





Citation: Xu G-L, Zhang F (2014) A new species of Dicranocentrus (Collembola, Entomobryidae) from China with comments on the systematic position of the genus. ZooKeys 417: 1–8. doi: 10.3897/zookeys.417.7373
Dicranocentrus liuae sp. n. is described from the northern subtropical region of China. The new species is most similar to D. wangi Ma & Chen, 2007, but differs from it in the relatively shorter Ant. V, the 1+1 central macrochaetae on Abd. III, the number of chaetae on tenaculum, and the absence of dental spines. The systematic position of Dicranocentrus is also discussed. Present evidence, particularly S-chaetotaxy, indicates that the genus is closer to Heteromurus than to the unscaled species of Orchesella and Orchesellides.
D. liuae sp. n. , chaetotaxy, Orchesellinae, Heteromurus
The genus Dicranocentrus was erected by
So far, three Dicranocentrus species have been reported from China: Dicranocentrus indicus Bonet, 1930 from Taiwan, Dicranocentrus chenae Ma, Chen & Soto-Adames, 2006 from Guangxi, and Dicranocentrus wangi Ma & Chen, 2007 from Gangdong (Fig. 1). Here, we describe a new species from the northern subtropical region of China, compare it with other orchesellids, and discuss the systematic position of the genus.
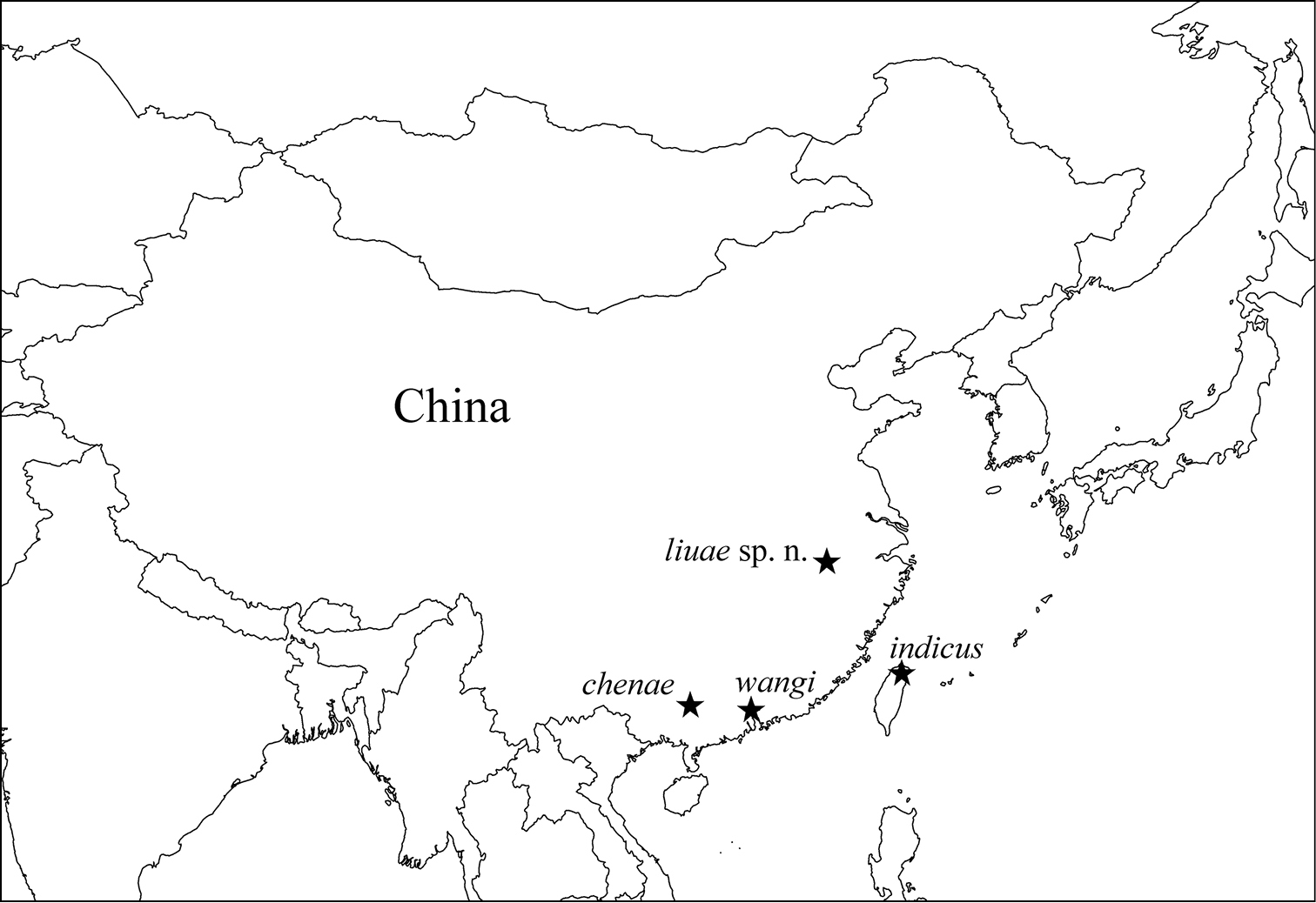
Geographical distribution of Dicranocentrus species from China.
Specimens were mounted in Marc André II solution after clearing in lactic acid and were studied using a Nikon E600 and SMZ-1000 microscope. Photographs were enhanced with Photoshop CS2/PC (Adobe Inc.). The number of macrochaetae is given by half-tergite in the descriptions. Dorsal cephalic chaetotaxy and interocular chaetae follow
Abbreviations. Th. I–III – thoracic segment I–III; Abd. I–VI – abdominal segment I–VI; Ant. I–IV – antennal segment I–IV; mac – macrochaeta/ae; mic – microchaeta/ae; ms – S-microchaeta/ae; sens – ordinary tergal S-chaeta/ae; post-labial quadrangle – PLQ.
http://zoobank.org/E9501B65-B6EC-4D84-AC1C-1BC07A65E5B8
Figs 2–14, Table 1China, Anhui, Shitai, Guniujiang, 30.092°N, 117.482°E, altitude 208m.
Holotype: ♀ on slide, China, Anhui, Shitai, Guniujiang, the entrance of Yan hamlet, 30.092°N, 117.482°E, altitude 208m, 15 August 2011, F Zhang, DY Yu and YH Ren leg. (#C9676). Paratypes: 3 ♀♀ and 1 ♂ on slides, 4 in alcohol, same data as holotype. Other material: 1 ♀ on slide, China, Anhui, Shitai, Gongxi, 27 August 1994, JX Chen leg. (#C8391); 1 ♀ on slide, China, Zhejing, Jin Xian, Tiantongshan National Natural Reserve, 10 June 1995, JX Chen leg. (#C8458).
Body length up to 1.8 mm.
Ground colour pale yellow. Eye patches dark. Blue pigment present on antennae and legs (Fig. 2). Scales brown, rounded, truncate, or pointed with numerous short striations; scales present on Ant. I‒IV, body, legs, both side of ventral tube and manubrium, and ventral side of dens.w

Dicranocentrus liuae sp. n. Habitus.
Antenna 2.0–2.7 times as long as cephalic diagonal. Ant. V and VI annulated and their length ratio as 1.2–0.7: 1. Four types of common chaetae observed: ciliate, thin (Fig. 3A) or thick (Fig. 3B) mic; smooth, straight, spiny mic on Ant. I and III (Fig. 3C); long, straight chaetae smooth or weakly ciliate (Fig. 3D). Most S-chaetae slightly curved, short (Fig. 3E–I) or long (Fig. 3J). Distal Ant. II with 1 rod-like S-chaeta ventrally (Fig. 3K), 2 strongly curved, thickened S-chaetae externally (Fig. 3L–M). Ant. VI apical bulb absent.
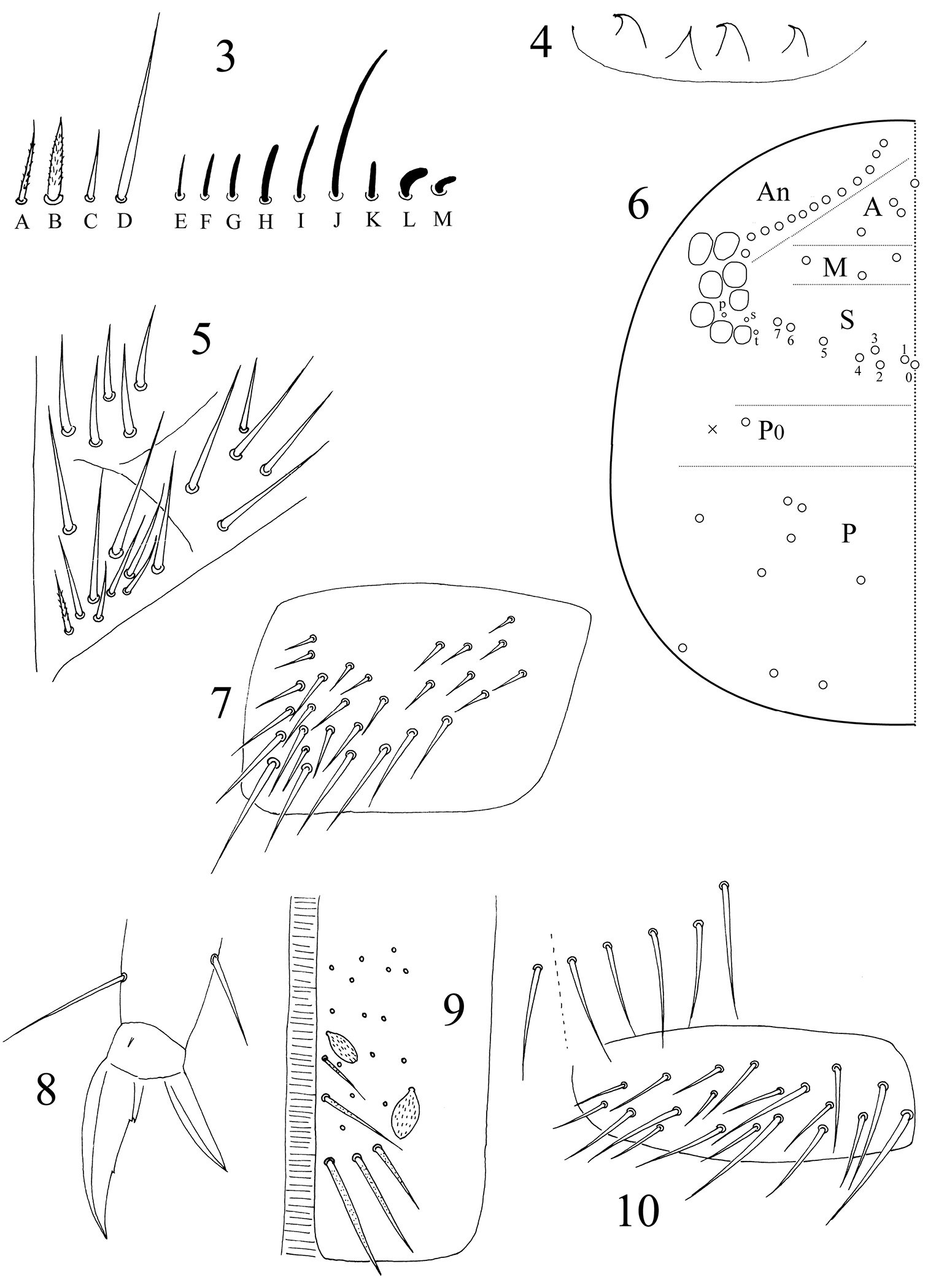
Dicranocentrus liuae sp. n. 3 antennal chaetae 4 labral papillae 5 labial chaetae 6 dorsal cephalic chaetotaxy 7 trochanteral organ 8 hind claw 9 anterior face of ventral tube 10 distal posterior and lateral flap chaetae of ventral tube.
Eyes 8+8, G and H smaller. Prelabral and labral chaetae 4/5, 5, 4, all smooth; prelabral ones stronger than labral ones. Labral papillae 4, cone-like with apical spine often curved (Fig. 4). Mandibles with 4+5 apical teeth. Lateral process of labial palp papillae E as thick as normal chaetae, with tip not reaching apex of labial papilla. Subapical chaeta of maxillary outer lobe thicker than apical; 3 smooth sublobal hairs on maxillary outer lobe. Labium with 5 smooth proximal chaetae and 8–10 submentum chaetae; the most external one A5 on mentum shorter than A1‒5; at most 1 ciliate chaeta on submentum (Fig. 5). PLQ chaetae smooth and 2+2 weakly ciliate chaetae posterior to PLQ. Dorsal cephalic chaetotaxy with10–13 antennal (An), 3 median (M), sutural S0–7, 1 postocular (P0) and 9 posterior (P) mac. Interocular chaetae 3 as p, s, t (Fig. 6).
Trochanteral organ with about 30 smooth spiny chaetae (Fig. 7). Some inner differentiated tibiotarsal chaetae smooth under light microscope. Unguis with 3 inner teeth, all minute. Unguiculus lanceolate with outer edge smooth. Tenent hairs acuminate (Fig. 8).
Abd. IV 1.66–1.89 times as long as Abd. III along dorsal midline. Ventral tube anteriorly with many weakly ciliate chaetae and some scales (Fig. 9); posteriorly with many smooth chaetae; both sides with scales; each lateral flap with about 20 smooth chaetae (Fig. 10). Tenaculum with 4+4 teeth, corpus with 2–4 smooth chaetae. Manubrial plaque with 3 pseudopores and 5–9 ciliate chaetae on each side (Fig. 11). Manubrium dorsally with rows of smooth chaetae but their number not clear. Dens without inner spines. Dental of lobe with 1+1 large blunt ciliate and about 5+5 small smooth chaetae. Smooth distal part of dens 4.4–5.7 times as long as mucro; mucro bidentate with two subequal teeth (Fig. 12).
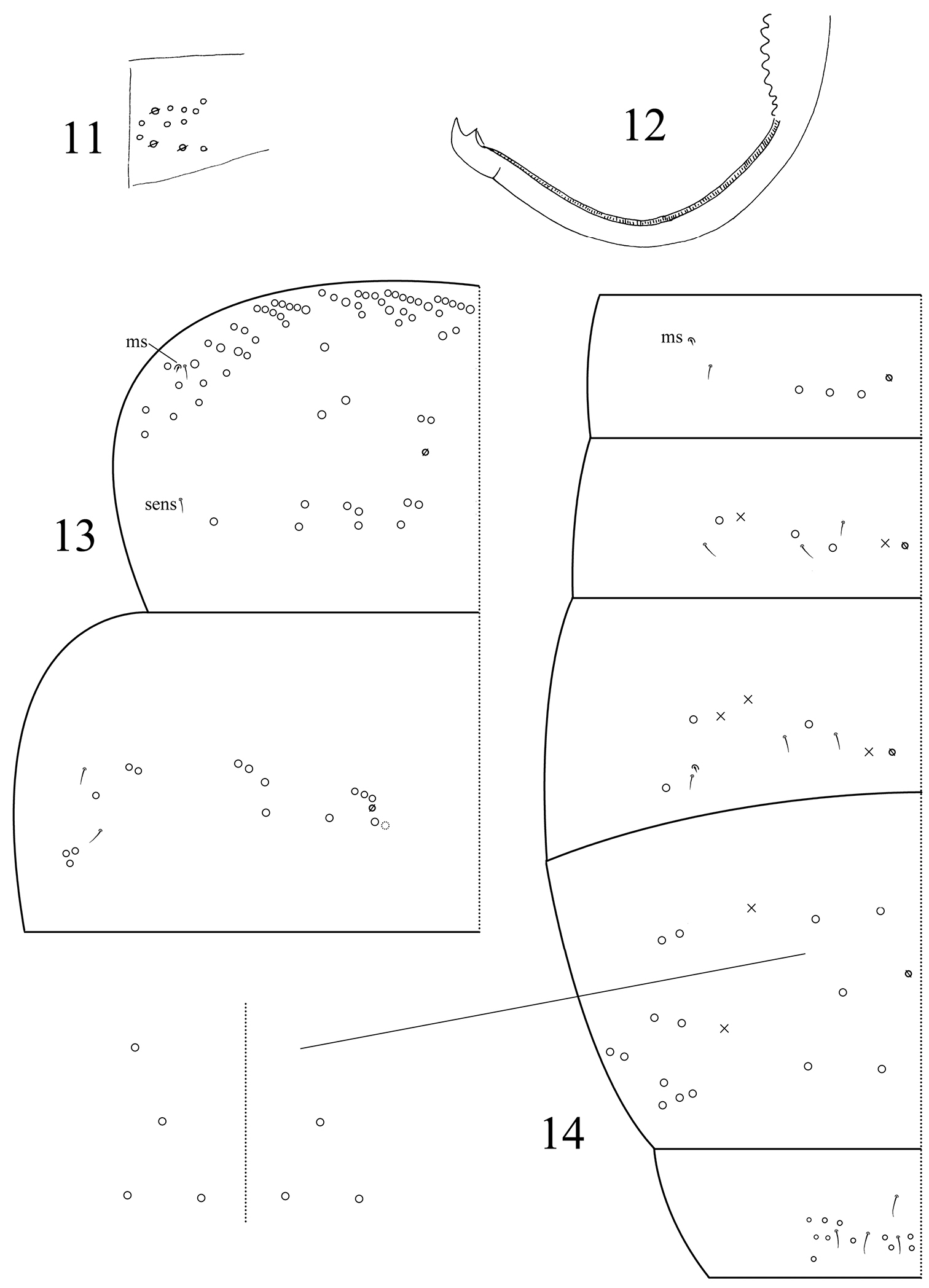
Dicranocentrus liuae sp. n. 11 manubrial plaque 12 mucro 13 thoracic chaetotaxy 14 abdominal chaetotaxy, inset, variation in number of inner mac.
Th. II with 2 inner and 2 outer mac on medial area, 9 posterior mac, 1 ms and 2 sens; anterior ms external to sens. Th. III with 15–16 mac and 2 lateral sens (Fig. 13). Abd. I with 3 mac, 1 ms and 1 sens; ms external to sens. Abd. II with 2 inner, 1 lateral mac and 3 sens. Abd. III with 1 inner, 2 lateral mac, 1 ms and 3 sens. Abd. IV with 3–5 inner, 10 lateral mac, and many (number undetermined) elongate sens. Abd. V with 4 sens (Fig. 14).
Named after the former member Ms L. Liu in our lab, who initiated the study of the genus in China.
In litter or on leaves of forest floor.
This new species belongs to sundanensis-group according to
Comparison between Dicranocentrus liuae sp. n. and Dicranocentrus wangi.
| Characters | Dicranocentrus liuae sp. n. | Dicranocentrus wangi |
|---|---|---|
| Ratio of Ant. V/VI | 0.7–1.2 | 1.2−2.3 |
| Posterior cephalic mac | 9 | 5 |
| Lateral flap of ventral tube | about 20 | 28−50 |
| Chaetae on tenaculum | 2−4 | 4−10 |
| Dental spines | absent | present |
| Inner mac on Abd. III | 1 | 2 |
The present study was supported by the National Natural Sciences Foundation of China (31101622, 40801096).