Research Article |
|
Corresponding author: Nataliya Yu. Dnestrovskaya ( ndnestro@mail.ru ) Academic editor: Christopher Glasby
© 2020 Nataliya Yu. Dnestrovskaya.
This is an open access article distributed under the terms of the Creative Commons Attribution License (CC BY 4.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Citation:
Dnestrovskaya NYu (2020) Identification key to Nephtyidae (Annelida) of the Black Sea. ZooKeys 908: 1-17. https://doi.org/10.3897/zookeys.908.38203
|
Abstract
Currently, nine species of Nephtyidae (Annelida) are known from the Black Sea. A new user-friendly identification key is presented with a brief description for each species based on type material and recently collected specimens from the Black Sea.
Keywords
East Atlantic, Inermonephtys, Micronephthys, Nephtys, Polychaeta
Introduction
The first data on Nephtyidae of the Black Sea were given by
The Black Sea nephtyid polychaetes are small to medium-sized bristle worms. The largest species, N. caeca, may reach a length of up to 250 mm (
The illustrated key provided herein allows for the identification of Nephtyidae species reported from the Black Sea and Sea of Azov. This key is based mainly on external morphological characters, which are best viewed using a stereomicroscope. Staining with methylene blue makes all morphological characters more visible. No slide preparation or a compound microscope is needed. Each species of Nephtyidae is provided with a brief description and distribution. The names of provinces are given according to
Abbreviations:
Materials and methods
Examined type material: N. caeca:
Additional material: over 200 specimens from the Black Sea were also examined: M. longicornis (
Four species, N. caeca (
Almost all samples were first fixed in 10% formaldehyde and then transferred to 70% ethanol (24 specimens of M. longicornis were fixed directly in 70% ethanol). Specimens were stained with methylene blue (water solution) and examined using stereomicroscopy. Pharynx characters were studied on worms with a fully everted pharynx. Photographs were taken using a Carton DSZT70 stereomicroscope equipped with a MDC 320 Microscope Digital Camera. Line drawings were prepared by tracing stereomicroscope photographs in CorelDRAW. To examine the ultrastructure of chaetae, some chaetigers were dissected, critical point dried, coated with 25 nm Au-Pd and observed with a Camscan S-2 Cambridge Scanning Electron Microscope (SEM).
All the descriptions and drawings are original except for that of I. foretmontardoi (after
Results
Remarks on the key
Nephtyids are rather similar in their morphology and often difficult to distinguish. The most used taxonomic characters to separate the species are: parapodial morphology, branchiae shape, number of branchiferous chaetigers, ornamentation of the chaetae (only visible under a compound microscope), and pharynx structure. The number of the most anterior chaetiger with developed postacicular lobes was also included in descriptions, as this is an important systematic character (
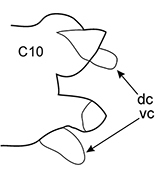
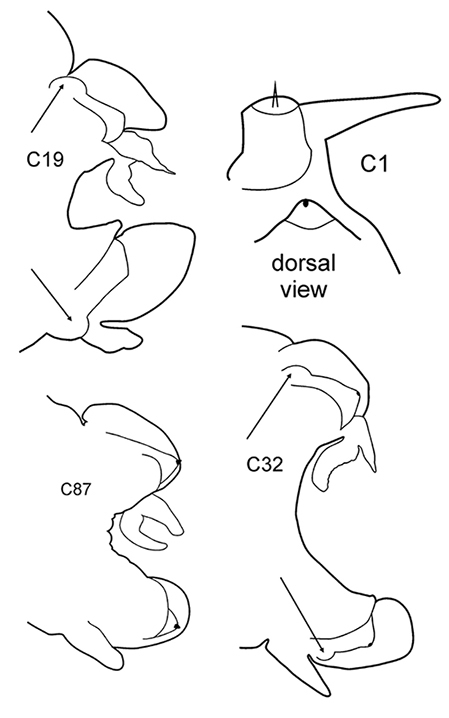
All parapodia in Nephtyidae are biramous. Both noto- and neuropodia consist of acicular, pre- and postacicular lobes, and dorsal (notopodial) and ventral (neuropodial) cirri. In Nephtys and Micronephthys species, the acicular lobes are supported by one acicula and may be conical, rounded, or bilobed (Fig.

The branchiae are inserted in the parapodia below the dorsal cirri and may be involute or recurved. They may be slender, digitiform, foliaceous, or rounded-fleshy. A small papilla may be present at the base of the branchiae under the notopodial cirrus. The shape and proportions of branchiae vary along the body, so they should be examined on the chaetigers that are recommended in the key. The chaetiger on which the branchiae begin should be checked on both sides of the worm.
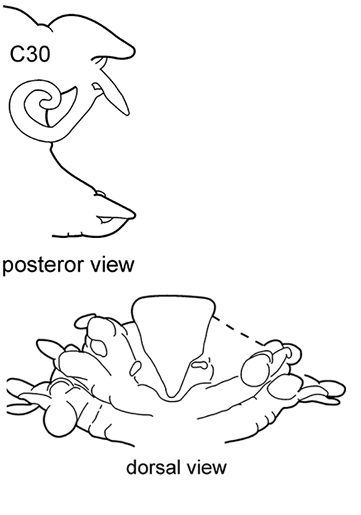
The prostomium is subquadrangular to subpentagonal (shape depends on whether the proboscis is everted or not). A pair of conical antennae is present in the anterior corners of the prostomium (absent in Inermonephtys). A pair of palps is inserted ventrolaterally. A pair of nuchal organs is located dorsolaterally on the posterior margin of the prostomium (Fig.

The pharynx is a large eversible muscular proboscis, usually covered with soft papillae located in different areas that can be seen when pharynx is everted or dissected (Fig.
Examining several specimens rather than a single individual is strongly recommended for identification. Staining with methylene blue (but not methyl blue!) will significantly highlight morphological characters of all structures.

Discussion
Nephtys caeca, N. ciliata, N. longosetosa, and N. paradoxa were absent not only in our collection from the Black Sea, but also in the collections of other museums where the Black Sea’s fauna traditionally was studied (
The mention of these species is based on identification by
The Nephtyidae of the Black Sea could be divided into two groups by the presence of different types of chaetae in the postacicular rows. The north-boreal species (N. caeca, N. ciliata, N. longosetosa, and N. paradoxa) have spinose chaetae, while south boreal-Lusitanian species (N. hombergii, and N. hystricis) have serrate chaetae with only single lateral rows of spines along one side of the chaeta (
No key is complete and perfect. The key given below should be used with caution and confirmed with descriptions of the species concerned.

Explanation of main chaetae terminology (all chaetae of postacicular rows) A, B geniculate chaeta of Nephtys cirrosa C spinose chaeta of Nephtys ciliata, frontal view D serrate chaeta of Nephtys hombergii, lateral view E lyrate chaeta of Inermonephtys foretmontardoi (after
List of Nephtyidae reported from the Black Sea
Inermonephtys Fauchald, 1968
Inermonephtys foretmontardoi Ravara, Cunha & Pleijel, 2010
Micronephthys Friedrich, 1939
Micronephthys longicornis (Perejaslavtseva, 1891)
Nephtys Cuvier, 1817
Nephtys caeca (Fabricius, 1780)
Nephtys ciliata (O.F. Müller, 1789)
Nephtys cirrosa Ehlers, 1868
Nephtys hombergii Savigny in Lamarck, 1818
Nephtys hystricis McIntosh, 1900;
Nephtys longosetosa Örsted, 1842
Nephtys paradoxa Malm, 1874
Key to nephtyid species from the Black Sea
| 1 | Branchiae absent, up to 11 mm long (usually shorter), no more than 50 chaetigers (usually 30–40) | Micronephthys longicornis | |
|
Small worms with body length up to 11 mm ( |
||
| – | Branchiae present, usually over several tens of chaetigers; up to 200 chaetigers or more and may be over 200 mm long in adults (in juveniles number of chaetigers may be low, but just before pygidium there is growth zone with numerous developing chaetigers) | 2 | |
| 2(1) | Branchiae of middle parapodia curved inward | Inermonephtys foretmontardoi | |
|
Body length more than 28.8 mm, more than 57 chaetigers (type specimen incomplete). Branchiae from C4, curved inward, thin and long, moderately ciliated, with conspicuous conical basal projections. Antennae and pharyngeal papillae absent. Nuchal organs cirriform. Preacicular lobes low, rounded. Acicular lobes rounded in anterior parapodia, conical in middle, acutely pointed in posterior. Postacicular lobes distinctly longer than acicular lobes, rounded in anterior notopodia and leaf-like in middle; always slender in neuropodia. Dorsal cirri conical in anterior parapodia, slender and cirriform in middle ones. Ventral cirri conical, as long as neuropodial postacicular lobes. Lyrate chaetae with subequal rami in postacicular rows. Anteriormost parapodia with up to 5 aciculae in neuropodia and 4 in notopodia. Pharynx smooth, without papillae (after |
||
|
– | Branchiae of middle parapodia curved outward | Nephtys 3 |
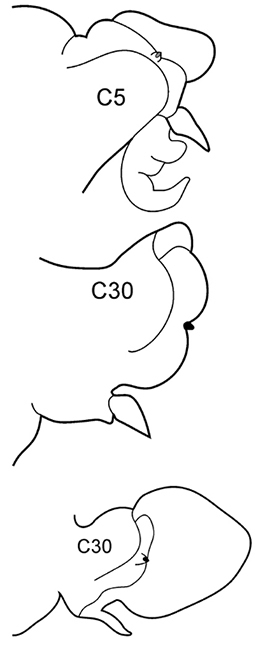
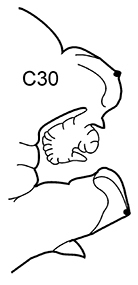
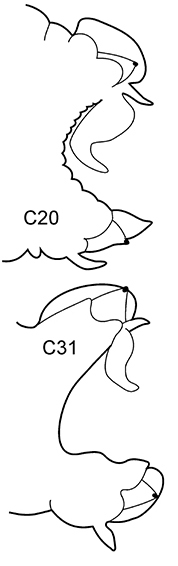
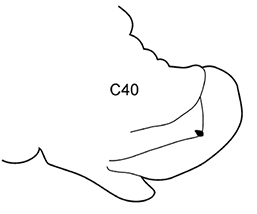
| 3(2) | Neuropodial postacicular lobes of middle chaetigers (after C30) almost equal or shorter than acicular lobes | 4 | |
| – | Neuropodial postacicular lobes of middle chaetigers (after C30) distinctly longer than acicular lobes | 6 | |
| 4(3) | In middle part of body (after C20) acicular lobes distinctly bilobed | N. ciliata | |
|
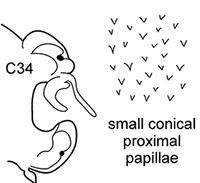
Body length up to 170 mm, up to 94 chaetigers. Branchiae from C8–C12 (rarely from C7), longer than dorsal cirri to C45–C55; decreasing in size to small knob posteriorly (shorter than dorsal cirrus), and then completely absent. Dorsal cirri of middle chaetigers long and cirriform. Notopodial preacicular lobes rudimentary, neuropodial preacicular lobes low, but distinct, in anterior and middle chaetigers surrounding acicular lobes interramally. Acicular lobes bilobed in anterior and middle region, rounded in posterior chaetigers. Postacicular lobes from C2 in neuropodia, C3 in notopodia. Notopodial postacicular lobes shorter or subequal in length to acicular lobes, neuropodial postacicular lobes subequal in length to, or slightly longer than, acicular lobes. Spinose chaetae in postacicular rows. Pharynx with long middorsal subterminal papilla, up to 7 subterminal papillae per row, proximal region covered with small conical papillae. Arcto-boreal, mainly lower shelf; reported from Black Sea near Bosphorus ( |
||
|
– | In middle part of body (after C20) acicular lobes rounded or conical | 5 |
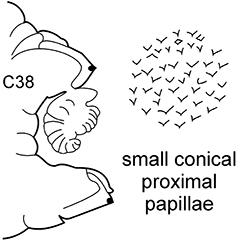
| 5(4) | Preacicular lobes rounded and rudimentary. Branchiae from C5–C20, very minute at first, in the middle chaetigers often (not always) more or less foliaceous | N. paradoxa | |
|
Body length up to 200 mm, up to 150 chaetigers ( |
||
| – | Preacicular lobes of middle chaetigers (between C15 and C45) distinctly bilobed. Branchiae from C6–C7 | N. hystricis | |
|
Body length up to 50 mm ( |
||
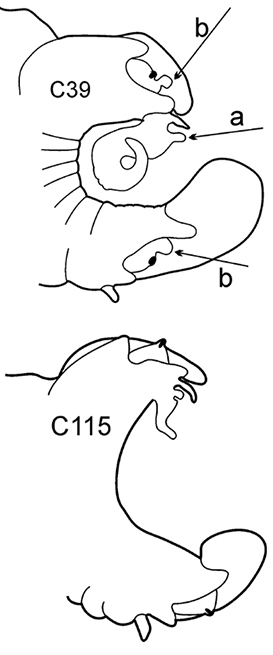
| 6(3) | In middle chaetigers: preacicular lobes distinctly bilobed, branchiae with visible basal outgrows under dorsal cirri (arrow a) | N. hombergii | |
|
Body length up to 200 mm up to 200 chaetigers ( |
||
| Remark: interramal outgrows in acicular lobes may be poorly expressed in specimens from Azov Sea and Kerch Strait. | |||
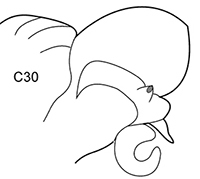
|
– | In middle chaetigers preacicular lobes rounded (rarely slightly bilobed in notopodia of very large worms). Branchiae without basal outgrows under dorsal cirri | 7 |
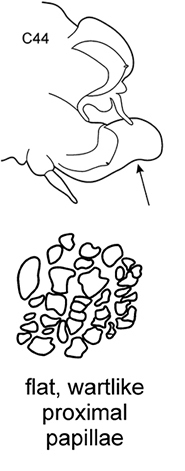
| 7(6) | Neuropodial postacicular lobes of middle chaetigers with distinct indentation on ventral side, distal to acicular lobes (arrow). Branchiae from C3 (rarely from C4; usually in small worms) | N. longosetosa | |
|
Body length up to 174 mm, up to 121 chaetigers. Branchiae from C3 to near posterior end. Preacicular lobes rounded, rudimentary, slightly bilobed in notopodia of very large worms. Acicular lobes of anterior chaetigers (and in middle chaetigers in large worms) bilobed. Postacicular lobes from C2 in neuropodia, C3 in notopodia. Notopodial postacicular lobes in anterior parapodia longer than acicular lobes, unequally bilobed with larger dorsal part; in middle and posterior parapodia shorter or slightly longer than acicular lobes. Neuropodial postacicular lobes of middle and posterior chaetigers much longer than acicular and notopodial lobes, with rounded tips and distinct indentation (arrow) on ventral side (best visible around C40). Spinose chaetae in postacicular rows. Pharynx with long middorsal subterminal papilla, up to 7 subterminal papillae per row, proximal region smooth or covered with flat warts in large specimens. Amphiboreal and Lusitanian, shelf depths, reported from Black Sea near Bosphorus ( |
||
|
– | In middle chaetigers: no indentation on ventral side of neuropodial postacicular lobes (distal to acicular lobes). Branchiae always from C4 or later | 8 |
| 8(7) | Dorsal cirri in C1 poorly developed or even absent; notopodial cirri in posterior chaetigers as long as branchiae or longer; in middle chaetigers postacicular lobes distally longer (but not twice as long) than acicular lobes; aciculae lobes of noto- and neuropodia unequally bilobed (near C25–C35); proximal region of pharynx smooth; geniculate chaetae in postacicular rows | N. cirrosa | |
|
Body length up to 110 mm ( |
||
| – | C1 with distinct dorsal cirri; notopodial cirri in all chaetigers half the length of the branchiae or even shorter; in middle chaetigers postacicular lobes more than twice as long as acicular lobes; aciculae lobes of noto-and neuropodia without any external outgrowths; proximal region of pharynx covered with small warts; geniculate chaetae absent | N. caeca | |
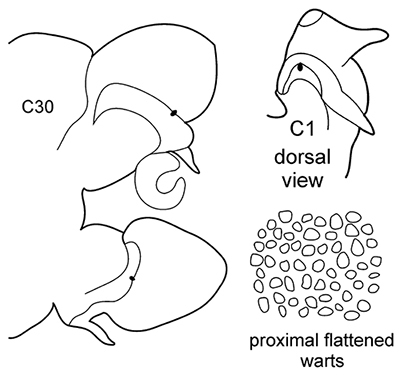
|
Body length up to 250 mm, up to 150 chaetigers ( |
||
Acknowledgements
The author is highly grateful to Dr A. Baldinger and Dr H. Dean (Museum of Comparative Zoology of Harvard University, Cambridge, MA, United States); Dr G. Paterson, Mr A.I. Muir and Ms E. Sherlock (National History Museum, London, United Kingdom) for the opportunity to work with type material. I would like to thank Professor A.I. Azovsky and Dr I. Jirkov (Dept of General Ecology and Hydrobiology, Biology Faculty Lomonosov Moscow State University, Russia), Dr G. Davidovich and Dr A. Bogdanov (User Facilities Center of Lomonosov Moscow State University, Russia); Dr G. Kolyuchkina (P.P. Shirshov Institute of Oceanology Russian Academy of Sciences, Moscow, Russia), Dr V. Semin (Institute of Arid Zones of Southern Scientific Center of the Russian Academy of Sciences, Rostov-on-Don, Russia), L.A. Zhivoglyadova (FSBSI “Azov Sea Research Fisheries Institute”); Dr S.Yu. Gagaev and Dr V.V. Potin (Zoological Institute of Russian Academy of Sciences, St Petersburg, Russia); Dr O. Bondarenko (Institute of Marine Biology of the NAS of Ukraine, Odessa, Ukraine); Dr. E. Lisitskaya (A.O.Kovalevsky Institute of Marine Biological Research of RAS, Sevastopol, Russia); Dr D. Hall and Dr T.Worsfold (APEM, Letchworth, United Kingdom); Dr J. Kongsrud (National History Museum, Bergen, Norway) for the numerous consultations, useful advice, critical remarks, equipment provided and for the loan of material. Special thanks are due to Mr A.I. Muir (National History Museum, London, United Kingdom) for correcting my English and many thanks to the reviewers, whose valuable comments highly improved this article.
This study was financially supported by a State Research Program granted to Moscow State University (project no. AAAA-A16-116021660062-9), and partially performed at the User Facilities Center of Moscow State University under financial support of the Ministry of Education and Science of the Russian Federation.
References
- Banse K (1959) Polychaeten aus Rovinj (Adria). Zoologischer Anzeiger 162: 296–313.
- Bobretsky N (1870) [Report of zoological researches conducted at the shore of the Black Sea in Summer 1869. In: Borschov I (Ed. ) Notes of the Kiev Society of Naturalists] 1: 1–18 [in Russian]
- Briggs JC, Bowen BW (2012) A realignment of marine biogeographic provinces with particular reference to fish distributions. Journal of Biogeography 39: 12–30. https://doi.org/10.1111/j.1365-2699.2011.02613.x
- Çinar ME, Dağlü E, Şahin GK (2014) Checklist of Annelida from the coasts of Turkey. Turkish Journal of Zoology 38: 734–764. https://doi.org/10.3906/zoo-1405-72
- Dnestrovskaya NYu (2013) The new species character of Nephtyidae (Polychaeta), taxonomic significance of morphological characters. “Marine biology, geology, oceanography – interdisciplinary research in marine hospitals”, dedicated to the 75th anniversary of the White Sea Biological Station of Moscow State University (Moscow, Feb 27 – Mar 1, 2013). KMK Scientific Press, Moscow, 8–12.
- Dnestrovskaya NYu, Jirkov IA (2001) Nephtyidae Grube 1850. In: Jirkov IA (Ed.) Polychaeta of the North Polar Basin. Yanus-K, Moscow, 181–212. [in Russian]
- Dnestrovskaya NYu, Jirkov IA (2011) Microscopical studies of nephtyid chaetae (Annelida: Polychaeta: Nephtyidae). Proceedings of the 10th International Polychaete Conference, Lecce (Italy), 20–26 June 2010. Italian Journal of Zoology 78(S1): 219–229. https://doi.org/10.1080/11250003.2011.589175
- Dnestrovskaya NYu, Jirkov IA (2019) Redescription of Micronephthys longicornis (Perejaslavtseva, 1891) (Annelida: Nephtyidae). Zootaxa 4550(3): 391–400. https://doi.org/10.11646/zootaxa.4550.3.6
- Ehlers E (1868) Die Borstenwürmer (Annelida Chaetopoda) Nach Systematischen und Anatomischen Untersuchungen Dargestellt. Wilhelm Engelmann, Leipzig, 748 pp. https://doi.org/10.5962/bhl.title.2081
- Fabricius O (1780) Fauna Groenlandica, Systematice Sistens, Animalia Groenlandiae Occidentalis Hactenus Indagata, Quoad Nomen Specificum, Triviale, Vernaculumque Synonyma Auctorum Plurium, Descriptionem, Locum, Victum, Generationem, Mores, Usum, Capturamque Singuli; Prout Detegendi Occasio Fuit, Maximaque Parte Secundum Proprias Observations. Hafniae et Lipsiae, Copenhagen, 452 pp. https://doi.org/10.5962/bhl.title.13489
- Fauvel P (1923) Polychètes Érrantes. Faune de France 5. Librairie de la Faculté des Sciences. Lechevallier, Paris, 488 pp. http://www.faunedefrance.org/
- Foret-Montardo P (1969) Étude systématique et écologique des Nephtyidae (Polychètes Errantes) des parages de Marseille PhD thesis, Marseille, France: Station marine d’Endoume 1(3): 807–832.
- Golikov AN, Dolgonenko MA, Maximovich NV, Scarlato OA (1990) Theoretical approaches to marine biogeography. Marine Ecology Progress Series 63: 289–301. https://doi.org/10.3354/meps063289
- Hartman O (1950) Polychaetous annelids. Goniadidae, Glyceridae and Nephtyidae. Allan Hancock Pacific Expeditions 15(1): 1–181. https://www.biodiversitylibrary.org/item/24858#page/7
- Hartmann-Schröder G (1996) Annelida, Borstenwürmer, Polychaeta. 2. neubearbeitete Auflage. Tierwelt Deutschlands 58. VEB Gustav Fischer, Jena, 648 pp.
- Ivanov VA, Belokopytov VN (2011) [Oceanography of the Black Sea. NAS of Ukraine. Marine Hydrophysical Institute, Sevastopol], 212 pp. [in Russian]
- Jirkov I (2018) Three new species of Thelepus Leuckart, 1849 from Europe and a re-description of T. cincinnatus (Fabricius, 1780) (Annelida, Terebellidae). ZooKeys 759: 29–56. https://doi.org/10.3897/zookeys.759.22981
- Jumars PA, Dorgan KM, Lindsay SM (2015) Diet of worms emended: an update of polychaete feeding guilds. Annual Review of Marine Science 7: 497–520. https://doi.org/10.1146/annurev-marine-010814-020007
- Kiseleva MI (2004) [Polychaetes (Polychaeta) of the Azov and Black Seas. Kola Science Centre RAS, Apatity] 409 pp. [in Russian]
- Laborda AJ (2004) Familia Nephtyidae Grube, 1850. In: Vieitez JM, Alos C, Parapar J, Besteiro C, Moreira J, Nunez J, Laborda AJ, San Martin G (Eds) Annelida, Polychaeta I. Fauna Iberica 25. Museo Nacional de Ciencias Naturales, CSIC, Madrid, 390–419.
- Lamarck JB (1818) Histoire Naturelle des Animaux Sans Vertèbres, Presentant les Caractères Generaux et Particuliers de ces Animaux, Leur Distribution, Leurs Classes, Leurs Familles, Leurs Genres, et la Citation des Principales Espèces qui s’y Rapportent; Précédés d’une Introduction Offrant la Détermination des Caractères Essentiels de L’animal, sa Distinction du Végétal et des Autres Corps Naturels, Enfin L’exposition des Principes Fondamentaux de la Zoologie 5. Deterville, Paris, 612 pp. http://biodiversitylibrary.org/page/12886879
- Malm AW (1874) Annulata i hafvet utmed Sveriges westkust och omkring Göteborg. Göteborgs Königlich vetenskaps–och Vitterhets Samhälles Handlingar 14: 67–105.
- Marinov T (1977) [Bristle Worms (Polychaeta). Fauna Bulgarica 6, Bulgarian Academy of Sciences, Sofia], 260 pp. [in Bulgarian]
- McIntosh WC (1900) Notes from the Gatty Marine Laboratory, St. Andrews. On the Nephthydidae of the Gulf of St. Lawrence, Canada XX (4). Annals and Magazine of Natural History (Series 7) 5: 254–268. https://doi.org/10.1080/00222930008678281
- Müller OF (1789) Zoologia Danica seu Animalium Daniae et Norvegiae Rariorum ac Minus Notorum Descriptiones et Historia, Tertium. Havniae, Copenhague, 274 pp. https://doi.org/10.5962/bhl.title.49477
- Örsted AS (1842) Udtag af en beskrivelse af Grønlands annulata dorsibranchiata. Naturhistorisk Tidsskrift 4: 109–127. http://www.biodiversitylibrary.org/item/19563#page/117/
- Perejaslavtseva SM (1891) [Supplementations to the fauna of the Black Sea. Proceedings of the Society of Naturalists at the Imperial University of Kharkov] 25: 235–274. [in Russian]
- Rainer SF (1990) The genus Nephtys (Polychaeta: Phyllodocida) in northern Europe: redescription of N. hystricis and N. incisa. Journal of Natural History 24: 361–372. https://doi.org/10.1080/00222939000770271
- Rainer SF (1991) The genus Nephtys (Polychaeta: Phyllodocida) of northern Europe: a review of species, including the description of N. pulchra sp. n. and a key to the Nephtyidae. Helgolander Meeresuntersuchungen 45: 65–96. https://doi.org/10.1007/BF02365637
- Ravara A, Cunha MR, Pleijel F (2010) Nephtyidae (Annelida, Polychaeta) from southern Europe. Zootaxa 2682: 1–68. https://doi.org/10.11646/zootaxa.2682.1.1
- Ravara A, Rizzo A, Lana P (2017) Nephtyidae Grube, 1850. In: Schmidt-Rhaesa A (Ed.) Handbook of Zoology Online. Berlin, Boston, De Gruyter. https://www.degruyter.com/view/Zoology/bp_029147-6_30 [Accessed on: 2017-9-19]
- Rullier F (1963) Les annélides polychètes du Bosphore, de la mer de Marmara et de la mer Noire, en relation avec celles de la Mediterranée. Extrait des Rapports et Procès-verbaux des réunions de la CIESMM 17: 161–260.
- Şahin GK, Çinar ME (2012) A check-list of polychaete species (Annelida: Polychaeta) from the Black Sea. Journal of the Black Sea/Mediterranean Environment 18(1): 10–48.
- San Martin G (1982) Una nueva especie de Nephtyidae (polyquetos: errantes) del Mediterraneo: Micronephthys maryae n. sp. Cachiers de Biologie Marine 23(4): 427–434.
- Schubert A, Reise K (1986) Predatory effects of Nephtys hombergii on other polychaetes in tidal flat sediments. Marine Ecology Progress Series 34: 117–124. https://doi.org/10.3354/meps034117
- Vinogradov KA, Losovskaya GV (1968) [Phylum Annelida: class Polychaeta]. In: Morduchai-Boltovskoi FD (Ed.) [Keys to the Fauna of the Black Sea and the Sea of Azov. Volume 1]. Naukova Dumka Press, Kiev, 251–359. [in Russian]