





(C) 2013 Jaime Troncoso-Palacios. This is an open access article distributed under the terms of the Creative Commons Attribution License 3.0 (CC-BY), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
For reference, use of the paginated PDF or printed version of this article is recommended.
In the current study, we review the taxonomic status of Liolaemus nigromaculatus. Despite being the nominal species of the nigromaculatus group and being the second species of the genus Liolaemus that was described, this species is of uncertain type locality and its true identification is a matter of discussion. After carefully analyzing several digital pictures of the holotype (juvenile male), reviewing all of the literature concerning the issue, examining specimens of nearly all recognized species of the nigromaculatus group, and determining the locations visited by the specimen collector, we are able to point out the following: 1) Liolaemus nigromaculatus was collected between Puerto Viejo and Copiapó of the Atacama region in Chile, and not in Huasco 2) Liolaemus bisignatus is a nomen nudum, and populations attributed to Liolaemus bisignatus should be referred to as Liolaemus nigromaculatus. 3) There is agreement that Liolaemus copiapoensis is indistinguishable from populations currently referred to as Liolaemus bisignatus (= Liolaemus nigromaculatus), 4) Populations found in Huasco (currently considered the type locality of Liolaemus nigromaculatus) are very similar to those found in Caldera (currently considered Liolaemus bisignatus) and should be designated as Liolaemus nigromaculatus, and 5) Liolaemus oxycephalus and Liolaemus inconspicuus are not synonymous with Liolaemus nigromaculatus, although their true identities are difficult to determine. We also detail several characteristic based on the holotype of Liolaemus nigromaculatus, in addition to drawing diagnostic comparisons between this species and others belonging to the nigromaculatus group.
Liolaemus, nigromaculatus, bisignatus, copiapoensis, Atacama
The genus Liolaemus is comprised of 230 species (
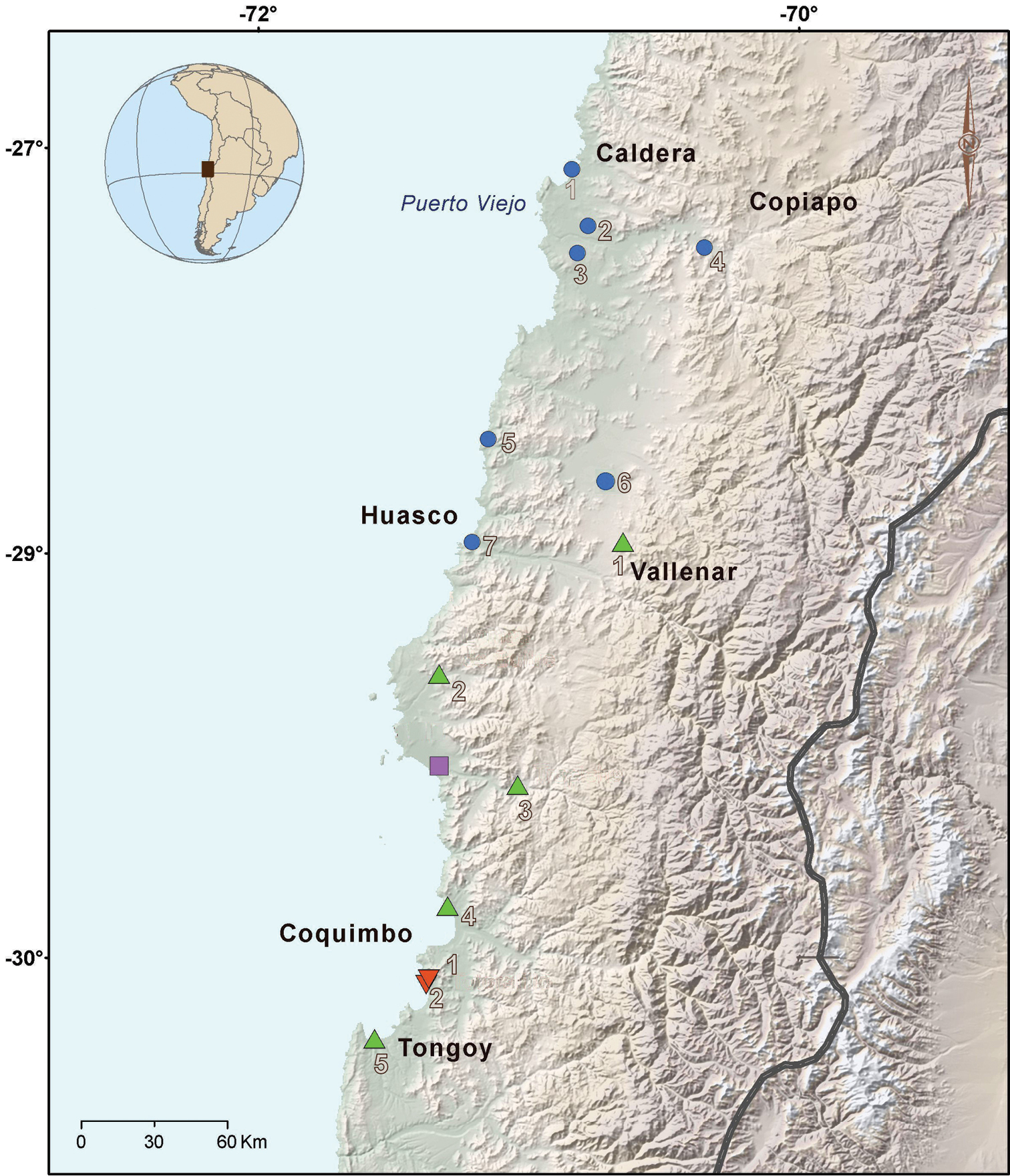
Distribution map of specimens used in this study. Blue circle: Liolaemus nigromaculatus (1 Caldera 2 Road between Caldera and Copiapó 3 Near to Puerto Viejo 4 Copiapó 5 Llanos de Challe 6 Road between Vallenar and Copiapó and 7 Huasco). Green triangle: Liolaemus atacamensis (1 Near to Vallenar 2 Lomas de Buitre 3 El Trapiche 4 Punta Teatinos and 5 Pachingo). Lilac square: Liolaemus silvai (Carrizalillo). Red triangle: Liolaemus zapallarensis (1 Totoralillo and 2 Las Tacas). The main localities are indicated.
Later,
With such uncertainty about the type locality,
Unfortunately,
Finally,
Therefore, in this paper we review the taxonomic status of Liolaemus nigromaculatus through the characterization of the holotype and clarification of the location in which it was collected. In addition, we provide a diagnosis respect of the other species of the nigromaculaus group.
The characteristics used for descriptions were taken from
Acronyms mentioned in this publication are: MNHN-CL (Museo Nacional de Historia Natural, Chile), MZUC (Museo de Zoología, Universidad de Concepción), MRC (Museo Regional de Concepción), SSUC Re (Colección de Flora y Fauna Patricio Sánchez Reyes, Pontificia Universidad Católica de Chile) and ZMB (Museum für Naturkunde).
Characteristics of the Liolaemus nigromaculatus holotype, ZMB 613. The holotype is a juvenile male (Figs 2, 3). The following measurements were taken from

Dorsal view of the Liolaemus nigromaculatus holotype (ZMB 613).

Ventral view of the Liolaemus nigromaculatus holotype (ZMB 613). Photograph by F. Tillack.
Color in alcohol. Dorsal and lateral views of the head are light brown in color, the same tone of the dorsum, and have numerous black spots which do not form a clear pattern. On the neck, these spots become smaller. The color of the dorsum is gray-brown (gray according to

Lateral shoulder view of some specimens studied. A Holotype of Liolaemus nigromaculatus (ZMB 613, juvenile male) with black “ג” shaped antehumeral spot and dorsal scales without mucrons (Photograph by F. Tillack) B Holotype of Liolaemus bisignatus (=Liolaemus nigromaculatus, adult male MNHN-CL 1477) with black “ג” shaped antehumeral spot and dorsal scales without mucrons C Male specimen of Liolaemus zapallarensis (MZUC 29118) with irregular antehumeral spot and dorsal scales with mucrons D Male specimen of Liolaemus atacamensis (SSUC Re 469) with rounded antehumeral spot and dorsal scales without mucrons.
Diagnosis of the nigromaculatus group. Currently, it is difficult to establish a diagnosis for this group. Preliminary evidence in an ongoing molecular phylogenetic study (Troncoso-Palacios and Schulte, in prep) shows that this group is composed of two clades. One clade includes Liolaemus atacamensis, Liolaemus ater, Liolaemus kuhlmanni, Liolaemus melaniceps, Liolaemus nigromaculatus, Liolaemus sieversi, Liolaemus silvai and Liolaemus zapallarensis; and the second includes Liolaemus hellmichi, Liolaemus platei and Liolaemus velosoi. A similar proposal is made in
However, since a formal study is lacking, the following diagnosis includes the currently considered species as members of the group nigromaculatus.
Diagnosis of Liolaemus nigromaculatus based on the holotype. Liolaemus nigromaculatus can be differentiated from the other species of the nigromaculatus group through the following combination of characteristics: 1) nasal and rostral scales separated by one scale, 2) presence of projected ciliary scales, 3) dorsum with abundant, black-spotted scales, 4) series of black spots on the paravertebral fields, from the base of the neck to the base of the tail, 5) marked “ג” shaped antehumeral black spot from the shoulder to the humeral zone, 6) dorsal scales are rounded or lanceolated, slightly keeled, and without mucrons. Diagnosis with respect to other species of the nigromaculatus group is listed below.
Liolaemus nigromaculatus differs from Liolaemus hellmichi (
Liolaemus nigromaculatus differs from Liolaemus melaniceps (
Liolaemus nigromaculatus differs from Liolaemus ater, Liolaemus kuhlmanni, Liolaemus zapallarensis (
Finally, Liolaemus nigromaculatus differs from its most similar species, Liolaemus atacamensis (
Scale and morphological characteristics of geographically close species to Liolaemus nigromaculatus (examined juveniles are excluded). M = males and F = females.
|
Liolaemus nigromaculatus (Puerto Viejo - Copiapó and surroundings) M=19, F=16 |
Liolaemus atacamensis M=7, F=9 |
Liolaemus kuhlmanni M=7, F=9 |
Liolaemus zapallarensis M=1, F=2 |
|
|---|---|---|---|---|
| Midbody Scales | 50–61 | 48–54 | 52–58 | 48–52 |
| Ventrals | 77–84 | 66–77 | 80–90 | 79–80 |
| Nasal Separated from Rostral | Yes | Yes | Yes | Yes |
| Color of Male | Gray (with olive or yellow shades) | Gray or brown | Black | Black |
| Shape of Dorsal Scales | Rounded or lanceolate | Rounded or lanceolate | Lanceolate | Lanceolate |
| Mucrons in Dorsal Scales | No | No | Yes | Yes |
| Black Spotted Dorsal Scales | Yes | No | No | No |
| Male with “ג” Shaped Antehumeral Spot | Yes | No | Indistinguishable | Indistinguishable |
| Maximum SVL (mm) | 83.0 | 67.2 | 81.2 | 72.4 |
The relationship between Liolaemus bisignatus (
We agree with
The relationship between Liolaemus nigromaculatus (
The second publication which deals with this species (

Some specimens listed by
Our review of twenty-three specimens of Liolaemus bisignatus (adults and juveniles, including the holotype) allows us to determine that the populations currently referred to as Liolaemus bisignatus should be referred to as Liolaemus nigromaculatus based on the following: 1) Of the species in the nigromaculatus group that have the nasal separated from the rostral, only Liolaemus bisignatus and Liolaemus atacamensis (Figs 6, 7) have dorsal scales without mucrons, and of these, only Liolaemus bisignatus overlaps with the diagnostic characters of Liolaemus nigromaculatus, 2) The color of the Liolaemus nigromaculatus holotype is brown-gray with a series of black spots over the paravertebral fields, as in juveniles of Liolaemus bisignatus, 3) Liolaemus nigromaculatus has a black“ג” shaped antehumeral spot, from the shoulder to the humeral zone, like Liolaemus bisignatus, 5) Liolaemus nigromaculatus has abundant black spotted scales on the dorsum, like Liolaemus bisignatus, 6) The holotype of Liolaemus nigromaculatus has at least 80 ventral scales, which is in the range of Liolaemus bisignatus but not for Liolaemus atacamensis (Table 1), 7) Of the species in the nigromaculatus group with the nasal separated from the rostral, only Liolaemus bisignatus and Liolaemus atacamensis are known to be from the zone in which Meyen collected the holotype of Liolaemus nigromaculatus (see below), and of these only Liolaemus bisignatus overlaps with the diagnostic characteristics of Liolaemus nigromaculatus, 8) In Huasco, the location currently accepted as the type locality of Liolaemus nigromaculatus, it is only possible to find two other species of Liolaemus (Liolaemus bisignatus and Liolaemus fuscus), this explains why

Variation of color pattern in Liolaemus nigromaculatus. A and B Adult males from 20 Km SE from Puerto Viejo C Adult female from the same location D Juvenile from Caldera.

Variation of color pattern in Liolaemus atacamensis. A Adult male from El Trapiche B Adult male from Lomas de Buitre C Adult female from Lomas de Buitre.
The true type locality of Liolaemus nigromaculatus.
According to
We can discard the area from San Fernando to Valparaíso as the type locality of Liolaemus nigromaculatus since no species of the nigromaculatus group has been documented for this area. On the other hand, all species of nigromaculatus group with the nasal scale separated from rostral and that inhabit the surroundings of Coquimbo (Liolaemus atacamensis, Liolaemus kuhlmanni and Liolaemus zapallarensis) are clearly distinguishable from Liolaemus nigromaculatus, so Liolaemus nigromaculatus is not found in Coquimbo.
Meyen never visited Huasco, so it is impossible that the specimen ZMB 613 was collected by him there.
In conclusion, the true type locality of Liolaemus nigromaculatus should be restricted to the route and surroundings from Puerto Viejo to Copiapó because in the entirety of this route specimens of Liolaemus nigromaculatus can be found and are being by far the most abundant species. Since this route covers approximately 60 Km, it is impossible to obtain a more precise location.
Variation of the species. Variation analysis was based on 40 specimens (19 males, 16 females, and 5 juveniles) from a transect of Puerto Viejo – Copiapó and the surroundings, including Caldera. The data are as follows: SVL adult males: 75.7 ± 5.4 mm. SVL adult females: 67.4 ± 7.2 mm. Tail length in adult males: 84.4 ± 15.5 mm. Tail length in adult females: 80.8 ± 9.7 mm. Head length in adult males: 19.3 ± 1.8 mm. Head length in adult females: 17.6 ± 2.7 mm. Head width in adult males: 14.3 ± 0.9 mm. Head width in adult females: 11.9 ± 0.4 mm. Head height in adult males: 9.5 ± 1.0 mm. Head height in adult females: 8.0 ± 0.8 mm. Forelimb length in adult males: 25.4 ± 3.8 mm. Forelimb length adult in females: 19.6 ± 1.7 mm. Hind limb in adult males: 40.2 ± 5.5 mmm. Hind limb in adult females: 32.1 ± 1.6 mmm.
Pentagonal or hexagonal interparietal, smaller than the parietals and surrounded by 6-7 scales; 7-9 scales between the interparietal and rostral; orbital semicircles are complete, formed by 8-10 scales, or incomplete; 3-5 supraoculars and 6 supercilliaries. The subocular is whitish with a vertical black line at the center. Two scales between nasal and canthal. The nasal is separated from the rostral by one scale and surrounded by six scales. There is one row of lorilabials between the supralabials and subocular. There are 4-5 supralabials, the last of which is curved upward without contact to the subocular; 5-6 infralabial scales. The mental is pentagonal and in contact with four scales. There are four pairs of post-mental shields; the second pair is in contact or is separated by one scale. There are 2-3 enlarged scales on the anterior edge of the ear but without covering the meatus. Temporal scales are subimbricated and smooth or slightly keeled. The lateral neck fold is “Y” shaped and antehumeral fold is present. There are 6-7 temporal scales between the level of supercilliaries and commissure. Midbody scales: 50-61. Dorsal scales are rounded or lanceolated, subimbricated or imbricated, slightly keeled and without mucrons, and some specimens have interstitial granules. Dorsal scales are similar or smaller than the ventrals. Ventrals : 77-84. Ventral scales are rounded, smooth, subimbricate, and without interstitial granules. There are 2-3 precloacal pores in the males. The suprafemoral scales are rounded, imbricated, and keeled without mucrons. Infrafemoral scales are rounded, smooth, and imbricated. Supraantebrachials scales are rounded, imbricated, and keeled without mucrons. Infraantebrachials are rounded, imbricated, and smooth. The ventral tail scales are lanceolate, imbricated, keeled, and with mucrons.
Color: There is a highly variable color pattern, which varies with the size and sex of individuals. The juveniles have a brown head, with the same shade on the dorsum and with some dark spots or no spots. The dorsum has a brown-yellow color. The vertebral line is fragmented or absent, always disappearing in larger juveniles. There are nine to ten dark spots on the paravertebral fields, from the neck to the base of the tail. These spots have white or yellow on the posterior edge. There is a yellow supraocular line, and black “ג” shaped antehumeral spot. The temporal band is formed by four to seven dark spots which have white or yellow on the posterior edge. The limbs are brown with small black and white spots. There is a whitish ventral color, sometimes with small dark spots. The throat is striated or spotted. The tail is brown, with vertebral line and ringed.
For adult males, the following observations were made: The supraoculars lines of juveniles disappear. The head is gray, has the same shade as the body, and which, in some specimens, also has yellow or olive shades. The dark spots of the temporal bands and paravertebral fields progressively merge until disappearing in the larger specimens, leaving a gray color on the dorsum without design, and sometimes with olive and yellow shades. On the dorsum there are abundant black spotted scales, dispersed and without forming a design. There is a black “ג” shaped antehumeral spot, and in some specimens it is accompanied by three to six round and smaller dark spots from shoulder towards the rear. There is an absence of a vertebral line. In some specimens, the flanks have yellow or orange color. The tail has either a vertebral line or no design. The limbs are gray with small black and white spots, and the belly is whitish. The throat is striated or spotted, and the cloacal region is orange or yellow.
Finally, for females observations were as follows: There is a gray or brown color and abundant black spotted scales dispersed on the dorsum. There is a black “ג” shaped antehumeral spot, but it is more diffuse than in males. Females differ from the males in that they have dark spots over the dorsum with white on the posterior edge. Also, females have a yellow supraocular line. There is an absence of vertebral line. The limbs are gray with small black and white spots. There is a whitish belly, but in some specimens there is orange in the middle. The throat is striated or spotted. The tail has a vertebral line and may be ringed.
Without a doubt, one of the most confused issues in the taxonomy of Chilean herpetology is the definition of Liolaemus nigromaculatus, whose taxonomic status and type locality have been uncertain for many years (
Several authors mentioned Coquimbo as the inhabiting locality of Liolaemus nigromaculatus (
The most similar species to Liolaemus nigromaculatus is Liolaemus atacamensis.
Use of digital pictures of type specimens has proved to be a powerful and useful tool for clarifying confusing taxonomic issues. Recently,
Here, we hope to have contributed to the clarification of the taxonomic identity of Liolaemus nigromaculatus in addition to providing new data and correcting some mistakes, all with the end of trying to understand the still uncertain semantics of the nigromaculatus group.
We thank M. Penna for his support. Frank Tillack (Museum für Naturkunde, Berlin, Germany) for sending us pictures of ZMB 613 and ZMB 615. P. Zabala (Pontificia Universidad de Católica de Chile) for allowing us to deposit materials into the collection under his care (SSUC Re). We thank the following colleagues and museums for allowing us to study specimens: J.F. Troncoso (Museo de Historia Natural de Concepción), J. Artigas & J.C. Ortiz (Museo de Zoología de la Universidad de Concepción), and H. Núñez (Museo Nacional de Historia Natural). We also thank the following colleagues and friends for their support in the field: Y. Marambio-Alfaro, D. Hiriart, O. Rabayo, F. Ferri, F. Urra and E. Riveros. C.F. Garin was supported by a CONYCIT-Chile Doctoral Fellowship. We thank A. VanCott for improving the English. We thank the two anonymous reviewers for greatly contributing to the improvement of this article. Finally, we thank the Servicio Agrícola y Ganadero (SAG, Chile) for authorizing the capturing of specimens through permission Nº 2411 (26/04/2012).
Liolaemus atacamensis. SSUC Re 469. 20 Km al norte de Vallenar. F. Ferri col. 2010. SSUC Re 470-71. El Trapiche, Coquimbo. J. Troncoso-Palacios, Y. Marambio & D. Hiriart cols. Mayo, 2012. SSUC Re 454, 464-68. Lomas de Buitre, Freirina, Atacama. J. Troncoso-Palacios, Y. Marambio & D. Hiriart cols. Mayo, 2012. SSUC Re 455-61. Playa Humedal Pachingo, entre Tongoy y Puerto Aldea, Coquimbo. C. Garin col. 10/12/2009. MZUC 30193, 30196. Punta Teatinos, sector de dunas costeras. J.C. Ortiz col. 14/09/1982.
Liolaemus fuscus. SSUC Re 258-59. Altos de Jahuel, Valparaíso. F. Ferri & J. Troncoso-Palacios. 19/12/2010. SSUC Re 255-57. Camino a Farellones. F. Ferri col. 18/12/2010. SSUC Re 260. Camino a Ovalle, Coquimbo. F. Ferri. 17/03/2012. MRC 229-34, 236. Huasco. J. Moreno col. May, 1983.
Liolaemus hellmichi. MZUC 25942-52. Cerro Moreno. J.C. Ortiz col. 02/04/2001.
Liolaemus kuhlmanni. MZUC 28829, 28838-41. Papudo. J. Simonetti & J.C. Ortiz col. 9/9/1977. SSUC Re 282-84. Dunas de Ritoque. F. Ferri col. 13/11/2010. SSUC Re 285-97. Caleta de La Ligua. F. Ferri col. 26/11/2010. SSUC Re 473. Dunas de Concón. J. Troncoso-Palacios & F. Urra cols. 04/01/2011.
Liolaemus nigromaculatus. SSUC Re 478 20 Km al sur este de Puerto Viejo. J. Troncoso-Palacios & Y. Marambio cols. Mayo, 2012. MRC 051-53. Caldera. J. Moreno col. Mayo, 1982. MRC 162, 273, 276, 282-83. Caldera. J. Moreno col. 1983. SSUC Re 306-15, 474-75. Caldera. F. Ferri col. Noviembre, 2011. SSUC Re 476-77. Caldera, Atacama. Y. Marambio col. Mayo, 2012. MNHN-CL 2237-38. 20 km de Caldera entre Copiapó y Caldera. H. Núñez col. 30/09/1991. SSUC Re 007. Llanos de Challe. G. Lobos, A. Channier & J. González cols. 2002. SSUC Re 453, 462. 50 Km al norte de Vallenar. J. Troncoso-Palacios col. 05/06/2011. MRC 087-94. Copiapó. C. Valdovinos col. 15/9/1984. MRC 514. Copiapó. Unknown collector. Noviembre, 1996. MNHN-CL 2249-55. Travesía, Copiapó. H. Núñez col. 28/09/1999. MRC 059. Huasco. J. Moreno col. 11/08/1982. MRC 225-28, 272, 274-75, 285. Huasco. J. Moreno col. 15/03/1983.
Liolaemus melaniceps. MNHN-CL 3601-03. Isla Chungungo. J.C. Torres-Mura col. January, 2002.
Liolaemus platei. MZUC 2152-53. Combarbalá. I. Peña col. Noviembre de 1961. SSUC Re 029. Llanos de Challe. G. Lobos, A. Channier & J. González cols. 2002. SSUC Re 316-20, 335-36, 380. Caldera. Francisco Ferri col. SSUC Re 321. Illapel. Francisco Ferri col. SSUC Re 420. Coquimbo. J. Troncoso-Palacios & Y. Marambio cols. 12/12/2011. SSUC Re 526. Coquimbo. J. Troncoso-Palacios & Y. Marambio cols. Mayo, 2012. MRC 058, 063. Chañaral. J. Moreno col. 28/7/1982.
Liolaemus sieversi. MZUC 8914, 12096-98, 12100, 12119-23, 12126, 12128. Isla Los Locos. R. Donoso-Barros col. 15/02/1954.
Liolaemus silvai. MZUC 30184. Carrizalillo. J.C. Ortiz col. February, 1983.
Liolaemus velosoi. MZUC 36612-14, 36618-20, 36624. Estación Paipote. J.C. Ortiz col. 16/02/1978. MZUC 32695, 32699, 32702, 32704, 32706. Copiapó. R. Moreno col. Febrero de 2000. MRC 054. Copiapó. Unknown collector. 16/06/1982. MRC 055. Copiapó. Unknown collector. 20/04/1982. MRC 061, 062, 066. Copiapó. Unknown collector. 19/07/1982. SSUC Re 322-26. Tierra Amarilla. J. Troncoso-Palacios & F. Ferri. cols. 23/11/2011. SSUC Re 330. Diego de Almagro. F. Ferri, J. Troncoso-Palacios cols. 09/12/2011. SSUC Re 327-29, 331-34, 419. Diego de Almagro. F. Ferri & J. Troncoso-Palacios cols. 12/12/2011.
Liolaemus zapallarensis. MZUC 29118. Las Tacas, Región de Coquimbo. J.C. Ortiz col. 15/10/1976. MZUC 29127. Las Tacas, Lagunillas, sector rocoso de la playa. J.C. Ortiz & J. Simonetti cols. 10/09/1977. SSUC Re 472. Totoralillo, Región de Coquimbo. J. Troncoso-Palacios & Y. Marambio cols. 12/12/2011.