





Citation: Song X-B, Li L-Z (2014) Descriptions of three new species of the Termitophilous tribe Termitopaediini in China (Coleoptera, Staphylinidae, Aleocharinae). ZooKeys 424: 91–100. doi: 10.3897/zookeys.424.7670
Three new species belonging to two genera of the aleocharine tribe Termitopaediini Seevers, viz., Dioxeuta rara Song & Li, sp. n., D. yunnanensis Song & Li, sp. n., and Termitopulex sinensis Song & Li, sp. n. from Baihualing Natural Reserve (Southwest China: Yunnan) are described and illustrated. This is the first record of Termitopulex Fauvel, 1899 from China.
Termitopaediini, Dioxeuta, Termitopulex, termitophily, taxonomy, new species, China, Oriental region
The termitophilous tribe Termitopaediini Seevers was established by
Holotypes and most of the paratypes are deposited in the Insect Collection of the Shanghai Normal University, Shanghai, China (SNUC), and some of paratypes are deposited in the Kyushu University Museum, Fukuoka, Japan (KUM).
Specimens were killed with ethyl acetate and preserved in 75% ethanol before dissection; photos of habitus were taken by a Canon EOS 7D with an MP-E 65mm macro photo lens; photos of characteristic pattern were taken by a Canon G9 Camera mounted on an Olympus CX31 microscope.
The following abbreviations are applied in the text: BL – body length, from the anterior margin of the head to the posterior margin of the abdominal tergite VIII; FBL – forebody length, from the clypeal anterior margin to the posterior margin of elytra; HL – head length, from the clypeal anterior margin to the occipital constriction; PL – length of the pronotum along the midline; HW – width of the head across the eyes; PW – maximum width of the pronotum.
The genus is similar to Neodioxeuta Seevers and Termitopulex Fauvel by the shape of head and thorax. It can be easily separated from Neodioxeuta by the abdomen only slightly physogastric and the abdominal segment II represented by a reduced tergite only. It can be distinguished from Termitopulex by the outer paratergites about twice the width of the inner paratergites (
Holotype: China: ♂ (on slide), labeled ‘CHINA: Yunnan, Baoshan City, Mangkuan Town (芒宽乡), Baihualing N. R. (百花岭), 25°17'47"N, 98°48'22"E, alt. 1400 m, 23-IV-2013, Xiao-Bin Song leg. / HOLOTYPE [red], Dioxeuta rara sp. n., Song & Li det. 2014, SNUC’. Paratype: China: 1 ♀, same data as holotype, bearing the following label: ‘PARATYPE [yellow], Dioxeuta rara sp. n., Song & Li det. 2014, SNUC’.
Dioxeuta rara is most similar to Dioxeuta rougemonti and Dioxeuta yunnanensis described below through its yellowish-brown color and the presence of macrochaetae on abdominal tergite VII. The new species can be readily separated from Dioxeuta rougemonti by the deferent abdominal macrochaetotaxy as well as the shape of aedeagal median lobe. It differs from Dioxeuta yunnanensis by the presence of short macrosetae on abdominal tergites VII; tergite VIII with 5 pairs of macrosetae; sternite VIII almost truncate at apex and the deferent shapes of aedeagal median lobe and spermatheca.
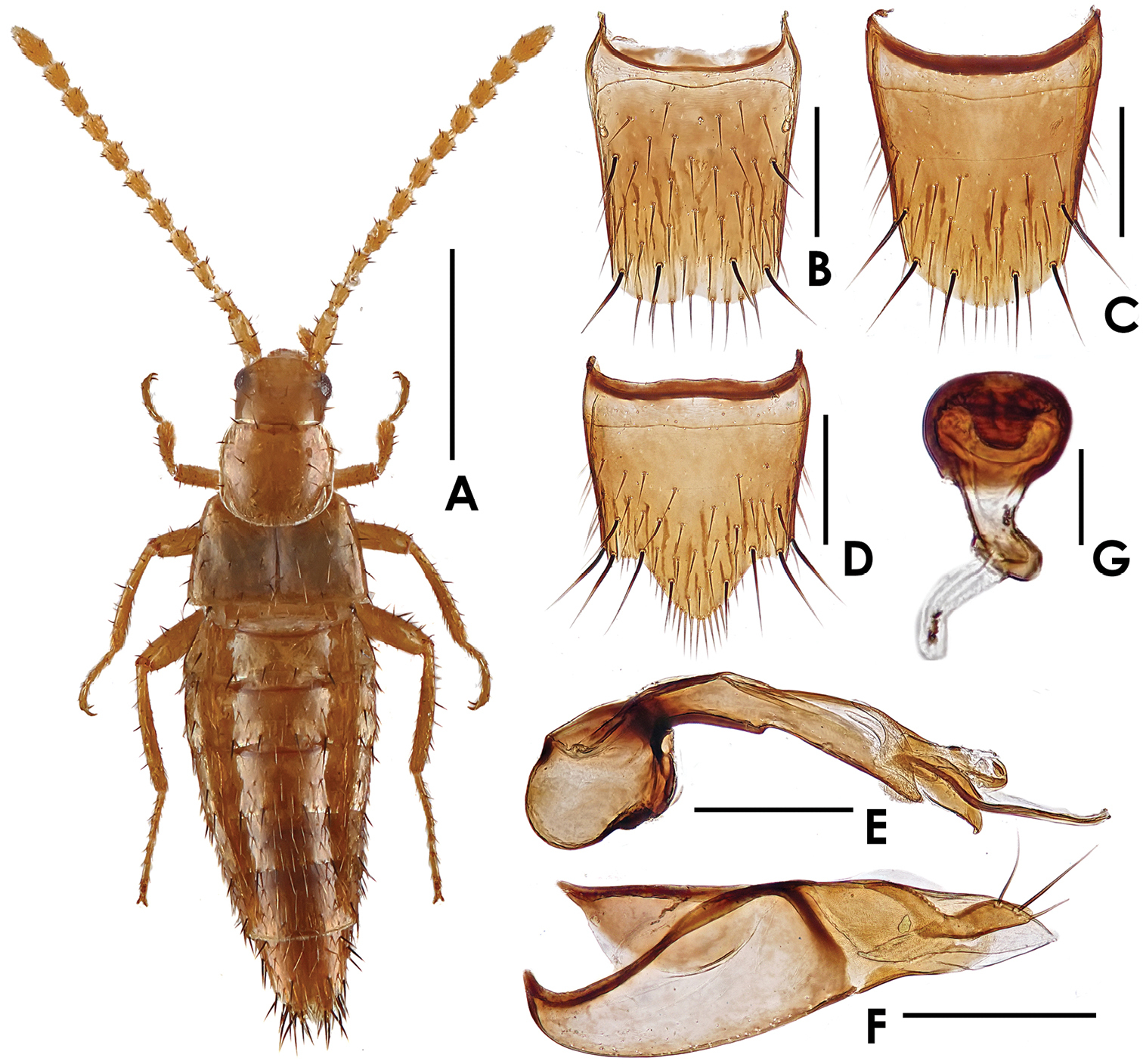
Body (Fig. 1A) shining, smooth. Coloration: fore body and legs yellowish-brown; antennae yellowish-brown to brown; abdomen yellowish-brown, with mesal area of tergites brown.

Dioxeuta rara sp. n. A holotype, female habitus B male tergite VII C female tergite VII D male tergite VIII E female tergite VIII F male sternite VIII G female sternite VIII. Scales (mm): A = 1; B–G = 0.2.
Head (Fig. 1A) subquadrate, slightly longer than wide, about 0.89 times as wide as long; sparsely covered with long setae. Pronotum (Fig. 1A) slightly wider than long, about 1.05 times as wide as long; with 7 pairs of macrosetae, with a row of 4 macrosetae along the anterior margin, 6 on disc and 2 each along the lateral margins. Elytra (Fig. 1A) wider than long; disc sparsely covered with macrosetae; hypomera with several long yellow setae along lateral margin. Abdomen slightly physogastric, widest at segments III–V; abdominal tergite VII shaped as in Figs 1B–C, sparsely covered with long yellow setae, posterior margin with a row of 8 very short setae; tergite VIII (Figs 1D–E) with 5 pairs of macrosetae. Macrochaetotaxy of abdominal tergites II–VIII: 4, 6, 6, 6, 6, 1–2 (short), 10; inner paratergites without macrosetae but sparsely covered with yellow setae; macrochaetotaxy of outer paratergites III–VII variable: 6–7, 5–7, 4–5, 2–3, 1.
Male. Sternite VIII almost truncate at apex, shaped as in Fig. 1F; with 5 pairs of macrosetae. Median lobe of aedeagus (Fig. 2A) with apical lobe distinctly curved; paramere shaped as in Fig. 2B.
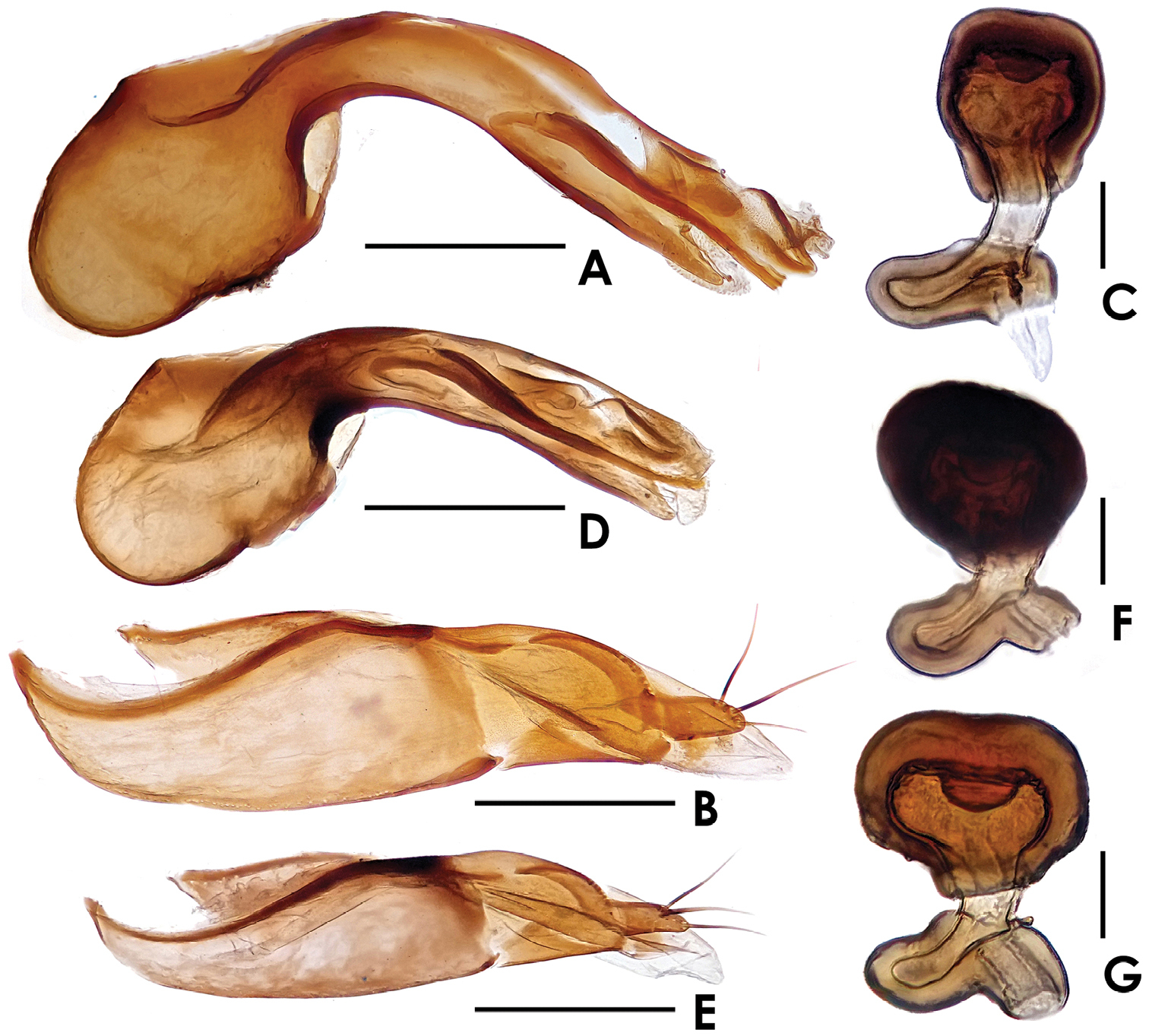
Dioxeuta rara sp. n. A median lobe of aedeagus, in lateral view B paramere C spermatheca Dioxeuta yunnanensis sp. n. D median lobe of aedeagus, in lateral view E paramere F spermatheca G ditto. Scales (mm): A, B, D, E = 0.2; C, F, G = 0.03.
Female. Sternite VIII slightly emarginate at apex, shaped as in Fig. 1G; with 5 pairs of macrosetae. Spermatheca shaped as in Fig. 2C.
Male: BL: 2.63; FBL: 1.06; HL: 0.38; HW: 0.35; PL: 0.41; PW: 0.45; HW/HL: 0.92; PW/PL: 1.05; HW/PW: 0.78. Female: BL: 2.65; FBL: 1.10; HL: 0.40; HW: 0.35; PL: 0.40; PW: 0.43; HW/HL: 0.88; PW/PL: 1.05; HW/PW: 0.81.
Southwest China: Yunnan.
Macrotermes sp.
The Latin adjective rāra means ‘rare’.
Holotype: China: ♂, labelled ‘CHINA: Yunnan, Baoshan City, Mangkuan Town (芒宽乡), Baihualing N. R. (百花岭), 25°17'47"N, 98°48'22"E, alt. 1400 m, 23-IV-2013, Xiao-Bin Song leg. / HOLOTYPE [red], Dioxeuta yunnanensis sp. n., Song & Li det. 2014, SNUC’. Paratypes: China: 1 ♂, 9♀♀, same data as holotype, bearing the following label: ‘PARATYPE [yellow], Dioxeuta yunnanensis sp. n., Song & Li det. 2014, SNUC’. (SNUC, KUM).
Dioxeuta yunnanensis is most similar to Dioxeuta rara described above, but can be readily separated from it by the different abdominal macrochaetotaxy as well as the shape of aedeagal median lobe and spermatheca.
Body (Figs 3A, 5B) shining, smooth. Coloration: fore body and legs yellowish-brown; antennae yellowish-brown to brown; abdomen yellowish-brown, with mesal area of tergites brown.

Dioxeuta yunnanensis sp. n. A male habitus B tergite VII C tergite VIII D ditto E male sternite VIII F female sternite VIII G ditto. Scales (mm): A = 1; B–G = 0.2.
Head (Fig. 3A) subquadrate in form; slightly longer than wide, about 0.86 times as wide as long; sparsely covered with long setae. Pronotum (Fig. 3A) slightly wider than long, about 1.02 times as wide as long; with 7 pairs of macrosetae, with a row of 4 macrosetae along the anterior margin, 6 on disc and 2 each along the lateral margins. Elytra (Fig. 3A) wider than long; disc sparsely covered with macrosetae; hypomera with several short yellow setae along lateral margin. Abdomen slightly physogastric, widest at segments III–V; abdominal tergite VII shaped as in Figs 3B; tergite VIII (Figs 3C, E) with 4–5 macrosetae, posterior margin with a row of 6–8 long yellow setae. Macrochaetotaxy of abdominal tergites II–VIII: 2, 6, 6, 6, 4–6, 0, 4–5; inner paratergites without macrosetae but sparsely covered with yellow setae; macrochaetotaxy of outer paratergites III–VII is variable: 5–8, 4–6, 3–4, 0, 0.
Male. Sternite VIII rounded at apex, shaped as in Fig. 3D; with 4 pairs of macrosetae. Median lobe of aedeagus (Fig. 2D) with apical lobe curved; paramere shaped as in Fig. 2E.
Female. Posterior margin of sternite VIII (Fig. 3F–G) nearly rounded; with 7–8 macrosetae. Spermatheca shaped as in Fig. 2F–G.
Male: BL: 2.34–2.46; FBL: 0.95–1.06; HL: 0.35; HW: 0.30; PL: 0.38; PW: 0.38; HW/HL: 0.87; PW/PL: 1.00; HW/PW: 0.79. Female: BL: 2.61–2.72; FBL: 1.02–1.06; HL: 0.35–0.38; HW: 0.32; PL: 0.37–0.40; PW: 0.39–0.41; HW/HL: 0.84–0.87; PW/PL: 1.03–1.04; HW/PW: 0.78–0.82.
Southwest China: Yunnan.
Macrotermes sp.
Named after its type locality of Yunnan Latinized.
The genus is similar to Polyteinia Bernhauer and Paratermitopulex Kistner through the overall shape. It can be easily separated from Polyteinia by the absence of easily visible exit pores from abdominal segment VII and the different shape of paramere. It can be distinguished from Paratermitopulex by the rounded lateral edges of the pronotum. Termitopulex is also related to Dioxeuta from which it can be distinguished the less physogastric abdomen and the paratergites approximately equal in width (
Holotype: China: ♂, labelled ‘CHINA: Yunnan, Baoshan City, Mangkuan Town (芒宽乡), Baihualing N. R. (百花岭), 25°17'47"N, 98°48'22"E, alt. 1400 m, 23-IV-2013, Xiao-Bin Song leg. / HOLOTYPE [red], Termitopulex sinensis sp. n., Song & Li det. 2014, SNUC’. Paratypes: China: 3 ♂♂, 8♀♀, same data as holotype, bearing the following label: ‘PARATYPE [yellow], Termitopulex sinensis sp. n., Song & Li det. 2014, SNUC’. (SNUC, KUM).
Termitopulex sinensis can be readily separated from the only Asian congener Termitopulex omaniensis Kistner by the posterior margin of abdominal tergite VIII being broadly concave and the different abdominal macrochaetotaxy. The new species differs from other species of the genus by the different macrochaetotaxy of abdomen as well as the shape of aedeagus and spermatheca.
Body (Figs 4A, 5C) shining, smooth. Coloration: fore body and legs yellowish-brown; abdomen yellowish-brown, with tergites VI–VIII darker.
Termitopulex sinensis sp. n. A male habitus B tergite VIII C female sternite VIII D male sternite VIII E median lobe of aedeagus, in lateral view F paramere G spermatheca. Scales (mm): A = 1; B–F = 0.2; G = 0.03.

A over view of the symbiotic host’s nest B Living Dioxeuta yunnanensis C Living Termitopulex sinensis.
Head (Fig. 4A) subquadrate in form; slightly longer than wider, about 0.92 times as wide as long; with 4 macrosetae on disc and 1 each along the lateral margins. Mandibles with apical teeth elongate. Pronotum (Fig. 4A) slightly wider than long, about 1.06 times as wide as long; with 6 pairs of macrosetae, with a row of 4 macrosetae along the anterior margin, 4 on disc and 2 each along the lateral margins. Elytra (Fig. 4A) wider than long; disc sparsely covered with macrosetae. Abdomen widest at segments III–V; posterior margin of tergite VIII (Fig. 4B) broadly concave. Macrochaetotaxy of abdominal tergites II–VIII: 2, 4, 4, 4, 4, 2 (very short), 6; inner paratergites without macrosetae but covered with brown setae; macrochaetotaxy of outer paratergites III–VII as follow: 2, 1–2, 1–2, 0, 0.
Male. Sternite VIII rounded at apex; shaped as in Fig. 4D; with 6 pairs of macrosetae. Median lobe of aedeagus and paramere shaped as in Fig. 4E–F.
Female. Posterior margin of sternite VIII (Fig. 4C) nearly rounded; with 4 pairs of macrosetae. Spermatheca shaped as in Fig. 4G.
Male: BL: 2.08–2.31; FBL: 0.88–0.92; HL: 0.32; HW: 0.29; PL: 0.32–0.33; PW: 0.34–0.35; HW/HL: 0.92; PW/PL: 1.06–1.08; HW/PW: 0.85–0.88. Female: BL: 2.38–2.76; HL: 0.32–0.35; HW: 0.31–0.32; PL: 0.32–0.37; PW: 0.34–0.38; HW/HL: 0.89–0.96; PW/PL: 1.03–1.08; HW/PW: 0.82–0.84.
Southwest China: Yunnan.
Macrotermes sp.
Named after the type locality.
We thank Cong-Chao Dai and Zhong Peng (both Shanghai, China) for their support during the field work. Two anonymous reviewers critically read the manuscript and provided helpful advice. The study is supported by the National Natural Science Foundation of China (No. 31101659 and No. 31172134, 31201734) and Shanghai Normal University (DZL125).