





Citation: Yu D, Zhang F, Deharveng L (2014) A peculiar cave species of Tomocerus (Collembola, Tomoceridae, Tomocerinae) from Vietnam, with a discussion of the postantennal organ and prelabral chaetae in Tomocerinae. ZooKeys 408: 61–70. doi: 10.3897/zookeys.408.7030
The first Tomocerus species with a postantennal organ (PAO) in the adult stage is described from Vietnam. Tomocerus postantennalis sp. n. differs from the other PAO-possessing tomocerid, Tomolonus reductus Mills, 1948, mainly in the morphology of PAO, the number of ocelli, the number of chaetae in trochantero-femoral organ and several features of the furca. The new species is placed in Tomocerus because of the presence of a toothlet on the outer basal mucronal tooth and the absence of the diagnostic character states of Plutomurus Yosii, 1956 and Aphaenomurus Yosii, 1956. Besides the presence of PAO, the new species is peculiar in having six prelabral chaetae, instead of four as in other Tomocerus species. The new species is similar to Tomocerus folsomi Denis, 1929 and Tomocerus ocreatus Denis, 1948 in the type of dental spines but different from them in the body colour, the relative length of antennae to body, the number of unguis inner teeth and the number of mucronal intermediate teeth.
New species, Southeast Asia, taxonomy, Tomocerus folsomi, Tomocerus ocreatus, Tomolonus
In Collembola, PAO is a paired organ located dorsally on the head, behind the antennae. It is probably a sensory organ of smell, humidity or temperature (
PAO in Tomocerinae was firstly recorded by
Until now, the only known species of Tomocerinae with PAO in the mature stage was Tomolonus reductus, from North America. The purpose of this paper is to describe a second species of this subfamily with well developed PAO, based on material from Vietnam.
All type specimens were collected in caves with aspirators. After being photographed with a Jenoptik ProgRes C10+ camera mounted on a Leica MZ 16 stereomicroscope, specimens were cleared in lactic acid and mounted in Marc André II solution. The head, furca and legs were cut off from the trunk and mounted separately for detailed observation. The slide-mounted specimens were studied using a Leica DMLB microscope.
The pattern of dorsal cephalic chaetotaxy used here is modified from that
http://zoobank.org/D2015E75-AF28-4A47-9FC2-7AC6D0568FD4
http://species-id.net/wiki/Tomocerus_postantennalis
Figs 1–18Vietnam, Tuyen Quang Province: Na Hang, Khuoi Sung, Hang Khuoi Sung, in cave, 22.5013°N, 105.3649°E, 24 Dec. 2003, Louis Deharveng and Anne Bedos leg.
Holotype female and three paratype females on slides, labelled with collectors’ sample number Vn0312-56. Deposited in MNHN (holotype and two paratypes) and NJAU (one paratype).
Body length 3.4–4.3 mm. Body with diffuse dark pigment all over; Ant. II, base of Ant. III and ventral side of Ant. I darker than other parts of antenna; eye patches black and small; clypeus, antero-dorsal region and posterior margin of head darker than other parts of head; anterior half of trunk darker than posterior half; head and trunk with bilaterally symmetrical white pattern formed by numerous unpigmented patches (Fig. 1).

Tomocerus postantennalis sp. n. Appearance in alcohol. Scale bar: 1000 μm.
Antenna longer than body, Ant. IV lost, Ant. I:II:III ≈ 1.0:1.8:21.0, Ant. I and II dorsally scaled, Ant. III unscaled. Antennal chaetae poorly preserved and not studied. PAO oval, with thickened inner margin, its long axis as long as the diameter of anterior ocelli (Fig. 2). Ocelli 6+6, anterior two larger than others (Figs 2, 3). Prelabral chaetae 3+3; labral chaetotaxy 5, 5, 4, distal four chaetae stronger, anterior margin of labrum with four papillae (Fig. 4). Mandibular head asymmetrical, left mandible with four teeth, right mandible with five teeth, molar plate of left mandible distally with a cone-like tooth (Fig. 5). Basal teeth of maxillary lamella 5 prolonged, distinct beard absent (Fig. 6). Mentum (baso-lateral area of labium) with 5 smooth chaetae, other parts of labium not clearly seen. Cephalic dorsal chaetotaxy: anterior area 2, 2; interocular area 2, 0, macrochaetae absent along transverse medial line; postocular area 2+2; posterior area 1+1; posterior margin with about 30+30 short chaetae (Fig. 3). Head scaled both dorsally and ventrally.
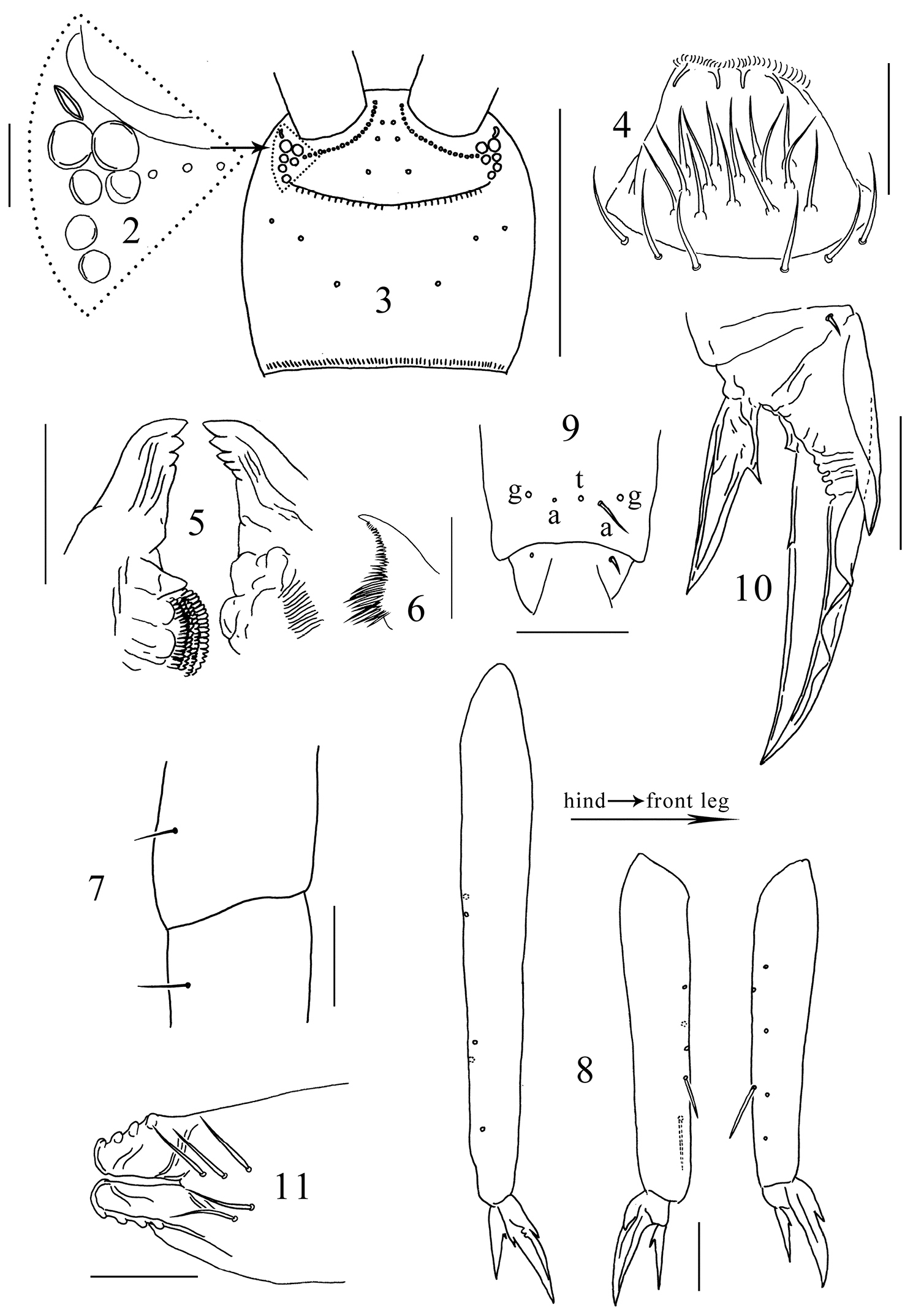
Tomocerus postantennalis sp. n. 2 PAO and ocelli 3 cephalic dorsal chaetotaxy 4 labrum 5 mandible 6 maxillary lamella five 7 trochanteral-femoral organ 8 tibiotarsus 9 anterior view of distal tibiotarsal chaetae (t: tenent hair, a: accessory chaetae, g: guard chaetae) 10 claw 11 tenaculum. Scale bars: 2, 7, 9, 10, 11 = 50 μm; 3 = 500 μm; 4, 5, 8 = 100 μm; 6= 20 μm.
Trochanteral-femoral organ with 1, 1 chaetae (Fig. 7); fore, middle and hind tibiotarsi ventrally with 6–7, 5, 5 spine-like chaetae (Fig. 8). Distal whorl of tibiotarsus with 11 chaetae, tenent hair thin and probably pointed (judging from its small socket), two small accessory chaetae beside tenent hair larger than pretarsal chaetae, sockets of two outer guard chaetae larger than tenent hair (Fig. 9). Unguis slender, with baso-internal ridging, two lateral teeth pointed, of moderate size; inner edge of unguis with one basal and one distal tooth, the distal tooth at about one third of the length of unguis from base. Unguiculus length 0.50–0.67 that of unguis, with one inner tooth larger than ungual teeth (Fig. 10). Scales present on all segments except pretarsus of all legs.
Ventral tube scaled both anteriorly and posteriorly, lateral flap unscaled. Each side of anterior face with ca. 50 chaetae, posterior face with ca. 90 chaetae, each lateral flap with ca. 60 chaetae; all chaetae smooth.
Tenaculum unscaled, with 4+4 teeth, anterior face with 5 small smooth chaetae (Fig. 11). Ratio manubrium:dens:mucro 3.3–4.0:4.8–5.4:1.0. Manubrium laterally with large scales and 11 chaetae, the proximal one chaeta small and smooth, the distal 10 chaetae enlarged and serrated; dorsal scales absent; each dorsal chaetal stripe with 250–300 smooth chaetae of different sizes, including 2+2 pointed prominent chaetae larger and straighter than other chaetae (arrowed in Fig. 12); 22–27 pseudopores on each side (Fig. 12); external corner chaetae as large as mesochaetae in the chaetal stripe (Fig. 13). Ventral side of manubrium covered only with scales. Dental spines formula 4–5/5–7, I, 1–2, I. Distal most spine strongest, about 0.1 times length of dens, sizes of the proximal spines increasing gradually from basal to distal. All spines compound, with numerous denticles of moderate size at basal half and smaller at distal half (Fig. 14). Dens internally divided into three subsegments, basally without outer strong chaetae or inner pointed scales, dorsally with ordinary smooth chaetae and a longitudinal central stripe of feathered chaetae (Fig. 15) from base of middle subsegment to apex of distal subsegment between ordinary chaetae, ventrally covered with small oval scales. Mucro elongate and multi-setaceous; both basal teeth with proximal lamellae, outer basal tooth with a toothlet (Fig. 16); apical and subapical teeth subequal; two dorsal lamellae beginning from subapical tooth, outer lamella ending in inner basal tooth, inner lamella ending freely beside inner basal tooth. Outer dorsal lamella with 5–7 subequal intermediate teeth (Fig. 17).
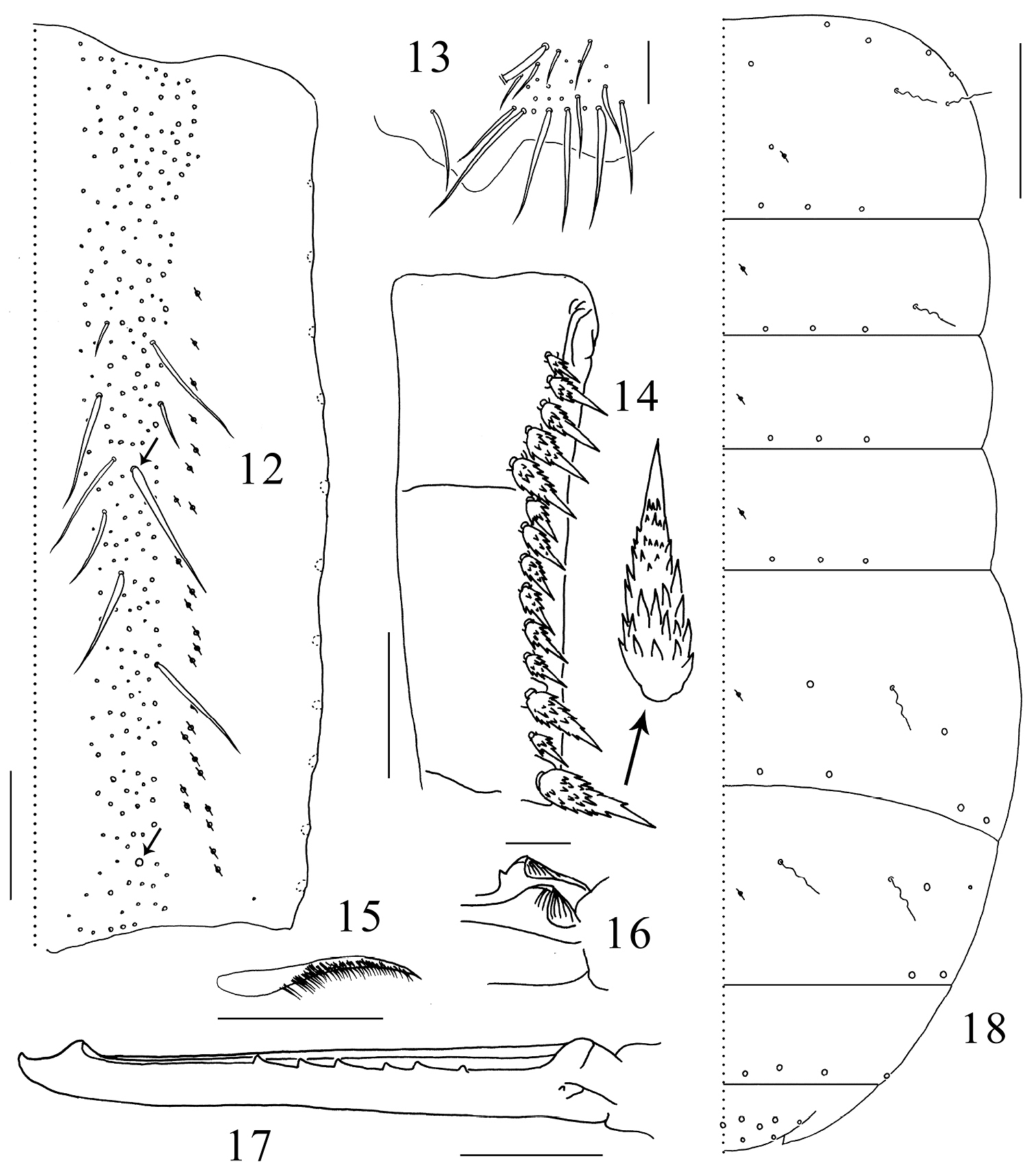
Tomocerus postantennalis sp. n. 12 dorsal face of manubrium (right side; prominent chaetae arrowed) 13 disto-dorsal chaetae on manubrium (left side) 14 dental spines (left side) 15 feathered chaeta on dens 16 basal teeth of left mucro 17 right mucro 18 body chaetotaxy. Scale bars: 12, 14 = 100μm; 13, 17 = 50 μm; 15, 16 = 21 μm; 18 = 400 μm. Large circles: macrochaetae; small circles: mesochaetae; wavy lines: bothriotricha; circles with a slash: pseudopores.
Macrochaetae distributed densely along anterior margin of Th. II (not drawn) and sparingly in posterior rows on terga. Th. II-Abd. V with 2, 1/0, 0, 1, 2, 0 bothriotricha and 3, 3/3, 3, 4, 2, 4 (3 central+1 lateral) posterior macrochaetae on each side; dorsal flap of Abd. VI with 13 macrochaetae (6+6 and 1 on middle line). Medial area of Th. II with two macrochaetae, the posterior one close to pseudopore; Abd. IV antero-laterally with one macrochaeta and one mesochaeta (Fig. 18). Most mesochaetae present at lateral margin of terga, especially from Abd. II to Abd. IV. Microchaetae uniformly distributed.
Named with reference to the presence of the postantennal organ.
Tomocerus postantennalis sp. n. is distinct from any other Tomocerus spp. in the presence of PAO and six prelabral chaetae. Besides, it is characterized by the unequal size of the ocelli, the lower number of cephalic macrochaetae and the reduction in the number of ungual inner teeth. Its compound dental spines are very similar to those of the Vietnamese species Tomocerus ocreatus Denis, 1948 and of the Chinese species Tomocerus folsomi Denis, 1929, which probably indicates a close phylogenetic relationship. The discrimination of these species is shown in Table 1.
Discrimination of Tomocerus postantennalis sp. n., Tomocerus ocreatus and Tomocerus folsomi on the basis of the original descriptions and notes (
| Species | Body colour | Length of antennae | Unguis teeth | Shape of dental spines | Dental spines formula | Number of mucronal intermediate teeth |
|---|---|---|---|---|---|---|
| Tomocerus folsomi | yellowish with dark pigment along lateral margin of anterior terga | shorter than body | 4–5 | compound with very fine denticles | 5–8/3–6, I, 1–2, I | 6–7 |
| Tomocerus ocreatus | pale | as long as body | 5 | compound with denticles of moderate size | 3/3–4, II | 8–9 |
| Tomocerus postantennalis sp. n. | dark grey | longer than body | 2 | compound with denticles of moderate size | 4–5/5–7, I, 1–2, I | 5–7 |
Most tomocerids have four prelabral chaetae, which is also common number in other groups of Entomobryomorpha. The taxonomic significance of the prelabral chaetae in Tomocerinae was firstly discovered by
So far, only two species of Tomocerinae have the PAO developed in adults: Tomolonus reductus (Mills) and Tomocerus postantennalis sp. n. The former has the most reduced eyes of non-cave species (3+3) and lives in soil, while the latter has the maximum number of ocelli for the subfamily (6+6) and lives in a cave, where its eyes are useless because of darkness. PAO development in adult may therefore be assumed to compensate for deficient vision performance in both species. Under this assumption, we would expect PAO among other cave species of Tomocerinae, a presence difficult to detect given the inconspicuousness of the organ when present.
The PAO of Tomocerus postantennalis sp. n. is similar in shape to those of juvenile Pogonognathellus longicornis (
Despite the presence of PAO, Tomolonus is more closely related to Plutomurus, since both have two posterior macrochaetae on the thoracic segments (
The genus Tomocerus Nicolet, 1841 is so far poorly defined by a single character: the presence of a toothlet on the outer basal mucronal tooth, thus all species conforming to this criterion and lacking the diagnostic character of other genera have been assigned to this genus, resulting in a wider range of intrageneric diversity than in other genera. For instance, all types of dental spines from simple to strongly furcated can be found in Tomocerus, whereas in Pogonognathellus, Plutomurus and Monodontocerus the shape of dental spines are constant within genus. On the other hand, the single generic character is not exclusive for Tomocerus since Aphaenomurus interpositus denticulatus Yosii, 1956 and Plutomurus vigintiferispina Lee, 1974 also have a toothlet on the mucronal outer basal tooth. The situation of Tomocerus will remain problematic until a comprehensive and detail investigate is carried out.
We thank Dr Mark Judson (MNHN, Paris) for reviewing the English style. The China Scholarship Council provided a grant to the first author for studying at MNHN, Paris.