





Citation: Khaustov AA (2014) A new genus and species in the mite family Eupodidae (Acari, Eupodoidea) from Crimea. ZooKeys 422: 11–22. doi: 10.3897/zookeys.422.7802
A new genus Pseudoeupodes Khaustov, gen. n. and new species Pseudoeupodes porosus sp. n. are described from moss in Crimea. The taxonomy of the Eupodidae and some other families and genera of Eupodoidea is reviewed. The genus Turanopenthalodes Barilo, 1988 is transferred from Penthalodidae to Penthaleidae. The family Cocceupodidae Jesionowska, 2010 and the genus Filieupodes Jesionowska, 2010 are considered as junior synonyms of Eupodidae Koch, 1842 and Cocceupodes Thor, 1934, respectively. A key to genera of the family Eupodidae is provided.
Acarina, Eupodoidea, Eupodidae, Pseudoeupodes, systematics, key, Crimea
Mites of the cosmopolitan superfamily Eupodoidea Koch, 1842 are fungivorous, phytophagous and predatory. The classification of the superfamily lacks stability (
This paper presents a description of a new genus and species of eupodid mite, Pseudoeupodes porosus gen. n., sp. n., collected from moss in Crimea, and discusses the taxonomy of some families and genera of Eupodoidea.
Mites were collected from moss using Berlese funnels and mounted in Hoyer’s medium. Notations for the prodorsal and leg setae follow
Pseudoeupodes porosus Khaustov, sp. n. Monotypic.
Female. Idiosomal dorsum (Figs 1, 11–13). Idiosoma oval. Cuticle soft and striated. Sejugal furrow well developed. Prodorsum with three pairs of tactile setae (v1, v2, sc2) and a pair of filiform trichobothria (sc1). Naso (epivertex) folded downward onto ventral surface of prodorsum, setae v1 situated on dorsal part of naso near anterior margin of prodorsum; naso defined by different pattern of striation from surrounding prodorsum (Fig. 11). Hysterosoma with eight pairs of dorsal setae (c1, c2, d1, e1, f1, f2, h1, h2) and three pairs of large round lyrifissures (ia, im, ip). Hysterosoma dorsally with two transverse furrows, between setae c1 and d1, and between e1 and f1. Setae f1 not trichobothrium-like.
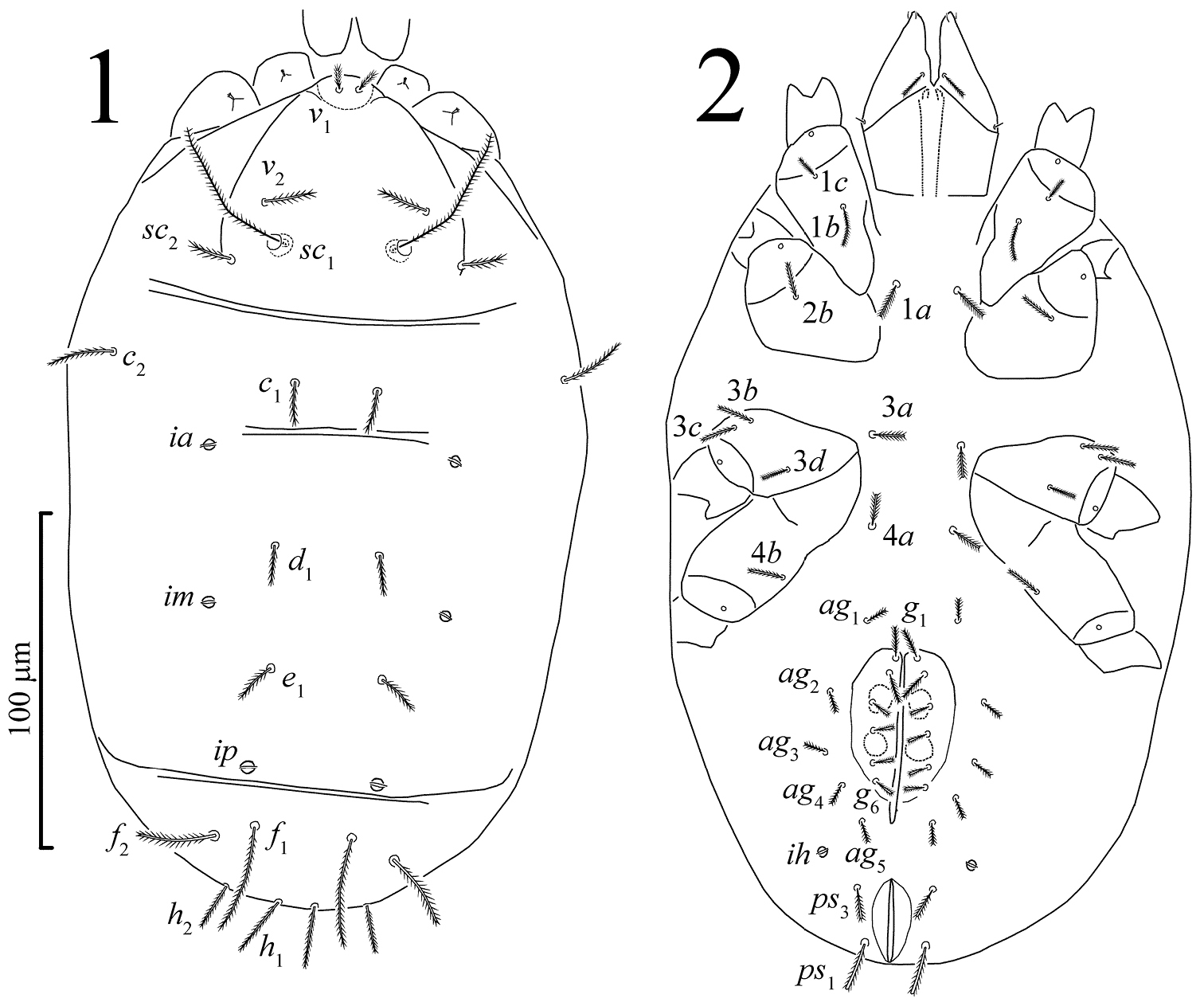
Pseudoeupodes porosus Khaustov, gen. n., sp. n., female: 1 idiosomal dorsum 2 idiosomal venter.
Idiosomal venter (Figs 2, 3, 14–15). Coxisternal setal formula 3-1-4-2; six pairs of eugenital setae; six pairs of genital setae; five pairs of aggenital setae; two pairs of pseudanal setae; one pair of lyrifissures (ih), same form as dorsals.
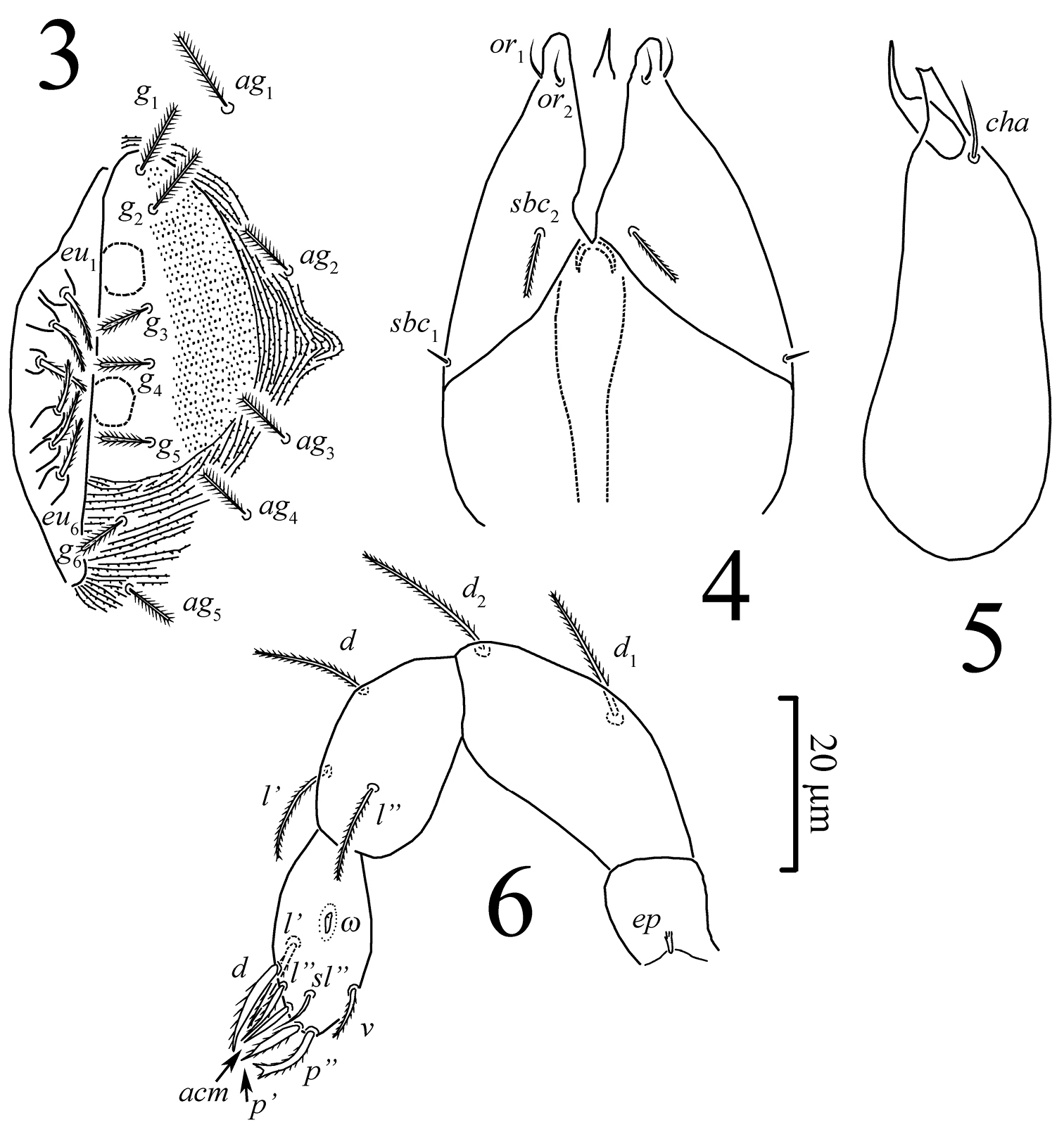
Pseudoeupodes porosus Khaustov, gen. n., sp. n., female: 3 genital area 4 subcapitulum 5 chelicera, antiaxial aspect 6 palp, antiaxial aspect.
Gnathosoma (Figs 4–6). Palp setal formula 0-2-3-8(ɷ), tarsus ovoid. Chelicerae: typical for eupodid mites, movable digit slender and acuminate distally, fixed digit distinctly shorter than movable digit and truncated distally; seta cha present.
Legs (Figs 7–10). All legs shorter than body. Soft cuticle separating coxisternal plates and trochanters of all legs with distinct pore-like structure (Fig. 14). Rhagidial organ I with two longitudinally arranged solenidia; rhagidial organ II with three longitudinally arranged rhagidial solenidia. Tarsus I with famulus (stellate setae) situated in shallow depression; tarsus II with spine-like famulus. Tibiae I and II with one distal rhagidial solenidion; tibiae I-III with proximal erect solenidion; genua I and II with one erect solenidion. Femur IV not enlarged. Trochanteral setal formula 1-1-1-1.
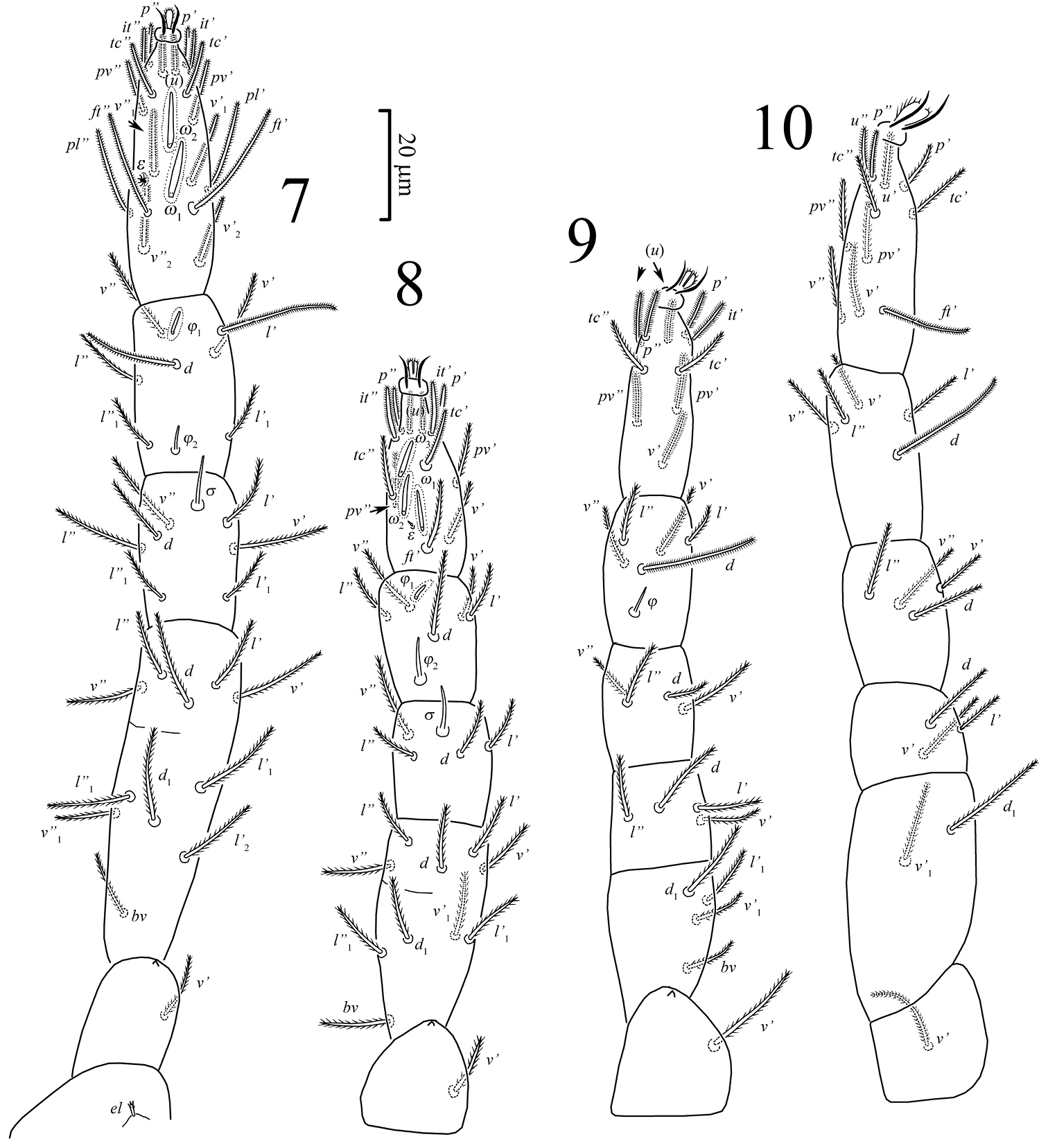
Pseudoeupodes porosus Khaustov, gen. n., sp. n., female: 7–10 legs I-IV, respectively.
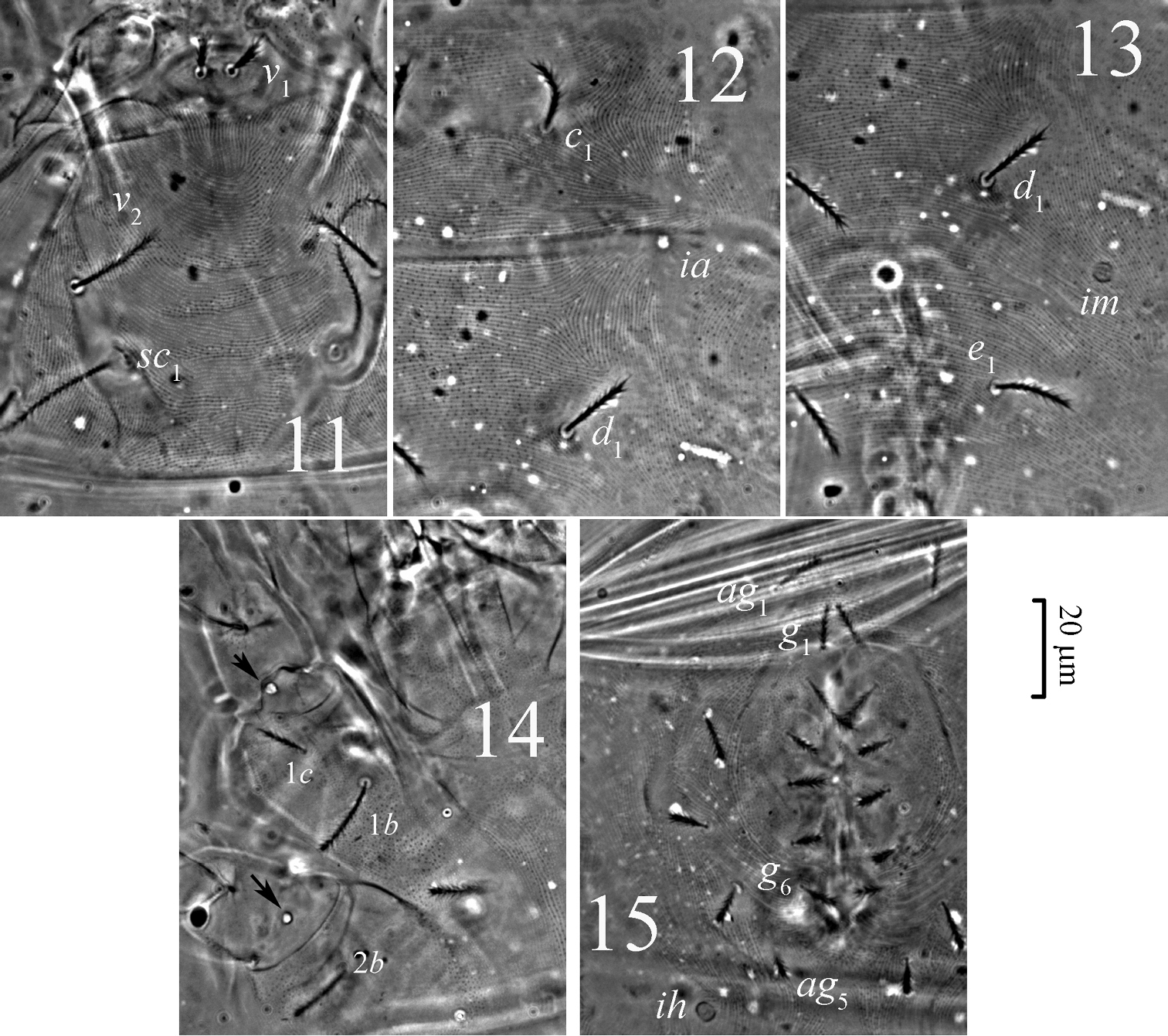
Pseudoeupodes porosus Khaustov, gen. n., sp. n., female: 11 prodorsum 12 striation in the area of setae c1 and d113 striation in the area of setae d1 and e114 venter of propodosoma, arrows point to pore-like structures 15 genital area.
Male and immatures. Unknown.
The genus name is derived from the related genus Eupodes and prefixed pseudo- (from Greek ψευδής) meaning false.
The new genus is most similar to Caleupodes Baker, 1987. Both genera are characterized by the naso folded down to the ventral surface of the prodorsum, the same palpal chaetotaxy, six pairs of eugenital and five pairs of aggenital setae, the presence of only two pairs of pseudanal setae, femur IV not enlarged, trochanteral setal formula 1–1–1–1, and legs I-IV shorter than idiosoma. The new genus differs from Caleupodes by having striated dorsal cuticle (reticulated in Caleupodes), the absence of a transverse furrow between segments D and E (all hysterosomal segments are clearly separated by transverse furrows in Caleupodes), coxisternal setal formula 3–1–4–2 (3–1–4–3 in Caleupodes), and genua I and II with a solenidion (absent in Caleupodes). The new genus is also closely related to Niveupodes Barilo, 1991 in the naso folded down to the ventral surface of the prodorsum, femur IV not enlarged, trochanteral setal formula 1–1–1–1, legs I-IV shorter than idiosoma, and only two pairs of pseudanal setae. It differs from Niveupodes by the presence of dorsal transverse furrows between segments C – D and E – F (dorsal hysterosoma without transverse furrows in Niveupodes), coxisternal setal formula 3–1–4–2 (3–1–4–3 in Niveupodes), five pairs of aggenital and six pairs of eugenital setae (four aggenital and five eugenital in Niveupodes), the absence of scapular lyrifissure isc (present in Niveupodes, according to
http://zoobank.org/3897531B-758D-4AC5-B0F0-F9D85048964C
Figs 1–15Female. Length of idiosoma 288 (280–300), width 163 (157–180).
Idiosomal dorsum (Figs 1, 11–13). Idiosoma with striae bearing microtubercles (Figs 11–13). All dorsal setae densely pilose, setae v1 slightly widening apically. Prodorsum with pair of longitudinal lines between the bases of setae sc2 and anterior margin of prodorsum near setae v1. Setae sc1 with large barbs, a weak reticulate subcuticular ornamentation visible posteriorly to bases of sc1 (Fig. 11). Lyrifissures ia situated posterolaterally to bases of setae c1; im situated posterolaterally to bases of setae d1; ip situated between setae e1 and f1. Length of dorsal setae: v1 9 (98-10), v2 16 (15–19), sc1 50 (47–55), sc2 15 (14–17), c1 13 (12–16), c2 21 (19–25), d1 16 (14–19), e1 15 (14–18), f1 40 (37–45), f2 25 (24–28), h1 23 (21–26), h2 17 (15–21). Setae f1 longest of dorsal hysterosomal setae but not of trichobothrial form.
Idiosomal venter (Figs 2, 3, 14–15). All ventral setae densely pilose. Setae 1a, 3a, 4a (10–13) slightly widened distally; setae 1c the shortest on coxal fields. Genital setae arranged in one longitudinal row, anterior five pairs (g1–g5) situated on non-striated genital covers bearing only microtubercles, posterior pair (g6) situated outside genital covers on striated cuticle (Figs 3, 15). Anterior two pairs of genital setae distinctly longer (10–11) than other genitals (6–7). Aggenital setae increasing in length from ag5 (7) to ag1 (10–11). Eugenital setae situated on protuberances and arranged in three groups: two anterior (eu1, eu2), one medial (eu3) and three posterior (eu4-eu6) (Fig. 3). Pseudanal setae ps1 15 (14–18) distinctly longer than ps3 11 (10–14). Lyrifissure ih located anterolaterally to bases of setae ps3.
Gnathosoma (Figs 4–6). Integument papillate. Subcapitulum (Fig. 4) roughly triangular, with two pairs of minute smooth adoral setae (or1, or2), located subapically; subcapitular setae sbc2 densely pilose, sbc1 smooth, located laterally at level of proximal margin of palp trochanters, about one-third as long as sbc2, sbc2 inserted ventrally one third to one quarter of distance between sbc1 and tip of subcapitulum, labrum acuminate. Chelicera (Fig. 5) 60 in length. Palps (Fig. 6) with supracoxal seta ep minute, brush-like, femorogenual and tibial setae densely pilose, tarsal setae acm and sl” smooth, other tarsal setae pilose, p” bifurcate distally.
Legs (Figs 7–10). Relative lengths of legs: I>IV>III>II. All leg setae densely pilose. Leg I (Fig. 7): Supracoxal setae ep of the same shape and length as palpal supracoxal setae ep. Femur incompletely divided into basi- and telofemur. Setal formula: Tr – 1, Fe – 6+5, Ge – 7(1σ), Ti – 7(2φ), Ta – 18 (2ω, 1ε). Famulus ε (stellate seta) located in a shallow depression clearly posterolaterally to basal part of rhagidial solenidion ω1 and anterior to seta ft”. Rhagidial solenidion φ1 obliquely oriented toward anterior lateral margin of leg, located anterodorsally and situated in shallow depression; solenidion φ2 located posterodorsally in the space between setae l’1 and l”1; solenidion σ located anterodorsally, about 1.5 times as long as φ2. All setae on tarsus, d and l’ on tibiae are eupathidia (as applied for Eupodoidea by
Holotype female, slide № AK210494, CRIMEA: Yalta mountain-forest Nature Reserve, moss on soil, 21 April 1994, coll. A.A. Khaustov. Paratypes: five females, same data as holotype; seven female paratypes, CRIMEA: Yalta, moss on soil, 5 March 1994, coll. A.A. Khaustov.
The name of the new species refers to the presence of unusual pore-like structures in the soft cuticle between the coxisternal plates and the trochanters of all legs.
The present systematic organisation of the family Eupodidae and the superfamily Eupodoidea is highly unstable. Preliminary morphological cladistic analyses (
The monotypic family Dendrochaetidae (
Currently I recognize 11 genera in the family Eupodidae: Xerophiles Jesionowska, 2003, Benoinyssus, Claveupodes Strandtmann & Prasse, 1976, Eupodes, Aethosolenia Baker and Lindquist, 2002, Neoprotereunetes, Linopodes, Cocceupodes, Niveupodes, Caleupodes, and Pseudoeupodes gen. n.
| 1 | Setae f1 trichobothrium-like | 2 |
| – | Setae f1 not trichobothrium | 3 |
| 2 | Setae ps2 present | Xerophiles |
| – | Setae ps2 absent | Benoinyssus |
| 3 | Setae ps2 present | 4 |
| – | Setae ps2 absent | 7 |
| 4 | Trichobothria (sc1) filiform | 5 |
| – | Trichobothria (sc1) clavate | Claveupodes |
| 5 | Femur IV not swollen | 6 |
| – | Femur IV swollen, adapted for jumping | Eupodes |
| 6 | Setae h1 trichobothrium-like, adanal setae present, tibia and tarsus I much thicker than other leg segments | Aethosolenia |
| – | Setae h1 not trichobothrium, adanal setae absent, tibia and tarsus I not enlarged | Neoprotereunetes |
| 7 | Leg I shorter or slightly longer than idiosoma, solenidia in rhagidial organs not T-shaped | 8 |
| – | Leg I more than 3 times longer than idiosoma, solenidia in rhagidial organs T-shaped | Linopodes |
| 8 | Leg I distinctly shorter than idiosoma, sejugal furrow well developed, femur IV not enlarged, naso folded to ventral surface of prodorsum | 9 |
| – | Leg I usually longer than idiosoma, sejugal furrow absent or poorly developed, femur IV enlarged, naso directed anteriorly | Cocceupodes |
| 9 | Hysterosoma dorsally with 3 pairs of lyrifissures | 10 |
| – | Hysterosoma dorsally with 4 pairs of lyrifissures (scapular lyrifissure present) | Niveupodes |
| 10 | Idiosoma dorsally reticulated, all hysterosomal segments delineated by distinct transverse furrows, solenidia on genua I and II absent | Caleupodes |
| - | Idiosoma dorsally striated, transverse furrows present only between segments C–D and E–F, solenidia on genua I and II present | Pseudoeupodes gen. n. |
The author thanks Dr. Bruce Halliday (CSIRO Entomology, Canberra, Australia) and Dr Evert Lindquist (Ottawa, Canada) for critical reviewing of the manuscript.