





Citation: Hita Garcia F, Fisher BL (2014) The ant genus Tetramorium Mayr in the Afrotropical region (Hymenoptera, Formicidae, Myrmicinae): synonymisation of Decamorium Forel under Tetramorium, and taxonomic revision of the T. decem species group. ZooKeys 411: 67–103. doi: 10.3897/zookeys.411.7260
In this study we synonymise the genus Decamorium Forel under Tetramorium Mayr, revise the new T. decem species group by providing a diagnosis of the group, an illustrated identification key to species level, and worker-based species descriptions for all five species, which include diagnoses, discussions, images, and distribution maps. The following species are revised in this study: T. decem Forel, comb. r., T. raptor sp. n., T. uelense Santschi, comb. r., T. ultor Forel, comb. r., stat. r. & stat. n., and T. venator sp. n. In addition, we also designate lectotypes for T. decem, T. uelense, and T. ultor.
Afrotropical region, Decamorium, taxonomy, Tetramoriini, Tetramorium, T. decem species
The genus Tetramorium Mayr is globally distributed and with 520 valid species it represents one of the most species-rich ant genera (
In this study we propose Decamorium as a junior synonym of Tetramorium and lower it to the arbitrary rank of a species group. Our decision is based on a critical analysis of the diagnostic characters previously defining Decamorium. In addition, we revise the alpha taxonomy of the Tetramorium decem species group. A diagnosis of the Tetramorium decem species group is given together with an illustrated identification key to species on the basis of the worker caste. In addition, all members of the species group are described/re-described including diagnoses, discussions, high-quality montage images and distribution maps.
The collection abbreviations follow
BMNH The Natural History Museum (British Museum, Natural History), London, U.K.
CASC California Academy of Sciences, San Francisco, U.S.A.
LACM Natural History Museum of Los Angeles County, Los Angeles, U.S.A.
MCZ Museum of Comparative Zoology, Harvard University, Cambridge, Massachusetts, U.S.A.
MHNG Muséum d’Histoire Naturelle de la Ville de Genève, Geneva, Switzerland
MNHN Muséum National d’Histoire Naturelle, Paris, France
MSNG Museo Civico di Storia Naturale “Giacomo Doria”, Genova, Italy
NHMB Naturhistorisches Museum, Basel, Switzerland
NMK National Museums of Kenya, Nairobi, Kenya
ZFMK Zoological Research Museum Alexander Koenig, Bonn, Germany
Most of the material examined in this study is located in the Hymenoptera collections of BMNH, CASC, MCZ, MHNG, and LACM. It includes much historical material collected prior to Bolton’s review of Decamorium (
HL Head length: maximum distance from the midpoint of the anterior clypeal margin to the midpoint of the posterior margin of head, measured in full-face view. Impressions on the anterior clypeal margin and the posterior head margin reduce head length.
HW Head width: width of the head directly behind the eyes measured in full-face view.
SL Scape length: maximum scape length excluding basal condyle and neck.
EL Eye length: maximum diameter of compound eye measured in oblique lateral view.
PW Pronotal width: maximum width of the pronotum measured in dorsal view.
WL Weber’s length: diagonal length of the mesosoma in lateral view from the posteroventral margin of propodeal lobe to the anterior-most point of pronotal slope, excluding the neck.
PSL Propodeal spine length: the tip of the measured spine, its base, and the centre of the propodeal concavity between the spines must all be in focus. Using a dual-axis micrometer the spine length is measured from the tip of the spine to a virtual point at its base where the spine axis meets orthogonally with a line leading to the median point of the concavity.
PTH Petiolar node height: maximum height of the petiolar node measured in lateral view from the highest (median) point of the node to the ventral outline. The measuring line is placed at an orthogonal angle to the ventral outline of the node.
PTL Petiolar node length: maximum length of the dorsal face of the petiolar node from the anterodorsal to the posterodorsal angle, measured in dorsal view excluding the peduncle.
PTW Petiolar node width: maximum width of the dorsal face of the petiolar node measured in dorsal view.
PPH Postpetiole height: maximum height of the postpetiole measured in lateral view from the highest (median) point of the node to the ventral outline. The measuring line is placed at an orthogonal angle to the ventral outline of the node.
PPL Postpetiole length: maximum length of the postpetiole measured in dorsal view.
PPW Postpetiole width: maximum width of the postpetiole measured in dorsal view.
OI Ocular index: EL / HW * 100
CI Cephalic index: HW / HL * 100
SI Scape index: SL / HW * 100
DMI Dorsal mesosoma index: PW / WL * 100
LMI Lateral mesosoma index: PH / WL * 100
PSLI Propodeal spine index: PSL / HL * 100
PeNI Petiolar node index: PTW / PW * 100
LPeI Lateral petiole index: PTL / PTH * 100
DPeI Dorsal petiole index: PTW / PTL * 100
PpNI Postpetiolar node index: PPW / PW * 100
LPpI Lateral postpetiole index: PPL / PPH * 100
DPpI Dorsal postpetiole index: PPW / PPL * 100
PPI Postpetiole index: PPW / PTW * 100
Pubescence and pilosity are often of high diagnostic value within the genus Tetramorium (e.g.
Tetramorium Mayr, 1855: 423. Type species: Formica caespitum, by subsequent designation of
Tetrogmus Roger, 1857: 10. Type species: Tetrogmus caldarius, by monotypy. [Tetrogmus junior synonym of Tetramorium:
Xiphomyrmex Forel, 1887: 385 [as subgenus of Tetramorium]. Type species: Tetramorium (Xiphomyrmex) kelleri, by subsequent designation of Wheeler, W.M. 1911: 175. [Xiphomyrmex junior synonym of Tetramorium:
Triglyphothrix Forel, 1890: cvi. Type species: Triglyphothrix walshi, by monotypy. [Triglyphothrix junior synonym of Tetramorium:
Atopula Emery, 1912: 104. Type species: Atopomyrmex nodifer, by original designation. [Atopula junior synonym of Tetramorium:
Decamorium Forel, 1913a: 121 [as subgenus of Tetramorium]. Type species: Tetramorium (Decamorium) decem, by monotypy. [Decamorium raised to genus:
Macromischoides Wheeler, W.M. 1920: 53. Type species: Macromischa aculeata, by original designation. [Macromichoides Santschi, 1924: 206, incorrect subsequent spelling.] [Macromischoides junior synonym of Tetramorium:
Lobomyrmex Kratochvíl, 1941: 84 [as subgenus of Tetramorium]. Type species: Tetramorium (Lobomyrmex) ferox silhavyi (junior synonym of Tetramorium ferox), by monotypy. [Lobomyrmex junior synonym of Tetramorium:
Sulcomyrmex Kratochvíl, 1941: 84 [as subgenus of Tetramorium]. Unavailable name. Proposed without designation of type species and therefore unavailable. Species included by
Apomyrmex Calilung, 2000: 66. Type species: Apomyrmex manobo, by original designation. [Apomyrmex junior synonym of Tetramorium:
Decamorium Forel–a junior synonym of Tetramorium Mayr
As outlined in the introduction, in the past various authors expressed very different opinions about the status of Decamorium. After examination of all available material and dissemination of all previous literature, we have come to the conclusion that Decamorium is best treated as a junior synonym of Tetramorium. Our reasons are summarised below:
As outlined above, the antennomere count was the main diagnostic character qualifying Decamorium as a genus (
In what is now considered to be Tetramorium one can find eleven-segmented and twelve-segmented antennae throughout all biogeographical regions, even though most of these forms were previously separated into Xiphomyrmex (11-segmented antennae) and Tetramorium (12-segmented antennae).
This is further supported by the presence of a very small species from India that possesses 10-segmented antennae: Tetramorium decamerum (Forel). This species was treated as Triglyphothrix by
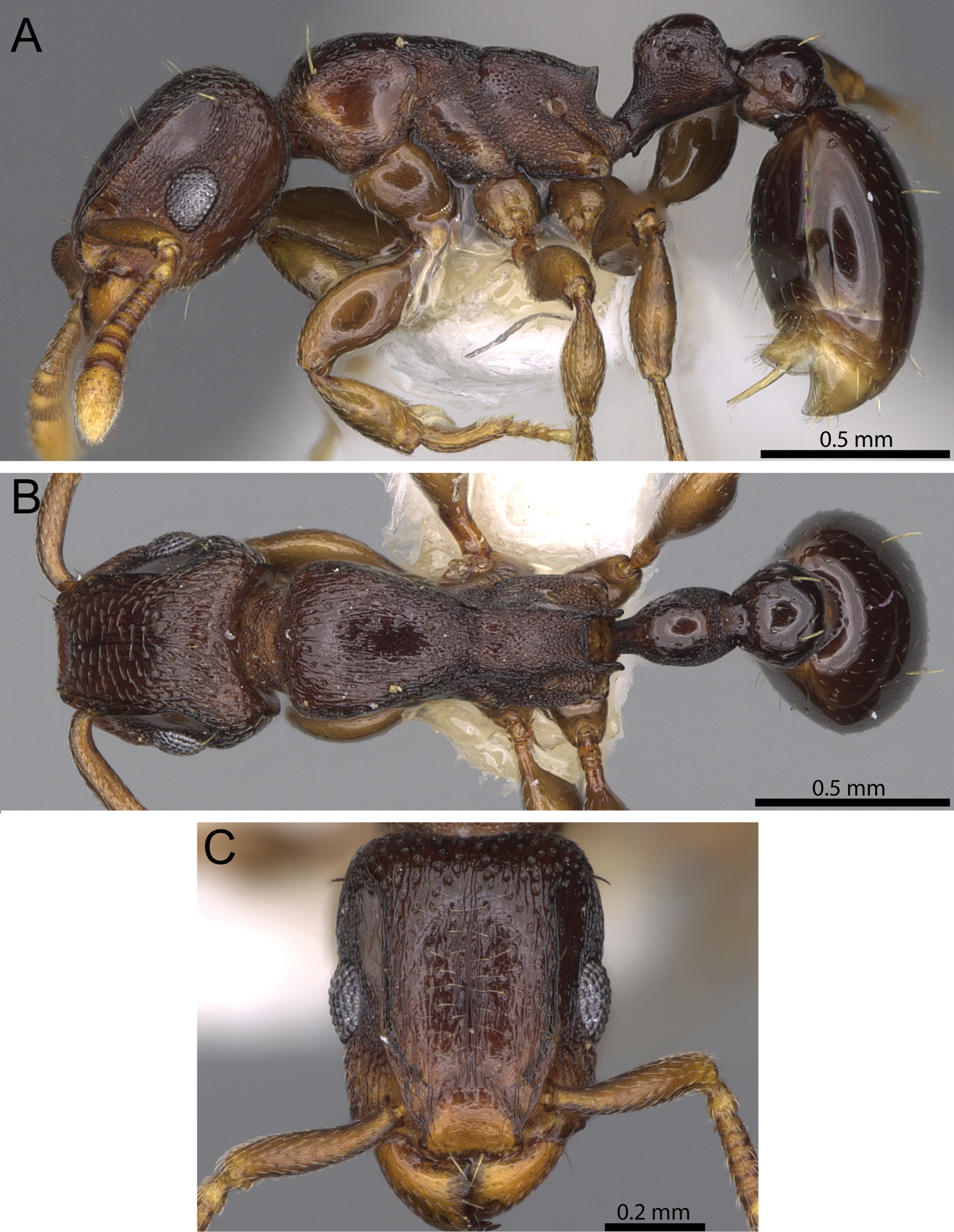
The reduced clypeal shield seen in Decamorium (Fig. 1A) is not unique to its species. Within the tropical Tetramorium fauna most species have a very well-developed and clearly distinctive clypeal shield (Figs 1F, 1G, 1F, 1I), but there are a number of species, such as Tetramorium nodiferum (Emery) (Fig. 1B), Tetramorium simulator Arnold (Fig. 1C), Tetramorium aculeatum (Mayr) (Fig. 1D), or Tetramorium anodontion Bolton (Fig. 1E), in which this shield is much less pronounced or almost reduced. The clypeal shield generally varies from species to species in its height and the sharpness of its dorsal edge. When the development of this character across several hundred Tetramorium species is considered, Decamorium emerges as one extreme of a cline that ranges from almost no clypeal shield to a very sharp and high shield, such as in the members of the Tetramorium sericeiventre Emery species group (Fig. 1I).

Anterior head showing varying development of the clypeal shield. A Tetramorium decem (CASENT0914088) B Tetramorium nodiferum (CASENT0217218) C Tetramorium simulator (CASENT0914089) D Tetramorium aculeatum (CASENT0235778) E Tetramorium anodontion Bolton (CASENT0102334) F Tetramorium diomandei Bolton (CASENT0901166) G Tetramorium hecate Hita Garcia & Fisher (CASENT0248334) H Tetramorium melanogyna Mann (CASENT0199931) I Tetramorium sericeiventre (CASENT0235773).
The mandibular dentition of Decamorium and Tetramorium seemed slightly different back in 1976, but as anticipated by Bolton, it has become clear that there is much more variation within Tetramorium. Currently there is no significant difference in mandibular dentition between Decamorium and Tetramorium. In Decamorium the mandibular count consists of three apical teeth followed by a series of four or five denticles, while in Tetramorium there are two to three apical teeth followed by a series of three to eight denticles (
If one considers the whole tribe Tetramoriini, then it becomes apparent that the specialised habitus of Decamorium is not unique. Several authors have stated that Decamorium are specialised termite hunters, and that their specialised morphology could be an adaptation to such a dangerous lifestyle (

Head in full-face view and body in profile. A, B Tetramorium decem (CASENT0914087) C, D Tetramorium simulator (CASENT0914089).
We do not intend to go into details of male morphology here, but so far there is not a single character that would separate the males of Decamorium from the males of Tetramorium; a result that agrees with Bolton’s findings (
In addition to our morphological analysis above, there is also molecular evidence supporting the synonymisation of Decamorium under Tetramorium. Based on a multi-gene dataset,
Tetramorium decem Forel, 1913a, comb. r.
Tetramorium raptor Hita Garcia, sp. n.
Tetramorium uelense Santschi, 1923, comb. r.
Tetramorium ultor Forel, 1913b, comb. r., stat. r. & stat. n.
Tetramorium venator Hita Garcia, sp. n.
Ten-segmented antennae; antennal scape relatively short (SI 67–76); anterior clypeal margin with distinct but often shallow impression; frontal carinae strongly developed and noticeably raised, forming dorsal margin of very well-developed antennal scrobes, curving down ventrally and anteriorly halfway between posterior eye margin and posterior head margin and forming posterior and usually ventral scrobe margins; antennal scrobes very well developed, deep and usually with clearly defined margins all around, median scrobal carina absent; eyes relatively large (OI 32–40); mesosoma relatively flat, low, and elongated, margination between lateral and dorsal mesosoma moderately developed (LMI 33–38); propodeum armed with short triangular to elongate-triangular teeth (PSLI 9–19); propodeal lobes short, rounded to triangular; tibiae and femorae strongly swollen; petiolar node nodiform with moderately rounded antero- and posterodorsal margins, petiolar dorsum weakly to strongly convex, node in profile between 1.0 to 1.3 times higher than long (LPeI 77–100), node in dorsal view around 1.1 to 1.3 times longer than wide (DPeI 76–92); postpetiole in profile globular, around 1.1 to 1.4 times higher than long (LPpI 71–88); mandibles and clypeus unsculptured, smooth, and shiny; sculpture on cephalic dorsum between frontal carinae and dorsal mesosoma variable, ranging from unsculptured, smooth, and shiny to longitudinally rugose/rugulose, often punctate or puncticulate; petiole usually weakly sculptured, postpetiole unsculptured to weakly sculptured; gaster unsculptured, smooth, and shiny; pilosity greatly reduced, head with several pairs of standing hairs, mesosoma with one pair, waist segments sometimes with one long pair each, and sometimes first gastral tergite with one pair; sting appendage triangular.
The Tetramorium decem species group is endemic to the Afrotropical region where it is widely distributed (Fig. 3). Tetramorium raptor and Tetramorium uelense are found in West and Central Africa and Tetramorium venator occurs through most of the equatorial rainforest belt from Liberia in West Africa to Western Kenya. By contrast, Tetramorium decem and Tetramorium ultor are species from eastern and southeastern Africa. Surprisingly, the group seems to be absent from South Africa based on the material available to us, but Tetramorium decem or Tetramorium ultor are likely to be found there or in neighbouring Botswana or Namibia. Furthermore, we expect the distribution ranges of Tetramorium decem, Tetramorium uelense, and perhaps Tetramorium ultor to expand with further ant inventory or collecting projects in Afrotropical savannahs, dry forests, and other arid habitats. These were sparsely sampled in sub-Saharan Africa in the past since most modern ant inventories have focused on rainforests (e.g.
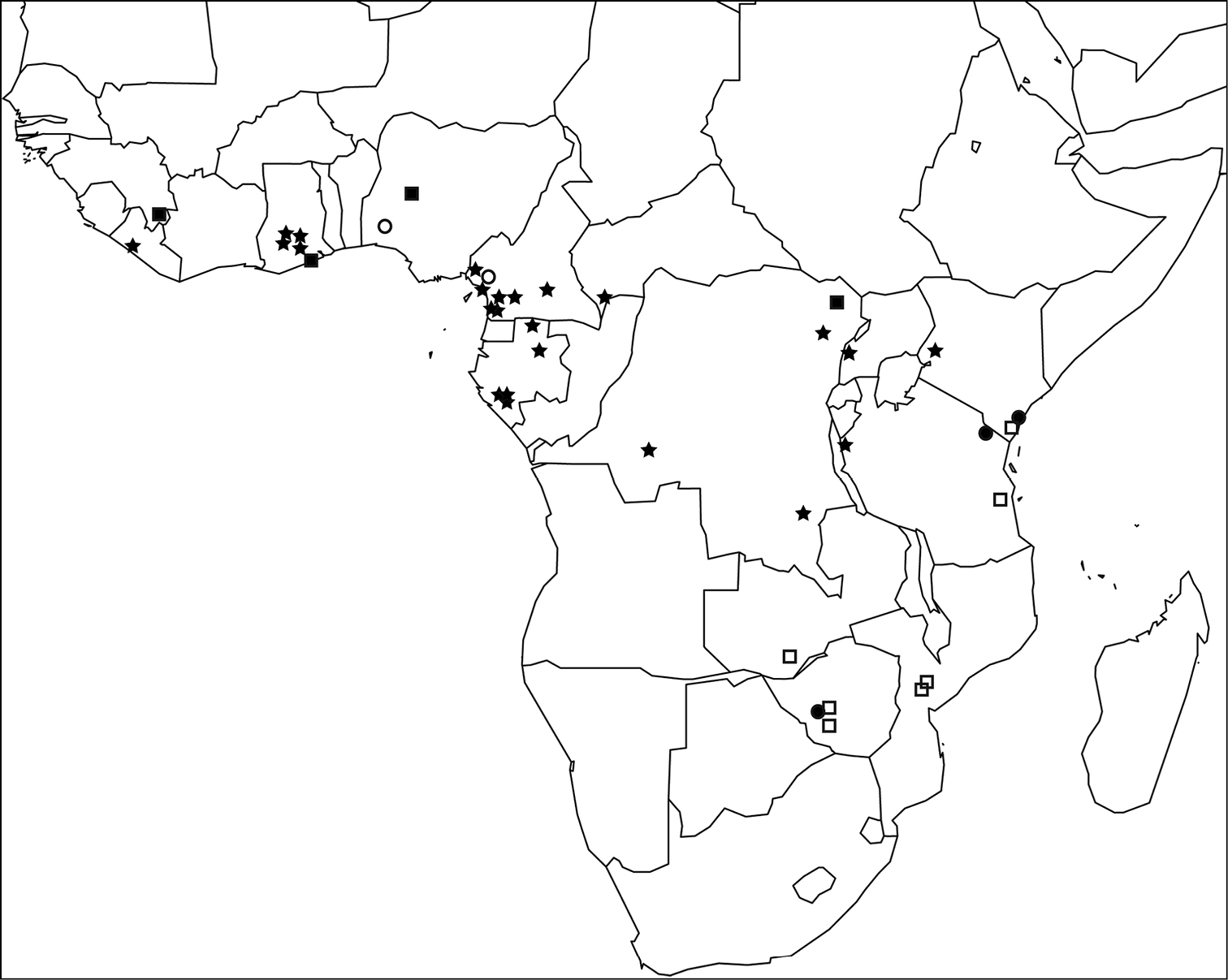
Map of sub-Saharan Africa showing the known distribution ranges of the five members of the Tetramorium decem species group: Tetramorium decem (filled circle), Tetramorium raptor (empty circle), Tetramorium uelense (filled square), Tetramorium ultor (empty square), and Tetramorium venator (star).
The separation of the Tetramorium decem species group from all other Tetramorium species groups is straightforward and easy. So far, only the members of the Tetramorium decem group have ten-segmented antennae, whereas all other Afrotropical Tetramorium have either eleven or twelve. Consequently, the Tetramorium decem species group is unlikely to be confused with another Afrotropical group. The morphology of the five species of the group is very uniform, likely due to their strongly specialised lifestyle, which makes the taxonomy of the group challenging at first sight. However, good diagnostic characters separate them fairly well from each other, especially eye size, propodeal spine/teeth length, petiolar node shape, mesosomal sculpture, and body colouration. These characters are remarkably consistent within each species throughout its whole distribution, as are the species-specific habitat preferences.
| 1 | Dorsum of promesonotum with conspicuous longitudinally rugose/rugulose sculpture (Fig. 4A, B) | 2 |
| – | Dorsum of promesonotum unsculptured, smooth, and usually very shiny (Fig. 4C, D) | 3 |
| 2 | Slightly smaller species (WL 0.88–0.93); propodeum armed with shorter, triangular, and acute teeth (PSLI 10–11); dorsum of promesonotum longitudinally rugulose with very little ground sculpture, lateral pronotum mostly unsculptured and shiny, only dorsally longitudinally rugulose; generally of uniform dark brown colour; rainforest species (Fig. 5A, B) [Cameroon, Nigeria] | Tetramorium raptor |
| – | Slightly larger species (WL 0.98–1.06); propodeum armed with longer, triangular to elongate-triangular, and acute teeth (PSLI 16–18); dorsum of promesonotum and lateral pronotum strongly longitudinally rugose with distinct punctate ground sculpture; strongly bicoloured species with dark brown or black gaster contrasting with light brown to reddish brown on remainder of body; savannah species (Fig. 5C, D) [Cameroon, Ghana, Guinea, Nigeria, and Republic of the Congo] | Tetramorium uelense |
| 3 | Generally larger species (WL 1.02–1.16); propodeal teeth relatively longer (PSLI 17–19); petiolar node in profile relatively higher, in profile 1.2 to 1.3 times higher than long (LPeI 77–82); strongly bicoloured species with dark brown or black gaster contrasting with light brown to reddish brown remainder of body (Fig. 6A) [Kenya, Tanzania, and Zimbabwe] | Tetramorium decem |
| – | Generally smaller species (WL 0.85–0.98); propodeal teeth relatively shorter (PSLI 9–13); petiolar node relatively lower, in profile around 1.0 to 1.2 times higher than long (LPeI 86–100); usually of uniform brown colour, if bicoloured, then only slightly so and never as well developed as above (Fig. 6B) | 4 |
| 4 | Smaller eyes (OI 33–36); body colouration uniformly light brown to chestnut brown (Fig. 7A, B) [Kenya, Mozambique, Tanzania, Zambia, and Zimbabwe] | Tetramorium ultor |
| – | Larger eyes (OI 37–40); body colouration uniformly dark brown to black, always darker than above (Fig. 7C, D) [Central African Republic, Cameroon, Democratic Republic of Congo, Gabon, Ghana, Kenya, Liberia, Tanzania, Uganda] | Tetramorium venator |

Mesosoma in dorsal view. A Tetramorium raptor (CASENT0195628) B Tetramorium uelense (CASENT0914084) C Tetramorium ultor (CASENT0235465) D Tetramorium venator (CASENT0401714).

Body in profile and mesosoma in dorsal view. A, B Tetramorium raptor (CASENT0280848) C, D Tetramorium uelense (CASENT0914084).

Body in profile. A Tetramorium decem (CASENT0914088) B Tetramorium venator (CASENT0195574).

Head and body in profile. A, B Tetramorium ultor (CASENT0235465) C, D Tetramorium venator (CASENT0401714).
Lectotype [designated here], pinned worker, ZIMBABWE, Redbank, 19.98333S, 28.37759E, 7.IV.1912 (G. Arnold) (MHNG: CASENT0909196) [examined]. Paralectotypes [designated here], seven pinned workers with same data as lectotype (BMNH: CASENT0901035; MHNG: CASENT0248316; MSNG: CASENT0904789) [examined].
[Note: the GPS data of the type locality was not provided by the locality label or the original description. The data presented above is based on our own geo-referencing of the town of Redbank located in the Matabeleland North Province. Consequently, the location should be considered as an approximation and not the exact position of the type locality.]
KENYA: Coastal Province, Malindi District, Arabuko Sokoke Forest, 3.28S, 39.97E, 75 m, Brachystegia forest, 26.V.2001 (R.R. Snelling & D.J. Martins); Coastal Province, Malindi District, Arabuko Sokoke Forest, 3.32111S, 39.92944E, ca. 50 m, VI.2009 (F. Hita Garcia & G. Fischer); TANZANIA: Mkomazi Game Reserve, Ibaya, 3.96667S, 37.8E, in burnt grassland, 19.–20.XI.1994 (A. Russel-Smith).

Tetramorium decem non-type worker (CASENT0914088). A Body in profile B Body in dorsal view C Head in full-face view.
Tetramorium decem can be recognised by the following combination of characters: relatively larger species (HW 0.59–0.62; WL 1.02–1.16); propodeal teeth relatively longer (PSLI 17–19); petiolar node in profile around 1.2 to 1.3 times higher than long (LPeI 77–82); dorsum of promesonotum unsculptured, smooth, and very shiny; strongly bicoloured species with dark brown or black gaster contrasting with light brown to reddish brown remainder of body.
(N=15). HL 0.71–0.74 (0.72); HW 0.59–0.62 (0.60); SL 0.42–0.45 (0.43); EL 0.19–0.21 (0.20); PH 0.33–0.37 (0.35); PW 0.47–0.50 (0.48); WL 1.02–1.16 (1.06); PSL 0.12–0.14 (0.13); PTL 0.25–0.27 (0.26); PTH 0.31–0.34 (0.33); PTW 0.22–0.24 (0.23); PPL 0.24–0.27 (0.25); PPH 0.32–0.36 (0.34); PPW 0.32–0.36 (0.34); CI 83–85 (84); SI 70–76 (72); OI 32–34 (33); DMI 41–47 (45); LMI 32–34 (33); PSLI 17–19 (18); PeNI 46–51 (48); LPeI 77–82 (80); DPeI 85–92 (88); PpNI 67–76 (70); LPpI 71–77 (75); DPpI 128–138 (133); PPI 143–149 (147).
Head much longer than wide (CI 83–85); posterior head margin weakly concave. Anterior clypeal margin with distinct, but often shallow median impression. Frontal carinae strongly developed and noticeably raised forming dorsal margin of very well-developed antennal scrobes, curving down ventrally and anteriorly halfway between posterior eye margin and posterior head margin and forming posterior and parts of ventral scrobe margins; antennal scrobes very well developed, deep and with clearly defined margins, but ventral margin less strongly developed, median scrobal carina absent. Antennal scapes short, not reaching posterior head margin (SI 70–76). Eyes very large (OI 32–34). Mesosomal outline in profile flat to weakly convex, relatively low and elongate (LMI 32–34), moderately to strongly marginate from lateral to dorsal mesosoma; promesonotal suture absent; metanotal groove present, distinct, and clearly impressed. Propodeal spines short, elongate-triangular, and moderately acute (PSLI 17–19), propodeal lobes short, triangular, and usually blunt, always significantly shorter than propodeal spines. Tibiae and femorae strongly swollen. Petiolar node nodiform with moderately rounded antero- and posterodorsal margins, around 1.2 to 1.3 times higher than long (LPeI 77–82), anterior and posterior faces approximately parallel, anterodorsal and posterodorsal margins situated at about the same height, petiolar dorsum clearly convex; node in dorsal view between 1.1 to 1.2 times longer than wide (DPeI 85–92), in dorsal view pronotum around 2.0 to 2.2 times wider than petiolar node (PeNI 46–51). Postpetiole in profile globular to subglobular, approximately 1.3 to 1.4 times higher than long (LPpI 71–77); in dorsal view around 1.3 to 1.4 times wider than long (DPpI 128–138), pronotum between 1.3 to 1.5 times wider than postpetiole (PpNI 67–76). Postpetiole in profile usually appearing less voluminous than petiolar node, postpetiole in dorsal view around 1.4 to 1.5 times wider than petiolar node (PPI 143–149). Mandibles and clypeus usually fully unsculptured, smooth, and shining, mandibles sometimes with few traces of rugulae apically; cephalic dorsum between frontal carinae mostly unsculptured and shiny, median ruga present and distinct, cephalic dorsum also puncticulate to punctate throughout its length, posteriorly close to posterior head margin especially pronounced; scrobal area partly unsculptured, smooth and shiny and partly merging with surrounding rugose sculpture on sides of head. Ground sculpture on head usually weak to absent. Dorsum of mesosoma mostly unsculptured, smooth and shiny with scattered punctures, rarely with few traces of rugulae; lateral mesosoma longitudinally rugose and very conspicuously reticulate-punctate except for mostly unsculptured lateral pronotum and katepisternum. Forecoxae unsculptured, smooth, and shining. Petiolar node and postpetiole superficially longitudinally rugulose or irregularly rugulose superimposed on conspicuous but relatively weak reticulate-punctate ground sculpture. Mesosoma and waist segments appearing mostly matt. First gastral tergite unsculptured, smooth, and shiny. Pilosity and pubescence greatly reduced: head with few pairs of moderately long, standing hairs, anterior pronotum with one long pair, waist segments sometimes with one long pair each, and sometimes first gastral tergite with one long pair; appressed pubescence present everywhere on body, but noticeable only on antennae, cephalic dorsum, legs, and first gastral tergite. Anterior edges of antennal scapes and dorsal (outer) surfaces of hind tibiae with appressed hairs. Body strongly bicoloured with dark brown to black gaster contrasting with light brown to reddish brown remainder.
The distribution range of Tetramorium decem is far smaller than previously thought (Fig. 3). Indeed, most of the material listed in the literature as Tetramorium decem or labelled as such in museum collections turned out to be either Tetramorium ultor or Tetramorium venator, while only a few collections proved to be genuine Tetramorium decem. Based on the redefined species definition, Tetramorium decem is only known from the type locality in Zimbabwe and two additional localities in East Africa: Arabuko Sokoke in Kenya and Mkomazi in Tanzania. Nevertheless, if more extensive sampling efforts are undertaken in East Africa, Tetramorium decem is likely to be found in more localities in Kenya, Tanzania, and Zimbabwe. Like Tetramorium uelense and Tetramorium ultor, Tetramorium decem prefers arid habitats, such as savannah and woodland. Based on
Tetramorium decem is the core species of the group, and was the type species for the description of the subgenus Decamorium by
Based on the available material we did not observe any significant form of intraspecific variation in Tetramorium decem.
http://zoobank.org/6A9F212B-8460-41C0-9F8C-792D9A4780C4
http://species-id.net/wiki/Tetramorium_raptor
Figs 3, 4A, 5, 9Holotype, pinned worker, CAMEROON, Sud-Ouest, Bakundu, 4.49222N, 9.375E, collection code ANTC27989, 8.XI.1990 (A. Dejean) (BMNH: CASENT0195628). Paratypes, 14 pinned workers with same data as holotype (BMNH: CASENT0195581; CASENT0195630; CASENT0195631; CASC: CASENT0195633; CASENT0195634; LACM: LACM_ENT_323500; MCZ: CASENT0195628; ZFMK: CASENT0195632).
Tetramorium raptor holotype worker (CASENT0195628). A Body in profile B Body in dorsal view C Head in full-face view.
[Note: the GPS data of the type locality was not provided by the locality label. The data presented above is based on our own geo-referencing of the Bakundu Forest located in the province Sud-Ouest. Consequently, it should be considered an approximation and not the exact position of the type locality.]
CAMEROON: Sud-Ouest, Bakundu, 4.49222N, 9.375E, 8.XI.1990 (A. Dejean); NIGERIA: Gambari, 10.VI.1969 (B. Bolton); Gambari, C.R.I.N., 17.VI.1975 (B. Taylor).
Tetramorium raptor is easily recognisable within the group on the basis of the following combination of characters: relatively smaller species (WL 0.88–0.93); very large eyes (OI 35); propodeum armed with very short triangular teeth (PSLI 10–11); petiolar node in profile around 1.1 times higher than long (LPeI 89–93); dorsum of mesosoma with longitudinally rugulose sculpture; body uniformly dark brown, appendages of lighter brown.
(N=12). HL 0.64–0.68 (0.67); HW 0.53–0.56 (0.54); SL 0.37–0.41 (0.39); EL 0.19–0.20 (0.XX); PH 0.31––0.34 (0.33); PW 0.40–0.43 (0.41); WL 0.88–0.93 (0.91); PSL 0.07–0.08 (0.07); PTL 0.23–0.25 (0.24); PTH 0.26–0.28 (0.27); PTW 0.19–0.21 (0.20); PPL 0.21–0.24 (0.22); PPH 0.25–0.28 (0.27); PPW 0.26–0.30 (0.28); CI 80–83 (82); SI 70–73 (72); OI 35; DMI 44–47 (45); LMI 35–37 (36); PSLI 10–11 (11); PeNI 47–51 (48); LPeI 89–93 (90); DPeI 80–85 (82); PpNI 64–70 (68); LPpI 81–88 (85); DPpI 123–130 (125); PPI 137–150 (142).
Head much longer than wide (CI 80–83); posterior head margin weakly concave. Anterior clypeal margin with distinct but often shallow median impression. Frontal carinae strongly developed and noticeably raised forming dorsal margin of very well-developed antennal scrobes, curving down ventrally and anteriorly halfway between posterior eye margin and posterior head margin and forming posterior and ventral scrobe margins; antennal scrobes very well developed, deep and with clearly defined margins all around, median scrobal carina absent. Antennal scapes short, far from reaching posterior head margin (SI 70–73). Eyes relatively large (OI 35). Mesosomal outline in profile relatively flat, elongate and low (LMI 35–37), moderately to strongly marginate from lateral to dorsal mesosoma; promesonotal suture absent; metanotal groove present and conspicuous, but relatively shallow. Propodeum armed with short, triangular, and usually acute teeth (PSLI 10–11), propodeal lobes short, well rounded, and usually larger than propodeal teeth. Petiolar node nodiform with moderately rounded antero- and posterodorsal margins, in profile around 1.1 times higher than long (LPeI 89–93), anterior and posterior faces approximately parallel, anterodorsal and posterodorsal margins situated at about same height and equally angled, petiolar dorsum weakly convex; node in dorsal view around 1.2 to 1.3 times longer than wide (DPeI 80–85), in dorsal view pronotum around 2.0 to 2.2 times wider than petiolar node (PeNI 47–51). Postpetiole in profile globular, approximately 1.1 to 1.2 times higher than long (LPpI 81–88); in dorsal view around 1.2 and 1.3 times wider than long (DPpI 123–130), pronotum around 1.4 to 1.6 times wider than postpetiole (PpNI 64–70). Postpetiole in profile appearing less voluminous than petiolar node, postpetiole in dorsal view around 1.4 to 1.5 times wider than petiolar node (PPI 137–150). Mandibles and clypeus unsculptured, smooth, and shining; cephalic dorsum between frontal carinae with fine irregularly longitudinally rugulose sculpture, rugulae running from posterior clypeal margin to posterior head margin, often interrupted or meandering, rarely with cross-meshes, cephalic dorsum also puncticulate to punctate throughout its length, otherwise without ground sculptured; scrobal area partly unsculptured, smooth and shiny and partly strongly reticulate-punctate; lateral head mainly reticulate-rugose with weak to moderately well developed punctate ground sculpture. Dorsum of mesosoma densely longitudinally rugulose, anteriorly without much ground sculpture, posteriorly on top of strong reticulate-punctate ground sculpture; lateral pronotum and katepisternum mostly unsculptured, smooth, and shiny, remainder of lateral mesosoma irregularly rugose and very conspicuously reticulate-punctate. Forecoxae unsculptured, smooth, and shining. Petiolar node laterally reticulate-punctate, dorsum of node mostly unsculptured, smooth, and shiny; postpetiole mostly unsculptured, smooth, and shiny with scattered punctures. First gastral tergite unsculptured, smooth, and shiny. Pilosity and pubescence greatly reduced: head with few pairs of moderately long, standing hairs, anterior pronotum with one long pair, waist segments sometimes with one long pair each, and sometimes first gastral tergite with one long pair; appressed pubescence present everywhere on body, but noticeable only on antennae, cephalic dorsum, legs, and first gastral tergite. Anterior edges of antennal scapes and dorsal (outer) surfaces of hind tibiae with appressed hairs. Body uniformly dark brown to black, appendages of lighter brown.
The name of the new species is Latin and means “thief, robber, or plunderer”. It refers to the predaceous lifestyle of Tetramorium raptor. The species epithet is a nominative noun, and thus invariant.
Tetramorium raptor is currently only known from the type locality Bakundu in the southeast of Cameroon and from Gambari in south-western Nigeria (Fig. 3). Based on the minimal collection label data, Tetramorium raptor lives in rainforest leaf litter.
Tetramorium raptor is an easily distinguishable member of the Tetramorium decem group, but was not recognised until this study. Indeed, all known material was collected in 1969 and 1990, but labelled as Tetramorium uelense on the basis of the distinctive sculpture on the mesosomal dorsum. The presence of conspicuous, longitudinally rugulose sculpture on the dorsum of the promesonotum distinguishes it from Tetramorium decem, Tetramorium ultor, or Tetramorium venator, since the latter three all lack sculpture on the promesonotal dorsum. Tetramorium uelense, however, shares the presence of sculpture on the mesosomal dorsum with Tetramorium raptor, which led to the abovementioned misidentifications. Nevertheless, careful examination of all material previously listed as Tetramorium uelense revealed the presence of two morphologically and ecologically different species. The most obvious differences are body size and colour. Tetramorium uelense is strongly bicoloured and larger (WL 0.98–1.06) than the smaller and uniformly-coloured Tetramorium raptor (WL 0.88–0.93). The latter also has shorter propodeal teeth (PSLI 10–11) than Tetramorium uelense (PSLI 16–18). Furthermore, Tetramorium raptor possesses a longitudinally rugulose promesonotal dorsum with very little ground sculpture and a mostly unsculptured and shiny lateral pronotum, whereas Tetramorium uelense has a promesonotal dorsum that is longitudinally rugose with distinct punctate ground sculpture and a lateral pronotum that is conspicuously rugose with prominent ground sculpture. In addition, both species also differ in habitat choice, as Tetramorium uelense seems to prefer savannah while Tetramorium raptor lives in rainforest.
Based on material from the two known localities, there is no intraspecific variation in Tetramorium raptor.
Of uelense: lectotype, pinned worker, D. R. CONGO (Congo belge), Uelé, Vankerhovenville, 3.0N, 29.5E (Degreef) (NHMB: CASENT0906826) [examined]. Paralectotype, pinned queen with same data as lectotype (MRAC) [not examined].
Of nimba: holotype, pinned worker, GUINEA, Kéoulenta, 7.714053N, 8.331786W, St. 1 Savane, (MNHN: CASENT0914084) [examined].
[Note: GPS data for neither of the type localities was included on the locality labels or the original descriptions. The data presented above is based on our own geo-referencing of Vankerhovenville located in Province Orientale and Kéoulenta located in the Nzérékoré Region. Consequently, they should be considered approximations and not the exact positions of the type localities.]
GHANA: Greater Accra Region, Accra Metropolis District, Legon, 23.VIII.1972 (D. Leston); NIGERIA: 16 km N. of Mokwa, 16.X.1976 (C. Longhurst).

Tetramorium uelense non-type worker (CASENT0195580). A Body in profile B Body in dorsal view C Head in full-face view.
The following character combination separates Tetramorium uelense from the other species of the Tetramorium decem species group: relatively larger species (WL 0.98–1.06); propodeum armed with short triangular to elongate-triangular teeth (PSLI 16–18); petiolar node in profile around 1.1 times higher than long (LPeI 88–93); dorsum of mesosoma conspicuously longitudinally rugose with distinctive reticulate-punctate ground sculpture; strongly bicoloured with dark brown to black gaster contrasting with light brown to reddish brown remainder of body.
(N=6). HL 0.67–0.72 (0.70); HW 0.54–0.59 (0.57); SL 0.39–0.42 (0.41); EL 0.19–0.20 (0.20); PH 0.36–0.38 (0.37); PW 0.43–0.47 (0.45); WL 0.98–1.06 (1.02); PSL 0.11–0.13 (0.10); PTL 0.27–0.29 (0.28); PTH 0.29–0.32 (0.31); PTW 0.21–0.23 (0.22); PPL 0.24–0.26 (0.25); PPH 0.28–0.34 (0.31); PPW 0.30–0.33 (0.31); CI 80–83 (81); SI 69–74 (72); OI 34–35 (35); DMI 43–44 (44); LMI 35–37 (36); PSLI 16–18 (17); PeNI 48–49 (49); LPeI 88–93 (90); DPeI 77–81 (79); PpNI 69–70 (70); LPpI 75–86 (80); DPpI 122–125 (124); PPI 141–145 (143).
Head much longer than wide (CI 80–83); posterior head margin weakly concave. Anterior clypeal margin with distinct, but often shallow median impression. Frontal carinae strongly developed and noticeably raised forming dorsal margin of very well-developed antennal scrobes, curving down ventrally and anteriorly halfway between posterior eye margin and posterior head margin and forming posterior and ventral scrobe margins; antennal scrobes very well developed, deep and with clearly defined margins, but ventral margin less strongly developed, median scrobal carina absent. Antennal scapes short, far from reaching posterior head margin (SI 69–74). Eyes relatively large (OI 34–35). Mesosomal outline in profile relatively flat, elongate and low (LMI 35–37), moderately to strongly marginate from lateral to dorsal mesosoma; promesonotal suture absent; metanotal groove present, distinct, but relatively shallow. Propodeum armed with short, triangular to elongate-triangular, and acute teeth (PSLI 16–18), propodeal lobes reduced, short, and well rounded, usually shorter than propodeal teeth. Tibiae and femorae strongly swollen. Petiolar node nodiform with moderately rounded antero- and posterodorsal margins, in profile around 1.1 times higher than long (LPeI 88–93), anterior and posterior faces approximately parallel, anterodorsal and posterodorsal margins situated at about same height and equally angled, petiolar dorsum clearly convex; node in dorsal view around 1.2 to 1.3 times longer than wide (DPeI 77–81), in dorsal view pronotum between 2.0 and 2.1 times wider than petiolar node (PeNI 48–49). Postpetiole in profile globular, approximately 1.2 to 1.3 times higher than long (LPpI 75–86); in dorsal view between 1.2 and 1.3 times wider than long (DPpI 122–125), pronotum around 1.4 times wider than postpetiole (PpNI 69–70). Postpetiole in profile more or less of same volume as petiolar node, postpetiole in dorsal view around 1.4 times wider than petiolar node (PPI 141–145). Mandibles and clypeus unsculptured, smooth, and shining; cephalic dorsum between frontal carinae with fine irregularly longitudinally rugulose/rugose sculpture, rugulae/rugae often interrupted, meandering, or with cross-meshes, cephalic dorsum also puncticulate to punctate throughout its length; scrobal area strongly reticulate-punctate; lateral head mainly reticulate-rugose with weak to moderately well developed punctate ground sculpture. Ground sculpture on head usually weak, except scrobal area (see above). Dorsum of mesosoma densely longitudinally rugose on top of strong punctate ground sculpture; lateral mesosoma longitudinally rugose and very conspicuously reticulate-punctate. Forecoxae unsculptured, smooth, and shining. Petiolar node and postpetiole superficially longitudinally rugulose or irregularly rugulose superimposed on conspicuous but relatively weak reticulate-punctate ground sculpture. Mesosoma and waist segments appearing matt. First gastral tergite unsculptured, smooth, and shiny. Pilosity and pubescence greatly reduced: head with few pairs of moderately long, standing hairs, anterior pronotum with one long pair, waist segments sometimes with one long pair each, and sometimes first gastral tergite with one long pair; appressed pubescence present everywhere on body, but noticeable only on antennae, cephalic dorsum, legs, and first gastral tergite. Anterior edges of antennal scapes and dorsal (outer) surfaces of hind tibiae with appressed hairs. Body strongly bicoloured with dark brown to black gaster contrasting with light brown to reddish brown remainder.
So far, Tetramorium uelense is known from a few collections in savannah habitats throughout a relatively wide geographical range from West to Central Africa (Fig. 3). The known distribution spans Guinea through Ghana and Nigeria to the northeast of the D. R. Congo close to South Sudan and Uganda. Compared to most other Afrotropical Tetramorium species, there is a wealth of information about the natural history of Tetramorium uelense (Longhurst, 1979).
Tetramorium uelense can be easily distinguished from the remainder of the Tetramorium decem species group. The presence of longitudinally rugose sculpture on the dorsum of the mesosoma separates Tetramorium uelense immediately from Tetramorium decem, Tetramorium ultor, and Tetramorium venator. In the latter three the mesosomal dorsum is completely unsculptured, smooth, and very shiny. The only other species with sculpture on the dorsum of the mesosoma, which could be confused with Tetramorium uelense, is Tetramorium raptor. Nevertheless, both are well separable in morphology and ecology. Most obviously, Tetramorium uelense is a larger species (WL 0.98–1.06) with distinct bicolouration while Tetramorium raptor (WL 0.88–0.93) is smaller and a uniform dark brown colour. In addition, Tetramorium uelense has longer and better developed propodeal teeth (PSLI 16–18) compared to Tetramorium raptor (PSLI 10–11), even though this might be difficult to see and may require measurements to confirm. Another, more visible character is the sculpture on the mesosomal dorsum, which is strongly longitudinally rugose with distinct punctate ground sculpture in Tetramorium uelense versus longitudinally rugulose with very little ground sculpture in Tetramorium raptor. Also, the lateral pronotum of the latter is mostly unsculptured, smooth, and shiny while in Tetramorium uelense the lateral pronotum is strongly rugose with conspicuous ground sculpture.
Despite the broad distribution range, we did not observe any significant intraspecific variation in Tetramorium uelense.
Lectotype [designated here], pinned worker, ZIMBABWE, Shiloh, 19.73333S, 28.55E, 12.V.1913 (G. Arnold) (MHNG: CASENT0909197) [examined]. Paralectotypes, seven pinned workers with same data as lectotype (BMNH: CASENT0901036; MHNG: CASENT0195688) [examined].

Tetramorium ultor paralectotype worker (CASENT0901036). A Body in profile B Body in dorsal view C Head in full-face view.
[Note: the GPS data of the type locality was not provided by the locality label or the original description. The data presented above is based on our own geo-referencing of the Shiloh locality located in Matabeleland North province. Consequently, it should be considered an approximation and not the exact position of the type locality.]
MOZAMBIQUE: Sofala Province, Gorongosa National Park, Limestone Gorge, 18°57'13"S, 34°10'37.6"E, 81 m, 15.V.2012 (G.D. Alpert & E.O. Wilson); Sofala Province, Gorongosa National Park, 5 km S Chitango, 18°59'28.8"S, 34°21'10"E, 10 m, secondary forest, 1.VI.2012 (G.D. Alpert); Sofala Province, Gorongosa National Park, Centracao Outpost (Piva-Joao), 18°30'20"S, 34°29'7"E, small forest along river, 11.VI.2012 (D. Muala & T. Torcida); Sofala Province, Gorongosa National Park, WP092, 18°56.1'3.1"S, 34°23'36.7"E, 51 m, open area, 26.VI.2012 (G.D. Alpert); KENYA: Kwale District, Shimba Hills, Longomagandi National Reserve, 4.23S, 39.43E, primary hardwood forest, 2.VI.2001 (R.R. Snelling); TANZANIA: Pwani, Rufiji District, Kichi Hills Forest Reserve, 8.23889S, 38.65023E, 499 m, primary forest, 5.–7.III.2008 (P. Hawkes, Y. Mlacha, & F. Ninga); ZAMBIA: Southern Province, 16.79533S, 26.93833E, 1330 m, Choma, Gwembe Lodge, miombo woodland, 3.XII.2005 (B.L. Fisher); ZIMBABWE: Balla-Balla, 20.45S, 29.03E, 1.IV.1945; Umtali, II.1917 (G. Arnold).
Tetramorium ultor can be recognised by the following combination of characters: relatively smaller species (WL 0.85–0.96); very large eyes (OI 33–36); propodeum armed with short teeth (PSLI 10–13); petiolar node in profile around 1.1 to 1.2 times higher than long (LPeI 86–92); dorsum of promesonotum unsculptured, smooth, and very shiny; body of uniform light to chestnut brown, appendages often lighter.
(N=25). HL 0.62–0.70 (0.66); HW 0.48–0.58 (0.53); SL 0.35–0.42 (0.37); EL 0.16–0.20 (0.19); PH 0.29–0.33 (0.30); PW 0.37–0.45 (0.41); WL 0.85–0.96 (0.89); PSL 0.07–0.09 (0.08); PTL 0.22–0.25 (0.24); PTH 0.25–0.29 (0.27); PTW 0.19–0.22 (0.20); PPL 0.19–0.23 (0.21); PPH 0.25–0.30 (0.27); PPW 0.24–0.30 (0.27); CI 77–82 (80); SI 67–73 (70); OI 33–36 (35); DMI 44–48 (46); LMI 32–35 (34); PSLI 10–13 (12); PeNI 46–50 (48); LPeI 86–92 (88); DPeI 79–86 (84); PpNI 60–71 (67); LPpI 73–81 (78); DPpI 126–132 (130); PPI 130–145 (139).
Head much longer than wide (CI 77–82); posterior head margin weakly concave. Anterior clypeal margin with distinct, but often shallow median impression. Frontal carinae strongly developed and noticeably raised forming dorsal margin of very well-developed antennal scrobes, curving down ventrally and anteriorly halfway between posterior eye margin and posterior head margin and forming posterior and ventral scrobe margins; antennal scrobes very well developed, deep and with clearly defined margins all around, median scrobal carina absent. Antennal scapes short, not reaching posterior head margin (SI 67–73). Eyes very large (OI 33–36). Mesosomal outline in profile relatively flat, long and low (LMI 32–35), moderately marginate from lateral to dorsal mesosoma; promesonotal suture absent; metanotal groove present and distinct, but relatively shallow. Propodeum armed with short, triangular, and mostly blunt teeth (PSLI 10–13), propodeal lobes short, triangular, and usually blunt, in profile usually longer and more voluminous than propodeal spines. Tibiae and femorae strongly swollen. Petiolar node nodiform with moderately rounded antero- and posterodorsal margins, in profile around 1.1 to 1.2 times higher than long (LPeI 86–92), anterior and posterior faces approximately parallel, anterodorsal and posterodorsal margins situated at about same height and equally angled, petiolar dorsum weakly convex; node in dorsal view around 1.1 to 1.2 times longer than wide (DPeI 79–86), in dorsal view pronotum between 2.0 to 2.2 times wider than petiolar node (PeNI 46–50). Postpetiole in profile globular, approximately 1.2 to 1.4 times higher than long (LPpI 73–81); in dorsal view around 1.3 times wider than long (DPpI 126–132), pronotum approximately 1.4 to 1.5 times wider than postpetiole (PpNI 60–71). Postpetiole in profile appearing less voluminous than petiolar node, postpetiole in dorsal view between 1.3 to 1.5 times wider than petiolar node (PPI 130–145). Mandibles and clypeus usually fully unsculptured, smooth, and shining; cephalic dorsum between frontal carinae mostly unsculptured and shiny, median ruga present and distinct, cephalic dorsum also puncticulate to punctate throughout its length, close to posterior head margin especially pronounced; scrobal area unsculptured, smooth, and very shiny; lateral head ventral of antennal scrobe mainly reticulate-rugose; ground sculpture on head usually weak to absent. Dorsum of mesosoma mostly unsculptured, smooth, and shiny with scattered punctures, rarely with few traces of rugulae; lateral mesosoma mostly unsculptured and shiny, posteriorly irregularly rugose and conspicuously reticulate-punctate. Petiolar node and postpetiole only weakly sculptured, laterally usually superficially rugulose and punctate on lower half and more unsculptured on upper half, node dorsally mostly smooth; postpetiole mostly unsculptured, smooth, and shiny with scattered punctures. First gastral tergite unsculptured, smooth, and shiny. Pilosity and pubescence greatly reduced: head with few pairs of moderately long, standing hairs, anterior pronotum with one long pair, waist segments sometimes with one long pair each, and sometimes first gastral tergite with one long pair; appressed pubescence present everywhere on body, but noticeable only on antennae, cephalic dorsum, legs, and first gastral tergite. Anterior edges of antennal scapes and dorsal (outer) surfaces of hind tibiae with appressed hairs. Body uniformly brown, appendages often lighter.
Tetramorium ultor is widespread in eastern and southern Africa (Fig. 3). It is distributed from Kenya south to Mozambique, and also found in Zambia and Zimbabwe. Most localities are tropical dry forest habitats or miombo woodland. Also, Tetramorium ultor seems to be a ground-active species nesting in or under rotten logs and is likely termitophagous like the other group members.
Since
Based on the available material, we did not observe any intraspecific variation in Tetramorium ultor.
http://zoobank.org/02C5E77F-FFD9-4204-843C-1E541B84972A
http://species-id.net/wiki/Tetramorium_venator
Figs 3, 4D, 6B, 7C, 7D, 12Holotype, pinned worker, KENYA, Western Kenya, Kakamega Forest, Bunyala Forest Fragment, 0.37889N, 34.69917E, 1448 m, disturbed primary forest, Kakamega 2008 survey, leaf litter, pitfall trap, Transect 35, position 10 m, 1.VIII.2008 (G. Fischer) (CASC: CASENT0195574). Paratypes, six pinned workers with same data as holotype (BMNH: CASENT0195625; CASC: CASENT0217165; BMNH: CASENT0195625; LACM: CASENT0195627; MCZ: CASENT0195624; NMK: CASENT0195626; ZFMK: CASENT0195623).

Tetramorium venator holotype worker (CASENT0195574). A Body in profile B Body in dorsal view C Head in full-face view.
CAMEROON: Centre, Mbalmayo, 1.XI.1993 (N. Stork); Centre, Ottotomo, 24.IV.1986 (A. Dejean); Est, Abong Mbang, 28.VI.1988 (A. Dejean); Sud, Bondé Forest, N’kolo village, 27.5 km 155°SSE Elogbatindi, 3.22167N, 10.24667E, 40 m, rainforest, 12.IV.2000 (B.L. Fisher); Sud, Res. de Faune de Campo, Massif des Mamelles, 15.1 km 84°E Ébodjé, 2.59417N, 9.9595E, 180 m, rainforest, 4.IV.2000 (B.L. Fisher); Sud, Res. de Faune de Campo, 2.16 km 106°ESE Ébodjé, 2.56783N, 9.84433E, 10 m, littoral rainforest, 9.IV.2000 (B.L. Fisher); Sud, P. N. Campo, 43.3 km 108°ESE Campo, 2.2825N, 10.20617E, 290 m, rainforest, 7.IV.2000 (B.L. Fisher); Sud-Ouest, Bimbia Forest, 7.4 km 119°ESE Limbe, 3.98183N, 9.2625E, 40 m, 14.IV.2000 (B.L. Fisher); Sud-Ouest, Korup N. P., 6.9 km 317°NW Mundemba, 5.016N, 8.864E, 110 m, rainforest, 19.IV.2000 (B.L. Fisher); CENTRAL AFRICAN REPUBLIC: Prefecture Sangha-Mbaéré, Parc National Dzanga-Ndoki, Mabéa Bai, 21.4 km 53°NE Bayanga, 3.03333N, 16.41E, 510 m, rainforest, 1.–7.V.2001 (B.L. Fisher); DEMOCRATIC REPUBLIC OF CONGO: Epulu, 750 m, 1.38333N, 28.58333E, rainforest, 1.XI.1995 (S.D. Torti); Kikwit, Kinzambi, 27.III.1984 (A. Dejean); 44 miles E. of Kileba, 1110 m, 16.I.1958 (E.S. Ross & R.E. Leech); GABON: La Makandé Forêt des Abeilles, 1.I.–1.II.1999 (S. Lewis); Ogooué-Ivindo, Makokou, C.N.R.S., 10.VII.1974 (W. Gotwald); Ogooue-Maritime, Réserve des Monts Doudou; 24.3km 307°NW Doussala, 2.2225S, 10.40583E, 370 m, coastal lowland rainforest, 5.–12.III.2000 (S. van Noort); Ogooue-Maritime, Aire d’Exploit. Rationnelle de Faune des Monts Doudou, 24.3 km 307°NW Doussala, 2.22639S, 10.40972E, 375 m, rainforest, 6.III.2000 (B.L. Fisher); Ogooue-Maritime, Reserve de la Moukalaba-Dougoua, 12.2 km 305°NW Doussala, 2.28333S, 10.49717E, 110 m, coastal lowland rainforest, sited within forest, 24.II.-3.III.2000 (S. van Noort); Ogooue-Maritime, Reserve de Faune de la Moukalaba-Dougoua, 10.8 km 214°SW Doussala, 2.42267S, 10.54533E, 110 m, rainforest, 29.II.2000 (B.L. Fisher); Ogooue-Maritime, Reserve de Faune de la Moukalaba-Dougoua, 12.2 km 305°NW Doussala, 2.31667N, 10.53333E, 110 m, rainforest, 1.III.2000 (B.L. Fisher); Woleu-Ntem, 31.3 km 108°ESE Minvoul, 2.08N, 12.40667E, 600 m, rainforest, 7.–15.II.1998 (B.L. Fisher); GHANA: Akwapim, Tafo, 19.I.1970 (B. Bolton); Ashanti, Poano, cocoa, 9.IX.1992 (R. Belshaw); Atewa Forest Reserve, near Kibi, primary forest, 24.III.1992 (R. Belshaw); Eastern, Kade, 1.I.1992 (R. Belshaw); Enchi, 17.V.1969 (D. Leston); Esunkawkaw Forest Reserve, primary forest, 27.X.1992 (R. Belshaw); Nkawanda near Nkawkaw, secondary forest, 12.XII.1991 (R. Belshaw); Portrase, 1.III.1992 (R. Belshaw); KENYA: Western Kenya, Kakamega Forest, Bunyala Forest Fragment, 0.37889N, 34.69917E, 1448 m, disturbed primary forest, 1.VIII.2008 (G. Fischer); LIBERIA: Monrovia, 5.VII.1957 (E.S. Ross & R.E. Leech); TANZANIA: Kigoma Region, Gombe Stream National Park, 4.7S, 29.616667E, 915–1012 m, 29.XII.2009–12.I.2010 (R. O’Malley); UGANDA: Semuliki NP, 00.83556, 30.15542 ± 200 m, 676 m, rainforest, 30.–31.VII.2012 (B.L. Fisher et al.); Semuliki NP, 00.84483, 30.15052 ± 200 m, 680 m, rainforest, 2.VIII.2012 (B.L. Fisher et al.).
Tetramorium venator can be recognised by the following combination of characters: relatively smaller species (WL 0.87–0.98); very large eyes, largest in the group (OI 37–40); propodeum armed with very short, triangular, and moderately acute (PSLI 9–12); petiolar node in profile between 1.0 to 1.2 times higher than long (LPeI 90–100); dorsum of promesonotum unsculptured, smooth, and very shiny; head, mesosoma, waist segments, and gaster uniformly very dark brown to black, appendages of lighter brown.
(N=25). HL 0.64–0.71 (0.67); HW 0.51–0.59 (0.54); SL 0.38–0.43 (0.40); EL 0.19–0.22 (0.21); PH 0.30–0.36 (0.32); PW 0.39–0.45 (0.41); WL 0.87–0.98 (0.92); PSL 0.06–0.09 (0.07); PTL 0.23–0.26 (0.25); PTH 0.25–0.29 (0.26); PTW 0.18–0.22 (0.20); PPL 0.21–0.25 (0.22); PPH 0.25–0.30 (0.26); PPW 0.25–0.30 (0.26); CI 79–83 (81); SI 70–75 (74); OI 37–40 (38); DMI 43–46 (44); LMI 33–37 (35); PSLI 9–12 (10); PeNI 45–51 (48); LPeI 90–100 (93); DPeI 76–85 (80); PpNI 63–67 (65); LPpI 80–86 (84); DPpI 115–124 (119); PPI 130–144 (135).
Head much longer than wide (CI 79–83); posterior head margin weakly concave. Anterior clypeal margin with distinct, but often shallow median impression. Frontal carinae strongly developed and noticeably raised forming dorsal margin of very well-developed antennal scrobes, curving down ventrally and anteriorly halfway between posterior eye margin and posterior head margin and forming posterior and ventral scrobe margins; antennal scrobes very well developed, deep and with clearly defined margins all around, median scrobal carina absent. Antennal scapes short, not reaching posterior head margin (SI 70–75). Eyes very large (37–40). Mesosomal outline in profile relatively flat, long and low (LMI 33–37), moderately marginate from lateral to dorsal mesosoma; promesonotal suture absent; metanotal groove present and distinct, but relatively shallow. Propodeum armed with very short, triangular, and moderately acute teeth (PSLI 9–12), propodeal lobes short, triangular to rounded, and usually blunt, in profile more or less of same length as propodeal teeth and appearing more voluminous than propodeal spines. Tibiae and femorae strongly swollen. Petiolar node nodiform with moderately rounded antero- and posterodorsal margins, in profile between 1.0 to 1.2 times higher than long (LPeI 90–100), anterior and posterior faces approximately parallel, anterodorsal and posterodorsal margins situated at about same height and equally angled, petiolar dorsum usually conspicuously convex, sometimes only weakly so; node in dorsal view around 1.2 to 1.3 times longer than wide (DPeI 76–85), in dorsal view pronotum between 2.0 to 2.2 times wider than petiolar node (PeNI 45–51). Postpetiole in profile globular, approximately 1.2 times higher than long (LPpI 80–86); in dorsal view around 1.2 times wider than long (DPpI 115–124), pronotum approximately 1.5 to 1.6 times wider than postpetiole (PpNI 63–67). Postpetiole in profile appearing less voluminous than petiolar node, postpetiole in dorsal view between 1.3 to 1.5 times wider than petiolar node (PPI 130–144). Mandibles and clypeus usually fully unsculptured, smooth, and shining; cephalic dorsum between frontal carinae mostly unsculptured and shiny, median ruga present and distinct, cephalic dorsum also puncticulate to punctate across its length, close to posterior head margin especially pronounced; scrobal area unsculptured, smooth and very shiny; lateral head ventral of antennal scrobe mainly reticulate-rugose; ground sculpture on head usually weak to absent. Dorsum of mesosoma mostly unsculptured, smooth and shiny with scattered punctures, rarely with few traces of rugulae; lateral mesosoma mostly unsculptured and shiny, posteriorly irregularly rugose and conspicuously reticulate-punctate. Petiolar node and postpetiole only weakly sculptured, laterally usually superficially rugulose and punctate on lower half and more unsculptured on upper half, node dorsally mostly smooth; postpetiole mostly unsculptured, smooth and shiny with scattered punctures. First gastral tergite unsculptured, smooth, and shiny. Pilosity and pubescence greatly reduced: head with few pairs of moderately long, standing hairs, anterior pronotum with one long pair, waist segments sometimes with one long pair each, and sometimes first gastral tergite with one long pair; appressed pubescence present everywhere on body, but noticeable only on antennae, cephalic dorsum, legs, and first gastral tergite. Anterior edges of antennal scapes and dorsal (outer) surfaces of hind tibiae with appressed hairs. Head, mesosoma, waist segments, and gaster uniformly very dark brown to black, appendages of lighter brown.
The name of the new species is Latin and means “hunter” referring to the predatory lifestyle of Tetramorium venator. The species epithet is a nominative noun, and thus invariant.
Tetramorium venator is the most widespread and abundant species of the group. It is found throughout much of the equatorial forest belt from Liberia in the west to Kenya in the east (Fig. 3). Even though there was no material from Benin, Togo, Nigeria, Equatorial Guinea or South Sudan available for this study, we expect that Tetramorium venator will be found in most or all of these countries. Based on the available data, this species lives in the leaf litter stratum of primary, secondary, or disturbed rainforests. Additionally, Tetramorium venator seems to be found at lower elevations in West and Central Africa, but also occurs at mid elevations further east in the eastern D.R. Congo, Tanzania, and Kenya, where it reaches its highest known elevation at the type locality at 1448 m. Based on unpublished stable isotope data from the type series, Tetramorium venator is a predatory species, and we assume that it feeds on termites. This is supported by some series from Cameroon that were collected while foraging in the nests of Cubitermes Wasmann.
Despite being common and collected fairly often prior to this study, most of the material of Tetramorium venator was identified and labelled as Tetramorium decem. Indeed, more than 90% of all the material listed as the latter species at the beginning of our revision turned out to be Tetramorium venator. Nevertheless, our revision shows that they are clearly not conspecific. Tetramorium venator is smaller in size (WL 0.87–0.98), has larger eyes (OI 37–40), shorter propodeal teeth (PSLI 9–12), a lower petiolar node (LPeI 90–100), and has a uniform body colouration. By contrast, Tetramorium decem is larger (WL 1.02–1.16), has smaller eyes (OI 32–34), longer propodeal spines (PSLI 17–19), a higher petiolar node (LPeI 77–82), and is distinctly bicoloured. Also, Tetramorium venator is a rainforest species while Tetramorium decem lives in savannah or woodland.
The abovementioned very large eyes of Tetramorium venator separate it also from Tetramorium ultor, which has smaller eyes (OI 33–36). In addition, Tetramorium ultor is also of a much lighter colour, usually light brown to chestnut brown, and prefers dry forest or woodland habitats. It should be noted, however, that Tetramorium ultor and Tetramorium venator are morphologically very close to each other and differ significantly only in eye size, colour and habitat preference. They could represent different ecotypes of the same species, one adapted to more shaded and humid forest versus one specialised to more arid savannah, woodland, and dry forest. Nevertheless, if this was true, then we would see some intermediate forms in transitional habitats, and there are none at present. As a matter of fact, Tetramorium venator is also found in secondary and disturbed rainforests. The type series was collected in a highly disturbed rainforest fragment in Kenya and the material from Gombe in Tanzania is from a rainforest-woodland mosaic. Both species are also separated by the Great Rift Valley, which separates different faunistic sub-regions of the Afrotropical region. We consider Tetramorium venator as a faunal element of the Guineo-Congolian forest zone, while we believe Tetramorium ultor is a species of the arid corridor running from East to Southern Africa. Based on the available material and African biogeography in general, we conclude that our two species hypothesis is more likely.
Furthermore, Tetramorium venator cannot be misidentified with either Tetramorium uelense or Tetramorium raptor since both possess strongly developed rugulose/rugose sculpture on the promesonotal dorsum that is absent in Tetramorium venator. At present, Tetramorium venator overlaps in its distribution with Tetramorium uelense and Tetramorium raptor in West and Central Africa. We think it might also overlap with Tetramorium decem and Tetramorium ultor in East Africa, even though it currently seems as if they are widely separated geographically. However, since the sampling is very patchy, especially in East Africa, much more Tetramorium decem and Tetramorium ultor material is likely to be collected with further inventories, and these two species will be found in close proximity to Tetramorium venator. Nevertheless, the latter species is restricted to more humid forest habitats, whereas Tetramorium decem and Tetramorium ultor clearly prefer more arid savannah, grassland, woodland and tropical dry forest.
Intriguingly, even though Tetramorium venator is very broadly distributed in Equatorial Africa, there seems to be no significant intraspecific variation.
First we thank Michele Esposito from CAS for her help with image processing and databasing. We are also indebted to Dr. Georg Fischer from CAS for his general help with Afrotropical ants and the good time in the field with FHG. He also collected and processed a good number of specimens from East Africa used in this study. We want to thank Dr. Gary D. Alpert from MCZ for providing very important material from Mozambique. In addition, we want to express our gratitude to Marek Borowiec, Barry Bolton, and one anonymous reviewer for suggestions, comments, and critique that have improved the manuscript. Furthermore, we are thankful to the following curators and/or curatorial staff, who either loaned important material or welcomed FHG to their collections: Mrs Suzanne Ryder, Dr. Gavin Broad, Mrs Natalie Dale-Skey Papilloud from BMNH; Dr. Stefan Cover and Dr. Gary D. Alpert from MCZ; Dr. Giulio Cuccodoro from MHNG; Mrs Isabelle Zürcher-Pfander from NHMB; Dr. Brian Brown and Mrs Giar-Ann Kung from LACM; and Dr. Eliane De Coninck from RMCA. This study was partially supported by the National Science Foundation under Grant No. DEB-0842395 granted to BLF and two Ernst Mayr Travel Grants from the MCZ granted to FHG to visit the collections in BMNH and MCZ.