 1, 2 | (4+4)/6 | A1, 2, M1, 2 P1, 2, 3 |
| II–III | 6/16 | A2, 4, M P1, 1a, 2, 2a, 3, 4, 5, 5a |
(7+2)/4 | Ac, 2, 3, 4, M Pc, 1, 2 |
|
| Abd. | I | 6/12 | A1, 2, 5 P1, 1a, 2, 2a, 3, 4 |
3/4 | Ac, 2 P1, 2 |
| II–III | 6/16 | A1, 2, 5 P1, 1a, 2, 2a, 3, 4, 4a, 5 |
3/5 | Ac, 2 Pc, 1a, 2 |
|
| IV–V | 6/16 | A1, 2, 5 P1, 1a, 2, 2a, 3, 4, 4a, 5 |
3/8 | Ac, 2 P1, 1a, 2, 3 |
|
| VI | 8/16 | A1, 2, 4, 5 P1, 1a, 2, 2a, 3, 4, 4a, 5 |
3/8 | Ac, 2 P1, 1a, 2, 3 |
|
| VII | 8/18 | A1, 2, 4, 5 P1, 1a, 2, 2a, 3, 3a, 4, 4a, 5 |
3/8 | Ac, 2 P1, 1a, 2, 3 |
|
| VIII | 6/16 | A2, 4, 5 M1, 2, 3, 4, P2, 3, 4, 5 |
4 | A1, 2 |
|
| IX | 14 | 1, 1a, 2, 2a, 3, 3a, 4 | 4 | 1, 2 | |
| X | 12 | 1, 1a, 2, 2a, 3, 4 | 4 | 1, 2 | |
| XI | 6 | 1, 3, 4 | 6 | 1, 1a, 2 | |
| XII | 9 | 6 | |||
Abdomen. Abdominal chaetotaxy given in Table 1. Tergite I with three pairs of anterior setae (A1, A2, A5) and six pairs of posterior setae (Fig. 2K). Tergites II–VI with eight pairs of posterior setae. Tergite VI and VII with four pairs of anterior setae (A1, A2, A4, A5). Tergites VII with nine pairs of posterior setae, P3a present. All accessory setae on tergites I–VII sensilliform, 5–6 µm on tergites I–VI and 7 µm on VII. Tergite VIII with paired setae M1 (Fig. 3G). Sternite IV–VII each with eight posterior setae, Pc absent (Fig. 3H, I, J). Sternite VIII with 4 setae.
Tergites I and VIII with pores psm only (Figs 2K, 3G), II–V with pores psm and al (Fig. 3E), VI–VII with pores psm, al and psl (Fig. 3F), IX–XI without pores, XII with single medial pore. Sternites I–III without pores, IV with 1+1 anteromembranal pores, V and VI each with 1+1 posterior pores anterior to seta P1 and 1+1 anteromembranal pores (Fig. 3H, I), VII with single posterior pore asymmetrical located left or right and 1+1 anteromembranal pores (Fig. 3J), VIII–XI without pores, XII with 1+1 pores al.
Abdominal appendages I, II, III with 2, 1, 1 segments and 4, 2, 2 setae respectively. On appendages II and III, subapical seta 15–17 µm, apical seta 11–12 µm in length. Striate band on abdominal segment VIII reduced, anterior margin regular wave shaped (Fig. 3G). Comb on abdomen VIII rectangular, with 12–13 teeth (Fig. 2J). Female squama genitalis with very short basal apodeme, extremely long and pointed acrostyli (Fig. 2F). Male unknown.
The species is named after Krabbe Peninsula where the type specimens were collected.
Known only from type locality.
Baculentulus krabbensis sp. n. is characterized by extremely long sensilum b on foretarsus, sensillum a’ located distal to t1, sensillum b’ absent, eight A-setae on tergite VI and VII, presence of P3a on tergite VII, and special female genitalia with short basal apodeme.
We placed the present new species in the genus Baculentulus because the baculiform sensillum t1 on foretarsus, reduced labial palpus with three setae and one sensillum, smooth calyx of maxillary gland, reduced striate band, two pairs of anterior setae on mesonotum and metanotum, abdominal appendages II and III each with two setae of different length, and only 4 setae on sternite VIII. It is similar to Baculentulus samchonri (Imadaté & Szeptycki, 1976) from North Korea in having eight A-setae on both tergites VI and VII, absence of sensillum b’ and extremely long sensillum b on foretarsus. They can be distinguished by the posterior setae on tergite VII (9 pairs of P-setae with P3a present in Baculentulus krabbensis sp. n. vs. 8 pairs of P-setae with P3a absent in Baculentulus samchonri), the anterior setae on tergite I (6 A-setae in Baculentulus krabbensis sp. n. vs. 4 in Baculentulus samchonri), the length of sensillum f (extremely long and surpassing the base of claw in Baculentulus krabbensis sp. n. vs. short and not reaching base of claw in Baculentulus samchonri), and the shape of female squama genitalis (basal apodeme very short in Baculentulus krabbensis sp. n. vs. basal apodeme in moderate length in Baculentulus samchonri).
http://zoobank.org/152D05AF-CE80-4BDA-B821-833163392DC1
Holotype, female (No. FE-2011051-2) (SEM), Russia, Far East, Primorsky Krai, Lazovsky area, nearby Preobrazheniye, from the mixed samples of humus (full of white mycelium) and a little soil under a broad-leaved forest close to the seashore, 42°54.16'N, 133°53.33'E, 22-IX-2011. coll. Y. Bu, C. W. Huang, M. Potapov & V. Alpatov. Paratypes, 5 females (Nos. FE-2011046-2, FE-2011052-4, FE-2011052-5, FE-2011052-6, FE-2011073-1) (SEM), 5 males (Nos. FE-2011071-1, FE-2011072-1, FE-2011074-1, FE-2011075-1, FE-211075-3) (SEM; MSPU), same data as holotype. Other materials, 3 male preimagos (Nos. FE-2011048-2, FE-2011048-3, FE-2011049-3) (SEM), same date as holotype.
Adult body length 1100–1300 µm (n=10), pale yellow in color (Fig. 5A).
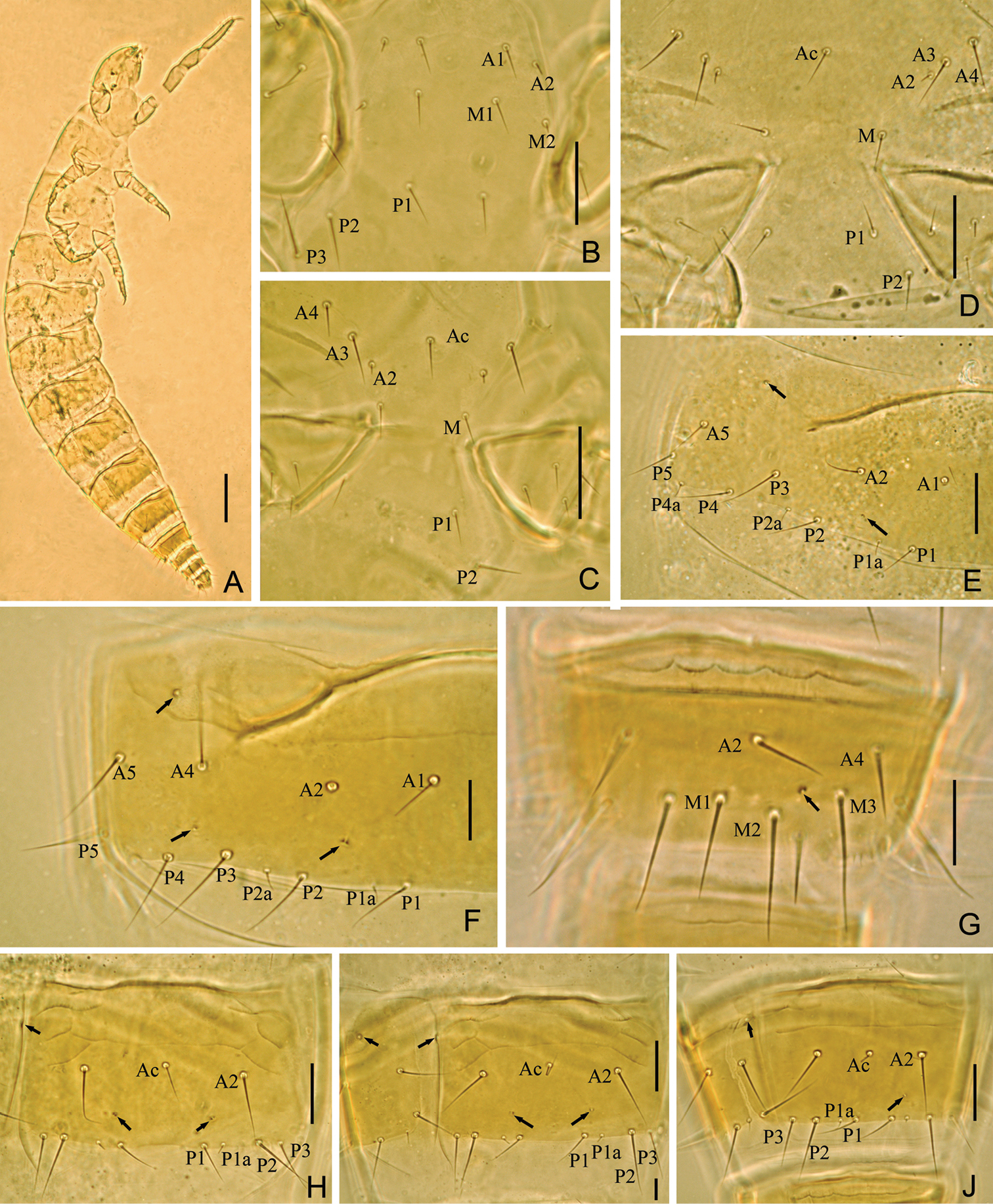
Head (Fig. 4A). Ovate, length 120–125 µm, width 85 µm. Setae d6 present, sd4 and sd5 short sensilliform. Setae d6 and d7 subequal in length. Seta sd7 18 µm in length. Clypeal pore cp and frontal pore fp present. Pseudoculus round, length 9–10 µm, with short posterior extension, some irregular lines visible under light microscope, PR=12–13 (Fig. 4B). Calyx of maxillary gland smooth, with one helmet-like dorsal appendix, blind end split into two leaves, and posterior filament 15 µm, CF=8 (Fig. 4C). Maxillary palpus with two tapering sensilla, dorsal one (8–10 µm) slightly longer than lateral one (7–9 µm) (Fig. 4D). Labial palpus reduced, with two-branched terminal tuft of setae, with one leaf-shape basal sensillum (Figs 4E, 5B).
Fjellbergella lazovskiensis sp. n. holotype. A Head, dorsal view B pseudoculus C canal of maxillary gland D maxillary palpus E labial palpus F female squama genitalis G male squama genitalis H foretarsus, exterior view I foretarsus, interior view J nota, right side K part of striate band L comb. Arrows indicate pores. Scale bar: 20 μm.
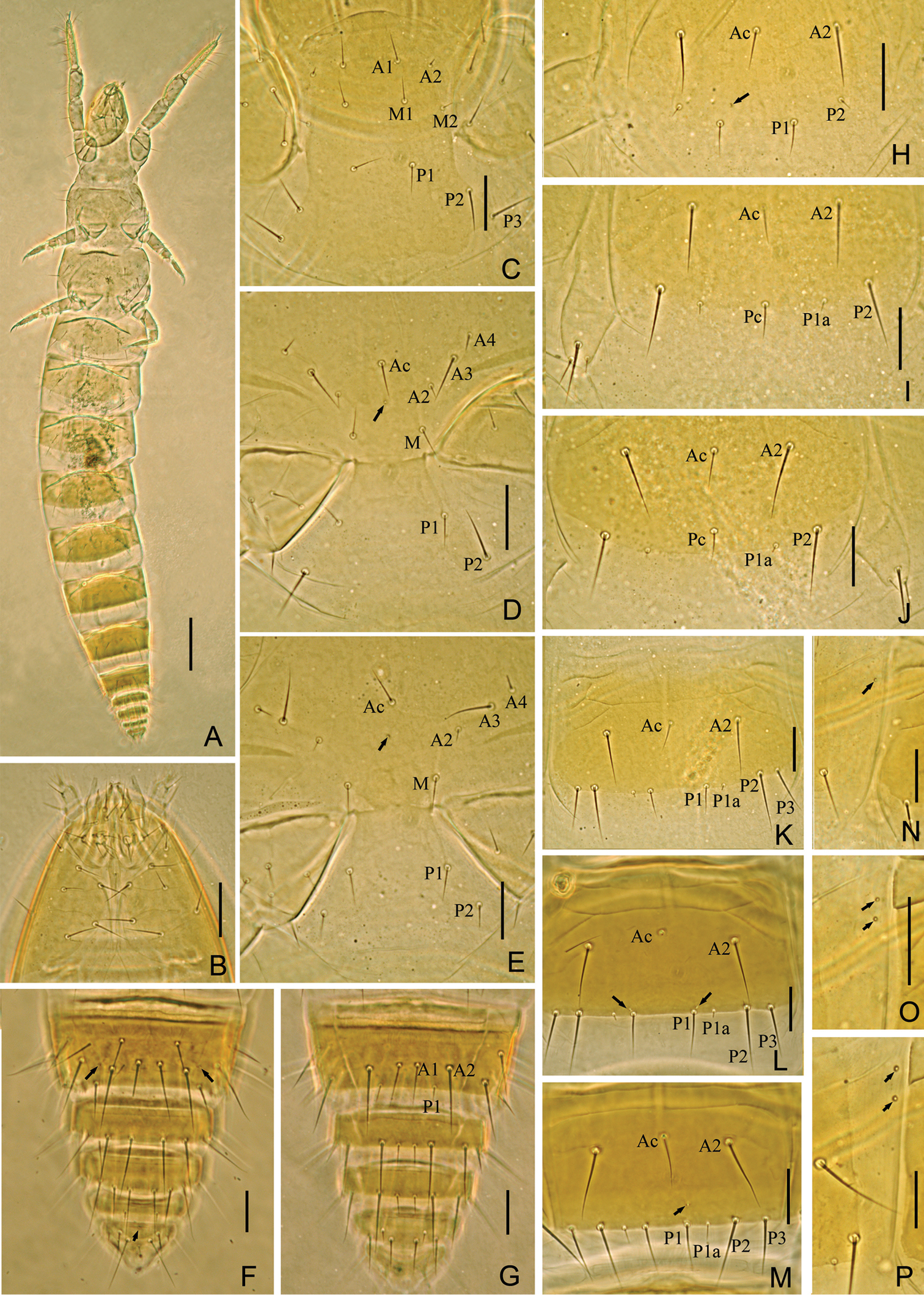
Fjellbergella lazovskiensis sp. n. holotype. A Habitus B ventral side of head C prosternum D mesosternum E metasternum F tergites VIII–XII G sternites VIII–XII H–J sternites I–III K sternite IV L sternite VI M sternite VII N–P laterotergites of abdominal segments IV, V and VI. Arrows indicate pores. Scale bar: 100 μm in A, others, 20 μm.
Foretarsus (Fig. 4H, I). Length 90–95 µm, claw length 20–21 µm, without inner flap, TR=4.3–4.7; empodium length 4 µm, EU=0.2. Dorsal sensilla t-1 claviform, t-2 slender and long (25–26 µm), BS=0.38–0.42, t-3 short and lanceolate. All other exterior sensilla slender, with except a slightly broad, surpassing base of c, b shorter than c and located in subequal level, d lower than c, e slender, f long, g short. Interior sensilla a’ short and broad, b’ slender and surpassing base of δ5, c’ reaching base of claw. Relative length of sensilla: t3 < t1< a’ < b < (a = g = d = b’ = c’) < (c = e = f = t2). Setae β1 and δ4 sensillum, 7 µm and 5 µm respectively. Pores close to base of sensilla a and t3 present. Length of middle tarsus 40–45 µm, claw length 18–20 µm. Length of hind tarsus 45–50 µm, claw length 20–23 µm.
Thorax. Thoracic chaetotaxy given in Table 2. Setae 1 and 2 on pronotum 20 µm and 15 µm length respectively, mesonotum and metanotum with eight pairs of P-setae, accessory setae short sensilliform, 2.5 µm; setae P1, P1a and P2 on mesonotum 18–20 µm, 2.5 µm and 22–25 µm respectively (Fig. 4J). Prosternum with two pairs of anterior seta, and setae A2 and M2 sensilliform (Fig. 5C). Mesosternum and metasternum each with 7 A-setae, and setae A2 and A4 sensilliform (Fig. 5D, E). Pronotum and prosternum without pores (Figs 4J, 5C). Mesonotum with pores sl and al, metanotum with pores sl only (Fig. 4J). Mesosternum and metasternum each with single median pore, situated anterioral to level of setae M (Fig. 5D, E).
Adult chaetotaxy of Fjellbergella lazovskiensis sp. n.
| Segment | Dorsal | Ventral | |||
|---|---|---|---|---|---|
| Formula | Setae | Formula | Setae | ||
| Th. | I | 4 | 1, 2 | (4+4)/6 | A1, 2, M1, 2 P1, 2, 3 |
| II–III | 6/16 | A2, 4, M P1, 1a, 2, 2a, 3, 4, 5, 5a |
(7+2)/4 | Ac, 2, 3, 4, M Pc, 1, 2 |
|
| Abd. | I | 6/12 | A1, 2, 5 P1, 1a, 2, 2a, 3, 4 |
3/4 | Ac, 2 P1, 2 |
| II–III | 6/16 | A1, 2, 5 P1, 1a, 2, 2a, 3, 4, 4a, 5 |
3/5 | Ac, 2 Pc, 1a, 2 |
|
| IV–V | 6/16 | A1, 2, 5 P1, 1a, 2, 2a, 3, 4, 4a, 5 |
3/8 | Ac, 2 P1, 1a, 2, 3 |
|
| VI | 8/16 | A1, 2, 4, 5 P1, 1a, 2, 2a, 3, 4, 4a, 5 |
3/8 | Ac, 2 P1, 1a, 2, 3 |
|
| VII | 8/18 | A1, 2, 4, 5 P1, 1a, 2, 2a, 3, 3a, 4, 4a, 5 |
3/8 | Ac, 2 P1, 1a, 2, 3 |
|
| VIII | 6/16 | A2, 4, 5 M1, 2, 3, 4, P2, 3, 4, 5 |
4/2 | A1, 2 P1 |
|
| IX | 14 | 1, 1a, 2, 2a, 3, 3a, 4 | 4 | 1, 2 | |
| X | 12 | 1, 1a, 2, 2a, 3, 4 | 4 | 1, 2 | |
| XI | 6 | 1, 3, 4 | 6 | 1, 1a, 2 | |
| XII | 9 | 6 | |||
Abdomen. Abdominal chaetotaxy given in Table 2. Tergite I with three pairs of anterior setae (A1, A2, A5) and six pairs of posterior setae. Tergites II–VI with eight pairs of posterior setae. Tergite VI and VII with four pairs of anterior setae (A1, A2, A4, A5). Tergite VII with nine pairs of posterior setae, P3a present. All accessory setae on tergites I–VII short sensilliform, 3 µm on tergites I–VI and 6 µm on VII. Tergite VIII with paired setae M1 (Fig. 5F). Sternites IV–VII each with eight posterior setae, Pc absent (Fig. 5K, L, M). Sternite VIII with two rows of setae (4/2) (Fig. 5G).
Tergites I and VIII with pores psm only, II–VII with pores psm and al, IX–XI without pores, XII with single medial pore (Fig. 5F). Sternites I–III without pores (Fig. 5H–J), IV with 1+1 anteromembranal pores (Fig. 5K, N), V and VI each with 1+1 posterior pores close to seta P1 and 2+2 anteromembranal pores (Fig. 5L, O, P), VII with single posterior pore asymmetrical located left or right (Fig. 5M), VIII–XI without pores, XII with 1+1 pores al.
Abdominal appendages I, II, III with 2, 1, 1 segments and 4, 3, 3 setae respectively. On appendages II and III, subapical seta 19–21 µm, two apical setae 11–12 µm, and 5–7 µm in length (Fig. 5I–J). Striate band on abdominal segment VIII reduced, anterior margin with sparse irregular teeth (Figs 4K, 5F, G). Comb on abdomen VIII rectangular, with 12 teeth (Fig. 4L). Female squama genitalis with short subuliform acrostyli (Fig. 4F). Male squama genitalis with 5+5 setae on dorsal side and 3+3 setae on ventral side (Fig. 4G).
Chaetal variability. Chaetal variations were observed in 4 specimens: on tergite VI, asymmetrical absence of A4 of right side (No. FE-2011071-1); on tergite VII, asymmetrical absence of P1 of right side (No. FE-2011052-4); on sternite VII, absence of Ac (No. FE-2011075-1), and asymmetrical absence of A2 of left side (No. FE-2011052-6).
The species is named after Lazovsky Nature Reserve where the type specimens were collected.
Known only from type locality.
Fjellbergella lazovskiensis sp. n. is characterized by three pairs of anterior setae on tergites II-V which is different to any other members of the genus, four pair of anterior setae, nine pairs of posterior setae on tergite VII, foretarsal sensilla b, c and d located in subequal level, b shorter than c, and tergite IX and X with 14, 12 setae respectively.
The present species is located in the genus Fjellbergella because three setae on abdominal legs, labial palpus with two-branched terminal tuft of setae, claviform sensillum t1 on foretarsus, reduced striate band, two pairs of anterior setae on mesonotum and metanotum, and 4/2 setae on sternite VIII. Fjellbergella lazovskiensis sp. n. is similar to Fjellbergella tuxeni Nosek, 1980 from Alaska in having 8 A-setae on tergite VII and presence of P1a on tergite I–VII. They can be distinguished by the chaetotaxy of tergite IX and X (14 and 12 setae in Fjellbergella lazovskiensis sp. n. respectively vs. 12 and 8 setae in Fjellbergella tuxeni), length and location of foretarsal sensilla b and c (b shorter than c and they located at the same level in Fjellbergella lazovskiensis sp. n. vs. b and c subequal in length and c located lower than b distinctly in Fjellbergella tuxeni), shape and length of sensillum a’ (broad and reaching base of sensillum b’ in Fjellbergella lazovskiensis sp. n. vs. slender and far surpassing base of sensillum b’ in Fjellbergella tuxeni), and shape of claw of foreleg (absence of inner flap in Fjellbergella lazovskiensis sp. n. vs. presence in Fjellbergella tuxeni). In addition, the new species has only 6 anterior setae on tergites II-V contrary to two other members of this genus, which have 8 anterior setae.
http://zoobank.org/513B6344-D78B-4777-90A9-AEB3BAEAB83E
Holotype, female (No. FE-2011035-1) (SEM), Russia, Far East, Primorsky Krai, Lazovsky area, nearby Preobrazheniye, from the mixed samples of soil and humus from mountains, 42°54. 48'N, 133°53. 96'E, 21-IX-2011, coll. Y. Bu, C. W. Huang, M. Potapov & V. Alpatov. Paratypes, 1 female (No. FE-2011035-6), 3 males (Nos. FE-2011035-4, FE-2011037-1, FE-2011045-3) (SEM; MSPU), 2 male preimagos (Nos. FE-2011038-1, FE-2011038-2) (SEM), same data as holotype.
Adult body length 1200–1350 µm (n=5).
Head (Fig. 6A). Ovate, length 120–130 µm, width 70–80 µm. Setae d6 present, sd4 and sd5 short sensilliform. Setae d6 11 µm, d7 10 µm and sd7 18 µm in length. Clypeal pore cp and frontal pore fp present. Pseudoculus round, length 7–8 µm, with short posterior extension, PR=16 (Fig. 6B). Calyx of maxillary gland smooth, without any appendix, blind end split into two leaves, posterior filament 16–17 µm, CF=7–8 (Fig. 6C). Maxillary palpus with two tapering sensilla, dorsal snsillum is evidently longer than lateral one (Fig. 6D). Labial palpus completed, with one-branched terminal tuft of setae, with three setae and one lanceolate basal sensillum (Fig. 6E).
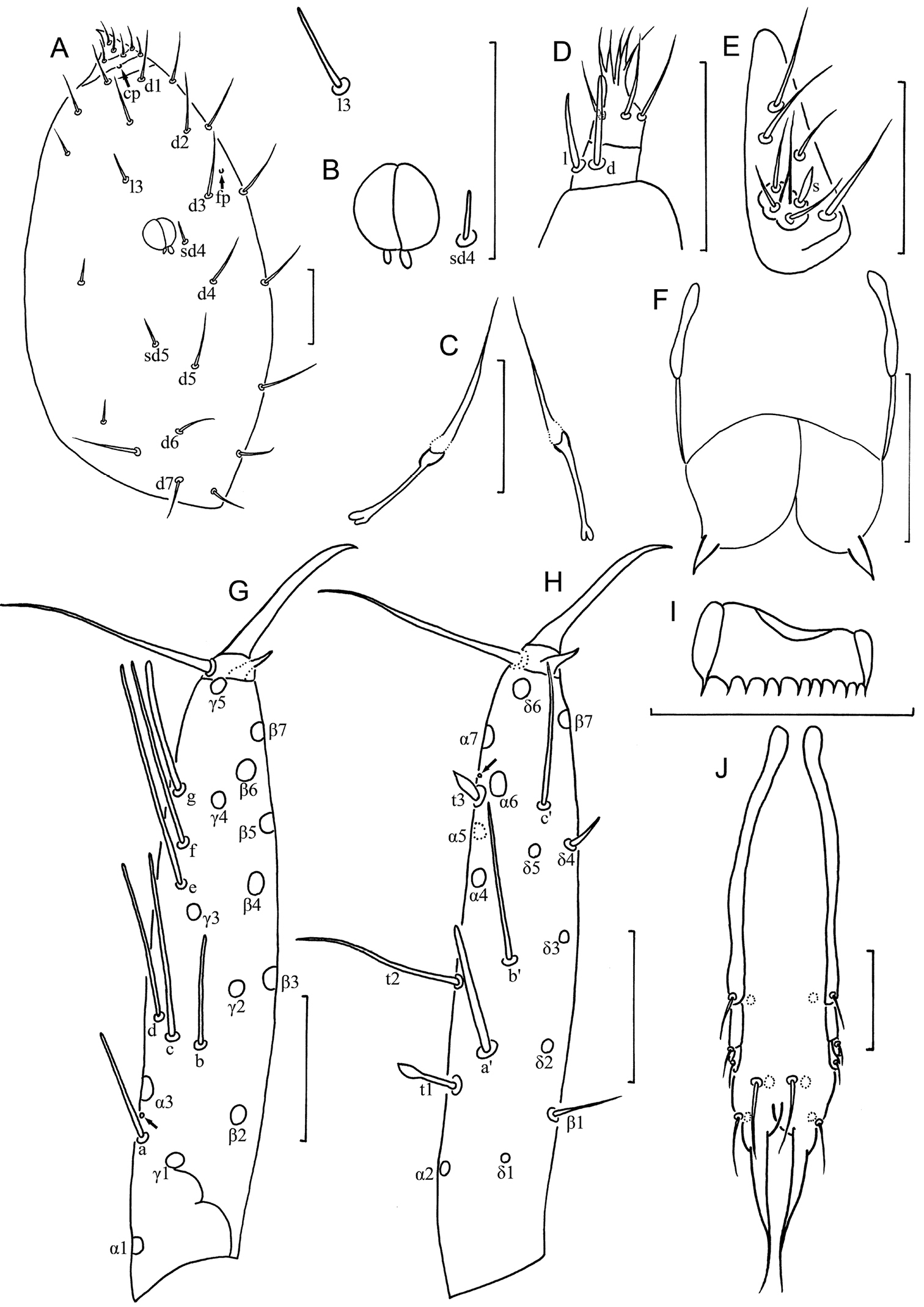
Yichunentulus alpatovi sp. n. holotype. A Head, dorsal view B pseudoculus C canal of maxillary gland D maxillary palpus E labial palpus F female squama genitalis G foretarsus, exterior view H foretarsus, interior view I comb J male squama genitalis. Arrows indicate pores. Scale bar: 20 μm.
Foretarsus (Fig. 6G, H). Length 86–93 µm, claw length 20–23 µm, without inner flap, TR=4; empodium length 4 µm, EU=0.18. Dorsal sensilla t-1 baculiform, t-2 slender and long (23 µm), BS=0.5, t-3 lanceolate. Exterior sensilla a surpassing base of b and c, b slender and short (16 µm), not reaching base of γ3, c reaching base of f and in subequal level to b, d located higher than b and c, e long and reaching base of claw (30 µm), f slender, g broad and short (18 µm). Interior sensilla a’ broad, b’ slender and reaching base of α6, c’ nearly reaching base of claw. Relative length of sensilla: t3 < t1 < b < (a = a’) < g < (b’ = c’) < d < t2 < c < e < f. Setae β1 setiform and δ4 sensilliform. Pores close to sensilla a and t3 present. Length of middle tarsus 40 µm, claw length 17 µm. Length of hind tarsus 45 µm, claw length 20 µm.
Thorax. Thoracic chaetotaxy given in Table 3. Setae 1 and 2 on pronotum 20 µm and 12–14 µm length respectively. Mesonotum and metanotum with eight pairs of P-setae, accessory setae short sensilliform, 2 µm in length; setae P1, P1a and P2 on mesonotum 17–20 µm, 3 µm and 20–24 µm respectively. Prosternum with two pairs of anterior seta, and setae A2 and M2 sensilliform. Mesosternum and metasternum each with 7 A-setae, and setae A2 sensilliform. Pronotum and prosternum without pores. Mesonotum with pores sl and al, metanotum with pores sl only. Mesosternum and metasternum each with single median pore, situated anterioral to level of setae M.
Adult chaetotaxy of Yichunentulus alpatovi sp. n.
| Segment | Dorsal | Ventral | |||
|---|---|---|---|---|---|
| Formula | Setae | Formula | Setae | ||
| Th. | I | 4 | 1, 2 | (4+4)/6 | A1, 2, M1, 2 P1, 2, 3 |
| II–III | 6/16 | A2, 4, M P1, 1a, 2, 2a, 3, 4, 5, 5a |
(7+2)/4 | Ac, 2, 3, 4, M Pc, 1, 2 |
|
| Abd. | I | 6/12 | A1, 2, 5 P1, 1a, 2, 2a, 3, 4 |
3/4 | Ac, 2 P1, 2 |
| II–III | 6/16 | A1, 2, 5 P1, 1a, 2, 2a, 3, 4, 4a, 5 |
3/5 | Ac, 2 Pc, 1a, 2 |
|
| IV–V | 6/16 | A1, 2, 5 P1, 1a, 2, 2a, 3, 4, 4a, 5 |
3/8 | Ac, 2 P1, 1a, 2, 3 |
|
| VI | 8/16 | A1, 2, 4, 5 P1, 1a, 2, 2a, 3, 4, 4a, 5 |
3/8 | Ac, 2 P1, 1a, 2, 3 |
|
| VII | 8/16 | A1, 2, 4, 5 P1, 1a, 2, 2a, 3, 3a, 4, 4a, 5 |
3/8 | Ac, 2 P1, 1a, 2, 3 |
|
| VIII | 6/16 | A2, 4, 5 M1, 2, 3, 4, P2, 3, 4, 5 |
4/2 | A1, 2 P1 |
|
| IX | 14 | 1, 1a, 2, 2a, 3, 3a, 4 | 4 | 1, 2 | |
| X | 12 | 1, 1a, 2, 2a, 3, 4 | 4 | 1, 2 | |
| XI | 6 | 1, 3, 4 | 6 | 1, 2 | |
| XII | 9 | 6 | |||
Abdomen. Abdominal chaetotaxy given in Table 3. Tergite I with three pairs of anterior setae (A1, A2, A5). Tergites II–VI with eight pairs of posterior setae. Tergites VI and VII with four pairs of anterior setae (A1, A2, A4, A5). Tergite VII with nine pairs of posterior setae, P3a present. All accessory setae on tergites I–VII short sensilliform, 3 µm on tergites I–V, 4 µm on tergite VI, and 5 µm on VII. Tergite VIII with paired setae M1. Sternite IV–VII each with eight posterior setae, Pc absent. Sternite VIII with two rows of setae (4/2).
Tergites I and VIII with pores psm only, II–V with pores psm and al, VI–VII with pores psm, al and psl, IX–XI without pores, XII with single medial pore. Sternites I–IV without pores, V and VI each with 1+1 posterior pores anterior to seta P1 and 1+1 anteromembranal pores, VII with single posterior pore asymmetrical located left or right and 1+1 anteromembranal pores, VIII–XI without pores, XII with 1+1 pores al.
Abdominal appendages I, II, III with 2, 1, 1 segments and 4, 2, 2 setae respectively. On appendages II and III, subapical setae 16–17 µm, apical setae 13–14 µm in length. Striate band on abdominal segment VIII reduced, anterior margin regular wave shaped. Comb on abdomen VIII rectangular, with 12–13 teeth (Fig. 6I). Female squama genitalis robust, with moderate basal apodeme and pointed acrostyli (Fig. 6F). Male squama genitalis with 5+5 setae on dorsal side and 3+3 setae on ventral side (Fig. 6J).
The species is named after Dr. V. Alpatov who accompanied us during our scientific trips.
Known only from type locality.
Yichunentulus alpatovi sp. n. is characterized by the presence of sensillum b’ on foretarus and short sensilum b on foretarsus, 6 anterior seate on tergites IV–V, swelled sensillum a’, presence of pores psl only on tergites VI and VII, and female squama genitalis with moderate basal apodeme and pointed acrostyli.
The present species is located in the genus Yichunentulus because the labial palpus with one-branched terminal tuft of setae, with three setae and one lanceolate basal sensillum, the baculiform sensillum t1 on foretarus, reduced striate band, two pairs of anterior setae on mesonotum and metanotum, 4/2 setae on sternite VIII, and abdominal appendages II and III each with two setae of different length. Yichunentulus alpatovi sp. n. is close to the type species Yichunentulus yichunensis Yin, 1980 in having short sensillum b and identical body chaetotaxy. However, it can be easily distinguished from Yichunentulus yichunensis and Yichunentulus borealis (Nakamura, 2004), comb. n. by the presence of sensillum b’ on foretarsus. It also differs from Yichunentulus yichunensis in the length of sensillum e (extremely long in Yichunentulus alpatovi sp. n. vs. short in Yichunentulus yichunensis) and sensilla c’ (long and surpassing base of δ6 in Yichunentulus alpatovi sp. n. vs. short and only reaching base of β7 in Yichunentulus yichunensis). It differs from Yichunentulus borealis in the chaetotaxy of tergites IV–V (each with 6 A-setae in Yichunentulus alpatovi sp. n. vs. 8 in Yichunentulus borealis), shape of sensillum a’ (slightly broad and as long as sensillum a in Yichunentulus alpatovi sp. n. vs. distinctly swell and shorter than sensillum a in Yichunentulus borealis), and the body porotaxy (pore psl present on tergites VI and VII only and sternites I–IV without pores in Yichunentulus alpatovi sp. n. vs. psl present on tergites III–VII and sternites II and IV each with 1+1 anteromembranal pores in Yichunentulus borealis).
Acerentomon confine Berlese, 1908.
Abdominal appendages II and III each with 3 setae, mesonotum and metanotum each with two pairs of anterior setae, foretarsal sensillum t1 claviform, sensillum b’ present, t3 willow-leaf shaped, sensillum b’ present, labial palpus with terminal tuft of setae, striate band on abdominal segment VIII well developed, and sternite VIII with 4/2 setae or with 4 setae only.
The genus Acerentulus has 47 species described in the world and most from Europe (
Fjellbergella tuxeni Nosek, 1980.
Mesonotum and metanotum each with two pairs of anterior setae, labial palpus with two-branched terminal tuft of setae, sensillum d located near t2 insertion, abdominal appendages II and III each with three setae, foretarsal sensillum t1 claviform, sensillum b’ present, striate band on abdominal segment VIII reduced and sternite VIII with 4/2 setae.
The genus Fjellbergella containes only two species so far: Fjellbergella tuxeni Nosek, 1980 from Alaska and Fjellbergella uteorum Shrubovych & Bernard, 2013 from Colorado (
| 1 | Tergite VII with 4 pairs of anterior setae, seta Pc absent | 2 |
| – | Tergite VII with 5 pairs of anterior setae, seta Pc present | Fjellbergella uteorum Shrubovych & Bernard, 2013; USA (Colorado) |
| 2 | Tergites I–VI without seta P1a | Fjellbergella jilinensis (Wu & Yin, 2007), comb. n.; China (Jilin) |
| – | Tergites I–VI with seta P1a | 3 |
| 3 | Tergites IX and X with 14 and 12 setae respectively, sensillum b shorter than c and they located at the same level, claw of foreleg without inner flap | Fjellbergella lazovskiensis sp. n.; Russia (Far East) |
| – | Tergites IX and X with 12 and 8 setae respectively, sensillum b and c subequal in length and c located lower than b distinctly, claw of foreleg with one inner flap | Fjellbergella tuxeni Nosek, 1980; USA (Alaska) |
Holotype, female, Paratype, 1 female, China, Jilin Province, Dehui city, 15-IX-2006, coll. D. H. Wu.
China (Jilin).
Yichunentulus yichunensis Yin, 1980.
Mesonotum and metanotum each with two pairs of anterior setae, abdominal appendages II and III each with two setae, labial palpus with one-branched terminal tuft of setae, with 3 setae and 1 sensillum, foretarsal sensillum t1 baculiform, sensillum b’ absent or present, sensillum a’ located distal to t1, maxillary gland simple and without appendages, sternite VIII with 4/2 setae, striate band on abdominal segment VIII reduced, and sternites II–VI each has 1+1 membranal pores.
The genus Yichunentulus Yin, 1980 has only one species described from Heilongjiang, Northeast China (
| 1 | Tergites IV and V each with 4 pairs of anterior setae | Yichunentulus borealis (Nakamura, 2004), comb. n. |
| – | Tergites IV and V each with 3 pairs of anterior setae | 2 |
| 2 | Foretarsal sensillum b’ absent | Yichunentulus yichunensis Yin, 1980 |
| – | Foretarsal sensillum b’ present | Yichunentulus alpatovi sp. n. |
1 female, 1 male, Locality 3, 25-IV-2010, coll. E. Sokolova & M. Potapov.
Russia (Far East, Khabarovsk Krai).
Body length 1300–1380 µm, foretarsus length 110 µm. We studied the new materials of Yichunentulus borealis collected from type locality and redescribe the head chaetotaxy and body porotaxy. Head with d6 seta present, sd4 and sd5 sensilliform. Pronotum and prosternum without pores. Mesonotum with pores sl and al, metanotum with pores sl only. Mesosternum and metasternum each with single medial pore. Tergites I and VIII with pores psm only, II with pores psm and al, III–VII with pores psm, al and psl, IX–XI without pores, XII with single medial pore. Sternites I with single medial pore, II–IV without 1+1 anteromembranal pores, V and VI each with 1+1 posterior pores anterior to seta P1 and 1+1 anteromembranal pores, VII with single posterior pore asymmetrical located left or right, VIII–XI without pores, XII with 1+1 pores al.
7 females (Nos. FE-2011062-1, FE-2011062-4, FE-2011062-5, FE-2011085-1, FE-2011085-2, FE-2011087-1, FE-2011089-1) (SEM; MSPU), 5 males (Nos. FE-2011062-2, FE-2011062-3, FE-2011086-1, FE-2011086-2, FE-2011087-2) (SEM; MSPU), 1 male preimago (No. FE-2011090-2), Russia, Far East, Khabarovsk Krai, Vaninsky area, nearby Vysokogorny, Valley of Mulinka, from Picea and Abies forest on Northeast slope, sample No. 27, 750 m alt., 30-IX-2011, 1 male preimago (No. FE-2011059-2), 1 maturus junior (No. FE-2011059-1), Russia, Far East, Khabarovsk Krai, Vaninsky area, nearby Datta, from coastal larch-wood on East slope (green moss), sample No. 24, 28-IX-2011. 1 female (No. FE-2011060-1), Russia, Far East, Khabarovsk Krai, Vaninsky area, nearby Vysokogorny, Valley of Mulinka, from spruce forest at pass, sample No. 26, 900 m alt., 29-IX-2011. 2 females (FE-2011067-2, FE-2011088-1), 1male (No. FE-2011088-2), Russia, Far East, Khabarovsk Krai, Vaninsky area, nearby Vysokogorny, Valley of Mulinka, from mixed sample of spruce-forest and rotten wood, sample No. 30, 600 m alt., 29-ix-2011. 1 female (No. FE-2011069-1), Russia, Far East, Khabarovsk Krai, Vaninsky area, nearly Vysokogorny (5 km South), Valley of Dopolnitel’ny Stream, from mixed sample of litter under old poplar-trees in floodplain, sample No. 32, 400 m alt., 30-IX-2011. All specimens are collected by M. Potapov & V. Alpatov.
Adult body length 1000–1200 µm (n=16), yellow in color (Fig. 8A).
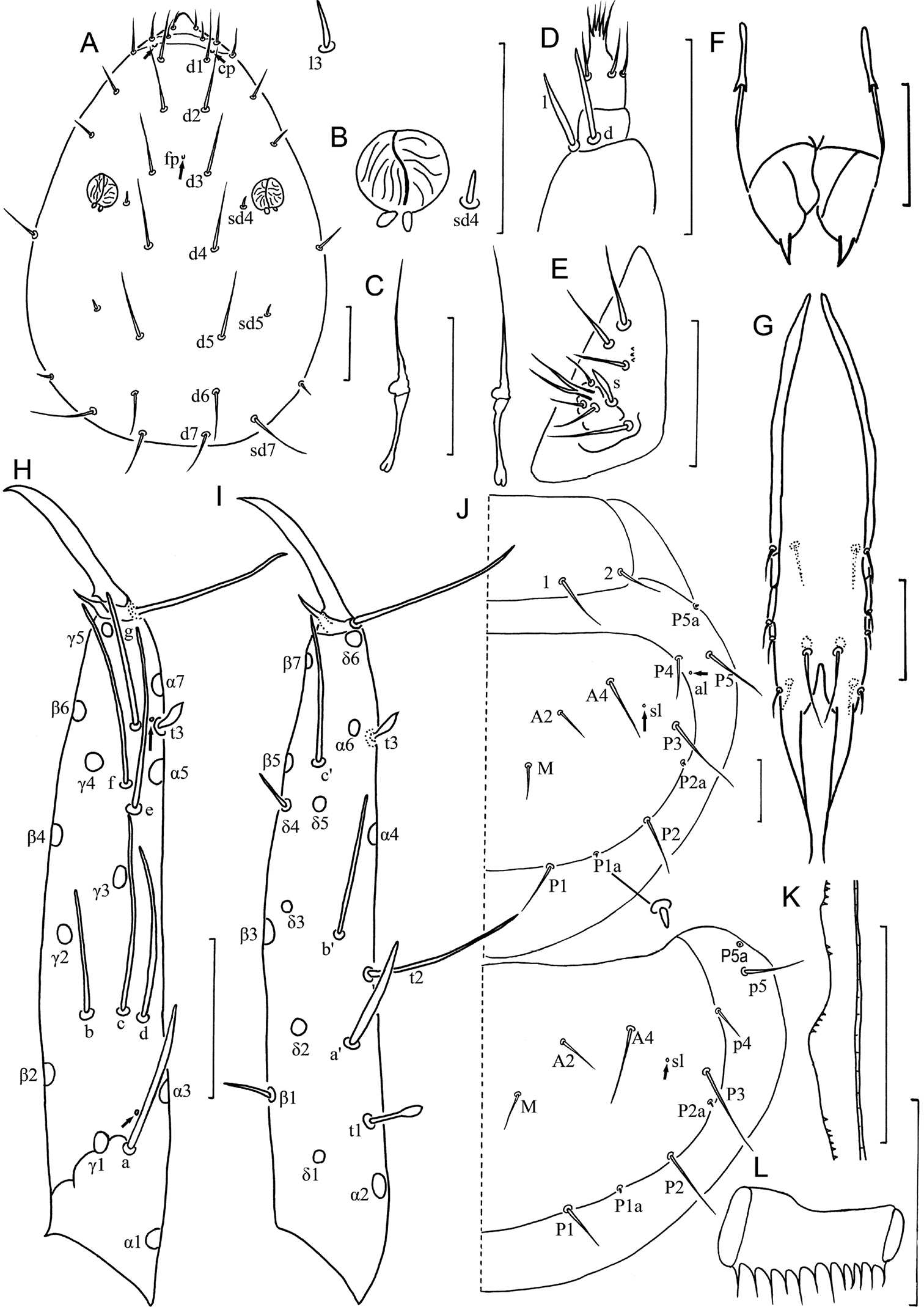
Head (Fig. 7A). Ovate, length 130–135 µm, width 80–85 µm. Setae d6 present, sd4 and sd5 short. Setae d6 14–15 µm, d7 16–17 µm and sd7 18–19 µm in length. Clypeal pore cp and frontal pore fp present. Pseudoculus round, length 7–8 µm, with short posterior extension, PR=16–17 (Fig. 7B). Maxillary gland large, calyx with lateral racemose appendices and one helmet-like dorsal appendix, and bilobed posterior dilation, posterior filament length 20–22 µm, CF=6–7 (Fig. 7C). Labial palpus reduced, with one-branched terminal tuft of setae, three setae and one leaf-shaped basal sensillum (Figs 7D, 8B), but well developed in five specimens (Figs 7E, 8C). Maxillary palpus with two tapering sensilla, subequal in length (Fig. 7F).
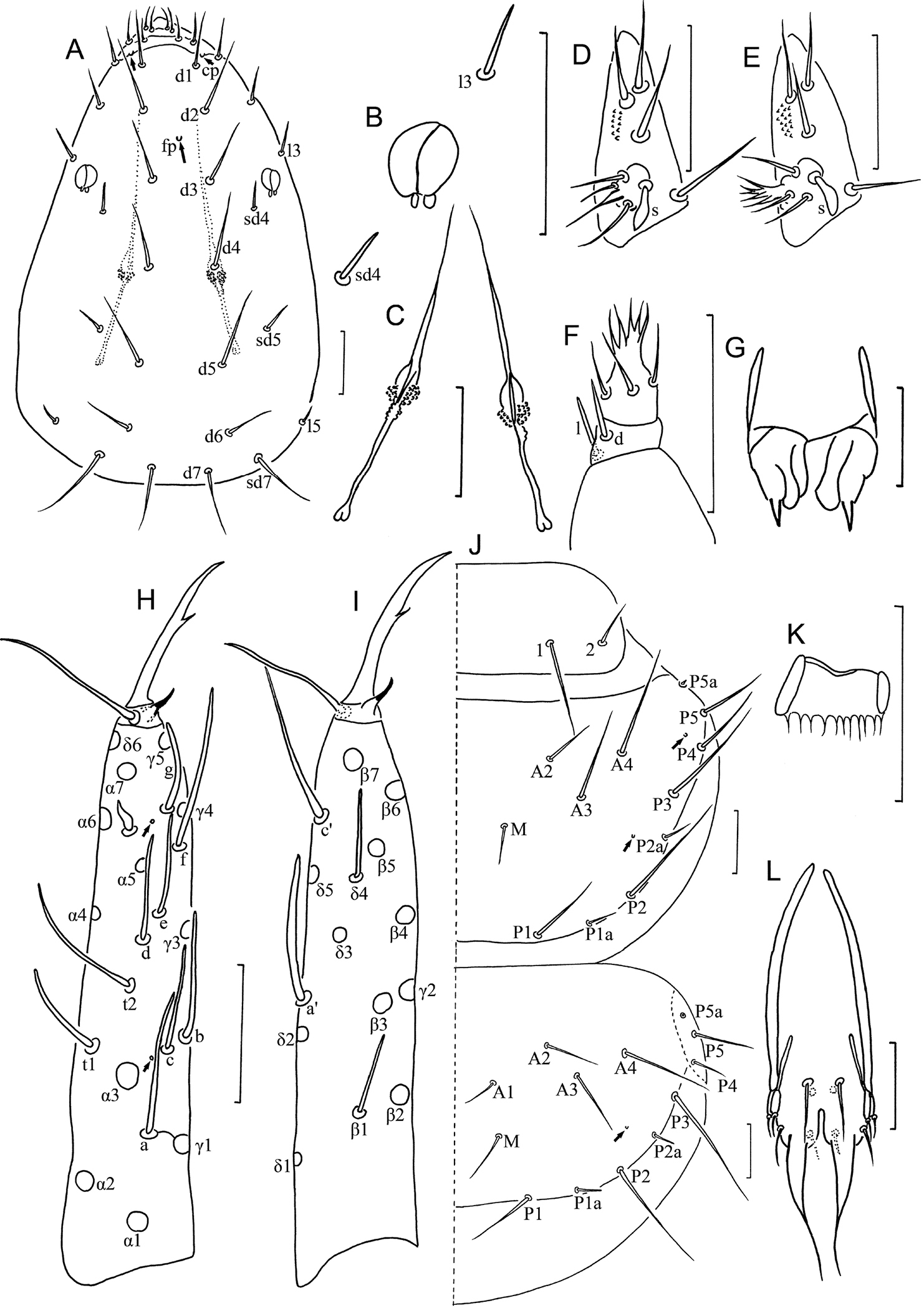
Imadateiella sharovi (Martynova, 1977) A Head, dorsal view B pseudoculus C canal of maxillary gland D labial palpus (specimens from samples 24 and 27) E labial palpus (specimens from samples 26, 30 and 32) F maxillary palpus G female squama genitalis H foretarsus, exterior view I foretarsus, interior view J nota, right side K comb L male squama genitalis. A–D and F–L specimen No. FE-2011062-4; E specimen No. FE-2011060-1. Arrows indicate pores. Scale bar: 20 μm.
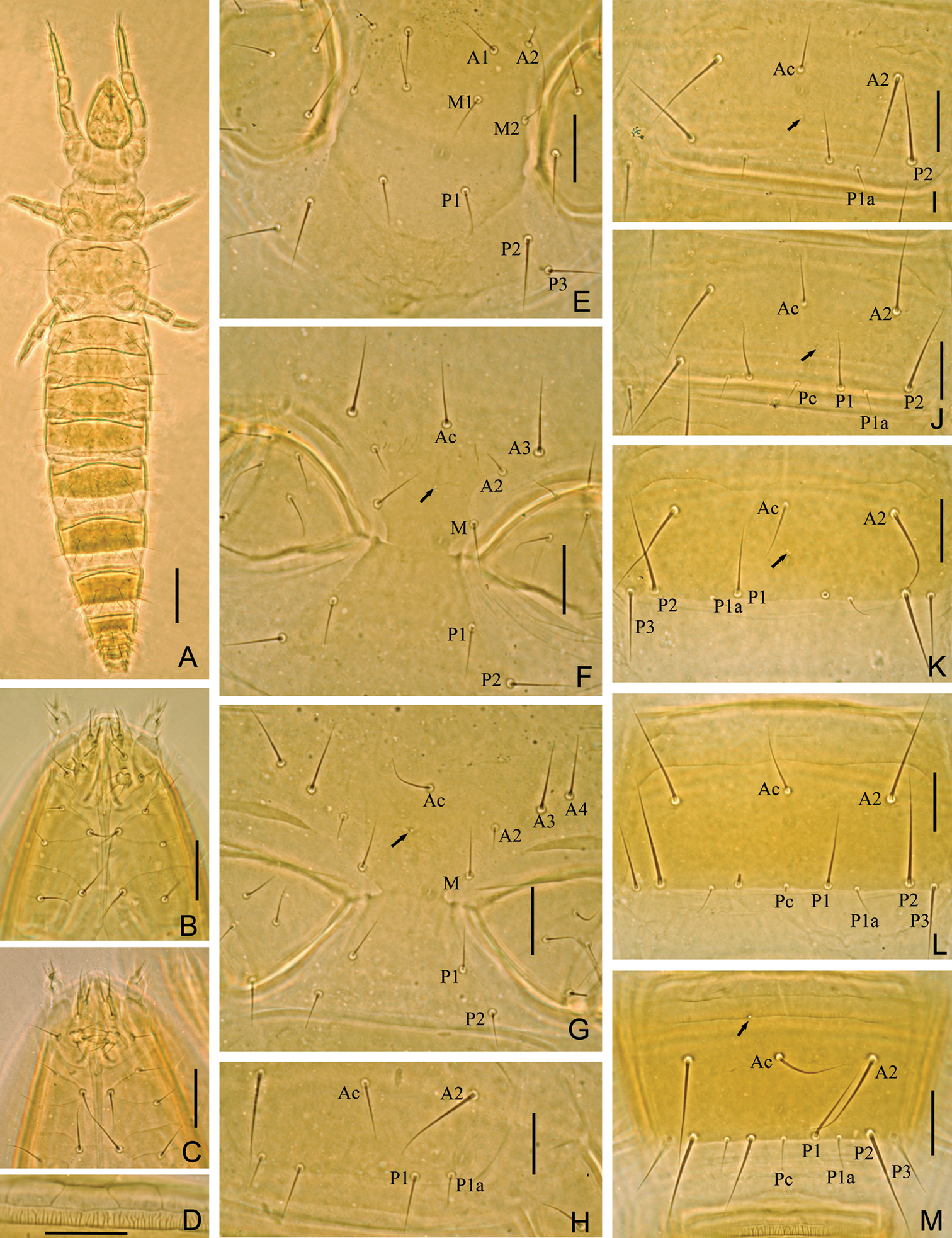
Imadateiella sharovi (Martynova, 1977) A Habitus B ventral side of head, shows reduced labial palpus C ventral side of head, shows well developed labial palpus D part of striate band E prosternum F mesosternum G metasternum H–M sternites I–III, IV, VI and VII. A–B and D–M specimen No. FE-2011062-4; C specimen No. FE-2011060-1. Arrows indicate pores. Scale bar: 100 μm in (A), others, 20 μm.
Foretarsus (Fig. 7H, I). Length 85–92 µm, claw length 23–29 µm, with one inner flap, TR=3.5–3.7; empodium length 4 µm, EU=0.15–0.18. Dorsal sensilla t-1 filiform, t-2 slender and long (17 µm), BS=0.33, t-3 lanceolate and short. All exterior sensilla slender, a far surpassing base of b and c, b slightly longer than c, c short and lower than b, d and e short, f long, g broad and short. Interior sensilla a’ broad and long, surpassing base of δ5, b’ absent, c’ slender and surpassing base of claw. Relative length of sensilla: t3 < g < t1 < (c = d = e) < b < a’ < a < t2 < c’ < f. Setae β1 and δ4 sensilliform, 12–13 µm in length. Pores close to sensilla c and t3 present. Length of middle tarsus 40 µm, claw length 15–20 µm. Length of hind tarsus 45 µm, claw length 18–20 µm.
Thorax. Thoracic chaetotaxy given in Table 4. Setae 1 and 2 on pronotum 26–35 µm and 16–17 µm length respectively, mesonotum and metanotum with eight pairs of P-setae, accessory setae sensilliform; setae P1, P1a and P2 on mesonotum 30 µm, 10 µm and 43 µm respectively (Fig. 7J). Prosternum with two pairs of anterior seta, and setae A2 and M2 sensilliform (Fig. 8E). Mesosternum with 5 A-setae, and metasternum each with 7 A-setae, and setae A2 sensilliform (Fig. 8F, G). Pronotum and prosternum without pores (Figs 7J, 8E). Mesonotum with pores sl and al, metanotum with pores sl only (Fig. 7J). Mesosternum and metasternum each with single medial pore (Fig. 8F, G). Single membranal pore present on membrane between each coxa and the body.
Adult chaetotaxy of Imadateiella sharovi (Martynova, 1977).
| Segment | Dorsal | Ventral | |||
|---|---|---|---|---|---|
| Formula | Setae | Formula | Setae | ||
| Th. | I | 4 | 1, 2 | (4+4)/6 | A1, 2, M1, 2 P1, 2, 3 |
| II | 8/16 | A2, 3, 4, M P1, 1a, 2, 2a, 3, 3a, 4, 5 |
(5+2)/4 | Ac, 2, 3, M P1, 2 |
|
| III | 10/16 | A1, 2, 3, 4, M P1, 1a, 2, 2a, 3, 3a, 4, 5 |
(7+2)/4 | Ac, 1, 2, 3, M P1, 2 |
|
| Abd. | I | 6/12 | A1, 2, 5 P1, 1a, 2, 2a, 3, 4 |
3/4 | Ac, 2 P1, 1a |
| II | 10/16 | A1, 2, 3, 4, 5 P1, 1a, 2, 2a, 3, 4, 4a, 5 |
3/5 | Ac, 2 Pc, 1a, 2 |
|
| III | 10/16 | A1, 2, 3, 4, 5 P1, 1a, 2, 2a, 3, 4, 4a, 5 |
3/6 | Ac, 2 P1, 1a, 2 |
|
| IV–V | 10/16 | A1, 2, 3, 4, 5 P1, 1a, 2, 2a, 3, 4, 4a, 5 |
3/8 | Ac, 2 P1, 1a, 2, 3 |
|
| VI | 10/16 | A1, 2, 3, 4, 5 P1, 1a, 2, 2a, 3, 4, 4a, 5 |
3/9 | Ac, 2 Pc, 1, 1a, 2, 3 |
|
| VII | 8/16 | A2, 3, 4, 5 P1, 1a, 2, 2a, 3, 4, 4a, 5 |
3/9 | Ac, 2 Pc, 1, 1a, 2, 3 |
|
| VIII | 6/15 | A1, 3, 5 Mc, 2, 3, 4, P2, 3, 4, 5 |
4/2 | 1, 2 1a |
|
| IX | 12 | 1, 1a, 2, 2a, 3, 4 | 4 | 1, 2 | |
| X | 10 | 1, 2, 2a, 3, 4 | 4 | 1, 2 | |
| XI | 6 | 1, 3, 4 | 6 | 1, 2, 3 | |
| XII | 9 | 6 | |||
Abdomen. Abdominal chaetotaxy given in Table 4. Tergite I with three pairs of anterior setae (A1, A2, A5) and six pairs of posterior setae. Tergites II–VII with eight pairs of posterior setae, P3a present. Tergite VII with four pairs of anterior setae (A2, A3, A4, A5). Tergite VII with nine pairs of posterior setae. All accessory setae on tergites I–VII sensilliform, 14–15 µm on tergites I–VI and 17 µm on VII. Tergite VIII with setae Mc). Sternite IV–V each with three anterior setae and eight posterior setae (Fig. 8K), VI–VII each with three anterior setae and nine posterior setae, Pc present (Fig. 8L, M). Sternite VIII with 4/2 setae. Hind margin of tergite IX smooth. Hind margin of tergites X–XI with very short fine ciliation. Hind margin of sternites IX–XI smooth. Posterior margin of tergite XII smooth, sternite with delicate serration in central part.
Tergites I and VIII with pores psm only, II–VII with pores psm and al, IX–XI without pores, XII with single medial pore. Sternites I and VI without pores (Fig. 8H, L), II–V each with single medial pore (Fig. 8I, J), VII with single anterior pore asymmetrical located left or right on the line (Fig. 8M), VIII–XI without pores, XII with 1+1 pores al.
Abdominal appendages I, II, III with 2, 1, 1 segments and 4, 2, 2 setae respectively. On appendages II and III, subapical seta 16–18 µm, apical seta 12–15 µm in length. Striate band on abdominal segment VIII well developed, anterior margin regular wave shaped (Fig. 8D). Comb on abdomen VIII rectangular, with 10–12 teeth (Fig. 7K). Female squama genitalis with short basal apodeme and pointed acrostyli (Fig. 7G). Male squama genitalis with 4+4 setae on dorsal side and 2+2 setae on ventral side (Fig. 7L).
Chaetal variability. Chaetal variations were observed in 7 specimens: on tergite II, absence of P4a (No. FE-2011067-2); on tergite IV, asymmetrical absence of A1 of right side (No. FE-2011060-1); on sternite II, absence of Pc and present of P1 on right side (No. FE-2011060-1, Fig. 70); on sternite III, present of Pc (No. FE-2011060-1, Fig. 71); on sternite VI, absence of Pc (Nos. FE-2011067-2, FE-2011088-2, FE-2011089-1); on sternite VIII, asymmetrical absence of P1 of left side (Nos. FE-2011062-3, FE-2011062-5, FE-2011069-1), or both side (No. FE-2011060-1).
Russia (Far East, Magadan Oblast; Khabarovsk Krai).
Imadateiella sharovi (Martynova, 1977) is characterized by the presence of setae Pc on sternite VI and 4/2 setae on sternite VIII.
Imadateiella sharovi (Martynova, 1977) is the first species of Protura described in RFE. We give the redescription of Imadateiella sharovi basing on our vast material since it shows minor differences from the redescription of
3 females, locality 4, 10-IX-2011, coll. Y. Bu, C. W. Huang, M. Potapov & V. Alpatov.
Northeast China; Russia (Far East: Khabarovsk Krai; Primorsky Krai). For Russia, it was already recorded from Khabarovsk Krai by
2 females, 2 males, locality 3, 25-IV-2010, E. Sokolova & M. Potapov collected; 2 females, 5 males, locality 5, 16-IX-2011, coll. Y. Bu, C. W. Huang, M. Potapov & V. Alpatov; 36 females, 39 males, 5 male preimagos, locality 7, 22-IX-2011, coll. Y. Bu, C. W. Huang, M. Potapov & V. Alpatov.
Widely distributed in eastern areas of Asia: China (Anhui, Taiwan, Xianggang, Yunnan, Zhejiang); Japan; Korea; Russia (Far East: Khabarovsk Krai: Khekhtsyr Range; Primorsky Krai: Shkotovsky area). New for Russia.
1 female, 1 male, locality 5, 16-IX-2011, 12 females, 7 males, locality 7, 22-IX-2011, coll. Y. Bu, C. W. Huang, M. Potapov & V. Alpatov.
Korea; Russia (Far East: Primorsky Krai). New for Russia.
3 females, 4 males, 2 maturi juniors, 1 prelarva, locality 4, 10-IX-2011, coll. Y. Bu, C. W. Huang, M. Potapov & V. Alpatov.
Russia (Far East: Primorsky Krai). It was described from Partizansky, Shkotovsky and Khasansky areas by
1 female, locality 7, 21-IX-2011. coll. Y. Bu, C. W. Huang, M. Potapov & V. Alpatov.
Japan; Russia (Far East: Primorsky Krai). New for Russia.
Body large and robust, 1700 µm in length. Foretarsus length 115 µm, setae β1 and δ4 sensilliform, 5 µm in length. Setae d6 present on head, 16 µm. Clypeal pore cp and frontal pore fp present. Pronotum and prosternum without pores, mesonotum with pores sl and al, metanotum with pores sl. Mesosternum and metasternum each with 1 medial pore. Tergites I and VIII with pores psm only, II–V with pores psm and al, VI–VII with pores psm, al, and psl. X–XI without pores, XII with single medial pore. Pores on sternites I–IV and VII not observed. Sternite V with 1 posterior pore. Sternite VI with 4 posterior pores composed by two groups, each group contains two close adjacent pores. VIII–XI without pores, XII with 1+1 pores al.
1 female, 2 maturi juniors, locality 5, 16-IX-2011. coll. Y. Bu, C. W. Huang, M. Potapov & V. Alpatov.
Japan; Russia (Far East: Primorsky Krai). New for Russia.
1 female, 2 maturi juniors, locality 4, 9-IX-2011, coll. Y. Bu, C. W. Huang, M. Potapov & V. Alpatov; 5 females, 2 males, 3 maturi juniors, 2 Larvae II, locality 2, 30-ix-2011, coll. M. Potapov & V. Alpatov.
Widely distributed in Asia: China (Anhui, Hebei, Jilin, Shanxi, Zhejiang); Japan; Korea; Russia (Far East: Primorsky Krai; Khabarovsk Krai). New for Russia.
1 female, 1 male, locality 2, sample No. 29, 30-IX-2011, coll. M. Potapov & V. Alpatov.
Northeast China; Japan; Russia (Far East: Khabarovsk Krai). New for Russia.
4 females, 2 males, 2 maturi juniors, 1 Larva LII, locality 3, 20-IX-2009. coll. O. Smirnova; 2 females, 1 larva I, 1 larva II, locality 3, 24-IV-2010, coll. E. Sokolova & M. Potapov; 2 females, 3 males, 2 larvae II, locality 4, 10-IX-2011; 3 females, 1 males, 1 maturus junior, locality 5, 16-IX-2011; 5 females, 4 males, 3 maturi juniors, 3 larvae II, 1 males preimago, locality 7, 22-IX-2011. coll. Y. Bu, C. W. Huang, M. Potapov & V. Alpatov.
Northeast China; Japan; Korea; Russia (Far East: Primorsky Krai; Khabarovsk Krai). It was reported from Shkotovsky and Khasansky areas by
5 females, locality 7, 22-IX-2011, coll. Y. Bu, C. W. Huang, M. Potapov & V. Alpatov.
Northeast China; Russia (Far East: Primorsky Krai, Lazovsky area). It was reported from Shkotovsky and Khasansky areas by
4 females, locality 4, 10-IX-2011, coll. Y. Bu, C. W. Huang, M. Potapov & V. Alpatov.
Russia (Far East, Primorsky Krai). It was recorded from Ussuriysky, Khasansky and Shkotovsky areas by
10 females, 9 males, locality 3, 20-IX-2009. coll. O. Smirnova; 2 females, 1 male, locality 3, 24-IV-2010, E. Sokolova & M. Potapov collected; 1 female, locality 4, 10-IX-2011, Y. Bu, C. W. Huang, M. Potapov & V. Alpatov collected.
Japan; Korea; Russia (Far East: Khabarovsk Krai; Prmorsky Krai). It was already recorded from Ussuriysky and Shkotovsky areas by
The description of the new material from Russian Far East is given above.
2 males, locality 3, 20-IX-2009, coll. O. Smirnova; 3 females, 2 males, locality 2, 30-IX-2011, coll. M. Potapov & V. Alpatov.
Russia (Far East, Khabarovsk Krai). New for Russia.
The present species is nearly identical to Nipponentomon bidentatum Nakamura, 2004 found from Korfovsky area of Khabarovsk Krai, they have the same shape of sensilla on foretarsus and the body chaetotaxy (seta P3a absent on tergites II–VII and A3 present on tergite I), our form only differs in the presence of seta d6 on head and 1 inner flap on the claw. Final decision on the status of our population calls for the additional study.
2 males, locality 5, 16-IX-2011, coll. Y. Bu, C. W. Huang, M. Potapov & V. Alpatov.
Russia (Far East, Primorsky Krai). New for Russia.
The present species has nearly the same body chaetotaxy (with P2a’ on mesonotum and metanotum) and the shape of sensilla on foretarsus as in Nipponentomon heterothrixi described from Northeast China, but differs by presence of setae P3a on tergites II–V which are absent in Nipponentomon heterothrixi. Insufficient material does not allow describing a new species.
2 females, 2 males, locality 7, 21-IX-2011, coll. Y. Bu, C. W. Huang, M. Potapov & V. Alpatov. 1 female, 1 male, locality 2, 30-IX-2011, coll. M. Potapov & V. Alpatov.
Russia (Far East, Khabarovsk Krai; Primorsky Krai ). New for Russia.
The present species was described from Korfovsky area of Khabarovsk Krai by
3 females, locality 4, 10-IX-2011, coll. Y. Bu, C. W. Huang, M. Potapov & V. Alpatov.
Russia (Far East, Primorsky Krai). It was recorded from Shkotovsky area by
1 female, locality 5, 16-IX-2011; 2 females, locality 7, 21-IX-2011. coll. Y. Bu, C. W. Huang, M. Potapov & V. Alpatov.
Russia (Far East, Primorsky Krai). It was already recorded from Partizansky and Khasansky areas by
1 male, locality 3, 2-X-2009, coll. O. Smirnova; 5 females, 1 male, 1 larva I, and 1 larva II, locality 4, 10-IX-2011, coll. Y. Bu, C. W. Huang, M. Potapov & V. Alpatov.
Russia (Far East: Khabarovsk Krai; Primorsky Krai). It is already recorded from Khabarovsk Krai by
1 female, locality 4, 10-IX-2011; 1 female, 1 male, 1 larva I, locality 7, 21-IX-2011, 1 female, 1 male, locality 7, 22-IX-2011; coll. Y. Bu, C. W. Huang, M. Potapov & V. Alpatov.
Widely distributed in China. Russia (Far East, Primorsky Krai). New for Russia.
4 females, 1 male, locality 7, 22-IX-2011, coll. Y. Bu, C. W. Huang, M. Potapov & V. Alpatov.
Widely distributed in China; Russia (Far East, Primorsky Krai). New for Russia.
The 31 species of Protura recorded from RFE so far are listed in Table 5. They belong to 12 genera and 4 families Berberentulidae, Acerentomidae, Nipponentomidae and Eosentomidae, half the species are known only from the RFE. The most species are representatives of Acerentomidae and Nipponentomidae, each with 11 species (70%). The family Eosentomidae is rare and consists of only 3 species. Palearctic genera Yamatentomon, Imadateiella, Yichunentulus, Callientomon and Holarctic genera Filientomon, Fjellbergella, Nipponentomon, Tuxenentulus, Verrucoentomon are the dominant taxa composing 66% of all species.
| Classification | Species | RFE | China | Korea | Japan |
|---|---|---|---|---|---|
| Acerentomata Yin, 1996 | |||||
| Berberentulidae Yin, 1983 | |||||
| Baculentulus Tuxen, 1977 | Baculentulus krabbensis sp. n. |
5 |
|||
| Baculentulus loxoglenus Yin, 1980 | 3, 4 | + | |||
| Baculentulus morikawai (Imdadaté & Yosii, 1956) | 3, 5, 7 | + | + | + | |
| Baculentulus pomorskii Shrubovych, 2010 |
4, 5, 6 | ||||
| Baculentulus potapovi Shrubovych, 2010 |
4, 5, 6 | ||||
| Baculentulus samchonri Imdadaté & Szeptycki, 1976 | 5, 7 | + | |||
| Acerentomidae Silvestri, 1907 | |||||
| Acerentulus Berlese, 1908 | Acerentulus kisonis Imdadaté, 1961 | 7 | + | ||
| Filientomon Rusek, 1974 | Filientomon duodecimsetosum Nakamura, 2004 |
3 | |||
| Filientomon gentaroanum Nakamura, 2001 | 5 | + | |||
| Filientomon takanawanum (Imdadaté, 1956) | 2, 4 | + | + | + | |
| Fjellbergella Nosek, 1978 | Fjellbergella lazovskiensis sp. n. |
7 | |||
| Tuxenentulus Imdadaté, 1974 | Tuxenentulus ohbai Imdadaté, 1974 | 2 | + | + | |
| Tuxenentulus solncevae Shrubovych & Bernard, 2013 |
8 | ||||
| Yamatentomon Imdadaté, 1964 | Yamatentomon kunnepchupi Imadaté, 1964 | 3 | + | ||
| Yamatentomon yamato Imadaté & Yosii, 1956 | 3, 4, 5, 7 | + | + | + | |
| Yichunentulus Yin, 1980 | Yichunentulus alpatovi sp. n. |
7 | |||
| Yichunentulus borealis (Nakamura, 2004), comb. n. |
3 | ||||
| Nipponentomidae Yin, 1996 | |||||
| Callientomon Yin, 1980 | Callientomon chinensis Yin, 1980 | 4, 5, 7 | + | ||
| Imadateiella Rusek, 1974 | Imadateiella sharovi (Martynova, 1977) |
1, 2 | |||
| Nipponentomon Imdadaté & Yosii, 1959 | Nipponentomon bidentatum Nakamura, 2004 | 3 | + | ||
| Nipponentomon cf. bidentatum Nakamura, 2004 |
2, 3 | ||||
| Nipponentomon cf. heterothrixi Yin & Xie, 1993 |
5 | ||||
| Nipponentomon khabarovskense Nakamura, 2004 |
2, 3, 7 | ||||
| Nipponentomon jaceki Shrubovych, 2009 |
4 | ||||
| Nipponentomon nippon (Yoshii, 1938) | 5, 6, 7 | + | + | + | |
| Verrucoentomon Rusek, 1974 | Verrucoentomon kawakatsui Imadaté, 1964 | 3 | + | ||
| Verrucoentomon louisanne Shrubovych, 2012 |
4 | ||||
| Verrucoentomon shirampa Imadaté, 1964 | 3, 4 | + | + | ||
| Eosentomata Yin, 1996 | |||||
| Eosentomidae Berlese, 1909 | |||||
| Eosentomon Berlese, 1909 | Eosentomon asahi Imadaté, 1961 | 3, 4 | + | + | + |
| Eosentomon brevicorpusculum Yin, 1965 | 3, 7 | + | |||
| Eosentomon orientale Yin, 1965 | 7 | + |
* Known only from RFE so far.
** Numbers indicate the localities given in the materials and methods.
Compared with neighbouring regions, the Protura fauna of RFE is closely related to the fauna of Northeast China, Korea, and Japan, sharing 11, 7, and 11 species with each respectively (
Based on the distributional data available so far, the species of Protura recorded in RFE can be classified into three groups:
1) Species widespread in the Eastern Asia including both its temperate and tropical parts (Fig. 9): Baculentulus morikawai (Imdadaté & Yosii, 1956), Filientomon takanawanum (Imdadaté, 1956), Eosentomon brevicorpusculum Yin, 1965 and Eosentomon orientale Yin, 1965.

Distribution of two widely distributed East-Asiatic species of RFE. In addition to locations listed in the text, other records are used after
2) Temperate East Palearctic species: Baculentulus loxoglenus Yin, 1980, Baculentulus samchonri Imdadaté & Szeptycki, 1976, Acerentulus kisonis Imdadaté, 1961, Filientomon gentaroanum Nakamura, 2001, Tuxenentulus ohbai Imdadaté, 1974, Yamatentomon yamato Imadaté & Yosii, 1956, Yamatentomon kunnepchupi Imadaté, 1964, Callientomon chinensis Yin, 1980, Nipponentomon bidentatum Nakamura, 2004, Nipponentomon nippon (Yoshii, 1938), Verrucoentomon kawakatsui Imadaté, 1964, Verrucoentomon shirampa Imadaté, 1964, and Eosentomon asahi Imadaté, 1961. The species are recorded in RFE, Northeast China, Korea and temperate part of Japan (Fig. 10).
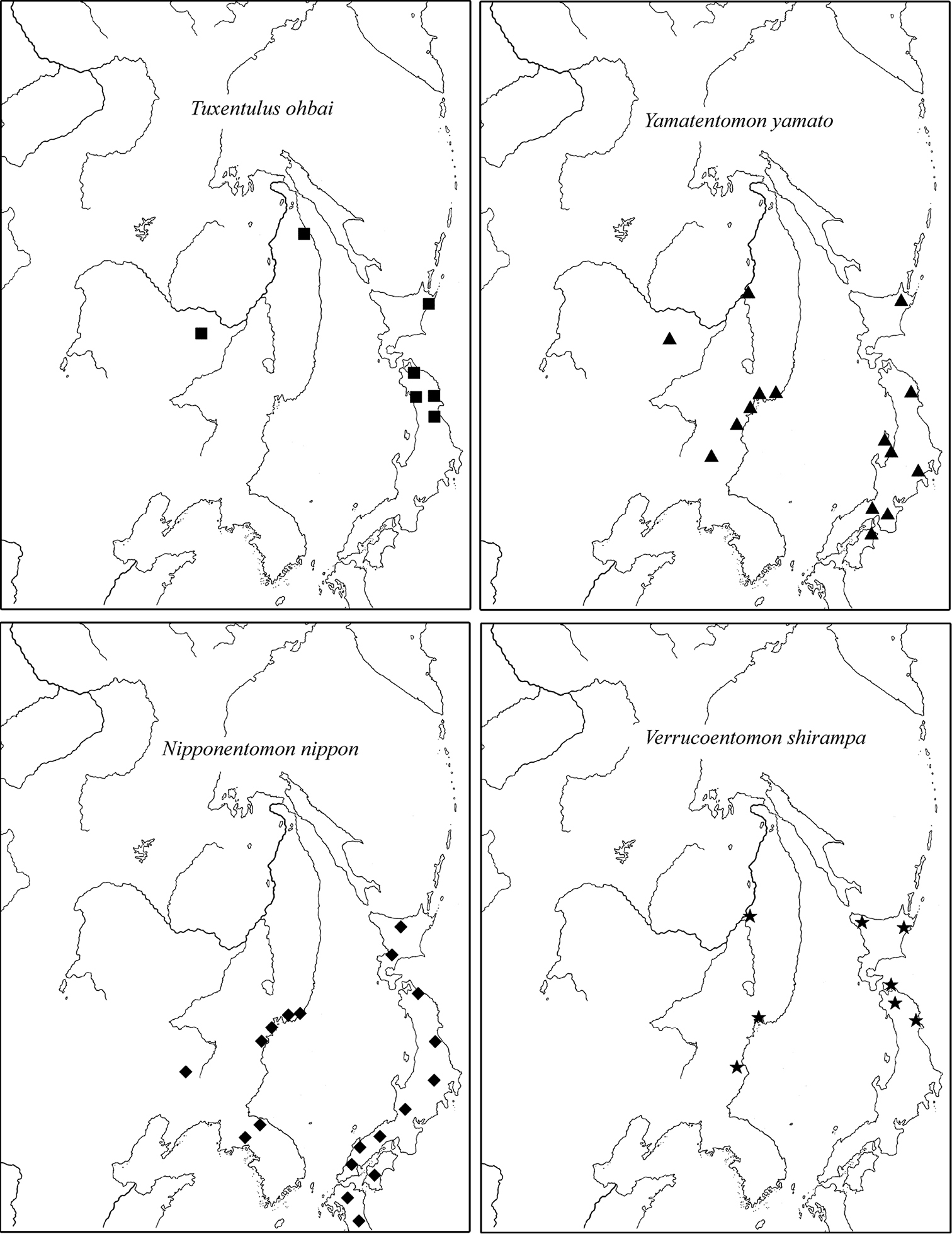
Distribution of typical East-Palearctic species of RFE. In addition to locations listed in the text, other records are used after
3) Local species: Baculentulus krabbensis sp. n., Baculentulus pomorskii Shrubovych, 2010, Baculentulus potapovi Shrubovych, 2010, Fjellbergella duodecimsetosum Nakamura, 2004, Fjellbergella lazovskiensis sp. n., Tuxenentulus solncevae Shrubovych & Bernard, 2013, Yichunentulus alpatovi sp. n., Yichunentulus borealis (Nakamura, 2004), comb. n., Imadateiella sharovi (Martynova, 1977), Nipponentomon cf. bidentatum Nakamura, 2004, Nipponentomon cf. heterothrixi Yin & Xie, 1993, Nipponentomon khabarovskense Nakamura, 2004, Nipponentomon jaceki Shrubovych, 2009, Verrucoentomon louisanne Shrubovych & Bernard 2012 and Eosentomon sp. Nakamura, 2004. So far they have been recorded only in the south of RFE apart from Imadateiella sharovi distributed wider penetrating to the northern part of RFE (Fig. 11).
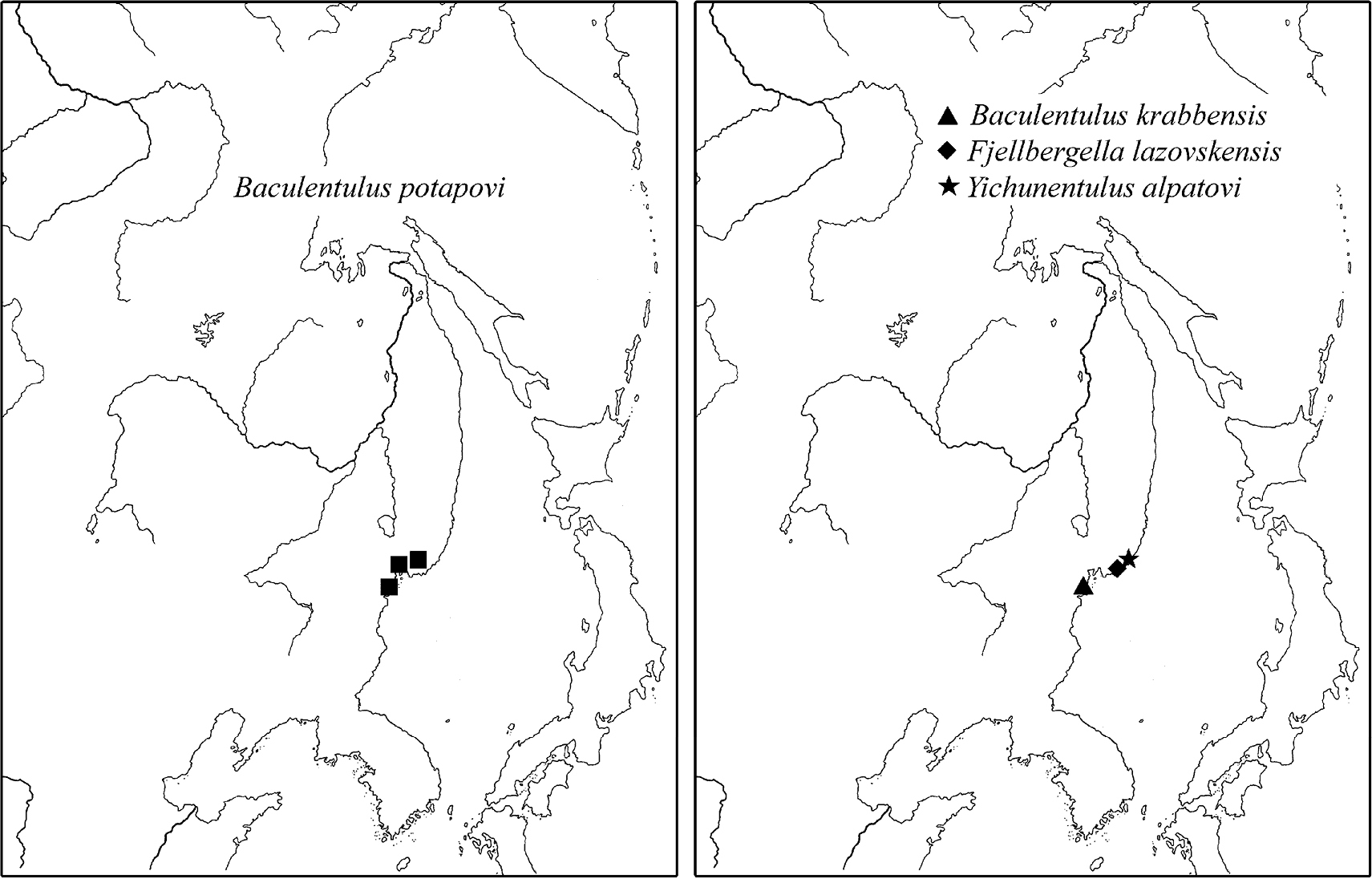
Distribution of some local species of RFE.
The biogeographical composition of Russian Far East, including Primorsky Krai and Khabarovsk Krai, has been described for many arthropods (
The portion of Holarctic and trans-Palearctic species in fauna of arthropods of Primorsky Krai.
| Taxa | Holarctic and trans-Palearctic species | References |
|---|---|---|
| Geometridae (Insecta: Lepidoptera) | ~28% | |
| Formicidae (Insecta: Hymenoptera) | ~14% | |
| Pompilidae (Insecta: Hymenoptera) | ~30% | |
| Syrphidae (Insecta: Diptera) | ~37% | |
| Spheciformes (Insecta: Hymenoptera) | ~45% | |
| Apiformes (Insecta: Hymenoptera) | ~43% | |
| Orthoptera (Insecta) | ~14% | |
| Plecoptera (Insecta) | ~10% | |
| Oribatei (Arachnida) | ~55% | |
| Protura (Hexapoda) | 0% | this study |
Only three widely distributed species of Protura are known so far: Acerentulus confinis (Berlese, 1908), Berberentulus capensis (Womersley, 1931), and Gracilentulus gracilis (Berlese, 1908). These species are cosmopolitans in the broad sense, but their distribution is not fully understood (
Several reasons can be proposed to explain the absence of the Holarctic and Palearctic species of Protura:
- High sensitivity of Protura to anthropogenic impact (
- In the Northern hemisphere, this group sharply declines at higher latitudes. In many groups of animals the portion of endemic species increases southwards in Eurasia (
- Low dispersal ability of Protura. This reason is possible but could not be substantiated since widely distributed species are known in other groups with low active movement but with high possibility of passive carriage (
In conclusion, the Protura appear to be a group with a low level of biogeographical “noise” (ruderal species), with high endemism and are candidate organisms for more detailed biogeographical analysis when more information is available for other regions.
We are grateful to Mr. C. W. Huang (China) and Dr. V. Alpatov (Russia) for their assistance in the field. We sincerely thank Dr. P. Greenslade (Australia) for her generous help in language revision. The authors are also grateful to the management and staff of State Reserve of Lazovsky who provided the collecting permit and suitable conditions for our field work, especially to A. Laptev, Yu. Sundukov, and V. Kirsanov. We would especially like to thank A. Lelej and E. Mikhaleva (Russia, Vladivostok), who’s hospitality has made our stay in Russian Far East very efficient. A. Babenko (Russia, Moscow) made several valuable critical remarks. We cordially thank Dr. O. Nakamura (Japan) and Dr. J. Shrubovych (Ukraine) for their constructive reviews of the manuscript and valuable comments. This study was supported by the National Natural Sciences Foundation of China (no: 31201706, 31301873, 31071887) and NSFC-RFBR Cooperative Research Project (31111120077 / 11-04-91179-GFENa, RFBR 14-04-91169M).