





Citation: Heads SW, Thomas MJ, Yinan Wang Y (2014) A remarkable new pygmy grasshopper (Orthoptera, Tetrigidae) in Miocene amber from the Dominican Republic. ZooKeys 429: 87–100. doi: 10.3897/zookeys.429.8020
A new genus and species of pygmy grasshopper (Orthoptera: Tetrigidae) is described from Early Miocene (Burdigalian) Dominican amber. Electrotettix attenboroughi Heads & Thomas, gen. et sp. n. is assigned to the subfamily Cladonotinae based on the deeply forked frontal costa, but is remarkable for the presence of tegmina and hind wings, hitherto unknown in this subfamily.
Orthoptera, Tetrigidae, pygmy locust, grouse locust, Hispaniola, Caribbean, amber, fossil
The Tetrigidae (pygmy grasshoppers, grouse locusts or ground hoppers) are a diverse group of small orthopterans characterized by their often remarkable morphological crypsis. These diminutive insects are primarily ground-dwelling and most commonly encountered among leaf litter on the forest floor, or in wet, marshy habitats bordering rivers, streams or standing water. With more than 1, 700 species in over 250 genera, the Tetrigidae are, among Caelifera, second only to Acrididae in terms of species diversity (
Members of the family are readily distinguished from other orthopterans by the marked posterior elongation of the pronotum which covers the entire dorsal surface of the abdomen and often extends well beyond it as an acuminate process (
Tetrigidae are extraordinarily rare in the fossil record. To date, only nine species have been described (Table 1) of which, five (including the new fossil described herein) are from fossil resins. The oldest fossil tetrigids are known from compression fossils in Early Cretaceous vulcano-sedimentary deposits of the Turga Formation, Transbaikalia, Russia.
Here, we describe a new genus and species of tetrigid from Dominican amber. Like other Dominican amber Tetrigidae, the new genus belongs to the subfamily Cladonotinae; a circumtropical group defined primarily by a deeply forked frontal costa (
Alphabetized list of fossil Tetrigidae described to date with their geological and geographical provenance. Asterisks indicate that the whereabouts of the type specimen is unknown.
| Species | Locality and age |
|---|---|
| Acrydium bachofeni Zeuner, 1937 | Baltic amber (Middle Eocene) |
| Antillotettix electrum Heads, 2009 | Dominican amber (Early Miocene) |
| Archaeotetrix locustopseiformis Sharov, 1968 | Turga Fm, Transbaikalia, Russia (Early Cretaceous) |
| Baeotettix lottiae Heads, 2009 | Dominican amber (Early Miocene) |
| Electrotettix attenboroughi Heads & Thomas, this paper | Dominican amber (Early Miocene) |
| Eotetrix unicornis Gorochov, 2012 | Green River Fm, Wyoming, USA (Middle Eocene) |
| Prototetrix reductus Sharov, 1968 | Turga Fm, Transbaikalia, Russia (Early Cretaceous) |
| Succinotettix chopardi Piton, 1938* | Baltic amber (Middle Eocene) |
| Tettigidea gracilis Heer, 1865* | Oeningen, Switzerland (Late Miocene) |
The holotype is deposited in the Paleontology Collection of the Illinois Natural History Survey (INHS), at the University of Illinois. The piece of amber contained multiple insect and plant inclusions and was cut into three pieces in order to better view the specimen. Cuts were made using a jeweler’s saw with care taken not to damage other inclusions. Facets were then ground flat and given a final polish with 50, 000 mesh diamond paste to remove visible scratches. The holotype of Electrotettix attenboroughi was studied using Olympus SZX12 zoom stereomicroscope with 1× and 2× objectives and a Zeiss SteREO Discovery V.20 stereomicroscope with 0.63× and 1.5× objectives. Photomicrographs were produced using an AxioCam HRc Rev. 3 digital camera attached to the Zeiss. Images were focus-stacked using Helicon Focus version 5.3 and the panorama was stitched in Adobe Photoshop CS5. Illustrations were produced using Adobe Illustrator CS5. The age and origin of Dominican amber is reviewed by
Electrotettix attenboroughi Heads & Thomas, gen. et sp. n.
The new genus is distinguished from all other Cladonotinae by the presence of tegmina and vestigial hind wings. Frontal costa forked just superior of antennal torulae. Anterior margin of frons at frontoclypeal margin broadly emarginate. Antennal flagellomere 3 approximately half as long as other flagellomeres. Pronotum with distinct lobe superior to humeral sinus. Posterior margin of pronotum forming a slightly upturned, blunt acuminate process.
The genus-group name is a combination of electrum (Latin from Greek, meaning “amber”) and tettix (Greek, meaning “grasshopper”).
http://zoobank.org/C2884326-0785-414A-B9AF-243A52B53F82
As for the genus (see above), by monotypy.
Female: Approximately 8.0 mm long measured from fastigium verticis to posterior apex of pronotum (Figs 1–3). Head hypognathous, robust and dorsoventrally elongate (Figs 4–5). Integument granulose; genae markedly so, bearing numerous tuberculae. Compound eyes large, globose, projecting somewhat dorsally; ventral margin acutely rounded. Vertex with low median carinula becoming lower as it crosses the fastigium, and two stronger, well-defined lateral carinae forming small dorsolaterally produced fastigial horns between compound eyes. Lateral foveae deep, longer than wide, deeper anteriorly than posteriorly. Fastigium verticis not projecting anteriorly beyond compound eyes. Interocular distance c. 0.30 mm. Frontal costa nascent immediately beneath fastigium, becoming prominent c. 0.25 mm from fastigial ridge and bifurcating at the lateral ocelli, diverging into two prominent ridge-like costal lobes between antennal torulae and ending at median ocellus. Frontal carina bifurcating c. 0.52 mm beneath median ocellus. Fronto-clypeal margin distinct, broadly emarginate Clypeus narrow; anterior margin with broad, shallow emargination. Labrum shield-like, markedly larger than clypeus with rounded apex. Mandible robust. Gena somewhat inflated with strongly granulose/tuberculate ornament; delimited anteriorly by a deep subocular furrow running the entire length of the fronto-genal region. Antennae filiform, with at least ten flagellomeres. Scape subcylindrical, approximately twice as large as pedicel and somewhat compressed laterally. Pedicel subspherical, narrower than scape but wider than flagellomeres. Flagellum at least 1.24 mm long. Flagellomeres cylindrical, longer than wide; flagellomere 3 approximately half as long as the others.
Pronotum robust, c. 6.55 mm long, with coarsely granulose ornament; anterior margin with small tectate process extending slightly above vertex of head; posterior process almost reaching apex of abdomen and terminating in a blunt and slightly upturned acuminate tip. Median carina forming distinct keel. Lateral carinae well-developed. Transverse sulci distinct, crossing and cutting the lateral carinae but not cutting the median carina. Thoracic sterna robust. Tegmen present, scale-like, longer than wide; venation indistinct, comprising numerous closed cells. Hind wing approx. 2.5 mm long, tightly folded (Fig. 6); costal lobe well-developed; Sc reaching costal margin almost reaching apex of wing; R and M entirely fused, running very close to Sc; area between R+M and Cu with numerous crossveins; CuA not visible; CuP approximating running close to 1A for its entire length; anal veins numerous.
Profemur 1.75 mm long, subquadrate in section with poorly developed carinae. Protibia at least 1.5 mm long and markedly more slender than profemur. Protarsus largely obscured by bubbles and debris in the amber. Mesofemur quadrate in section and similar in length to profemur but with carinae well-developed and complete for entire length of femur. Mesotibia 1.5 mm long, more slender than mesofemur but not as slender as protibia. Mesobasitarsus 0.2 mm long with bilobed euplantulae; second tarsomere 0.7 mm long, apically inflated with two strong pretarsal claws. Metafemur large and robust (Fig. 7), 5.5 mm long, with prominent upper and lower carinulae, dorsal keel and ventral carinae; superior and inferior marginal areas with transverse patches of rugose integument separated by smooth cuticle; medial area with prominent herringbone ornamentation comprising roughly diamond-shaped ‘cells’ of smooth cuticle, delimited by raised areas of rugose integument; femoral lobe low and distinctly rounded, not forming a spine; genicula bulbous with prominent dorsal process. Metatibia 5.0 mm long with prominent genicular bulb; 6 inner and 6 outer robust dorsal spines; 2 inner and 2 outer curved apical spurs, with inner spurs longer than outer spurs. Metabasitarsus robust, 1.0 mm long, with distinct, dorsal apical spine and two bilobed euplantulae situated in its basal half; second tarsomere much shorter, 0.3 mm long; third tarsomere almost as long as basitarsus, distinctly curved and somewhat inflated apically, bearing two pretarsal claws.
Abdomen at least 4.0 mm long, though apical damage prevents accurate measurement. Subgenital apically bilobed. Ovipositor approximately 1.0 mm long, with strong denticles on dorsal valvulae and fewer, smaller denticles on the ventral valvulae. Dorsal parts of terminalia (epiproct, cerci, etc.) obscured by bubbles and detritus.
Male: Unknown.
INHS 10175, Early Miocene (Burdigalian) amber from the La Toca region, near Santiago de los Caballeros, Santiago Province, Dominican Republic. Well-preserved adult female in a piece of amber approximately 20 × 15 × 12 mm. Numerous syninclusions are also present within the piece, including: an indet. chalcid wasp (Hymenoptera: Chalcidoidea); an indet. proctotrupoid wasp (Hymenoptera: Proctotrupoidea); a single worker ant of the genus Solenopsis (Hymenoptera: Formicidae); numerous smaller ants, possibly of the genus Azteca (Hymenoptera: Formicidae); a springtail (Collembola); three net-winged midges (Diptera: Blephariceridae); numerous botanicals including a well-preserved flower bud and a leaf fragment with possible epiphytic fungus.
The specific epithet is a patronym honouring Sir David Attenborough, British naturalist and film maker, who has been an inspiration not only to the authors of this paper, but to an entire generation of natural scientists.
Electrotettix is very distinctive among Neotropical cladonotines in that it possesses tegmina and rudimentary hind wings. Although the hind wings are much reduced, they nevertheless have complete venation and demonstrate full rotation, confirming that the holotype is a brachypterous adult and not a nymph with wing pads. Electrotettix is similar to Baeotettix in the large eyes projecting dorsally above the fastigium, but differs in the smaller fastigial horns and the absence of superior lobes on the frontal costa. Both Baeotettix and Electrotettix share features in common with extant Antillean cladonotines such as Bahorucotettix, Haitianotettix, Mucrotettix and Hottotettix (

Electrotettix attenboroughi Heads & Thomas, gen. et sp. n. 1 holotype in oblique right lateral view (scale bar 1.0 mm) 2 holotype in oblique left lateral view (scale bar 1.0 mm).
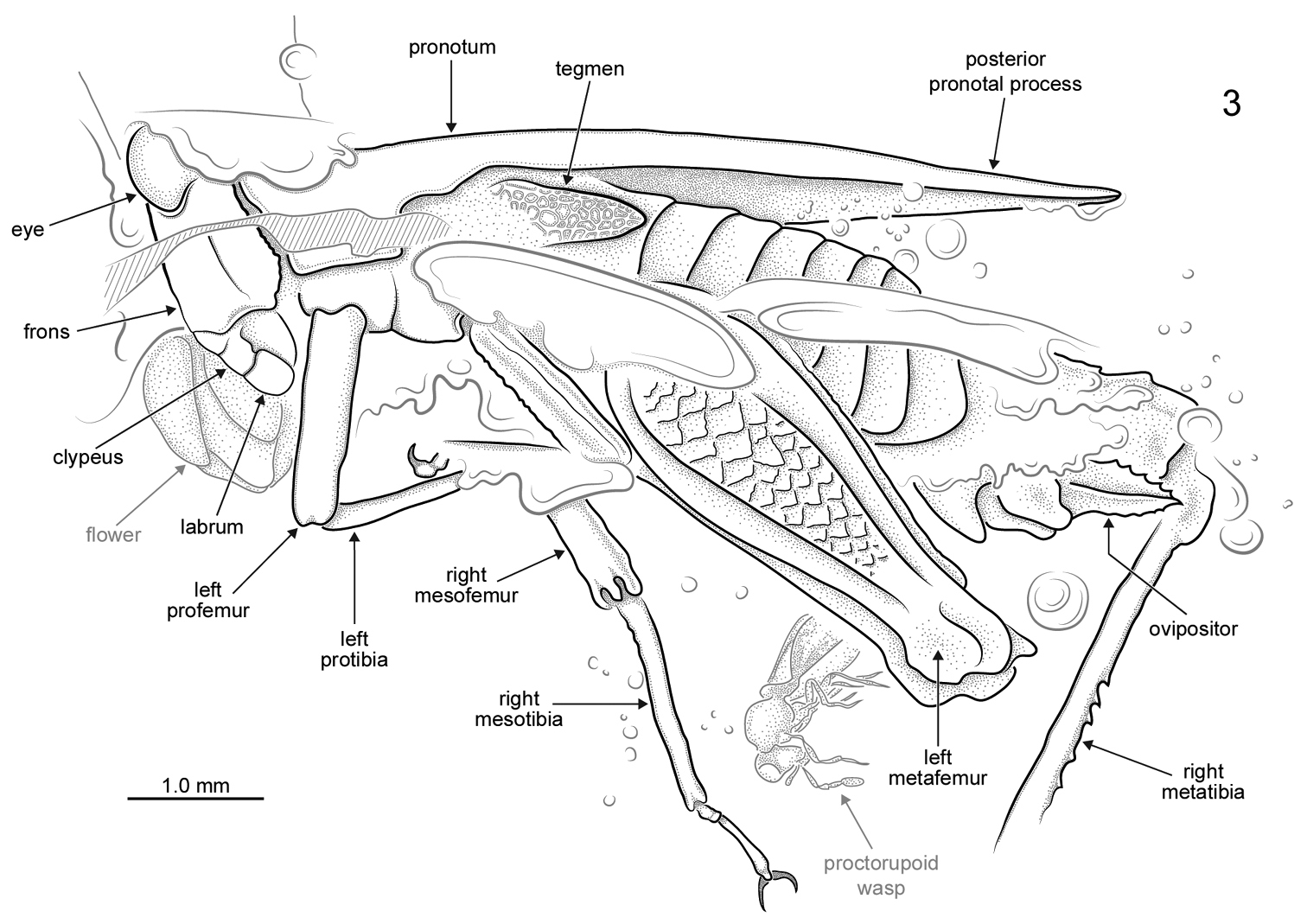
Electrotettix attenboroughi Heads & Thomas, gen. et sp. n., explanatory drawing of holotype in oblique left lateral view (scale bar 1.0 mm).

Electrotettix attenboroughi Heads & Thomas, gen. et sp. n., frontal view of head capsule; the flower bud preserved alongside the head is visible to the left (scale bar 0.5 mm).
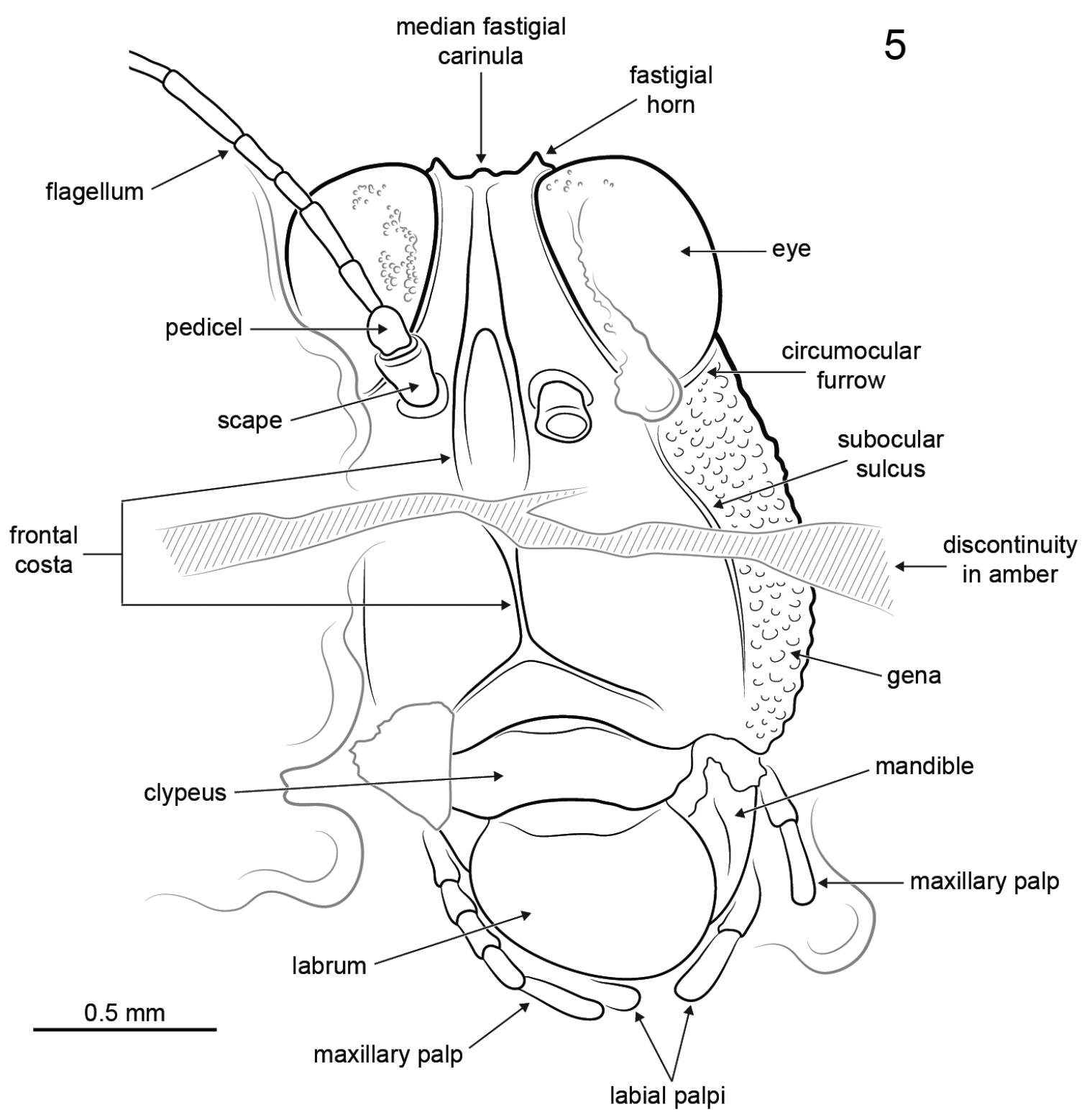
Electrotettix attenboroughi Heads & Thomas, gen. et sp. n., explanatory drawing of head capsule in frontal view (scale bar 0.5 mm).
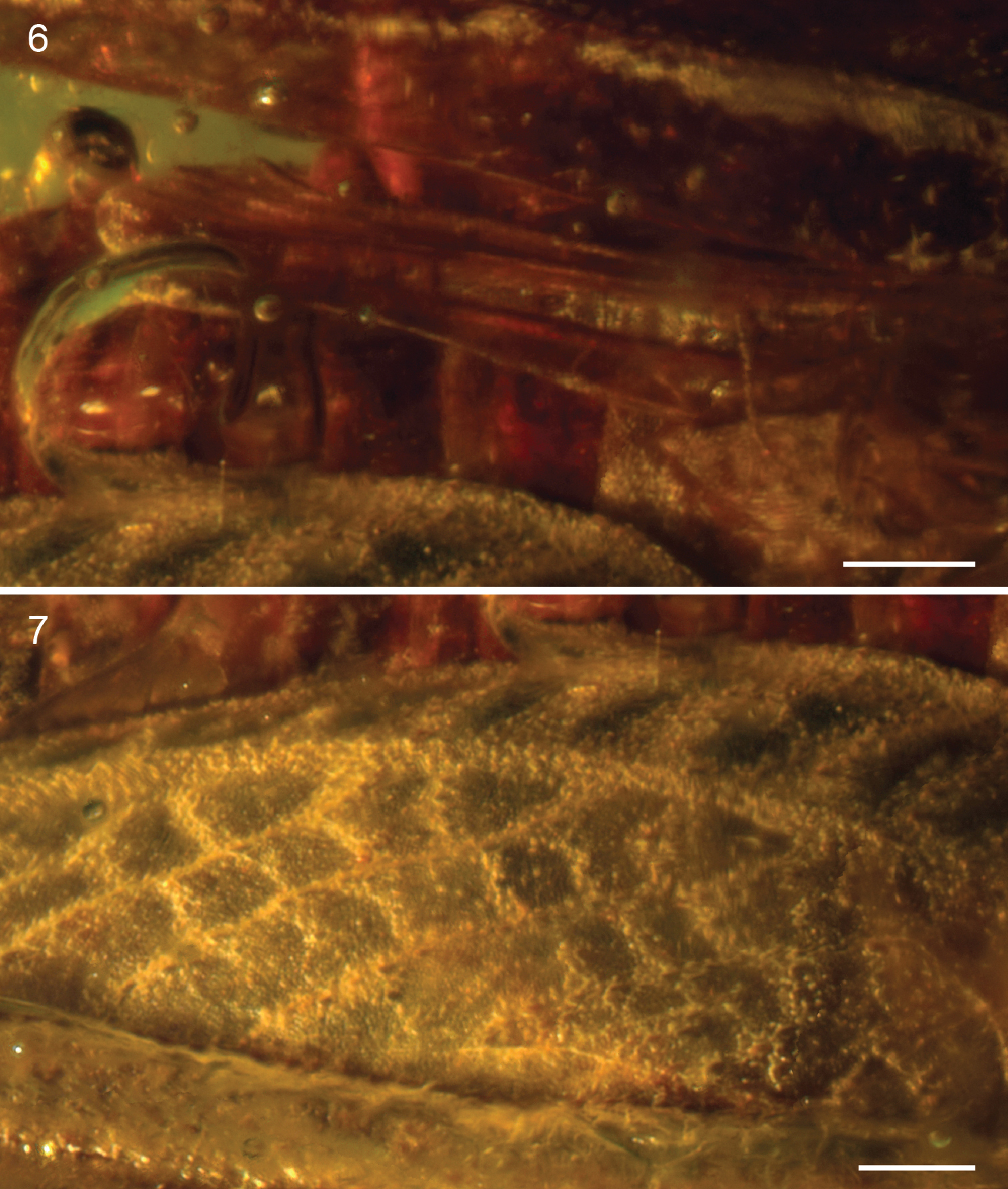
Electrotettix attenboroughi Heads & Thomas, gen. et sp. n. 6 right hind wing (scale bar 0.25 mm) 7 detail of the superior marginal and medial areas of right metafemur (scale bar 0.25 mm).
Fossil taxa are widely recognized as a valuable source of data concerning the morphology and evolution of their extant relatives. Such taxa often present novel combinations of plesiomorphic and derived character states, which provide unique insight into the acquisition and transformation of morphological characters through deep time (
While a robust comparative phylogenetic analysis is not yet available for Cladonotinae, there is evidence to support the existence of an Antillean clade characterized by coarsely granulose integument, presence of fastigial horns or tubercles, and low, non-foliaceous pronotal crests. Such a clade would comprise the fossil genera Baeotettix and Electrotettix as well as all extant Antillean cladonotines with the exception of the leaf-mimics Choriphyllum and Phyllotettix (see
The new species is named in honour of Sir David Attenborough, British naturalist and film maker, in recognition of his many important contributions to natural history. His work has been an inspiration not only to us, but to countless others, scientist and layperson alike. For their assistance in a variety of ways, we thank Mary Best, Synan Nicholson, Danielle Ruffatto, Patricia Dickerson, Mike Jeffords, Sue Post, Catherine Dana and Tina Heads (all INHS, University of Illinois). Many thanks also go to Dave Grimaldi and Paul Nascimbene (AMNH), and to Daniel Pérez-Gelabert (NMNH) for useful discussion. Partial support was provided by NSF grant EF 1304622 (to SWH).