





Citation: Kavanaugh DH, Hieke F, Liang H, Dong D (2014) Inventory of the carabid beetle fauna of the Gaoligong Mountains, western Yunnan Province, China: species of the tribe Zabrini (Coleoptera, Carabidae). ZooKeys 407: 55–119. doi: 10.3897/zookeys.407.7353
A ten-year multidisciplinary, multi-national and multi-institutional biodiversity inventory project in the Gaoligong Shan region of western Yunnan Province, China generated more than 35, 000 specimens of the beetle (Coleoptera) family Carabidae. In this report, first of a planned series, we focus on diversity in tribe Zabrini. Our study of just over 1300 specimens of zabrine carabids from the project, all in genus Amara Bonelli, found a total of 13 species, all previously described, to occur in the study area, with none of them strictly endemic. We present a key for identification of adults of these species, as well as nomenclatural data, diagnoses, illustrations of dorsal habitus and male genitalia, and information about geographical, altitudinal and habitat distributions within the study area and overall geographical distribution for each species. Distributions of the species within the study area are compared, and broader geographical range patterns are characterized. We also discuss a possible role of the Gaoligong Shan region as one source area for the present-day fauna of the Himalaya and southern edge of the Qinghai-Xizang (Tibetan) Plateau.
Coleoptera, Carabidae, Zabrini, Amara, China, Yunnan, Gaoligong Shan, Himalaya, Qinghai-Xizang (Tibetan) Plateau, distribution, biodiversity hotspot
The Gaoligong Shan (Gaoligong Mountains) of extreme western Yunnan province, China, form the westernmost range of the Hengduan Mountains system of southeastern Xizang Autonomous Region (Tibet), northern and western Yunnan, and western Sichuan (Fig. 1). They extend north to south for more than 600 km, and, in the central part of the range, their crest forms the border between China and Myanmar. They also separate and form parts of the watersheds of two of Southeast Asia’s major rivers, the Irrawaddy and the Salween (known in China as the Nujiang). Elevations within the region range from a low of about 650 m in the south to more than 5000 m in the north.
Map of Asia with study region outlined; scale line = 500 km. Modified from Wikimedia Commons, World Atlas of the World, at URL: http://upload.wikimedia.org/wikipedia/commons/8/8f/Whole_world_-_land_and_oceans_12000.jpg
In late 1997, the California Academy of Sciences was invited to participate in a joint project with the Kunming Institutes of Botany and Zoology of the Chinese Academy of Sciences to conduct a biodiversity inventory of the Gaoligong Mountains. Scientists from several additional institutions, including the Institute of Zoology, Beijing, and Royal Botanical Garden (Edinburgh) joined in the collaboration. Principal target groups for the inventory included bryophytes and vascular plants, all vertebrate groups, and arachnids, myriapods, and insects, especially the Neuropteroidea, Mecoptera, and Coleoptera (the Carabidae in particular). Multidisciplinary and multi-institutional teams carried out biotic sampling through more than 25 separate expeditions during the period 1998 to 2007. More than 100 reports on the project have been published to date, including partial results for bryophytes (e.g.,
Prior to the start of the project the carabid beetle fauna of the region was very poorly known. The faunal for the entire Hengduan region included only about 50 species (
This report, on the tribe Zabrini, represents the first of an intended series of treatments on the carabid beetle fauna of the Gaoligong Shan region, each dealing with one or more tribes or hyper-diverse genera represented in the fauna. These will appear as taxonomic work on each group is completed and not in any particular taxonomic or phylogenetic order.
Zabrini is a moderately diverse taxon, including nearly 700 described species (
Amara is most diverse in temperate parts of the Holarctic Region. Along with members of the tribe Harpalini, zabrines are unusual among Carabidae in feeding mainly on seeds (
As is the case with most other terrestrial anthropod groups, the Amara fauna of the study area has not been well documented previously. Most of our current knowledge of the Southeast Asian regional fauna is from the works of
The natural physiographic limits of the study area for the project are as shown in Fig. 2 and include areas in eastern Myanmar and southern Xizhang (Tibet); but we had permission to survey only those parts in Yunnan Province. Specialists for all taxonomic groups concentrated their efforts on seven core areas within the project region (Fig. 3), selected to facilitate comparisons of possible north to south and east to west spatial differences within the regional biota, as well as recognition of areas of local endemism. Other areas were sampled as time and opportunity permitted. The entomological team made a total of 13 expeditions to the Gaoligong region. Our sampling sites within the region are shown in Fig. 4. Habitats included in the study area range from subtropical lowland rainforest to the margin of glaciers and snowfields. In all, more than 35, 000 carabid specimens were collected during the project by using a variety of collecting methods, including hand collecting both day and night, beating vegetation, sifting litter with subsequent extraction by hand or by mini-Winkler units, and Malaise flight traps and pitfall traps. All specimens were sorted to morphospecies (i.e., presumptive species units based on features of external structure and male and female genitalic traits) and detailed systematic studies of taxonomic groups are ongoing.
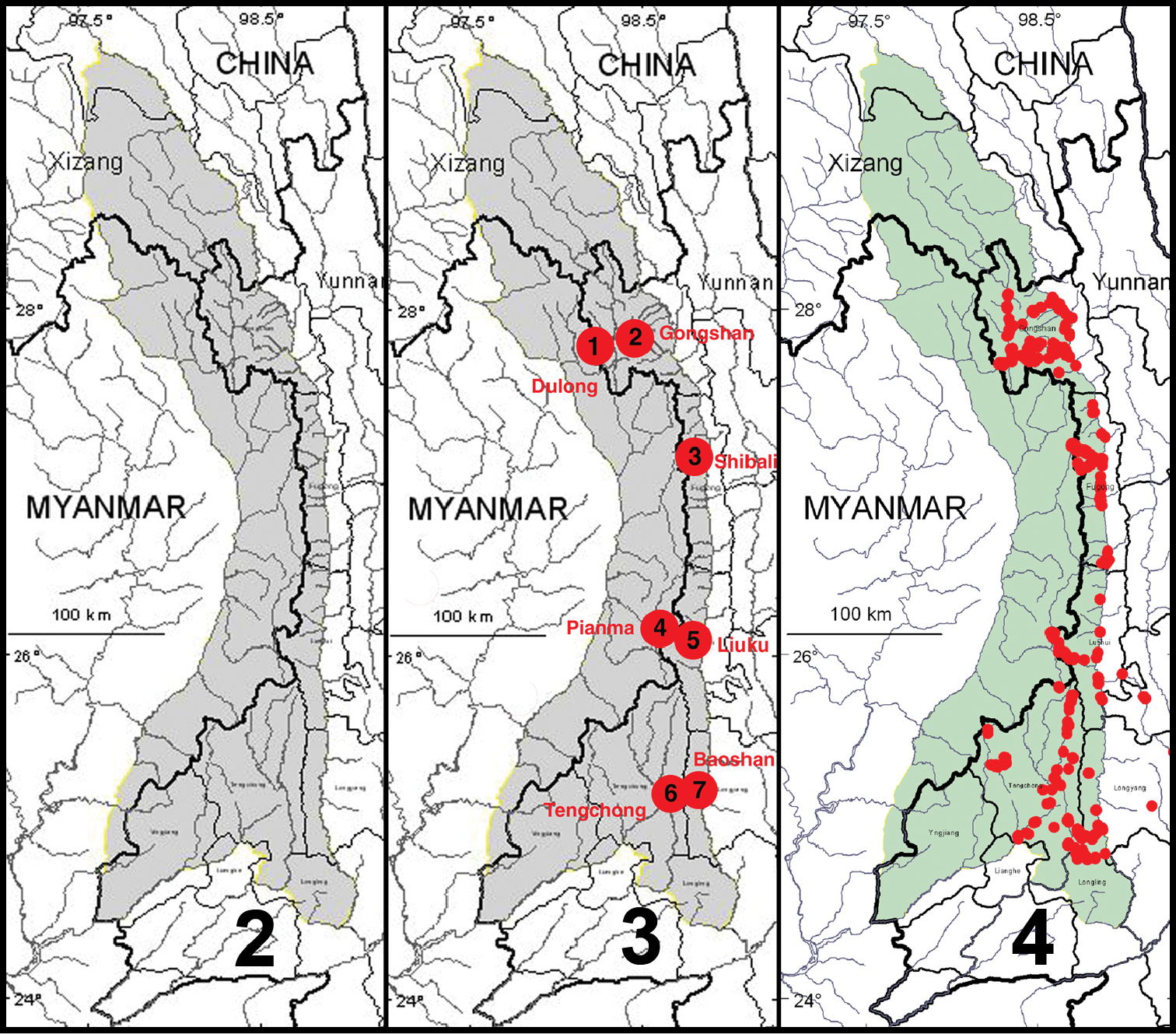
2 Map showing natural extent of study area, colored in gray (however, sampling was permitted only in those portions in Yunnan Province 3 Map showing location of core sampling areas 4 Map showing locations of all entomological sampling sites.
A total of 1, 327 specimens representing zabrine species were collected during the project. All of these specimens have been divided among and are deposited in collections of our home institutions. Codens used throughout this report for collections in which specimens, including primary types, are deposited are as follows:
BMNH British Museum (Natural History), London, United Kingdom
CAS California Academy of Sciences, San Francisco, U.S.A.
CMEY Collection of P. Meyer, Darmstadt, Germany
CWRA Collection of D. Wrase, Berlin, Germany
DEI Deutsches Entomologisches Institut, Eberswalde, Germany
FRSDD Forest Research Institute, Dehra Dun, India
IOZ National Zoological Museum of China, Institute of Zoology, Beijing, China
IRSNB Institut Royal des Sciences Naturelles de Belgique, Brussels, Belgium
MCSNG Museo Civico di Storia Naturale, Genoa, Italy
MNHN Muséum National d’Histoire Naturelle, Paris, France
NHRS Naturhistoriska Riksmuseet, Stockholm, Sweden
NMPC National Museum (Natural History), Prague, Czech Republic
NZSI National Zoological Collection, Zoological Survey of India, Calcutta, India
ZIN Zoological Institute Academy of Sciences, St.Petersburg, Russia
ZFMK Zoologisches Forschungsmuseum “Alexander Koenig”, Bonn, Germany
ZMHB Museum für Naturkunde an der Humboldt-Universität, Berlin, Germany
ZMMU Zoological Museum, Moscow University, Moscow, Russia
The only measurements taken were of body length, measured from the anterior margin of the clypeus to the apex of the longer elytron. Digital images of dorsal habitus of a typical representative member of each morphospecies were taken using an Automontage imaging system from Synchroscopy with a JVC KY-F-75U digital camera and a Leica M420 dissecting microscope. The “CASENT” number associated with each image, as noted in figure captions, is a unique identifier that refers to the particular specimen photographed and its CAS database record. Distribution maps for each species were generated from geographical coordinate data maintained in a Biota Version 3.0 database (
Adult specimens of species represented in the Gaoligong Shan region can be distinguished using the following key.
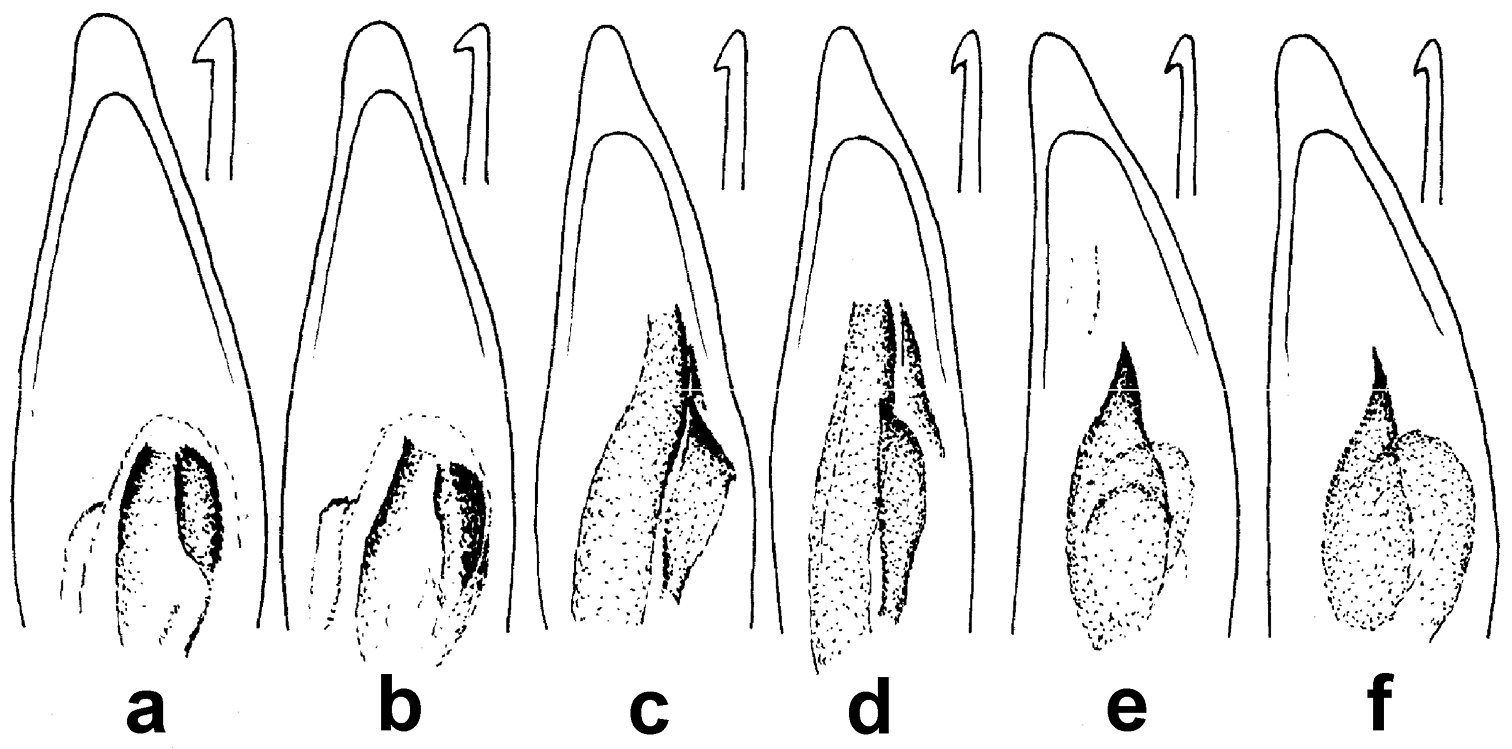
| 1 | Medial protibial spurs trifid (Fig. 5a); dorsal surface dark metallic green, non-metallic black in a very few specimens; body length 7.0–8.5 mm; elytron with parascutellar pore-puncture present | Amara (Zezea) davidi Tschitschérine, 1897 |
| – | Medial protibial spurs simple (Fig. 5b) | 2 |
| 2 | Last abdominal sternite of male with two pairs of setiferous punctures (Fig. 5d) near hind margin (second seta absent from one side in a few males for total of three setae); dorsal surface dark piceous; body length 7–8 mm; pronotum with both inner and outer basal impressions deep and broadly foveate (Fig. 14a) | Amara (Xenocelia) sikkimensis Andrewes, 1930 |
| – | Last abdominal sternite of male with one pair of setiferous punctures (Fig. 5c) near hind margin (Fig. 5d) | 3 |
| 3 | Body length more than 11 mm; tarsomere 5 of hind tarsi with five or six setal pairs ventrally (Fig. 6a); dorsal surface dark piceous; body length 11–13 mm, body form stout; pronotum (Fig. 20a) with lateral margins markedly rounded, basal area punctate, in most specimens also punctate laterally in basal half and near anterior margin | Amara (Bradytus) pingshiangi Jedlička, 1957 |
| – | Body length less; tarsomere 5 of hind tarsi with two (in a few specimens three) setal pairs ventrally (Fig. 6b) | 4 |
| 4 | Elytron with parascutellar pore-puncture present; dorsal surface with metallic luster | 5 |
| – | Elytra without parascutellar pore-puncture | 7 |
| 5 | Base of pronotum evenly convex from one side to the other, outer basal impressions absent or only very faintly suggested; body length 9–10 mm; sclerites of internal sac of median lobe of male aedeagus as in Fig. 7e–f | Amara (Amara) congrua Morawitz, 1863 |
| – | Base of pronotum slightly flattened at the sides, only the middle part evenly convex, outer basal impressions evident, either shallow and obliquely linear (Fig. 11a) or deep and broadly foveate (Fig. 12a); sclerites of internal sac of median lobe of male aedeagus with different form (Fig. 7a–d) | 6 |
| 6 | Base of pronotum coarsely punctate; outer basal impressions shallow and obliquely linear (Fig. 11a); male aedeagus with median lobe distinctly broader in apical one-third than more basally, apical lamella shorter, broadly rounded apically and with sides only slightly convergent subapically (Fig. 11c); sclerites of internal sac of median lobe of male aedeagus as in Fig. 7a–b; body length 9–10 mm | Amara (Amara) silvestrii Baliani, 1937 |
| – | Base of pronotum finely punctate; outer basal impressions deep and broadly foveate (Fig. 12a); male aedeagus with median lobe not or only slightly broader in apical one-third than more basally, apical lamella longer, narrowly rounded apically and with sides more distinctly convergent apically (Fig. 12c); sclerites of internal sac of median lobe of male aedeagus as in Fig. 7c–d; body length 8.5–10.0 mm | Amara (Amara) shaanxiensis Hieke, 2002 |
| 7 | Dorsal surface light-brown to brownish black, without metallic reflection, entire legs and antennae pale; body length 6.5–8.0 mm; male aedeagus with apical third of median lobe broader than middle third (Fig. 21c), apical hook of right paramere large and slightly subapical (Fig. 8a) | Amara (Reductocelia) lucidissima Baliani, 1932 |
| – | Dorsal surface darker, with or without distinct metallic reflection, at least femora dark (piceous to black) | 8 |
| 8 | Pronotum with lateral margins straight or faintly to distinctly sinuate just anterior to basal angles, rounded near middle, less rounded or nearly straight also in anterior one-third in most specimens, anterior angles distinctly and narrowly projected anteriorly beyond anterior margin; dorsal surface dark with distinct metallic blue-green reflection in most specimens, non-metallic black in a few specimens | 9 |
| – | Pronotum with lateral margins more or less evenly rounded from apical to basal angle, anterior angles not or only faintly and broadly projected anteriorly beyond anterior margin; dorsal surface with or without metallic reflection | 10 |
| 9 | Pronotum (Fig. 18a) with lateral explanation narrow throughout, outer basal impressions foveate and distinct from lateral groove, punctation of base not extended anteriorly along sides beyond basal one-third; elytral microsculpture effaced or nearly so in both males and females; body length 7.0–7.5 mm | Amara (Bradytus) elegantula Tschitschérine, 1899 |
| – | Pronotum (Fig. 16a) with lateral explanation distinctly broader basally, outer basal impressions indistinct from lateral groove in most specimens, punctation of base extended anteriorly along sides to pronotal mid-length; elytral microsculpture comprised of isodiametric meshes, faintly impressed or nearly effaced in males, deeply impressed and distinct in females; body length 7.5–9.0 mm | Amara (Bradytus) chalciope (Bates, 1891) |
| 10. | Elytral microscuplture comprised of distinctly transverse meshes in both males and females (more transverse and less deeply impressed in males than in females); pronotum (Fig. 19a) only slightly narrower anteriorly than basally, anterior margin almost as wide as posterior margin, pronotal base very coarsely punctate, outer basal impressions sharply delimited laterally by narrow, slightly oblique raised (but not carinate) areas | Amara (Bradytus) simplicidens Morawitz, 1863 |
| – | Elytral microscuplture comprised of distinctly isodiametric meshes in both males and females (more deeply impressed in females than in males); pronotum (Figs 13a, 15a, 17a) more distinctly narrowed anteriorly than basally, anterior margin clearly narrower than posterior margin, pronotal base moderately coarsely punctate, outer basal impressions either not sharply delimited laterally by raised areas or, if so, then the raised area broader | 11 |
| 11 | Pronotum (Fig. 15a) with posterior angles distinctly rounded (narrowly so in some individuals), slightly to moderately obtuse; elytra with slight sub-basal depressions centered on striae 6 (visible on right elytron in Fig. 15a), also on striae 4 and/or 5 in some individuals; dorsal surface black, with very faint metallic blue or green metallic reflection in most individuals, more vivid (as in Fig. 15a) in or lacking from a few specimens; body length 8.5–9.0 mm | Amara (Harpaloamara) latithorax Baliani, 1934 |
| – | Pronotum (Figs 13a, 17a) with posterior angles either obtusely angulate and slightly denticulate or narrowly rounded (if the latter, then body length 7.5 mm or less); elytra without evident sub-basal depressions; dorsal surface with or without metallic reflection | 12 |
| 12 | Dorsal surface with distinct metallic copper or bronze reflection, non-metallic black in very few specimens; pronotum (Fig. 13a) with posterior angles narrowly rounded, basal impressions deeply foveate but small in diameter, outer basal impressions not distinctly delimited laterally by a broad convexity; elytral striae distinct throughout but only shallowly impressed in most individuals; metatibiae of males without a brush-like patch of setae medially in the apical half; body length 6.5–7.5 mm | Amara (Pseudoamara) birmana Baliani, 1934 |
| – | Dorsal surface black or piceous, without or with only very faint metallic green reflection; pronotum with posterior angles obtusely angulate and slightly denticulate, basal impressions broadly and deeply foveate, outer basal impression distinctly delimited laterally by a broad convexity; elytral striae distinct throughout and deeply impressed; metatibiae of males with a brush-like patch of setae medially in the apical half; body length 7.7–8.7 mm | Amara (Bradytus) dissimilis Tschitschérine, 1894 |
http://species-id.net/wiki/Amara_davidi
Figs 5a, c, e, 6b, 9, 22a, 27–30Adults of this species (Fig. 9) can be distinguished from those of all other species in the region by their trifid medial protibial spurs (Fig. 5a). Amara davidi is the only member of subgenus Zezea, members of which share this feature, known from the region.

Tibial and abdominal apices. a–b right tibial apices, lateroventral aspect; a. Amara davidi Tschitshérine b Amara sikkimensis Andrewes, typical of all other Amara species of the area c–e abdominal apices, ventral aspect, with white arrows indicating insertion points for setae c Amara sikkimensis, typical of all Amara females d Amara sikkimensis Andrewes male e Amara latithorax Baliani, typical of males of all other Amara species of the area.

Tarsomere 5 of left hind tarsus, medial aspect. a Amara pingshiangi Jedlička b Amara sikkimensis Andrewes, typical of all other Amara species of the area.
Specimens of this species were collected from under stones in open roadside and waste areas (Fig. 22a) with scattered grasses and shrubs at elevations ranging from 2020 to 2440 m, syntopic (together in the same habitat) with adults of Amara birmana, Amara shaanxiensis, Amara sikkimensis and Amara silvestrii at one or more sites.
Fig. 9d. We examined a total of 3 specimens (all females) from the following localities: Gongshan County: Heiwadi (16.8 km W of Cikai on Dulong Valley Road, 27.79584°, 98.58443°, 2020 m, 20 April 2002, H.B. Liang, W.D. Ba, G.D. Yang & X.Q. Li collectors [2 females; CAS, IOZ]). Longyang County: Bawan-Tengchong Road Km 41 (near yakou, 24.93972°, 98.75333°, 2440 m, 15 October 2003, H.B. Liang & J.J. Yang (1 female; CAS).
Members of this species were collected in both the northern and southern parts of the study area (Core Areas 2, 6 and 7) but not in the central part. This gap in distribution is most likely an artifact of inadequate sampling and not a real disjunction.
Fig. 27. This species has been recorded from Beijing, Gansu, Hubei, Qinghai, Shaanxi, Sichuan and Yunnan Provinces in China. Its occurrence in the study area represents the southwestern limit of its known geographical range.
http://species-id.net/wiki/Amara_congrua
Figs 5b–c, e, 6b, 7e–f, 10, 22b, 27–30Adults of this species (Fig. 10) can be distinguished from those of all other species in the region by the following combination of character states: body length 9–10 mm; base of the pronotum evenly convex from one side to the other, outer basal impressions absent or only very faintly suggested; elytra with parascutellar pore puncture present; medial protibial spurs simple; tarsomere 5 of hind tarsi with two or (in a few specimens) three pairs of setae ventrally (Fig. 6b); last abdominal sternite of male with one pair (Fig. 5e) and female with two pairs (Fig. 5c) of setiferous punctures near hind margin; sclerites of internal sac of median lobe of male aedeagus with form as in Fig. 7e–f.
Apical and subapical region of median lobe (dorsal aspect) and apex of right paramere (medial aspect) of aedeagus of males. a–b Amara silvestrii Baliani c–d Amara shaanxiensis Hieke e–f Amara congrua Morawitz.
Specimens of this species were collected in daytime from under stones and other cover in open roadside areas with scattered grasses and shrubs (Fig. 22b), at the edges of agricultural fields, including wet and dry rice paddies, and on open banks of streams, and in these same habitats at night, when beetles were found active on bare substrate. Specimens were found at elevations ranging from 892 to 2000 m. The only other Amara species members of which were found syntopic with those of Amara congrua was Amara latithorax, with adults of both species found together at only one site at an elevation of 1223 m.
Fig. 10d. We examined a total of 33 specimens (17 males and 16 females) from the following localities: Fugong County: Lumadeng Township (Chihengdi village, 26.98749°, 98.86310°, 1230 m, 29 April 2004, D.H. Kavanaugh & H.B. Liang collectors [3 males and 2 females; CAS, IOZ]); Shangpa Township (road on west side of Nu Jiang S of Shangpa, 26.88952°, 98.86539°, 1223 m, 22 April 2004, D.H. Kavanaugh & C.E. Griswold collectors [1 male and 1 female; CAS]). Gongshan County: Cikai Township (27.27064°, 98.66557°, 1500 m, 28 June 2000, D.H. Kavanaugh & H.B. Liang collectors [1 male; CAS]); Heiwadi village (27.78594°, 98.60117°, 1880-2000 m, 28 June 2000, D.H. Kavanaugh & H.B. Liang collectors [1 female; IOZ]). Lushui County: Liuku Township (Laimo village, 25.82767°, 98.85120°, 892 m, 15 May 2004, H.B. Liang & X.Y. Li collectors [1 male and 1 female; CAS, IOZ]). Tengchong County: Hehua Township (3.8 km S of Hehua along Daying Jiang at Nangyan village, 24.93873°, 98.38444°, 1140 m, 2 June 2006, D.H. Kavanaugh, R.L. Brett & D.Z. Dong collectors [1 female; CAS]), (5.4 km S of Hehua along Daying Jiang at Dengma village, 24.92346°, 98.38612°, 1105 m, 2 June 2006, D.H. Kavanaugh, R.L. Brett & D.Z. Dong collectors [1 female; IOZ]; Jietou Township (stream 0.7 km N of Jietou, 25.43128°, 98.64773°, 1564 m, 22 May 2006, D.H. Kavanaugh, R.L. Brett & H.B. Liang collectors [3 males and 2 females; CAS, IOZ]); Qingshui Township (Liangyang village in Rehai area, 24.94919°, 98.44921°, 1450 m, 1 June 2006, D.H. Kavanaugh, R.L. Brett, H.D. Liang, D.Z. Dong & P. Hu collectors [1 female; IOZ]); Qushi Township (Longchuan Jiang at Qinqiao in Qinqiao village, 25.27250°, 98.60083°, 1464 m, 6 June 2006, D.H. Kavanaugh, R.L. Brett & D.Z. Dong collectors [7 males and 5 females; CAS, IOZ, ZMHB]); Shangying Township (Bawan-Tengchong Road KM 63, 25.02917°, 98.66917°, 1360 m, 19 October 2003, H.B. Liang & X.C. Shi collectors [1 female; IOZ]); Wuhe Township (west bank of Longchuan Jiang at Tongjiazhuang village, 24.89284°, 98.67439°, 1210 m, H.B. Liang collector 24 May 2005 [1 male; IOZ]).
Members of this species were collected from the northern to the southern parts of the study area (Core Areas 2, 3, 5 and 6), but they were found only on the eastern side of the mountain range in northern and central areas (Core Areas 2, 3 and 5) and only on the western versant in the southern part (Core Area 6). This distribution pattern is most likely an artifact of inadequate sampling on the western slope of the mountain range, much of which is in Myanmar.
Fig. 27. This species has been recorded from China (Beijing, Fujian, Gansu, Guizhou, Hebei, Heilongjiang, Hongkong, Hubei, Jiangsu, Jiangxi, Jilin, Liaoning, Nei Mongol, Shaanxi, Shanghai, Sichuan, Yunnan and Zhejian Provinces), Japan (southern half), Laos, North and South Korea, Myanmar (extreme north), Russia (Primorskij Krai), Taiwan and Vietnam (extreme north).
http://species-id.net/wiki/Amara_silvestrii
Figs 5b–c, 5e, 6b, 7a–b, 11, 23a, 27–30Adults of this species (Fig. 11) can be distinguished from those of all other species in the region by the following combination of character states: body length 9–10 mm; base of pronotum slightly flattened at the sides, only the middle part evenly convex, coarsely punctuate, outer basal impressions shallow and obliquely linear (Fig. 11a); elytra with parascutellar pore puncture present; medial protibial spurs simple; tarsomere 5 of hind tarsi with two or (in a few specimens) three pairs of setae ventrally (Fig. 6b); last abdominal sternite of male with one pair (Fig. 5e) and female with two pairs (Fig. 5c) of setiferous punctures near hind margin; male aedeagus with median lobe distinctly broader in apical one-third than more basally, apical lamella shorter, broadly rounded apically and with sides only slightly convergent subapically (Fig. 11c), sclerites of internal sac with form as in Fig. 7a–b.
Specimens of this species were collected in daytime from under stones and other cover in open roadside areas (Fig. 23a) and meadows with scattered grasses and shrubs, at the edges of agricultural fields, including wet and dry rice paddies, under clods of soil in recently tilled fields and on open banks of streams, and in these same habitats at night, when beetles were found active on bare substrate. They were also collected under debris and in leaf litter in deciduous forests and also in crevices between stones in a talus slope. Members of this species were found at elevations ranging from 1515 to 3000 m, syntopic with adults of Amara birmana, Amara chalciope, Amara davidi, Amara lucidissima, Amara shaanxiensis, and Amara sikkimensis at one or more sites.
Fig. 11d. We examined a total of 34 specimens (18 males and 16 females) from the following localities: Fugong County: Lishadi Township (Shibali area, 27.16536°, 98.78003°, 2535 m, D.H. Kavanaugh, P. Paquin & D.Z. Dong collectors [1 female; CAS]), (0.5 km W of Shibali, 27.16665°, 98.77936°, 2537 m, 18 August 2005, P. Paquin collector [1 male and 1 female; CAS]), (below Shibali on Yaping Road, 27.16520°, 98.77980°, 2530 m, 24 April 2004m, H.B. Liang & X.Y. Li collectors [1 male; IOZ]), (7.5 km below Shibali on Yaping Road, 27.14627°, 98.81559°, 2030 m, 3 May 2004, H.B. Liang & M. Xi collectors [1 female; IOZ]); Lumadeng Township (0.5 km W of Lao Shibali on Lao Shibali Road, 27.08072°, 98.76920°, 2305 m, 22 August 2008, P. Paquin collector [1 male; CAS]), (South Fork of Yamu He at Yejiadi, 27.08994°, 98.77325°, 2307 m, 10 May 2004, H.B. Liang collector [2 males and 1 female; CAS, IOZ); Maji Township (Majimi village near power station on Gaxie He, 27.39630°, 98.81701°, 1567 m, 28 April, 2004, H.B. Liang collector [1 male; IOZ]). Gongshan County: Cikai (27.74972°, 98.66444°, 1515 m, 5 October 2002, H.B. Liang & W.D. Ba [1 female; IOZ]); Dabadi (41 km W of Cikai on Dulong Valley Road, 27.79655°, 98.50562°, 3000 m, 27 September to 6 October 2002, D.H. Kavanaugh, P.E. Marek, H.B. Liang & D.Z. Dong collectors [5 males and 6 females; CAS, IOZ, ZMHB]); Heiwadi (16.8 km W of Cikai on Dulong Valley Road, 27.79584°, 98.58443°, 2020 m, 15 and 20 April 2002, H.B. Liang, W.D. Ba & C.G. Jin collectors [2 males and 1 female; IOZ, ZMHB]); Qiqi area (27.71542°, 98.56529°, 2000-2020 m, 9-14 July 2000, D.H. Kavanaugh & H.B. Liang collectors [1 female; CAS]); Cikai Township (8.3 to 13.1 km NW of Cikai on Dulong Valley Road, 27.75653°, 98.58214°, 2620-3000 m, 23 September 2002, D.H. Kavanaugh, P.E. Marek & D.Z. Dong collectors [1 female; CAS]). Lushui County: Luzhang Township (Yaojiaping He at Pianma Road, 25.97722°, 98.71091°, 2527 m, 20 May 2005, D.H. Kavanaugh, C.E. Griwold, H.B. Liang, D.Z. Dong & G. Tang collectors [1 male; CAS]); Pianma Township (6 km ESE of Pianma, 26.00808°, 98.65921°, 2210 m, 15 May 2005, H.B. Liang & D.Z. Dong collectors [2 males and 2 females; CAS, IOZ]). Tengchong County: Jietou Township (Cha He at Shaba village, 25.39256°, 98.70488°, 1840 m, 25 May 2006, D.H. Kavanaugh, R.L. Brett & D.Z. Dong collectors [1 male; CAS]); Qushi Township (Longchuan Jiang at Longkou village, 25.28167°, 98.59167°, 1500 m, D.H. Kavanaugh & C.E. Griswold collectors [1 male; CAS]).
Members of this species were collected from the northern to the southern parts of the study area (Core Areas 2, 3, 5 and 6), but they were found only on the eastern side of the mountain range in northern half of the study area (Core Areas 2 and 3), on both side in the central part (Core Areas 3 and 4) and only on the western versant in the southern part (Core Area 6). This distribution pattern is most likely an artifact of inadequate sampling on the western slope of the mountain range in the north, some of which is in Myanmar.
Fig. 27. This species has been recorded from China (Gansu, Hubei, Shaanxi, Sichuan and Yunnan Provinces), Myanmar (extreme north) and Taiwan. Its occurrence in the study area represents the southwestern limit of its known geographical range.
http://species-id.net/wiki/Amara_shaanxiensis
Figs 5b–c, 5e, 6b, 7c–d, 12, 22a, 25b, 27–30Adults of this species (Fig. 12) can be distinguished from those of all other species in the region by the following combination of character states: body length 8.5–10 mm; base of pronotum slightly flattened at the sides, only the middle part evenly convex, finely punctate; outer basal impressions deep and broadly foveate (Fig. 12a); elytra with parascutellar pore puncture present; medial protibial spurs simple; tarsomere 5 of hind tarsi with two or (in a few specimens) three pairs of setae ventrally (Fig. 6b); last abdominal sternite of male with one pair (Fig. 5e) and female with two pairs (Fig. 5c) of setiferous punctures near hind margin; male aedeagus with median lobe not or only slightly broader in apical one-third than more basally, apical lamella longer, narrowly rounded apically and with sides more distinctly convergent apically (Fig. 12c), sclerites of internal sac with form as in Fig. 7c-d.
Specimens of this species were collected in daytime from under stones and other cover in open roadside areas (Figs 22a, 25b), meadows and marshy areas with scattered grasses and shrubs, at the edges of agricultural fields, including wet and dry rice paddies and on open banks of streams, and in these same habitats at night, when beetles were found active on bare substrate. They were also collected under debris in daytime and on the surface of leaf litter at night in deciduous forests. Members of this species were found at elevations ranging from 1837 to 3000 m, syntopic with adults of Amara birmana, Amara chalciope, Amara davidi, Amara lucidissima, Amara pingxiangi, Amara sikkimensis and Amara silvestrii at one or more sites.
Fig. 12d. We examined a total of 88 specimens (36 males and 52 females) from the following localities: Gongshan County: Dabadi (41 km W of Cikai on Dulong Valley Road, 27.79655°, 98.50562°, 3000 m, D.H. Kavanaugh, P.E. Marek, H.B. Liang, D.Z. Dong & X.C. Liang collector [1 male and 4 females; CAS, IOZ]). Longling County: Xiaheishan Forest Reserve (Guchengshan at 1.2 km SSE of Km 23.5 on Route 23.5, 24.82888°, 98.76001°, 2020 m, 25-26 May 2005, D.H. Kavanaugh, C.E. Griswold, H.B.Liang and D.Z. Dong collectors [4 males and 4 female; CAS]). Longyang County: Bawan-Tengchong Road (Km 29-35, 24.92916°, 98.75861°, 2000-2350 m, 12 October 2003, D.Z. Dong collector [1 female; IOZ]), (Km 36-37, 24.93417°, 98.77944°, 2150 m, H.B. Liang & X.C. Shi collectors [2 males and 1 female; IOZ, ZMHB]), (Km 40–41, 24.92694°, 98.75278°, 2404 m, 12 October 2003, H.B. Liang & X.C. Shi collectors [1 female; IOZ]), (Km 41 near yakou, 24.93972°, 98.75333°, 2440 m, 15 October 2003, H.B. Liang & J.J. Yang collectors [1; female; IOZ]), (Km 42 at Sanchawa, 24.94750°, 98.75556°, 2300 m, 13 October 2003, H.B. Liang & X.C. Shi collectors [1 male; CAS]; Luoshuidong area (at Sancha He, 24.94833°, 98.75667°, 2300 m, 30 May 2005, D.H. Kavanaugh & H.M. Yan collectors [1 female; CAS]; 3 June 2005, D.H. Kavanaugh, C.E. Griswold, H.B. Liang, D.Z. Dong & H.M Yan collectors [1 male; CAS]); Nankang Forestry Station (at Km 19.8 on Route S317, 24.82284°, 98.78207°, 2060 m, 23 May 2005, D.H. Kavanaugh, C.E. Griswold, H.B. Liang, D.Z. Dong, H.M. Yang & G. Tang collectors [3 males and 7 females; CAS, IOZ, ZMHB]; Nakang Yakou (24.82587°, 98.76832°, 2148 m, 22 May 2005, H.B. Liang collector [(just N of yakou, 24.83178°, 98.76472°, 2180 m, 22 May 2005, D.H. Kavanaugh, C.E. Griswold & D.Z. Dong collectors [2 males and 2 females; CAS, IOZ]; 25 May 2005, D.H. Kavanaugh & C.E. Griswold [1 female; CAS]). Lushui County: Luzhang Township (Lusai He, 25.96378°, 98.77032°, 1873 m, 20 May 2005, H.B. Liang & D.Z. Dong collectors [5 males; CAS, IOZ]). Tengchong County: Houqiao Township (5.9 airkm NE of Houqiao below Guyong Forestry Station, 25.36562°, 98.31610°, 2030 m, 27 May 2006, D.H. Kavanaugh, R.L. Brett, H.B. Long, D.Z. Dong & Z.C. Liu collectors [1 male and 3 females; CAS, IOZ]); Jietou Township (2.0 km N of Dahetou Ligganjiao on Longtang He, 25.75743°, 98.69457°, 2080 m, 16 May 2006, D.Z. Dong collector [1 male; CAS]), (1.4 km S of Dahetou Ligganjiao along Longchuan Jiang, 25.72717°, 98.69322°, 1960 m, 16 May 2006, H.B. Liang collector [2 males and 3 females; CAS, IOZ]), (Longtang He at Dahetou Lingganjiao, 25.73947°, 98.69630°, 2010 m, 14-20 May 2006, D.H. Kavanaugh, R.L. Brett, H.B. Liang & P. Hu collectors [8 males and 15 female; CAS, IOZ]); Shangying Township (Bawan-Tenchong Road Km 42-46, 24.95361°, 98.74222°, 2290 m, 14 and 17 October 2003, H.B. Liang & X.C. Shi collectors [4 males and 2 females; CAS, IOZ]).
Members of this species were collected in both the northern and southern parts of the study area (Core Areas 2, 5, 6 and 7), but not in the central part (Core Area 3). This gap in distribution is most likely an artifact of inadequate sampling and not a real disjunction.
Fig. 27. This species has been recorded from Shaanxi and Yunnan Provinces in China, and its occurrence in the study area represents the southwestern limit of its known geographical range.
http://species-id.net/wiki/Amara_birmana
Figs 5b–c, 5e, 6b, 13, 22a, 23a, 25a, 25b, 27–30Adults of this species (Fig. 13) can be distinguished from those of all other species in the region by the following combination of character states: body length 6.5-7.5 mm; dorsal surface with distinct metallic copper or bronze reflection, non-metallic black in very few specimens, at least femora and outer antennomeres dark (piceous to black); elytral microscuplture comprised of distinctly isodiametric meshes in both males and females (more deeply impressed in females than in males); pronotum (Fig. 13a) more distinctly narrowed anteriorly than basally, anterior margin clearly narrower than posterior margin, lateral margins more or less evenly rounded from apical to basal angle, anterior angles not or only faintly and broadly projected anteriorly beyond anterior margin, posterior angles narrowly rounded, basal impressions deeply foveate but small in diameter, outer basal impressions not distinctly delimited laterally by a broad convexity; elytra without evident sub-basal depressions, parascutellar pore puncture absent, elytral striae distinct throughout but only shallowly impressed in most individuals; medial protibial spurs simple; metatibiae of males without a brush-like patch of setae medially in the apical half; tarsomere 5 of hind tarsi with two or (in a few specimens) three pairs of setae ventrally (Fig. 6b); last abdominal sternite of male with one pair (Fig. 5e) and female with two pairs (Fig. 5c) of setiferous punctures near hind margin; male aedeagus with apical third of median lobe about as broad as middle third, apical hook of right paramere smaller and closer to apex (Fig. 8a).

Apical region of median lobe (dorsal aspect) and apex of right paramere (medial aspect) of aedeagus of males. a Amara lucidissima Baliani b all other species of subgenus Reductocelia (for comparative purposes only, no other species of this subgenus in the area).
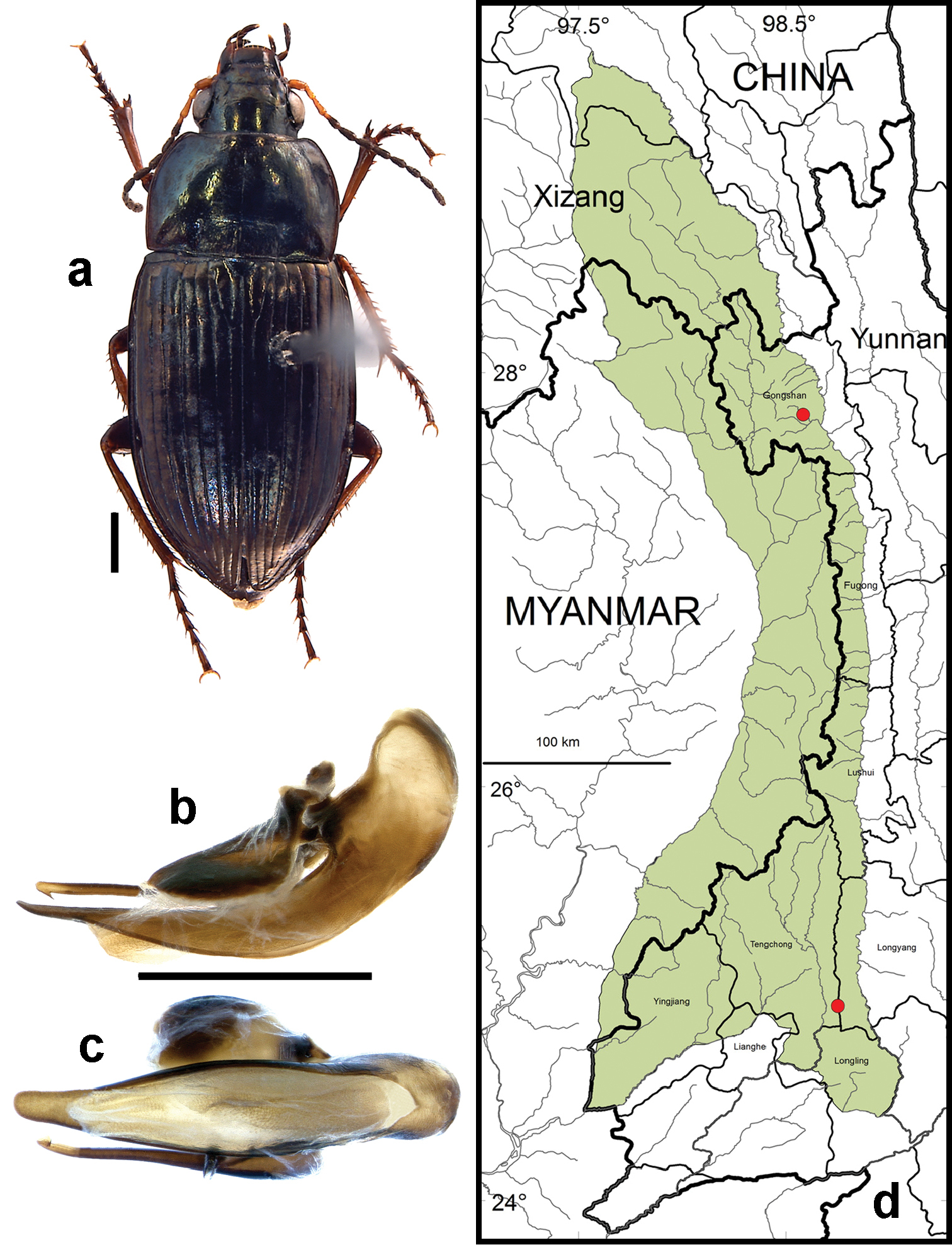
Amara (Zezea) davidi Tschitschérine. a dorsal habitus (CASENT1010925) b–c median lobe of aedeagus of male (CASENT8125447) b left lateral aspect c dorsal aspect; scale lines = 1.0 mm d Map of localities records (red circles) for Amara davidi in the Gaoligong Shan region, scale line = 100 km.
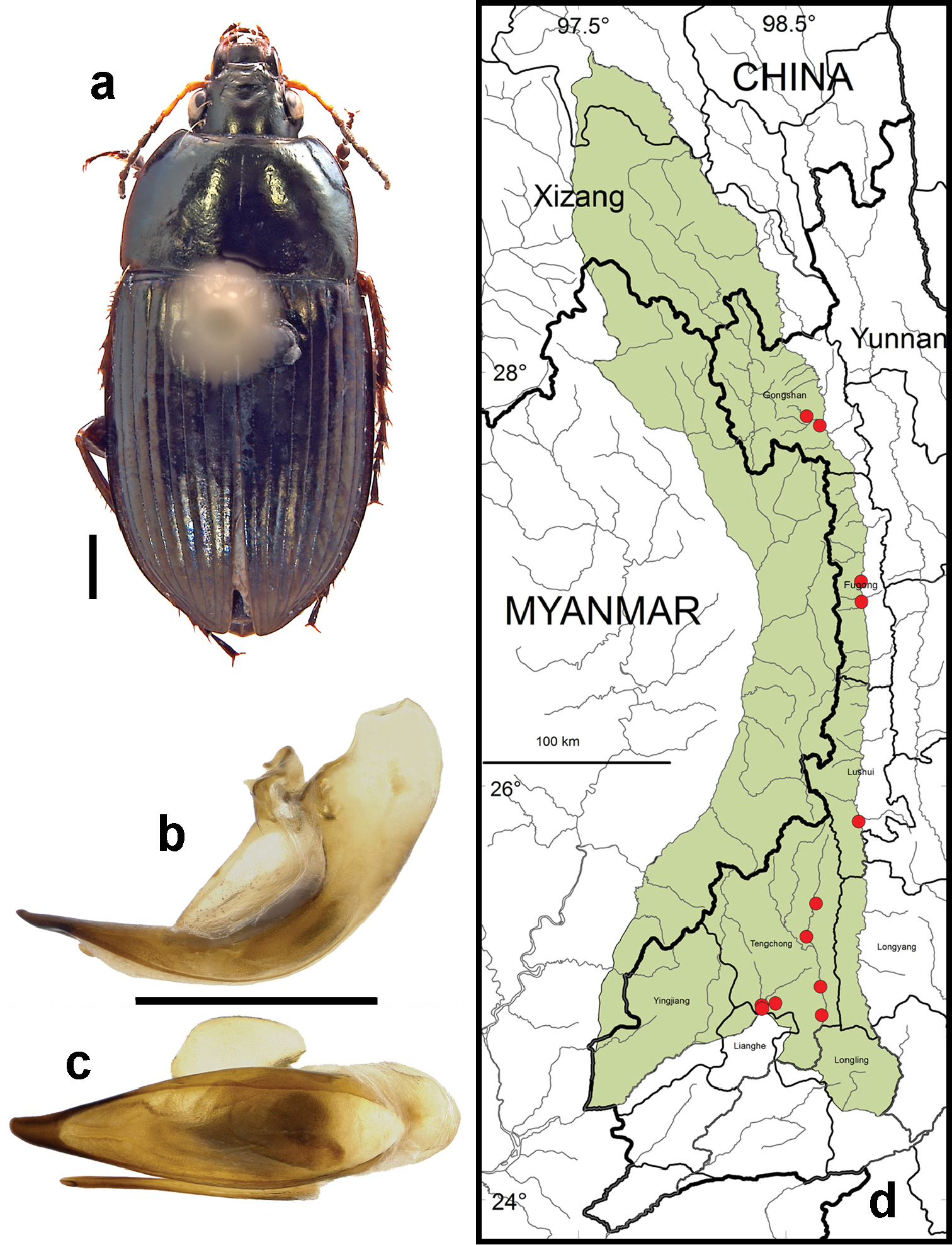
Amara (Amara) congrua Morawitz. a dorsal habitus (CASENT1039647) b–c median lobe of aedeagus of male (CASENT1039647) b left lateral aspect c dorsal aspect; scale lines = 1.0 mm d Map of localities records (red circles) for Amara congrua in the Gaoligong Shan region, scale line = 100 km.

Amara (Amara) silvestrii Baliani. a dorsal habitus (CASENT1035225) b–c median lobe of aedeagus of male (CASENT1035225) b left lateral aspect c dorsal aspect; scale lines = 1.0 mm d Map of localities records (red circles) for Amara silvestrii in the Gaoligong Shan region, scale line = 100 km.
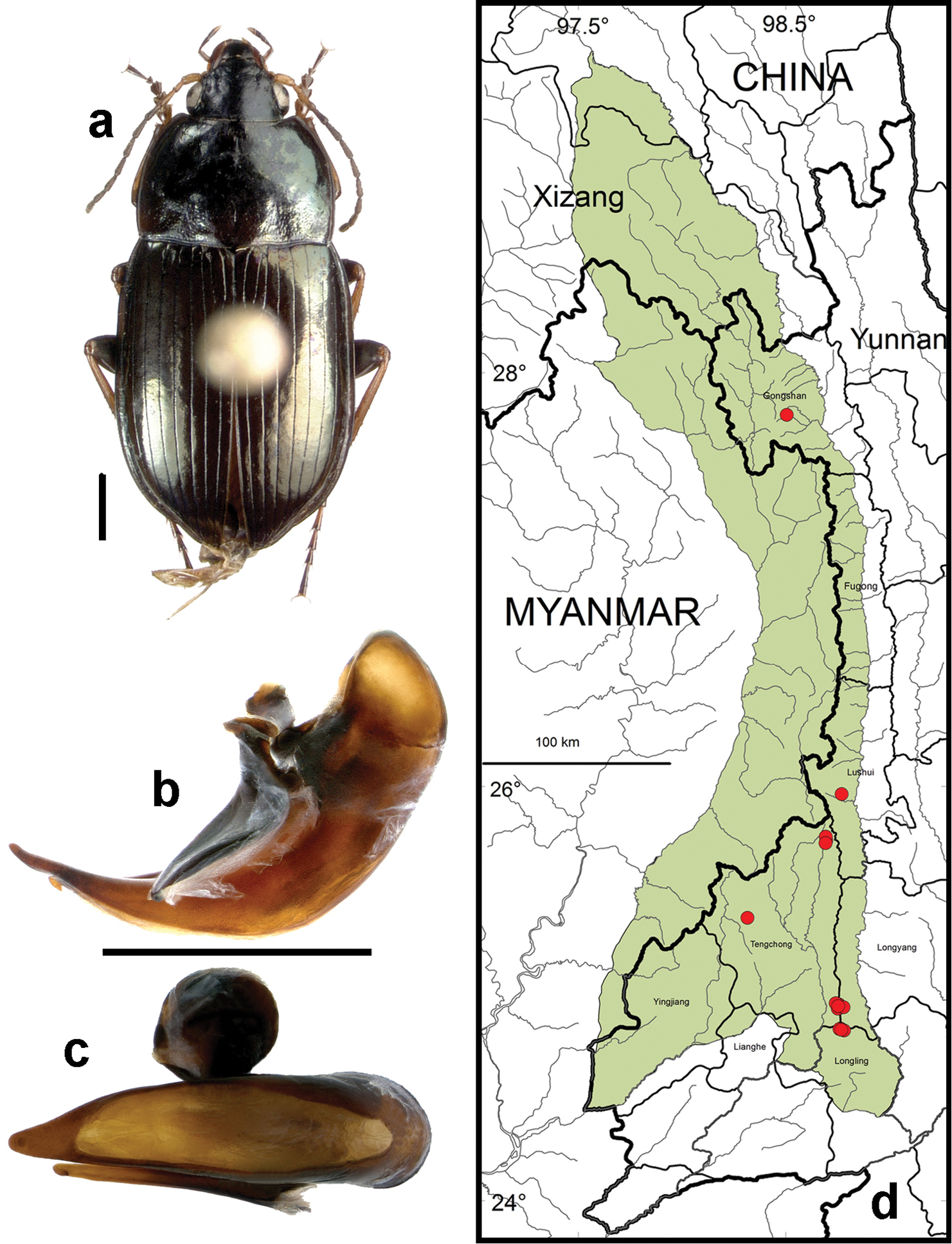
Amara (Amara) shaanxiensis Hieke. a dorsal habitus (CASENT1013215) b–c median lobe of aedeagus of male (CASENT1024088) b left lateral aspect c dorsal aspect; scale lines = 1.0 mm d Map of localities records (red circles) for Amara shaanxiensis in the Gaoligong Shan region, scale line = 100 km.

Amara (Pseudoamara) birmana Baliani. a dorsal habitus (CASENT1011859) b–c median lobe of aedeagus of male (CASENT1039066) b left lateral aspect c dorsal aspect; scale lines = 1.0 mm d Map of localities records (red circles) for Amara birmana in the Gaoligong Shan region, scale line = 100 km.
Specimens of this species were collected, often in great abundance, in daytime from under stones and other cover in open roadside areas (Figs 22a, 25a) and waste areas around human settlements (Figs 23a, 25b) with scattered grasses and shrubs, at the edges of agricultural fields and on open banks of streams, and in these same habitats at night, when beetles were found active on bare substrate. They were also collected under rotting logs on a deciduous forest floor. Members of this species were found at elevations ranging from 1590 to 3150 m, syntopic with adults of Amara chalciope, Amara davidi, Amara dissimilis, Amara lucidissima, Amara pingxiangi, Amara shaanxiensis, Amara sikkimensis and Amara silvestrii at one or more sites.
Fig. 13d. We examined a total of 244 specimens (126 males and 118 females) from the following localities: Longling County: Longjiang Township (small stream along road at 1.2 km SSE of Km 23.5 on Route 23.5, 24.82888°, 98.76001°, 2020 m, 25–27 May 2005, D.H. Kavanaugh, C.E. Griswold, H.B. Liang & D.Z. Dong collectors [4 males and 5 females; CAS, IOZ]), (Xiaoheishan Forest Station, 24.83671°, 98.76185°, 2067 m, 28 May 2005, H.B. Liang, K.J. Guo & H.M. Yan collectors [2 females; CAS, IOZ]). Longyang County: Bawan Township (mountain near Nankang Yakou, 24.83250°, 98.76944°, 2245 m, 27 October 2003, H.B. Liang & X.C. Shi collectors [3 males and 5 females; CAS, IOZ]), (Km 24 on Baoshan-Tengchong Highway near Nankang Yakou, 24.82583°, 98.77222°, 2130 m, 26 October 2003, H.B. Liang & X.C. Shi collectors [6 males and 5 females; CAS, IOZ]), (Km 36–37 on Bawan-Tenchong Road, 24.93417°, 98.77944°, 2150 m, 12 October 2003, H.B. Liang & X.C. Shi collectors [1 female; IOZ]), (Km 40 on Bawan-Tenchong Road at Dasheyao Plant Protection Station, 24.92944°, 98.75861°, 2320 m, 16 October 2003, H.B. Liang & J.J Yang collectors [1 female; IOZ], 3 June 2005, D.H. Kavanaugh, D.Z. Dong & J.J. Yang collectors [1 male and 1 female; CAS, IOZ]), 90 km W of Baoshan, 26–28 May 1995, S. Bečvar collector [1 female; ZMHB]), (Luoshuidong area at Sancha He, 24.94833°, 98.75667°, 2300 m, 26–31 October 1998, D.H. Kavanaugh & C.E. Griswold collectors [6 males and 4 females; CAS, IOZ, ZMHB], 3 June 2005, D.H. Kavanaugh, C.E. Griswold, H.B. Liang, D.Z. Dong & H.M. Yan collectors [1 female; CAS], 24.94865 0176/98.75193°, 2350 m, 30 May 2005, H.B. Liang & J.J. Yang collectors [3 males and 1 female; CAS, IOZ]), (Nankang Forest Station (24.82444°, 98.77889°, 2085 m, 27 October 2003, H.B. Liang & X.C. Shi collectors [9 males and 12 females; CAS, IOZ]), (Nankang Yakou (24.83167°, 98.76667°, 2130 m, 4–7 November1998, D. H. Kavanaugh collector [25 males and 20 females; CAS, IOZ, ZMHB], 24.82587°, 98.76832°, 2148 m, 22 May 2005, H.B. Liang collector [1 male and 1 female; CAS, IOZ], 24.83124°, 98.76843°, 2210 m, 23 May 2005, H.B. Liang collector [2 females; CAS, IOZ]), (just N of Nankang Yakou, 24.83178°, 98.76472°, 2180 m, 22 and 26 May 2005, H.B. Liang & D.Z. Dong collectors [5 males and 4 females; CAS, IOZ]). Lushui County: Luzhang Township (Km 44.7 on Pianma Road at Yaojiaping, 25.97538°, 98.71006°, 2516 m, 11 May 2005, D.H. Kavanaugh, H.B. Liang, D.Z. Dong & G. Tang collectors [1 female; CAS]), (Fengxue Yakou at Pianma Road, 25.97228°, 98.68336°, 3150 m, D.H. Kavanaugh, P.E. Marek & H.B. Liang collectors [1 female; CAS]), (Lusai He, 25.96378°, 98.77032°, 1873 m, 20 May 2005, H.B. Liang & D.Z. Dong collectors [3 amles and 1 female; CAS, IOZ]), (Yaojiaping He at Pianma Road, 25.97722°, 98.71091°, 2527 m, 20 May 2005, D.H. Kavanaugh, C.E. Griswold, H.B. Liang, D.Z. Dong & G. Tang collectors [1 female; IOZ]); Pianma Township (6 km ESE of Pianma, 26.00808°, 98.65921°, 2310 m, 15 May 2005, H.B. Liang & D.Z. Dong collectors [5 males and 6 females; CAS, IOZ]), (20 km N of Pianma along Gangfang He at Gulang Village, 26.10321°, 98.58094°, 1590 m, 14 May 2005, H.B. Liang collector [1 female; IOZ]), (Xia Pianma Village, 26.01137°, 98.61788°, 1850 m, 13 May 2005, H.B. Liang collector [1 male; IOZ]). Tengchong County: Houqiao Township (5.9 airkm NE of Houqiao below Guyong Forestry Station, 25.3562°, 98.31610°, 2030 m, 27 May 2006, D.H. Kavanaugh, R.L. Brett & D.Z. Dong collectors [1 female; IOZ]), (8.5 airkm NNE of Houqiao at Gaoshidong, 25.39858°, 98.30533°, 2580 m, 27 May 2006, D.H. Kavanaugh, R.L. Brett & D.Z. Dong collectors [1 male and 1 female; CAS, IOZ]); Jietou Township (Dahetou Lingganjiao, Longtang He, 25.73947°, 98.69630°, 2010 m, 14–16 and 18 May 2006, D.H. Kavanaugh, R.L. Brett & H.B. Liang collectors [16 males and 11 female; CAS, IOZ], 0.3 km S in Longchuan Jiang valley, 25.73678°, 98.69639°, 2005 m, 18 May 2006, D.Z. Dong collector [4 males and 5 females; CAS, IOZ], 1.4 km S along Longchuan Jiang, 25.72717°, 98.69322°, 1960 m, 19 May 2006, D.H. Kavanaugh, R.L. Brett & D.Z. Dong collectors [2 females; CAS, IOZ], 0.75 km N on Longtang He, 25.74622°, 98.69612°, 2030 m, 18 May 2006, D.H. Kavanaugh & R.L. Brett collectors [1 female; CAS]), (Longchuan Jiang from Dahetou Village to Dahetou Lingganjiao, 25.67125°, 98.68016°, 1838–2010 m, 14 May 2006, H.B. Liang collector [2 males and 1 female; CAS, IOZ]), (Shaba Village, Cha He, 25.39256°, 98.70488°, 1840 m, 25 May 2006, D.H. Kavanaugh, R.L. Brett & D.Z. Dong collectors [1 male and 1 female; CAS]); Shangying Township (Km 41 on Bawan-Tenchong Road near Yakou, 24.93972°, 98.75333°, 2440 m, 15 October 2003, H.B. Liang, X.C. Shi & D.Z. Dong collectors [2 males and 1 female; CAS, IOZ]), (Km 42–46 on Bawan-Tenchong Road, 24.95361°, 98.74222°, 2290 m, 14 and 17 October 2003, H.B. Liang & X.C. Shi collectors [7 males and 2 female; CAS, IOZ]), (Km 46–51 on Bawan-Tenchong Road, 24.95722°, 98.73667°, 2220 m, 17 October 2003, H.B. Liang & X.C. Shi collectors [1 male; IOZ]); Wuhe Township (Km 24 on Baoshan-Tengchong Highway, 24.82889°, 98.76028°, 2008 m, 29 October 2003, N.D. Penny, T. Briggs & X.Y. Li collectors [6 males; CAS, IOZ]), (Km 28.8 on Route S317 at Zhengding Forestry Station, 24.84855°, 98.73761°, 1834 m, 23 May 2005, D.Z. Dong & H.M. Yan collectors [1 male and 2 females; CAS, IOZ]), (31 km SE of Tengchong, 24.88639°, 98.75611°, 26 August 2009, D.W. Wrase collector [1 male and 2 females; ZMBH]), (33 km SE of Tengchong, 24.85611°, 98.76000°, 2100–2200 m, 31 May and 4 June 2007, A. Pütz collector [2 males and 1 female; ZMHB], 31 May 2007, D.W. Wrase collector [1 male; ZMHB]), (100 km W of Baoshan, 14–21 June 1993, E. Jendek & O. Sausa collectors [1 male; ZMHB]), (Xiaodifang Village, 24.85722°, 98.75917°, 2150 m, 29 October 2003, D.Z. Dong collector [3 males and 1 female; CAS, IOZ]), (Xiaoheishan Forest Station, 24.82889°, 98.76000°, 2025 m, 29 October 2003, H.B. Liang collector [6 males and 7 females; CAS, IOZ]).
Members of this species were collected only in the southern half of the study area (Core Areas 4, 5, 6 and 7), on both eastern and western slopes of the mountain range.
Fig. 27. This species has been recorded from China (Sichuan and Yunnan Provinces), India (Assam, Arunashal Pradesh and Sikkim) and Myanmar. Its occurrence in the study area represents the southern limit of its known geographical range.
http://species-id.net/wiki/Amara_sikkimensis
Figs 5b–d, 6b, 14, 23a, 23b, 27–30Adults of this species (Fig. 14) can be distinguished from those of all other species in the region by the following combination of character states: body length 7-8 mm; dorsal surface dark piceous; pronotum with both inner and outer basal impressions deeply impressed; elytra with parascutellar pore puncture absent; medial protibial spurs simple; tarsomere 5 of hind tarsi with two or (in a few specimens) three pairs of setae ventrally (Fig. 6b); last abdominal sternite of both male (Fig. 5d) and female (Fig. 5c) with two pairs of setiferous punctures near hind margin (second seta absent from one side in a few males for total of three setae).
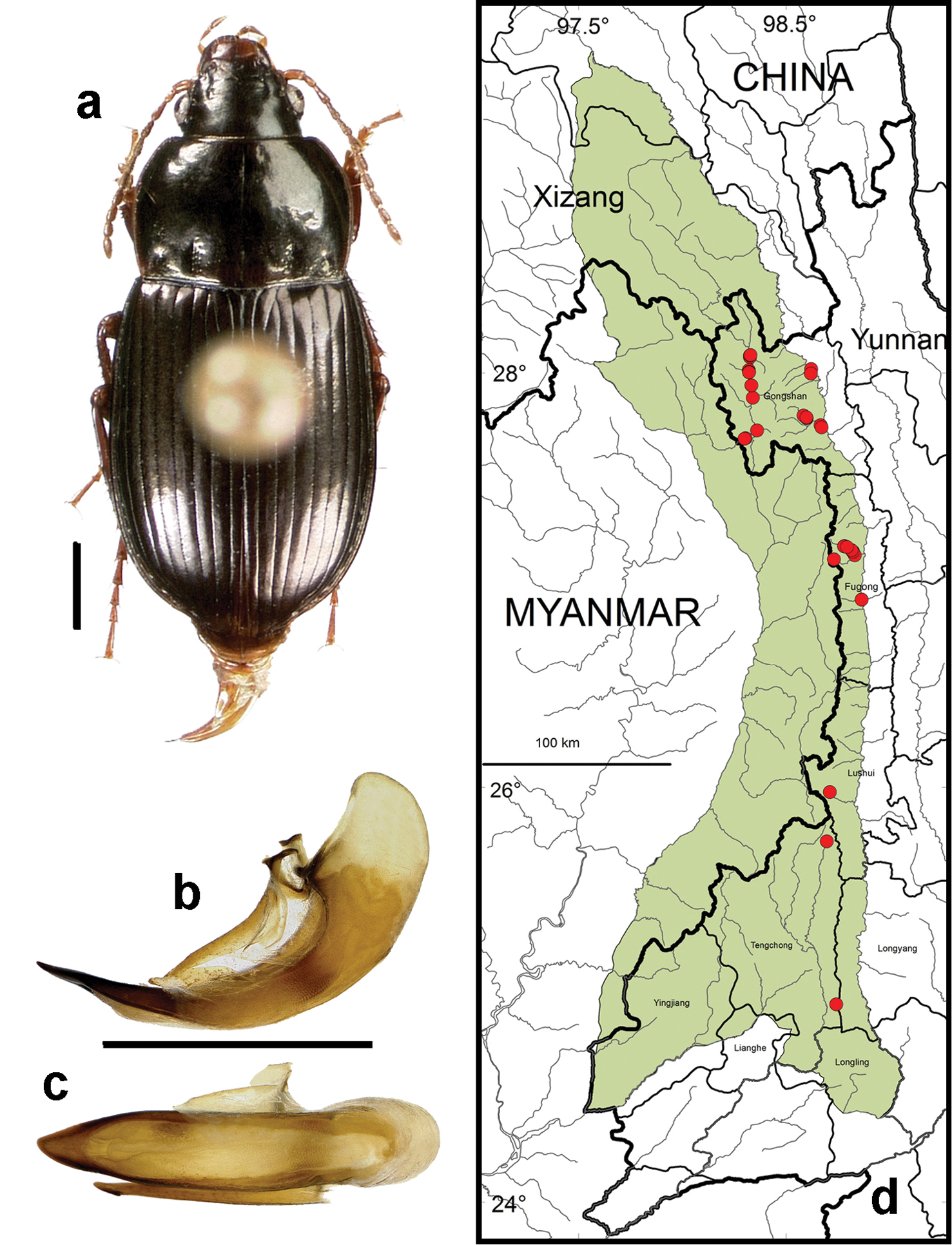
Amara (Xenocelia) sikkimensis Andrewes. a dorsal habitus (CASENT1015288) b–c median lobe of aedeagus of male (CASENT1025342) b left lateral aspect c dorsal aspect; scale lines = 1.0 mm d Map of localities records (red circles) for Amara sikkimensis in the Gaoligong Shan region, scale line = 100 km.
Specimens of this species were collected in daytime from under stones and other cover in open roadside areas (Fig. 23b) and other waste areas around human settlements (Fig. 23a) with scattered grasses and shrubs, at the edges of agricultural fields, and on open rocky, graveled and sandy banks of streams, and in these same habitats at night, when beetles were found active on bare substrate. They were especially conspicuous on open sandy beaches at night. Members of this species were found at elevations ranging from 1175 to 2800 m, syntopic with adults of Amara birmana, Amara chalciope, Amara davidi, Amara dissimilis, Amara latithorax, Amara lucidissima, Amara shaanxiensis and Amara silvestrii at one or more sites.
Fig. 14d. We examined a total of 164 specimens (92 males and 72 females) from the following localities: Fugong County: Lishadi Township (Shibali area, 27.16536°, 98.78003°, 2535 m, 6 October 2007, H.L. Shi collector [1 female; IOZ]), (0 to 2 km E of Shibali on Shibali Road, 27.16100°, 98.79370°, 2300–2350 m, 18 August 2005, D.Z. Dong collector [1 male and 1 female; IOZ]), (0.1 km below Shibali on Shibali Road, 27.16577°, 98.78091°, 2545 m, 5 May 2004, D.H. Kavanaugh & C.E. Griswold collectors [1 male; CAS]), (0.3 km SE of Shibali on North Fork Yamu He, 27.16337°, 98.78208°, 2475 m, 7 May 2004, D.H. Kavanaugh & C.E. Griswold collectors [1 male and 1 female; CAS]), (1.5 km below Shibali on Yaping Road, 27.16284°, 98.78989°, 2420 m, 2 May 2004, H.B. Liang & G.X. Peng collectors [1 male and 1 female; CAS, IOZ]), (Shilajia Village at Shibali Road, 27.13947°, 98.82184°, 1800–1900 m, 24–25 April 2004, D.H. Kavanaugh & C.E. Griswold collectors [2 females; CAS, IOZ]), (11 km above Nu Jiang on Yaping Road at Shimowa Village, 27.13839°, 98.82147°, 1850–1928 m, 25 April 2004, H.B. Liang collector [4 males and 5 females; CAS, IOZ]); Lumadeng Township (7.0 km SW of Lao Shibali on Lao Shibali Road at tributary of South Fork Yamu He, 27.10220°, 98.73107°, 2800 m, 13 August 2005, D.H. Kavanaugh & P. Paquin collectors [1 male; CAS]), (7.5 km below Shibali on Yaping Road, 27.14627°, 98.81559°, 2030 m, 3 May 2004, H.B. Liang & M. Xie collectors [2 males and 2 female; CAS, IOZ]), (South Fork Yamu He above Shilajia Village, 27.12101°, 98.83173°, 1630–1790 m, 26 April 2004, D.H. Kavanaugh collector [1 female; CAS]), (Yaping Road below Shibali, 27.16520°, 98.77980°, 2530 m, 24 April 2004, H.B. Liang & X.Y. Li collectors [1 male; IOZ]); Shangpa Township (Nu Jiang at west end of footbridge in Shangpa, 26.90743°, 98.86391°, 1175–1180 m, 20 April 2004, H.B. Liang collector [1 female; IOZ]). Gongshan County: Bingzhongluo Township (Bingzhongluo Village, 28.01940°, 98.62106°, 1760 m, 21 April 2002, H.B. Liang & W.D. Ba collectors [2 males and 1 female; IOZ, ZMHB]), (Gongdangshenshan, 27.99725°, 98.62003°, 2489 m, 12 November 2004, H.B. Liang collector [1 male; CAS]); Cikai Township (Heiwadi, 16.8 km W of Cikai on Dulong Valley Road, 27.79584°, 98.58443°, 2020 m, 20 April 2002, H.B. Liang & W.D. Ba collectors [2 males; IOZ]), (Heiwadi Village, 27.79584°, 98.58443°, 1965 m, 14 November 2004, H.B. Liang, D.Z. Dong & G. Tang collectors [26 males and 14 females; CAS, IOZ]), (North Fork Pula He above Heiwaidi Village, 27.78644°, 98.59831°, 1890 m, 15 November 2004, D.H. Kavanaugh & V.F. Lee collectors [3 males; CAS, IOZ]), (Nu Jiang in Cikai at Dashaba, 27.73845°, 98.67092°, 1430 m, 11 November 2003, D.H. Kavanaugh, H.B. Liang, D.Z. Dong & G. Tang collectors [1 female; CAS]), (Pula He just above Nu Jiang Road, 27.74861°, 98.66675°, 1440 m, 11 November 2004, D.H. Kavanaugh, H.B. Liang, D.Z. Dong & G. Tang collectors [1 female; CAS]); Dulongjiang Township (Bapo, Miliwang, 27.72383°, 98.36117°, 1956 m, 31 October 2004, H.B. Liang collector [21 males and 8 females; CAS, IOZ, ZMHB]), (0.6 km N of Dizhengdang Village on Dulong Jiang, 28.08442°, 98.32652°, 1880 m, 29–30 October 2004, D.H. Kavanaugh, D.Z. Dong & G. Tang collectors [4 males and 5 females; CAS, IOZ]), (S of Dizhengdang Village at Silalong He, 28.07654°, 98.32603°, 1890 m, 30 October 2004, D.H. Kavanaugh, D.Z. Dong & G. Tang collectors [2 males and 3 females; CAS, IOZ]), (Dizheng Wang, 28.08686°, 98.32840°, 1900–1970 m, 30 October 2004, D.H. Kavanaugh & D.Z. Dong collectors [1 female; CAS]), (Dulong Jiang at Elideng Village, 28.00287°, 98.32145°, 1640 m, 3 November 2004, D.H. Kavanaugh, H.B. Liang, D.Z. Dong & G. Tang collectors [1 male and 2 females; CAS, IOZ]), (Dulong Jiang at Xiajiudang Village, 27.94092°, 98.33340°, 1580 m, 4 November 2004, D.H. Kavanaugh, D.Z. Dong & G. Tang collectors [2 females; CAS, IOZ]), (0.5 km N of Kongdang, 27.88111°, 98.34063°, 1500 m, 25 October 2004, D.H. Kavanaugh, H.B. Liang, D.Z. Dong & G. Tang collectors [1 female; CAS], 5 November 2004, H.B. Liang collector [1 female; IOZ]), (2.3–3.3 airkm S of Longyuan Village on Dulong Jiang, 28.00532°, 98.32145°, 1685–1720 m, 2 November 2004, D.H. Kavanaugh, D.Z. Dong & G. Tang collectors [2 males and 1 female; CAS, IOZ]), (Maku, 27.68553°, 98.30425°, 1823 m, 2 November 2004, H.B. Liang collector [14 males and 13 females; CAS, IOZ]), (Siran Wang, 0.2 km above confluence with Dulong Jiang, 28.01347°, 98.32117°, 1720 m, 2 November 2004, D.H. Kavanaugh & D.Z. Dong collectors [1 female; CAS]). Lushui County: Luzhang Township (Yaojiaping He at Pianma Road, 25.97722°, 98.71091°, 2527 m, 20 May 2005, D.H. Kavanaugh, C.E. Griswold, H.B. liang, D.Z. Dong & G. Tang collectors [1 male; CAS]). Tengchong County: Jietou Township (Longtang He at Dahetou Lingganjiao, 25.73947°, 98.69630°, 2010 m, 20 May 2005, H.B. Liang & P. Hu collectors [1 female; IOZ]); Shangying Township (Km 42–46 on Bawan-Tengchong Road, 24.95361°, 98.74222°, 2290 m, 14 October 2003, H.B. Liang & X.C. Shi collectors [1 male; IOZ]).
Members of this species were collected from the northern to the southern parts of the study area (Core Areas 1, 2, 3, 5 and 6), but they were found only on the western side of the mountain range in the southern part (Core Area 6). This distribution pattern may be an artifact of inadequate sampling on the eastern slope of the mountain range in the south.
Fig. 27. This species has been recorded from Bhutan, China (Gansu, Sichuan and Yunnan Provinces and Xizang Autonomous Region), India (Assam, Himachal Pradesh, Kashmir and Jammu, Sikkim and Uttar Pradesh), Nepal and Pakistan (northern). Its occurrence in the study area represents the southern limit of its known geographical range.
http://species-id.net/wiki/Amara_latithorax
Figs 5b–c, 5e, 6b, 15, 22b, 23b, 27–30Adults of this species can be distinguished from those of all other species in the region by the following combination of character states: body length 8.5-9.0 mm; dorsal surface black, with very faint metallic blue or green metallic reflection in most individuals, more vivid (as in Fig. 15a) in or lacking from a few specimens, at least femora and outer antennomeres dark (piceous to black); pronotum (Fig. 15a) only slightly narrowed anteriorly with anterior margin nearly straight and clearly narrower than posterior margin, without or with only very slightly projected anterior angles, posterior angles distinctly rounded (narrowly so in some individuals), slightly to moderately obtuse, pronotal base moderately coarsely punctate, outer basal impressions either not sharply delimited laterally by raised areas or, if so, then the raised area broader; elytra elytra with slight sub-basal depressions centered on striae 6 (visible on right elytron in Fig. 15a), also on striae 4 and/or 5 in some individuals, parascutellar pore puncture absent; medial protibial spurs simple; tarsomere 5 of hind tarsi with two or (in a few specimens) three pairs of setae ventrally (Fig. 6b); last abdominal sternite of male with one pair (Fig. 5e) and female with two pairs (Fig. 5c) of setiferous punctures near hind margin.
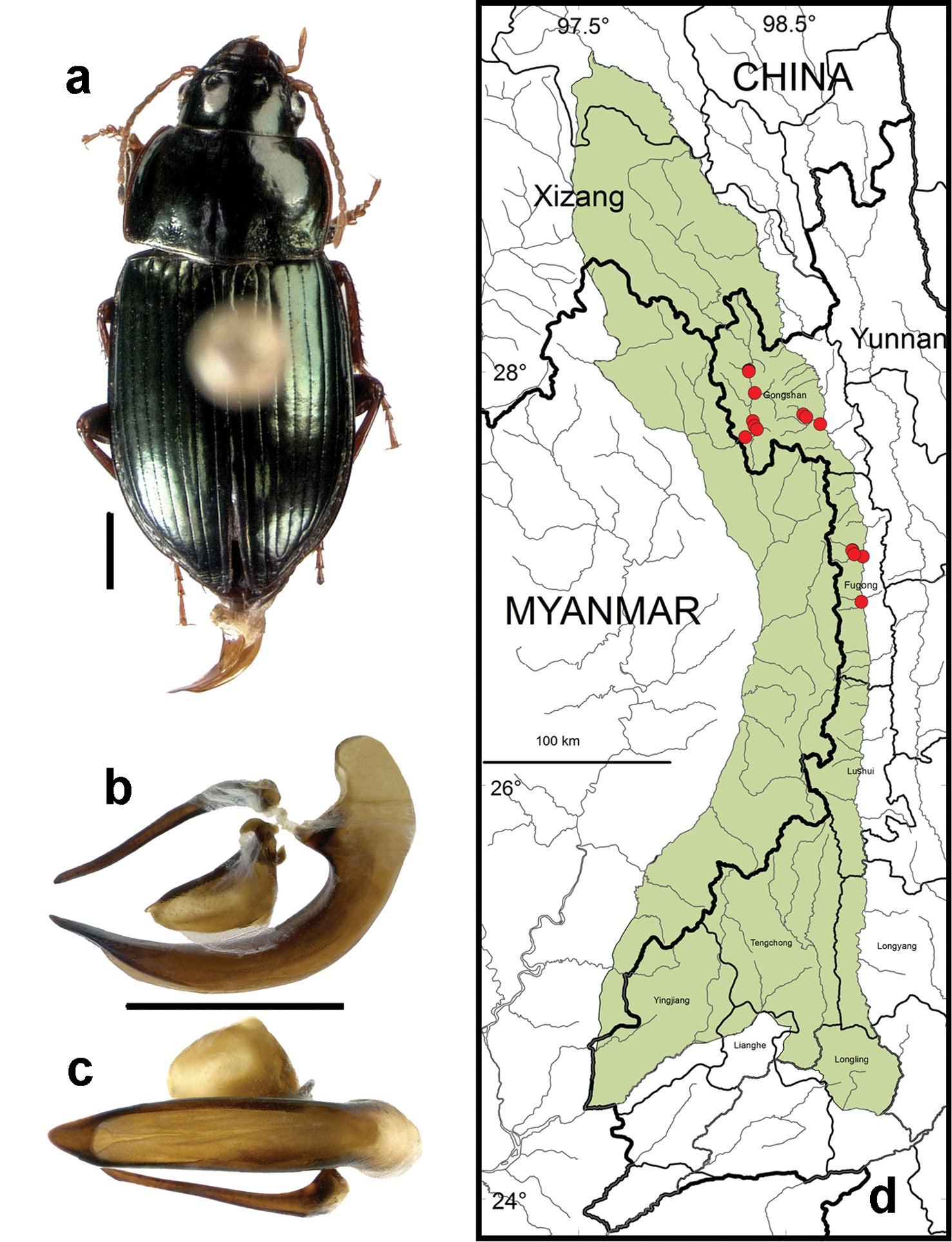
Amara (Harpaloamara) latithorax Baliani. a dorsal habitus (CASENT1015304); b-c. median lobe of aedeagus of male (CASENT10115091) b left lateral aspect c dorsal aspect; scale lines = 1.0 mm d Map of localities records (red circles) for Amara in the Gaoligong Shan region, scale line = 100 km.
Specimens of this species were collected in daytime from under stones and other cover in open roadside areas (Figs 22b, 23b) and meadows with scattered grasses and shrubs, at the edges of agricultural fields, including wet and dry rice paddies and on open sandy banks of streams, and in these same habitats at night, when beetles were found active on bare substrate. Members of this species were found at elevations ranging from 1195 to 2022 m, syntopic with adults of Amara chalciope, Amara congrua, Amara dissimilis, Amara lucidissima and Amara sikkimensis at one or more sites.
Fig. 15d. We examined a total of 37 specimens (12 males and 25females) from the following localities: Fugong County: Aludi Village (Nu Jiang, 27.10834°, 98.87218°, 1195-1250 m, 22 April 2004, D.H. Kavanaugh collector [1 female; CAS]); Lishadi Township (Shilajia Village at Shibali Road on North Fork Yamu He, 27.13947°, 98.82184°, 1800–1900 m, 24-25 April 2004, D.H. Kavanaugh & C.E. Griswold collectors [1 female; CAS]), (11 km above Nu Jiang on Yaping Road at Shimowa Village, 27.13839°, 98.82147°, 1850-1928 m, 25 April 2004, H.B. Liang collector [1 male and 2 females; CAS, IOZ, ZMHB]); Shangpa Township (road on west side of Nu Jiang S of Shangpa, 26.88952°, 98.86539°, 1223 m, 22 and 27 April 2004, D.H. Kavanaugh & C.E. Griswold collectors [1 male and 1 female; CAS, IOZ]). Gongshan County: Cikai Township (15 km W of Cikai on Dulong Valley Road, 27.79584°, 98.58443°, 2022 m, 10 October 2002, D.H. Kavanaugh, P.E. Marek & H.B. Liang collectors [1 female; CAS]), (Heiwadi Village, 27.72250°, 98.59902°, 1965 m, 14 November 2004, H.B. Liang, D.Z. Dong & G. Tang collectors [2 males; IOZ, ZMHB]), (Pula He just above Nu Jiang Road, 27.74861°, 98.66675°, 1440 m, 23 October 2004, D.H. Kavanaugh & H.B. Liang collectors [1 female; IOZ], 11 November 2004, D.H. Kavanaugh, H.B. Liang, D.Z. Dong & G. Tang collectors [1 male and 2 females; CAS, IOZ]); Dulongjiang Township (Bapo, 27.73902°, 98.34975°, 1412 m, 20 October 2004, H.B. Liang collector [1 female; IOZ], 3 November 2004, H.B. Liang collector [2 males and 1 female; CAS, IOZ], at Miliwang, 27.72383°, 98.36117°, 1956 m, 31 October 2004, H.B. Liang collector [6 females; CAS, IOZ]), (Dulong Jiang at Elideng Village, 28.00287°, 98.32145°, 1640 m, 3 November 2004, D.H. Kavanaugh, D.Z. Dong & G. Tang collectors [1 female; CAS]), (2.3-3.3 airkm S of Longyuan Village on Dulong Jiang, 28.00532°, 98.32145°, 1685-1720 m, 2 November 2004, D.H. Kavanaugh, D.Z. Dong & G. Tang collectors [1 female; IOZ], 2.8 km S of Longyuan Village on Dulong Jiang, 28.00905°, 98.32204°, 1660 m, 2 November 2004, D.H. Kavanaugh & D.Z. Dong collector [1 male; CAS]), (main road between Mabidang and Kongdang, 27.76361°, 98.34111°, 1329 m, 5 November 2004, V.F. Lee & D.G. Long collectors [1 female; CAS]), (Maku, 27.68553°, 98.30425°, 1823 m, 2 November 2004, H.B. Liang collector [2 males and 3 females; CAS, IOZ]), (Moqie Wang at KM 91 on Gongshan-Dulong Road, 27.89934°, 98.34999°, 1550 m, 6 November 2004, D.H. Kavanaugh & H.B. Liang collectors [1 male and 1 female; CAS, IOZ]).
Members of this species were collected only in the northern half of the study area (Core Areas 1, 2 and 3), on both sides of the mountain range (Core Areas 1 and 2).
Fig. 27. This species has been recorded from Bhutan, China (Yunnan Province), India (Sikkim, Uttar Pradesh and West Bengal), Myanmar and Nepal. Its occurrence in the study area represents the southern and eastern limits of its known geographical range.
http://species-id.net/wiki/Amara_chalciope
Figs 5b–c, 5e, 6b, 16, 24a, 24b, 25a, 26a, 27–30Adults of this species (Fig. 16) can be distinguished from those of all other species in the region by the following combination of character states: body length 7.5–9.0 mm; dorsal surface dark with distinct metallic blue-green reflection in most specimens, non-metallic black in a few specimens, at least femora and outer antennomeres dark (piceous to black); elytral microsculpture comprised of isodiametric meshes, faintly impressed or nearly effaced in males, deeply impressed and distinct in females; pronotum (Fig. 16a) with lateral margins straight or faintly to distinctly sinuate just anterior to basal angles, rounded near middle, less rounded or nearly straight also in anterior one-third in most specimens, anterior angles distinctly and narrowly projected anteriorly beyond anterior margin, lateral explanation distinctly broader basally, outer basal impressions indistinct from lateral groove in most specimens, punctation of base extended anteriorly along sides to pronotal mid-length; elytra with parascutellar pore puncture absent; medial protibial spurs simple; metatibia of male without brush-like setae medially in the apical half; tarsomere 5 of hind tarsi with two or (in a few specimens) three pairs of setae ventrally (Fig. 6b); last abdominal sternite of male with one pair (Fig. 5e) and female with two pairs (Fig. 5c) of setiferous punctures near hind margin.
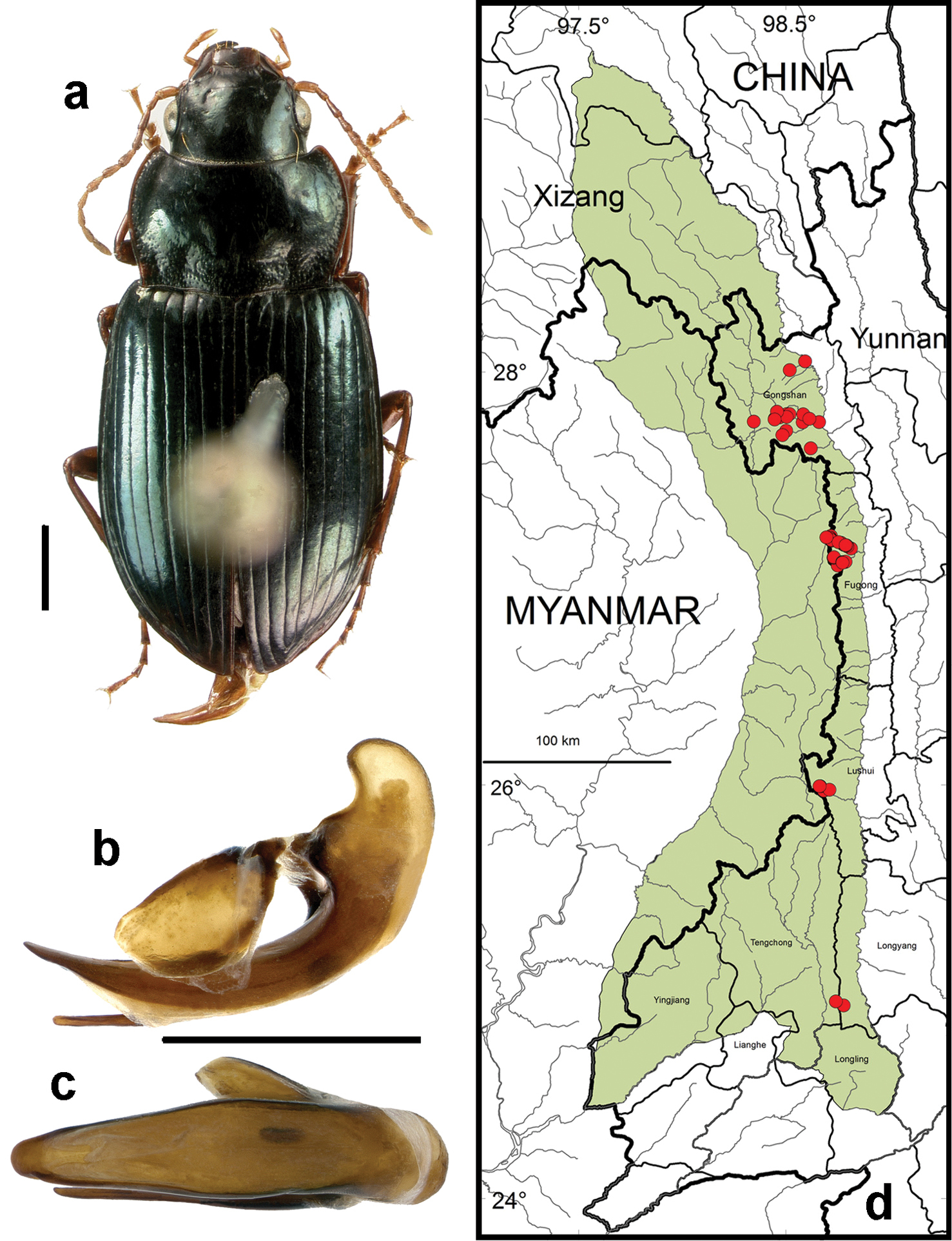
Amara (Bradytus) chalciope Bates. a dorsal habitus (CASENT1028265) b–c median lobe of aedeagus of male (CASENT1028375) b left lateral aspect c dorsal aspect; scale lines = 1.0 mm d Map of localities records (red circles) for Amara chalciope in the Gaoligong Shan region, scale line = 100 km.
Specimens of this species were collected in daytime from under stones and other cover in open roadside areas (Figs 24b, 25a, 26a), particularly on bare granitic sand substrates on roadcuts, landslides, stream banks, flood outwash flats and open slopes above treeline (Fig. 24a), and in these same habitats at night, when beetles were found active on bare substrate. Members of this species were found at elevations ranging from 1500 to 3611 m (most abundantly between 2500 and 3200 m), and syntopic with adults of Amara birmana, Amara dissimilis, Amara elegantula, Amara latithorax, Amara lucidissima, Amara shaanxiensis, Amara sikkimensis, Amara silvestrii at one or more sites.
Fig. 16d. We examined a total of 634 specimens (296 males and 338 females) from the following localities: Fugong County: Lishadi Township (Shibali area, 27.16553°, 98.77854°, 2550 m, 5 and7 May 2004, D.H. Kavanaugh, H.B. Liang, X.Y. Li, X.Q. Li and H.M. Yan collectors [12 females; IOZ]), 27.16536°, 98.78003°, 2535 m, 4–17 August 2005, D.H. Kavanaugh, H.B. Liang, P. Paquin & D.Z. Dong collectors [9 males and 7 females; CAS, IOZ], 4–5 October 2007, D.H. Kavanaugh, H.B. Liang & H.L. Shi collectors [1 male and 5 females]), (0.3 km SE of Shibali at North Fork Yamu River, 27.16337°, 98.78208°, 2475 m, 7 May 2004, D.H. Kavanaugh & C.E. Griswold collectors [2 males; CAS]), (1.5 km below Shibali on Yaping Road, 27.16284°, 98.78989°, 2420 m, 2 May 2004, H.B. Liang & G.X. Peng collectors [2 males and 6 females; CAS, IOZ]), (2.0 km E of Shibali on Shibali Road, 27.16276°, 98.78927°, 2430 m, 6 August 2005, H.B. Liang collector [5 males and 2 females; CAS, IOZ], 27.16100°, 98.79370°, 2300–2350 m, 18 August 2005, D.Z. Dong collector [3 males and 7 females; CAS, IOZ]), (2.7 km above of Shibali on Shibali Road, 27.17368°, 98.76684°, 2735 m, 10 August 2005, D.H. Kavanaugh collector [2 females; CAS, IOZ]), (2.8 km above of Shibali on Shibali Road, 27.17405°, 98.76722°, 2750 m, 9 August 2005, D.Z. Dong collector [1 male and 1 female; CAS, IOZ]), (4 km E of Shibali on Shibali Road, 27.15727°, 98.79784°, 2280 m, 11 August 2005, D.Z. Dong collector [1 male and 1 female; CAS IOZ]), (4 km W of Shibali on Shibali Road, 27.17740°, 98.75490°, 2800 m, 16 August 2005, D.Z. Dong collector [2 males and 1 female; CAS IOZ]), (8.4 to 9.5 km W of Shibali on Shibali Road, 27.18740°, 98.71936°, 3160–3195 m, 13 August 2005, D.H. Kavanaugh, H.B. Liang & D.Z. Dong collectors [1 male and 1 female; CAS, IOZ]), (8.5 km above Shibali on Shibali Road at North Fork Yamu River, 27.18416°, 98.72026°, 3100 m, 7 May 2004, D.H. Kavanaugh & B.X. Zhu collectors [1 male; CAS]), (9.5 to10.0 km W of Shibali on Shibali Road, 27.19438°, 98.71486°, 3195–3200m, 12 August 2005, D.H. Kavanaugh, H.B. Liang & D.Z. Dong collectors [15 males and 12 females; CAS, IOZ, ZMHB]), (10 km above Shibali on Shibali Road, 27.20055°, 98.71399°, 3121 m, 5–6 August 2005, D.H. Kavanaugh, P. Paquin & D.Z. Dong collectors [6 males and7 females; CAS, IOZ]), (10 km above Shibali on Shibali Road, 27.19980°, 98.71375°, 3200 m, 16 August 2005, J.F. Zhang collector [10 males and 9 females; CAS, IOZ]), (10 to 11 km above Shibali on Shibali Road, 27.19980°, 98.71375°, 3200–3280 m, 8 August 2005, D.H. Kavanaugh, H.B. Liang, P. Paquin & D.Z. Dong collectors [2 males and 3 females; CAS, IOZ]), (10.1 km above Shibali on Shibali Road, 27.20049°, 98.71354°, 3225 m, 6 May 2004, D.H. Kavanaugh, C.E. Griswold, H.B. Liang & B.X Zhu collectors [3 males and 1 female; CAS, IOZ]), (10.1 to11.5 km above Shibali on Shibali Road, 27.20676°, 98.71763°, 3225–3290 m, 8 May 2004, D.H. Kavanaugh, C.E. Griswold, H.B. Liang, X.Y. Li & Z.B. Xi collectors [1 male; IOZ]); Lumadeng Township (8.5 km above Shibali on Shibali Road at North Fork Yamu He, 27.18416°, 98.72026°, 3100 m, 8 August 2005, D.H. Kavanaugh, P. Paquin, D.Z. Dong & J.F. Zhang collectors [1 female; CAS], (7.5 km below Shibali on Yaping Road, 27.14627°, 98.81559°, 2030 m, 3 May 2004, H.B. Liang & M. Xie collectors [2 males; CAS, IOZ]), (Lao Shibali wood inspection station, 27.07831°, 98.77416°, 2305 m, 21 August 2005, H.B. Liang & J.F. Zhang collectors [2 males; CAS, IOZ]), (1 km E of Lao Shibali on Lao Shibali Road at South Fork Yamu He, 27.08141°, 98.78273°, 2275 m, 15 August 2005, D.H. Kavanaugh collector [1 males; CAS]), (1.3 km E of Lao Shibali on Lao Shibali Road at South Fork Yamu He, 27.08180°, 98.78670°, 2250 m, 15 August 2005, D.H. Kavanaugh, H.B. Liang & J.F. Zhang collectors [2 males and 6 females; CAS, IOZ]), (1.6 km E of Lao Shibali on Lao Shibali Road at South Fork Yamu He, 27.08260°, 98.78877°, 2240 m, 21 August 2005, D.H. Kavanaugh collector [5 males and 9 females; CAS, IOZ]), (6.7 km E of Lao Shibali on Lao Shibali Road at tributary of South Fork Yamu He, 27.10437°, 98.73253°, 2805 m, 13 August 2005, D.H. Kavanaugh, H.B. Liang, D.Z. Dong & J.F. Zhang collectors [1 male and 3 females; CAS, IOZ]), (7.0 km E of Lao Shibali on Lao Shibali Road at tributary of South Fork Yamu He, 27.10220°, 98.73107°, 2800 m, 13 August 2005, D.H. Kavanaugh & P. Paquin collectors [5 males; CAS, IOZ]); Lao Shibali Yakou (27.06429°, 98.75123°, 3270 m, 13 August 2005, D.H. Kavanaugh & D.Z. Dong collectors [1 male and 2 females; CAS, IOZ]); ridge and cirques S of Shibali Yakou (27.20035°, 98.69604°, 3599–3611 m, 5 October 2007, H.L. Shi, H.B. Liang & X.J. Peng collectors [1 female; IOZ]); South Fork Yamu He (at Lao Shibali, 27.07978°, 98.77328°, 2305 m, 15 August 2005, D.H. Kavanaugh & D.Z. Dong collectors [ 2 females; IOZ]), (at Yejiadi, 27.08004°, 98.77325°, 2307 m, 10 May 2004, H.B. Liang, X.Y. Li and B.X. Zhu collectors [3 males and 2 females; CAS, IOZ]). Gongshan County: Bapo (2 km N along Dulong Jiang, 27.76000°, 98.34611°, 1510 m, 16–17 July 2000, P. Thomas collector [2 males; CAS, IOZ]), Bingzhongluo Township (Guocai He at Fucai, 28.00855°, 98.51886°, 2800 m, 16 August 2006, D.Z. Dong collector [1 male; IOZ]), (Niwaluo He just above Nu Jiang Road, 28.05140°, 98.59319°, 1630 m, 8 October 2002, D.H. Kavanaugh, P.E. Marek & D.Z. Dong collectors [1 female; CAS]); Cikai Township (8.3 to 13.1 km NW of Cikai on Dulong Valley Road, 27.75653°, 98.58214°, 2630–3000 m, 23 September 2002, D.H. Kavanaugh, P.E. Marek & D.Z. Dong collectors [6 males and 10 females; CAS, IOZ]), (Km 49 on Gongshan-Dulong Road, 27.78075°, 98.47000°, 3330m, 1 October 2002, D.H. Kavanaugh collector [5 males and 6 females; CAS, IOZ]), (53 km W of Cikai on Dulong Valley Road, 27.77422°, 98.44716°, 3380 m, 1 October 2002, D.H. Kavanaugh & H.B. Liang collectors [2 females; CAS, IOZ]), (57 km W of Cikai on Dulong Valley Road, 27.80789°, 98.45736°, 3162 m, 2 October 2002, H.B. Liang collector [2 males and 1 female; CAS, IOZ]); Heipu Yakou ((southeast slope, 27.77032°, 98.44674°, 3365 m, 11and 13 August 2006, D.H. Kavanaugh, J.A. Miller, D.Z. Dong & Y. Liu collectors [6 males and 4 females; CAS, IOZ]); Dabadi (40 km W of Cikai on Dulong Valley Road, 27.79619°, 98.51867°, 3900 m, 29 September 2002, H.B. Liang collector [38 males and 28 females; CAS, IOZ, ZMHB]), (41 km W of Cikai on Dulong Valley Road, 27.79655°, 98.50562°, 3000 m, 27 September – 6 October 2002, D.H. Kavanaugh, P.E. Marek, H.B. Liang, D.Z. Dong & X.C. Li collectors [67 males and 70 females; CAS, IOZ, ZMHB]), (45 km W of Cikai on Dulong Valley Road, , 27.78253°, 98.50444°, 3133 m, 2 October 2002, H.B. Liang collector [2 males and 2 females; CAS, IOZ]); Danzhu He drainage (27.63063°, 98.62074°, 2700 m, 30 June-5 July 2000, D.H. Kavanaugh, C.E. Griswold & H.B. Liang collectors [20 males and 34 females; CAS, IOZ, ZMHB]); Heiwadi (15 km W of Cikai on Dulong Valley Road, 27.79584°, 98.58443°, 2022 m, 4 October 2002, H.B. Liang, W.D. Ba and C.G. Jin collectors [1 male and 1 female; CAS, IOZ], 10 October 2002, D.H. Kavanaugh, P.E. Marek & H.B. Liang collectors [11 males and 4 females; CAS, IOZ]), (27.77415°, 98.61382°, 1767 m, 5 November 2004, H.B. Liang collector [1 male; IOZ]); Qiqi He (27.75748°, 98.66073°, 1500 m, 30 September-1 October 2007, D.H. Kavanaugh, H.B. Liang & H.L. Shi collectors [1 female; CAS]); Qiqi Trail (at No. 12 Bridge Camp, 27.71502°, 98.50244°, 2775 m, 15–19 July 2000, D.H. Kavanaugh & H.B. Liang collectors [2 males and 1 female; CAS, IOZ]), (at Dongshaofang area, 27.69504°, 98.48433°, 3230–3680 m, 16–17 July 2000, D.H. Kavanaugh & H.B. Liang collectors [8 male and 10 females; CAS, IOZ]). Longyang County: Bawan-Tengchong Road Km 36–37 (24.93417°, 98.77944°, 2150 m, 12 October 2003, H.B.Liang & X.C. Shi collectors [1 female; IOZ]). Lushui County: Luzhang Township (Fengxue Yakou at Pianma Road, 25.97228°, 98.68336°, 3150 m, 15 October 2002, D.H. Kavanaugh P.E. Marek & H.B. Liang collectors [8 males and 10 females], 11 May 2005, D.H. Kavanaugh, H.B. Liang, C.E. Griswold, D.Z. Dong & G. Tang collectors [1 female; CAS]), (Yaojiaping He at Pianma Road, 25.97722°, 98.71091°, 2527 m, 19–20 May 2005, D.H. Kavanaugh, C.E. Griswold, H.B. Liang, D.Z. Dong & G. Tang collectors [23 males and 33 females; CAS, IOZ]); Pianma (9 km ESE on Pianma Road at Changya He, 25.99414°, 98.66336°, 2450 m, 14 May 2005, H.B. Liang collector [2 females; CAS, IOZ]), (9.3 km ESE on Pianma Road, 25.99363°, 98.66651°, 2460–2470m, 15–18 October 1998, D.H. Kavanaugh collector [6 males and 6 females; CAS, IOZ]), (Changya He, 25.99363°, 98.66651°, 2460–2470m, 13 October 1998, D.H. Kavanaugh collector [3 males and 6 females; CAS, IOZ]). Tengchong County: Bawan-Tengchong Road Km 42–46 (24.95361°, 98.74222°, 2290 m, 14 October 2003, H.B. Liang & X.C. Shi collectors [2 males and 1 female; CAS, IOZ].
This is the only species recorded from all seven Core Areas in the Gaoligong Shan region; but in the southern half of the study area, it is restricted to only the highest elevations, where members are found mainly on the passes over the crest of the mountain range and on the slopes just below the passes on both sides.
Fig. 27. This species has been recorded from Bhutan and China (Fujian, Sichuan and Yunnan Provinces and Xizang Autonomous Region).
http://species-id.net/wiki/Amara_dissimilis
Figs 5b–c, 5e, 6b, 17, 22a, 24b, 27–30Adults of this species (Fig. 17) can be distinguished from those of all other species in the region by the following combination of character states: body length 7.7–8.7 mm; dorsal surface dark brown to black, without or with only very faint metallic green reflection, at least femora and outer antennomeres dark (piceous to black); elytral microscuplture comprised of distinctly isodiametric meshes in both males and females (more deeply impressed in females than in males); pronotum (Fig. 17a) distinctly narrowed anteriorly, with anterior margin clearly narrower than the hind margin, lateral margins more or less evenly rounded from apical to basal angle, anterior angles slightly projected anteriorly beyond anterior margin, posterior angles obtusely angulate and slightly denticulate, basal impressions broadly and deeply foveate, outer basal impression distinctly delimited laterally by a broad convexity; elytra without evident sub-basal depressions, pore puncture absent, elytral striae distinct throughout and deeply impressed; medial protibial spurs simple; metatibiae of males with a brush-like patch of setae medially in the apical half; tarsomere 5 of hind tarsi with two or (in a few specimens) three pairs of setae ventrally (Fig. 6b); last abdominal sternite of male with one pair (Fig. 5e) and female with two pairs (Fig. 5c) of setiferous punctures near hind margin.

Amara (Bradytus) dissimilis Tschitschérine. a dorsal habitus (CASENT1033318) b–c median lobe of aedeagus of male (CASENT1002115) b left lateral aspect c dorsal aspect; scale lines = 1.0 mm d Map of localities records (red circles) for Amara dissimilis in the Gaoligong Shan region, scale line = 100 km.
Specimens of this species were collected in daytime from under stones and other cover in open roadside areas (Fig. 24b) and other waste areas (Fig. 22a) around human settlements with scattered grasses and shrubs, at the edges of meadows and agricultural fields, and on open rocky banks of streams, and in these same habitats at night, when beetles were found active on bare substrate. Members of this species were found at elevations ranging from 1500 to 3611 m (most abundantly between 1500 and 3150 m), and syntopic with adults of Amara birmana, Amara chalciope, Amara latithorax, Amara lucidissima, Amara pingshiangi, Amara sikkimensis and Amara silvestrii at one or more sites.
Fig. 17d. We examined a total of 14 specimens (7 males and 7 females) from the following localities: Fugong County: Lumadeng Township (7.5 km below Shibali on Yaping Road, 27.14627°, 98.81559°, 2030 m, 3 May 2004, H.B. Liang & M. Xie collectors [1 female; IOZ]). Gongshan County: Bingzhongluo Township (Gongdong, 27.99858°, 98.61933°, 2506 m, P.E. Marek collector [1 female; CAS]); Cikai Township (27.74939°, 98.66453°, 1515 m, 25 September 2002, H.B. Liang & W.D. Ba collectors [1 male and 1 female; IOZ]), (Pula He just above Nu Jiang Road, 27.74861°, 98.66675°, 1440m, 23 October 2004, D.H. Kavanaugh & H.B. Liang collectors [1 female; IOZ]); Dulongjiang Township (0.6 km N of Dizhengdang village on Dulong Jiang, 28.08442°, 98.32652°, 1880m, 29 October 2004, D.H. Kavanaugh, D.Z. Dong & G. Tang collectors [2 males; CAS, IOZ]), Dulongjiang Township (S of Dizhengdang village at Sialong He, 28.07654°, 98.32603°, 1890 m, 30 October 2004, D.H. Kavanaugh, D.Z. Dong & G. Tang collectors [1 female; CAS]), (2.3 to 3.3 airkm S of Longyuan village on Dulong Jiang, 28.00532°, 98.32145°, 1685–1720 m, 2 November 2004, D.H. Kavanaugh, D.Z. Dong & G. Tang collectors [2 females; CAS, IOZ]); Heiwadi (16.8 km W of Cikai on Dulong Valley Road, 27.79584°, 98.58443°, 2150 m, 4 October 2002, H.B. Liang, W.D. Ba & C.G. Jin collectors[1 male; IOZ]; 10 October 2004, D.H. Kavanaugh, P.E. Marek & H.B. Liang collectors [1 male; CAS]). Longyang County: Nankang Yakou (24.83167°, 98.76667°, 2130 m, 4 November 1998, D.H. Kavanaugh collector [1 males; CAS]). Lushui County: Luzhang Township (Fengxue Yakou at Pianma Road, 25.97228°, 98.68336°, 3150 m, 11 October 1998, D.H. Kavanaugh collector [1 male; CAS]).
This species was recorded from all but one Core Area (Core Area 5) in the Gaoligong Shan region. In the southern half of the study area, it is restricted to only the highest elevations, where members are found mainly on the passes over the crest of the mountain range.
Fig. 27. This species has been recorded from Gansu, Qinghai, Shaanxi, Sichuan and Yunnan Provinces and Xizang Autonomous Region in China. Its occurrence in the study area represents the southern limit of its known geographical range.
http://species-id.net/wiki/Amara_elegantula
Figs 5b–c, 5e, 6b, 18, 26a, 26b, 27–30Adults of this species (Fig. 18) can be distinguished from those of all other species in the region by the following combination of character states: body length 7.0–7.5 mm; dorsal surface dark with distinct metallic blue-green reflection in most specimens, non-metallic black in a few specimens, at least femora and outer antennomeres dark (piceous to black); elytral microsculpture effaced or nearly so in both males and females; pronotum (Fig. 18a) with lateral margins straight or faintly to distinctly sinuate just anterior to basal angles, rounded near middle, less rounded or nearly straight also in anterior one-third in most specimens, anterior angles distinctly and narrowly projected anteriorly beyond anterior margin, lateral explanation narrow throughout, outer basal impressions foveate and distinct from lateral groove, punctation of base not extended anteriorly along sides beyond basal one-third; elytra with parascutellar pore puncture absent; medial protibial spurs simple; tarsomere 5 of hind tarsi with two or (in a few specimens) three pairs of setae ventrally (Fig. 6b); last abdominal sternite of male with one pair (Fig. 5e) and female with two pairs (Fig. 5c) of setiferous punctures near hind margin; male aedeagus with apical third of median lobe about as broad as middle third, apical hook of right paramere smaller and closer to apex (Fig. 8a).

Amara (Bradytus) elegantula Tschitschérine. a dorsal habitus (CASENT1026671) b–c median lobe of aedeagus of male (CASENT1026255) b left lateral aspect c dorsal aspect; scale lines = 1.0 mm d Map of localities records (red circles) for Amara elegantula in the Gaoligong Shan region, scale line = 100 km.
Specimens of this species were collected in daytime from under stones and other cover in open roadside areas near the crest of the mountain range (Fig. 26a), on the rocky heath tundra in areas with scattered vegetation, and on stabilized scree slopes between persistent snow patches in glacial cirques (Fig. 26b). At night, specimens were found in these same areas active on the bare substrate. Specimens were found at elevations ranging from 3350 to 4000 m, the highest sites sampled. The only other Amara species members of which were found syntopic with those of Amara elegantula was Amara chalciope, with adults of both species found together at only one site at an elevation of 3365 m (Fig. 26a), near the lower altitudinal limit of the former and the upper altitudinal limit of the latter.
Fig. 18d. We examined a total of 60 specimens (30 males and 30 females) from the following localities: Gongshan County: Heipu Yakou (northwest slope along road W of tunnel, 27.77447°, 98.44793°, 3350 m, 13 August 2006, D.H. Kavanaugh & J.A. Miller collectors [2 females; CAS, IOZ]), (southeast slope, 27.77032°, 98.44674°, 3365 m, 11–13 August 2006, D.H. Kavanaugh, J.A. Miller, D.Z. Dong & Y. Liu collectors [15 males and 9 females; CAS, IOZ]); southwest slope of Kawakarpu Shan (on slope NE of Chukuai Lake, 27.98206°, 98.48027°, 3950 m, 20 August 2006, Y. Liu, P. Hu, D.Z. Dong & J. Wang collectors [4 males and 3 females; CAS, IOZ]), (0.9 km N of Chukuai Lake, 27.98981°, 98.47392°, 4000 m, 21 August 2006, D.H. Kavanaugh, J. Xiong & C.H. Li collectors [2 females; CAS, IOZ]), (0.3 km NNE of Chukuai Lake, 27.98393°, 98.47491°, 3745 m, 19 August 2006, D.H. Kavanaugh, J.A. Miller, D.Z. Dong, J. Xiong & C.H. Li collectors [7 males and 8 females; CAS, IOZ, ZMHB]), (0.4 km NW of Chukuai Lake, 27.98231°, 98.47069°, 3808 m, 21 October 2006. D.Z. Dong collector [2 males; CAS, IOZ]), (0.75 km NW of Chukuai Lake, 27.98631°, 98.47069°, 3820 m, 21 August 2006, Y. Liu, P. Hu & J. Wang collectors [1 male and 2 females; CAS, IOZ]), (0.3 km SW of Chukuai Lake, 27.97686°, 98.47799°, 3750 m, 19 August 2006, Y. Liu collector [2 males and 3 females; CAS, IOZ]).
This species was recorded only from the northern part of the study area (Core Areas 1 and 2), where it is restricted to the highest elevations sampled, along the crest of the mountain range and both east and west slopes just below the crest.
Fig. 27. This species has been recorded from Bhutan, China (Yunnan Province and Xizang Autonomous Region), India (Sikkim and West Bengal) and eastern Nepal. Its occurrence in the study area represents the southern and eastern limits of its known geographical range.
http://species-id.net/wiki/Amara_simplicidens
Figs 5b–c, 5e, 6b, 19, 27–30Adults of this species (Fig. 19) can be distinguished from those of all other species in the region by the following combination of character states: body length 8.3–8.4 mm; dorsal surface dark brown to black, without metallic reflection, at least femora and outer antennomeres dark (piceous to black); elytral microscuplture comprised of distinctly transverse meshes in both males and females (more transverse and less deeply impressed in males than in females); pronotum (Fig. 19a) only slightly narrower anteriorly than basally, anterior margin almost as wide as posterior margin, lateral margins more or less evenly rounded from apical to basal angle, anterior angles slightly projected anteriorly beyond anterior margin, posterior angles sharp, not rounded, outer basal impressions sharply delimited laterally by narrow, slightly oblique raised (but not carinate) areas, pronotal base very coarsely punctate; elytra with parascutellar pore puncture absent; medial protibial spurs simple; metatibia of male with brush-like setae medially in the apical half; tarsomere 5 of hind tarsi with two or (in a few specimens) three pairs of setae ventrally (Fig. 6b); last abdominal sternite of male with one pair (Fig. 5e) and female with two pairs (Fig. 5c) of setiferous punctures near hind margin.
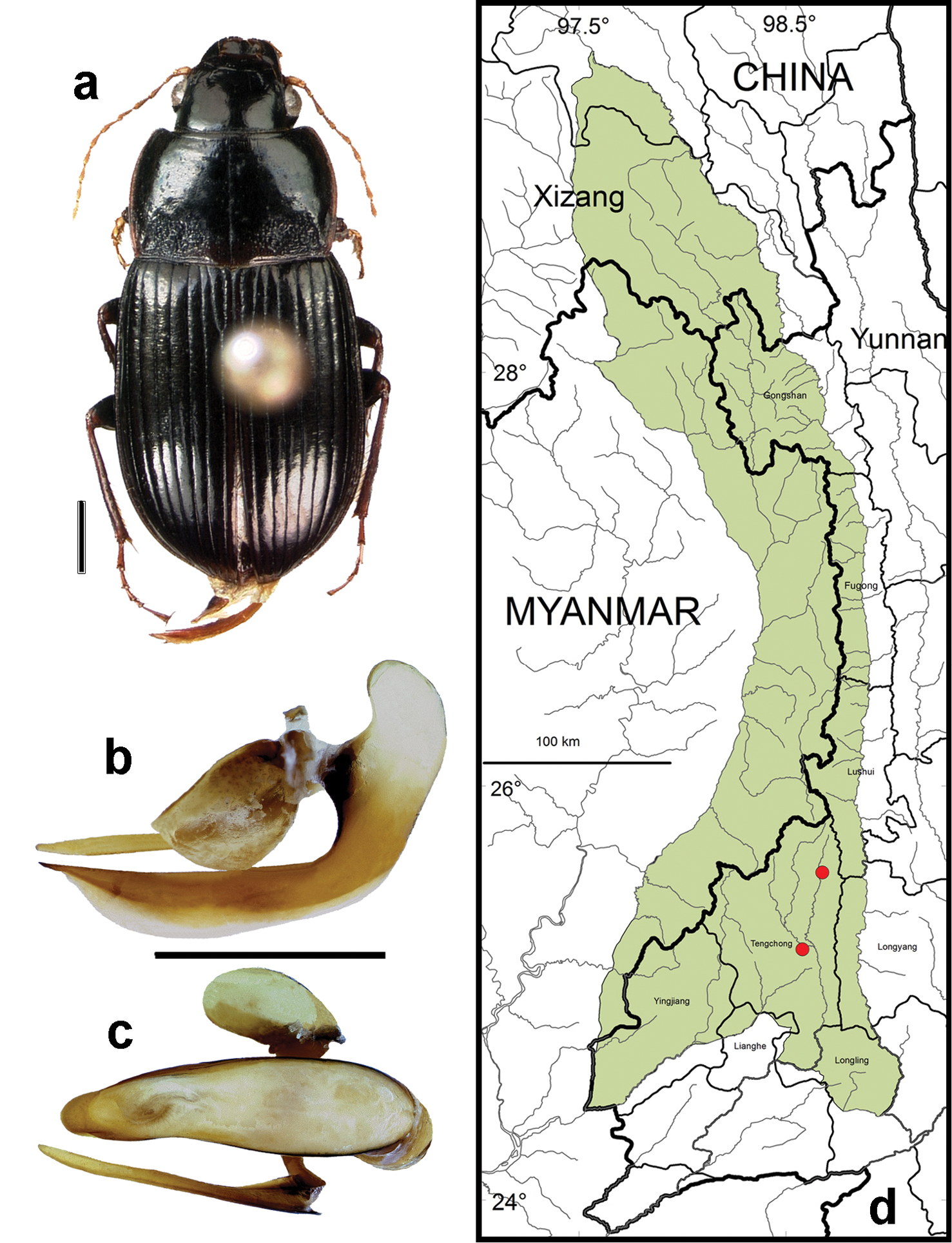
Amara (Bradytus) simplicidens Morawitz a dorsal habitus (CASENT1013861) b–c median lobe of aedeagus of male (CASENT1013861) b left lateral aspect c dorsal aspect; scale lines = 1.0 mm d Map of localities records (red circles) for Amara simplicidens in the Gaoligong Shan region, scale line = 100 km.
Specimens of this species were collected in daytime from under stones on the open bank of a small stream at an elevation of 1740 m and at night on wet stones along a roadside at 1515 m elevation. The only other Amara species members of which were found syntopic with those of Amara simplicidens was Amara lucidissima, with adults of both species found together at the 1740 m site.
Fig. 19d. We examined a total of 2 specimens (1 male and 1 female) from the following localities: Tengchong County: Datang Village (Maluchong, 25.58194°, 98.67583°, 1740 m, 24 October 2003, H.B. Liang collector [1 male; CAS]); Longchuan Jiang (at Xiangyang Bridge, 25.21056°, 98.58028°, 1515 m, 23 October 2003, H.B. Liang & X.C. Shi collectors [1 female; IOZ]).
This species was recorded only from the western slope of the southern part of the study area (Core Area 6).
Fig. 27. This species has been recorded from China (Fujian, Heilongjiang, Henan, Hubei, Jiangsu, Jiangxi, Sichuan, Yunnan, and Zhejian Provinces), Japan, North Korea, and Russia (Khabarovsk and Primorsky Kraya, Sakhalinskaya Oblast, and the Kuril Islands). Its occurrence in the study area represents the western limit of its known geographical range.
http://species-id.net/wiki/Amara_pingshiangi
Figs 5b–c, 5e, 6a, 20, 25b, 27–30Adults of this species (Fig. 20) can be distinguished from those of all other species in the region by the following combination of character states: body length 11–13 mm, body form stout; dorsal surface dark piceous, at least femora and outer antennomeres dark (piceous to black); pronotum with lateral margin markedly rounded, pronotal base punctate, lateral areas in basal half and near anterior margin also punctuate in most specimens; elytra with parascutellar pore puncture absent; medial protibial spurs simple; tarsomere 5 of hind tarsi with five or six pairs of setae ventrally (Fig. 6a); last abdominal sternite of male with one pair (Fig. 5e) and female with two pairs (Fig. 5c) of setiferous punctures near hind margin.
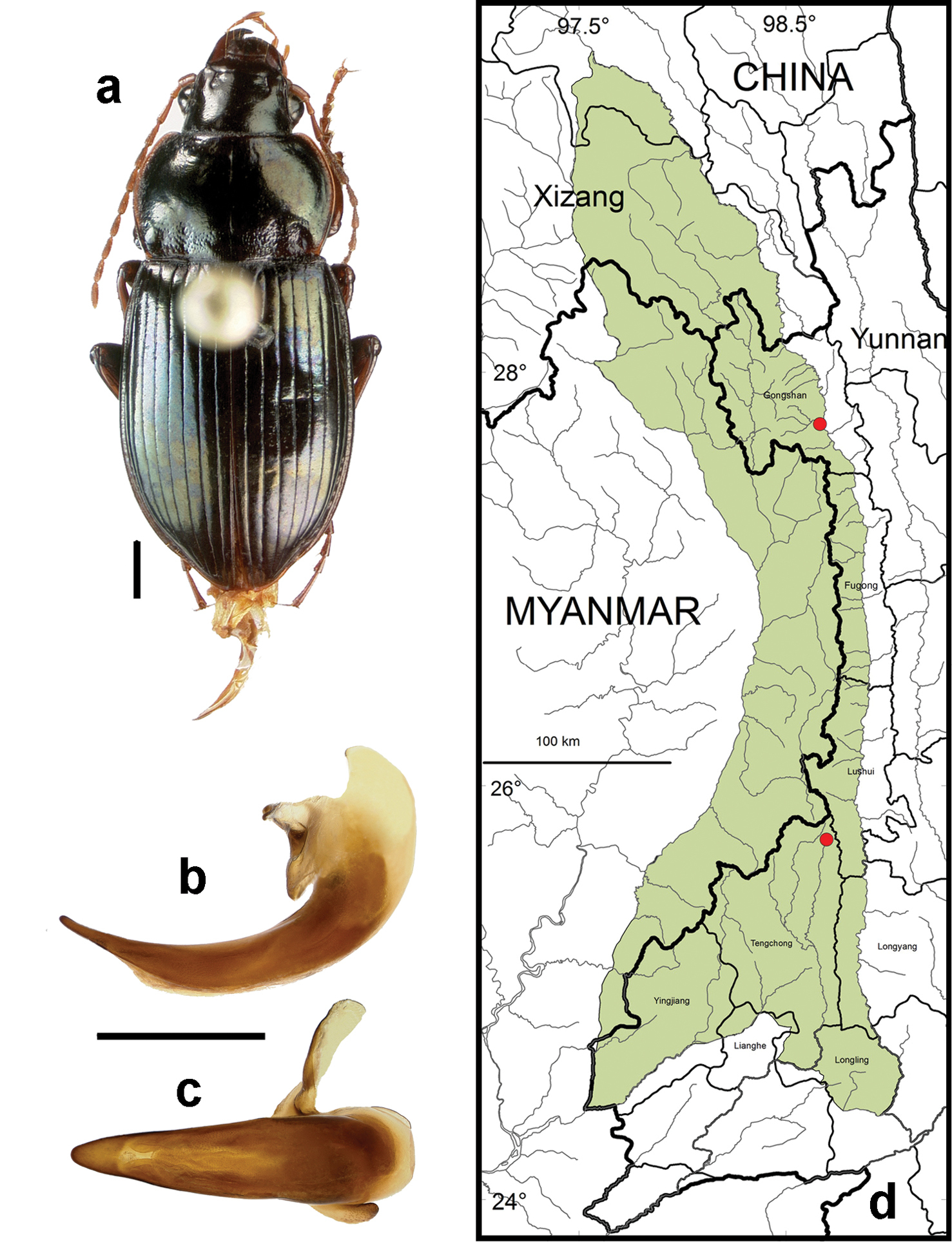
Amara (Bradytus) pingshiangi Jedlička. a dorsal habitus (CASENT1038324) b–c median lobe of aedeagus of male (CASENT1038324) b left lateral aspect c dorsal aspect; scale lines = 1.0 mm d Map of localities records (red circles) for Amara pingshiangi in the Gaoligong Shan region, scale line = 100 km.
Specimens of this species were collected only at night, active on the ground in marshy meadow margin areas (Fig. 25b) near forest edges, at elevations ranging from 1515 to 2010 m, and syntopic with adults of Amara birmana, Amara dissimilis and Amara shaanxiensis at one or more sites.
Fig. 20d. We examined a total of 3 specimens (2 males and 1 female) from the following localities: Gongshan County: Cikai Township (27.74939°, 98.66453°, 1515 m, 25 Sep. 2002, H.B. Liang & W.D. Ba collectors [1 female; IOZ]). Tengchong County: Longtang He (at Dahetou Lingganjiao, 25.73947°, 98.6963°, 2010 m, 16 May 2006, D.H. Kavanaugh collector [1 male; CAS], 18 May 2006, D.H. Kavanaugh & R.L. Brett collectors [1 male; CAS]).
Members of this species were collected at only two sites, one in the northeastern part (Core Area 2) and other in the southwestern part (Core Area 6) of the study area, but not in intervening areas. This gap in distribution is most likely an artifact of inadequate sampling and not a real disjunction.
Fig. 27. This species has been recorded from Fujian, Jiangxi, Sichuan, Yunnan and Zhejiang Provinces in China. Its occurrence in the study area represents the western limit of its known geographical range.
http://species-id.net/wiki/Amara_lucidissima
Figs 5b–c, 5e, 6b, 8a, 21, 22a, 27–30Adults of this species (Fig. 21) can be distinguished from those of all other species in the region by the following combination of character states: body length 6.5–8.0 mm; dorsal surface light-brown to brownish black, without metallic reflection, entire legs and antennae pale; elytra with parascutellar pore puncture absent; medial protibial spurs simple; tarsomere 5 of hind tarsi with two or (in a few specimens) three pairs of setae ventrally (Fig. 6b); last abdominal sternite of male with one pair (Fig. 5e) and female with two pairs (Fig. 5c) of setiferous punctures near hind margin; male aedeagus with apical third of median lobe broader than middle third, apical hook of right paramere large and slightly subapical (Fig. 8a).
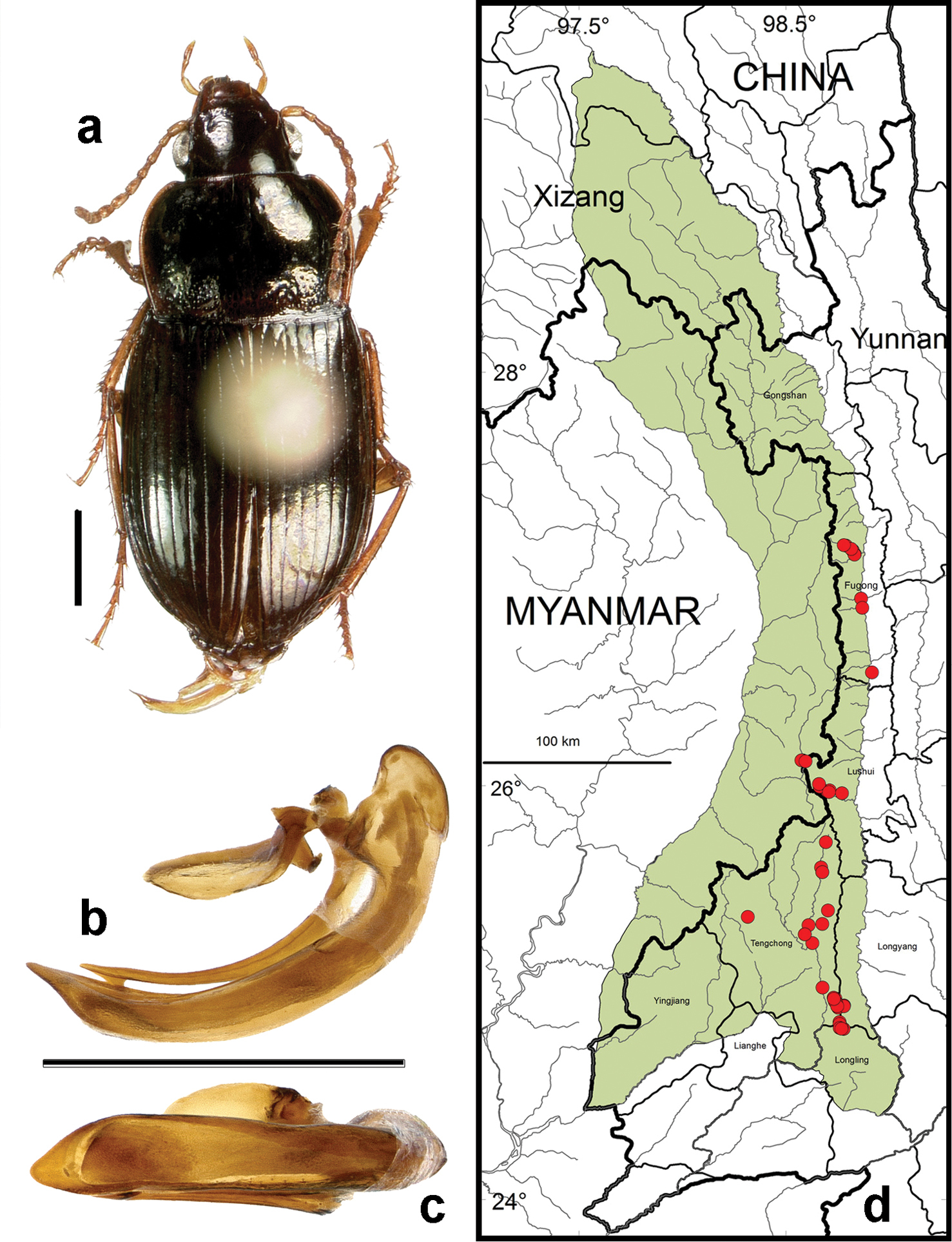
Amara (Reductocelia) lucidissima Baliani. a dorsal habitus (CASENT1011863) b–c median lobe of aedeagus of male (CASENT1002221) b left lateral aspect c dorsal aspect; scale lines = 1.0 mm d Map of localities records (red circles) for Amara lucidissima in the Gaoligong Shan region, scale line = 100 km.
Specimens of this species were collected, often in great abundance, in daytime from under stones and other cover in open roadside areas and waste areas (Fig. 22a) around human settlements with scattered grasses and shrubs, at the edges of agricultural fields and on open banks of streams, and in these same habitats at night, when beetles were found active on bare substrate. Members of this species were found at elevations ranging from 1185 to 3140 m and syntopic with adults of Amara birmana, Amara chalciope, Amara dissimilis, Amara latithorax, Amara shaanxiensis, Amara sikkimensis, Amara simplicidens and Amara silvestrii at one or more sites.

Photographs of habitats for Amara species in the Gaoligong Shan region. a. Baoshan-Tengchong Road at Nankang Yakou, elevation 2130 m; habitat in which specimens of Amara birmana, Amara davidi, Amara dissimilis, Amara lucidissima and Amara shaanxiensis were collected. b. Gongshan County, Cikai Township, Heiwadi Village area, elevation 1800 m; habitat in which specimens of Amara congrua and Amara latithorax were collected.
Fig. 21d. We examined a total of 581 specimens (276 males and 305 females) from the following localities: Fugong County: Mugujia Village (Guquan He, 26.86177°, 98.86900°, 1175 m, 21 April 2004, D.H. Kavanaugh collector [1 female; CAS]); Shibali Road (above Shilajia village, 27.13947°, 98.82184°, 1900 m, 24 April 2004, D.H. Kavanaugh collector [2 females; CAS]), (above Shilajia Village at North Fork Yamu He, 27.13947°, 98.82184°, 1800–1900 m, 25 April 2004, D.H. Kavanaugh & C.E. Griswold collectors [18 males and 8 females; CAS, IOZ]); South Fork Yamu He (above Shilajia Village, 27.12101°, 98.83173°, 1630–1790 m, D.H. Kavanaugh collector [1 male; CAS]). Longyang County: Bawan-Tengchong Road Km 36 (24.9375°, 98.78028°, 2075 m, 11 October 2003, H.B.Liang collector [2 males and 2 females; CAS, IOZ]); Bawan-Tengchong Road Km 48–51 near Dahaoping Forest Station (24.97556°, 98.73000°, 2014 m, 18 October 2003, H.B. Liang collector [1 male, 3 females; CAS, IOZ]); Bawan-Tengchong Road Km 40–41 (24.92694°, 98.75278°, 2404 m, 12 October 2003, H.B.Liang collector [7 males and 3 females; CAS, IOZ]); Bawan-Tengchong Road Km 51 near Dahaoping Forest Station (24.9725°, 98.73889°, 2170 m, 18 October 2003, H.B. Liang & X.C. Shi collectors [1 female; IOZ]); Nankang Forest Station (24.82444°, 98.77889°, 2085 m, 27 October 2003, H.B. Liang & X.C. Shi collectors [12 males, 6 females; CAS, IOZ]); Nankang Yakou (24.83167°, 98.76667°, 2130 m, 4–7 November 1998, D.H. Kavanaugh collector [23 males and 44 females; CAS, IOZ]), (at Baoshan-Tengchong Road Km 24, 24.82583°, 98.77222°, 2130 m, 26 October 2003m, H.B. Liang & X.C. Shi collectors [11 males and 10 females; CAS, IOZ]), (mountain NW of Nankang Yakou, 24.83250°, 98.76944°, 2245 m, 27 October 2003, H.B. Liang & X.C. Shi collectors [1 male and 1 female; CAS, IOZ]). Lushui County: Gangfang Sancha Lukou (26.12167°, 98.575°, 1550 m, 14–15 October 1998, D.H. Kavanaugh collector [3 males and 5 females; CAS, IOZ]). Tengchong County: Bawan-Tengchong Road Km 42–46 (24.95361°, 98.74222°, 2290 m, 14 October 2003, H.B. Liang & X.C. Shi collectors [2 males and 1 female; IOZ], 17 October 2003, H.B. Liang collector [1 male and 2 females; IOZ]); Bawan-Tengchong Road Km 46–51 (24.95722°, 98.73667°, 2220 m, 17 October 2003, D.Z. Dong, H.B. Liang & X.C. Shi collectors [16 males and 25 females; CAS, IOZ]); Bawan-Tengchong Road Km 65 at Longwenqiao (25.02396°, 98.67675°, 1285 m, 19 October 2003, D.Z. Dong collector [4 males and 1 female; CAS, IOZ]); Baoshan-Tengchong Road Km 24 (24.82889°, 98.76028°, 2008 m, 29 October 2003, N.D. Penny & T.S. Briggs collectors [5 males and 6 females; CAS, IOZ]); 5–8 km E of Dahaoping (24.93417°, 98.7475°, 2358 m, 18 October 2003, N.D. Penny, T.S. Briggs, & D.Z. Dong collectors [1 male; CAS]); Datang Village (Maluchong, 25.58194°, 98.67583°, 1740 m, 24 October 2003, H.B. Liang collector [1 male; IOZ]), (Danlonghe Bridge, 25.60500°, 98.67028°, 1768 m, 24October 2003, H.B. Liang & X.C. Shi collectors [3 males and 3 females; CAS, IOZ]); Longchuan Jiang (at Longkou Village, 25.28167°, 98.59167°, 1500 m, 2 November 1998, D.H. Kavanaugh & C.E. Griswold collectors [4 males and 11 females; CAS, IOZ]), (at Xiaojiangqiao (25.23944°, 98.62722°, 1445 m, 21 October 2003, H.B. Liang & X.C. Shi collectors [1 male and 2 females; CAS, IOZ]), (at Yonganqiao (25.32556°, 98.60944°, 1500 m, 22 October 2003, H.B. Liang & X.C. Shi collectors [10 male and 10 females; CAS, IOZ]), (at Yonganqiao (25.60500°, 98.67583°, 1500 m, 22 October 2003, H.B. Liang & X.C. Shi collectors [2 females; IOZ]); Longkou Village, 25.28167°, 98.59167°, 1500 m, 22 October 2003, H.B. Liang & X.C. Shi collectors [15 males and 26 females; CAS, IOZ]); Shaba Village (25.39639°, 98.70306°, 1850 m, 23 October 2003, H.B. Liang & X.C. Shi collectors [6 males and 7 female; CAS, IOZ]); Xiaodifang Village (24.85722°, 98.75917°, 2150 m, 29 October 2003, D.Z. Dong collector [2 males and 1 female; CAS, IOZ]); Xiaoheishan Forest Station (24.82889°, 98.76000°, 2025 m, 29 October 2003, H.B. Liang collector [32 males and 41 females; CAS, IOZ]); Zhoujia-po Village, 25.33222°, 98.67611°, 1740 m, 24 October 2003, D.Z. Dong collector [2 males; CAS]); near Zhoujia-po village (25.58194°, 98.67611°, 1725 m, 24 October 2003, N.D. Penny & T.S. Briggs collectors [2 females; CAS, IOZ].
This species was recorded from the southern two-thirds of the study area (Core Areas 2, 4, 5, 6 and 7), on both eastern and western slopes of the mountain range.
Fig. 27. This species has been recorded from China (Fujian, Hubei, Sichuan, Yunnan, and Zhejiang Provinces) and Taiwan. Its occurrence in the study area represents the western limit of its known geographical range.

Photographs of habitats for Amara species in the Gaoligong Shan region. a Lushui County, Yaojiaping just off Pianma Road, elevation 2600 m; habitat in which specimens of Amara birmana, Amara sikkimensis and Amara silvestrii were collected b Gongshan County, Dulong Valley, 2.8 km S of Longyuan Village, elevation 1660 m; habitat in which specimens of Amara latithorax and Amara sikkimensis were collected.

Photographs of habitats for Amara species in the Gaoligong Shan region. a Gongshan County, Dongshaofang area just below pass into Dulong Valley, elevation 3500 m; habitat in which specimens of Amara chalciope were collected b Lushui County, Pianma Road just W of Fengxue Yakou, elevation 3150 m; habitat in which specimens of Amara dissimilis and Amara chalciope were collected.

Photographs of habitats for Amara species in the Gaoligong Shan region. a Lushui County, Pianma Road just E of Fengxue Yakou, elevation 3150m; habitat in which specimens of Amara birmana and Amara chalciope were collected b Tengchong County, Longtang He at Dahetou Lingganjiao, elevation 2010 m; habitat in which specimens of Amara birmana, Amara pingshiangi and Amara shaanxiensis were collected.

Photographs of habitats for Amara species in the Gaoligong Shan region. a Gongshan County, Heipu Yakou just E of Tunnel, elevation 3365 m; habitat in which specimens of Amara chalciope and Amara elegantula were collected b Gongshan County, southwest slope of Kawakarpu Shan on slope abiove and NE of Chukuai Lake, elevation 3950 m; habitat in which specimens of Amara elegantula were collected.
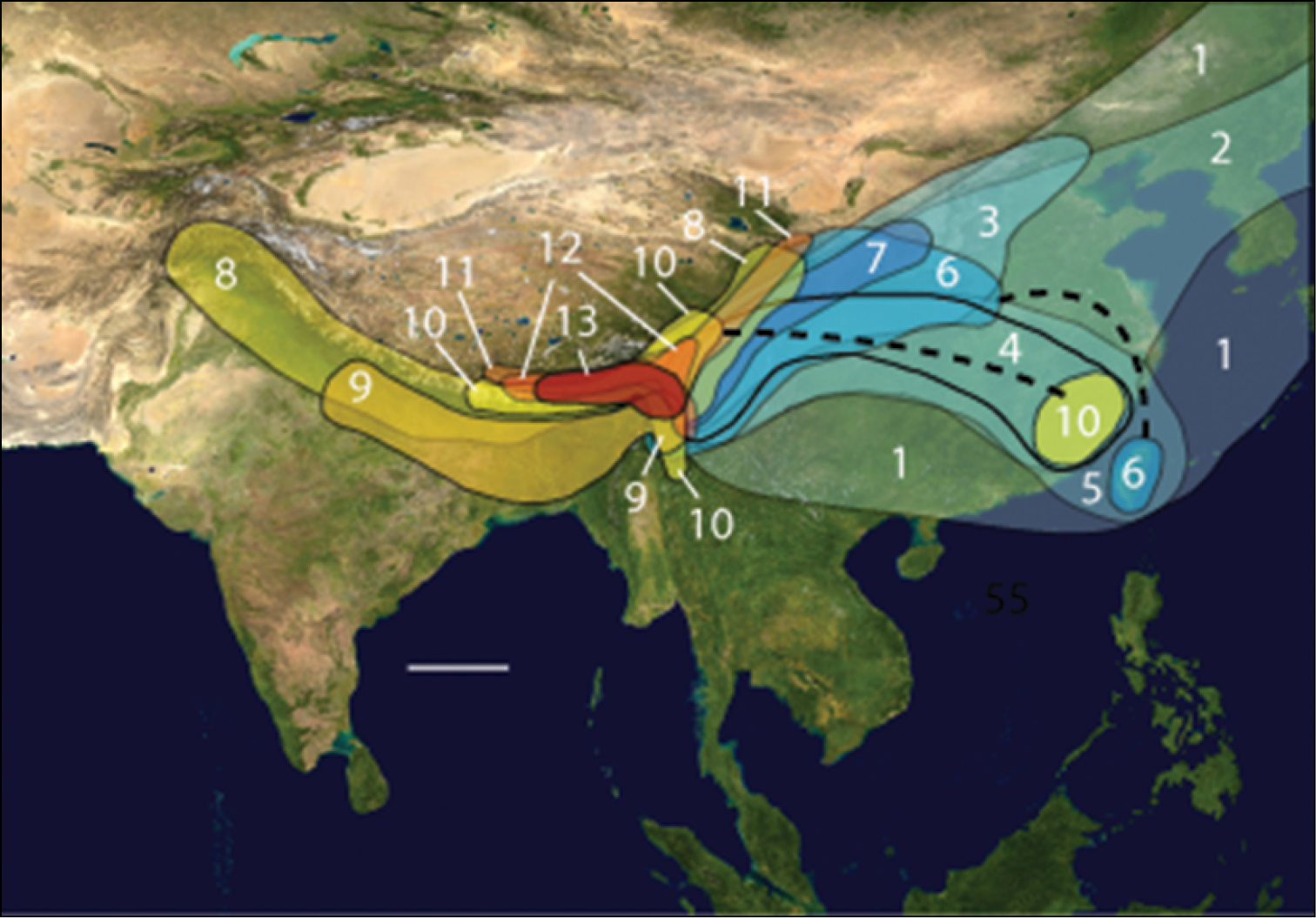
Map showing approximate overall geographical distributions of species occurring in the Gaoligong Shan region (ranges that extend to the upper right corner of the map are truncated there but continue northeast to Japan and the Russian Far East); dashed lines connect apparently significant disjunct areas of the range of a species; scale line = 500 km. 1 Amara (Amara) congrua Morawitz 2 Amara (Bradytus) simplicidens Morawitz 3 Amara (Zezea) davidi Tschitschérine 4 Amara (Bradytus) pingshiangi Jedlička 5 Amara (Reductocelia) lucidissima Baliani 6 Amara (Amara) silvestrii Baliani 7 Amara (Amara) shaanxiensis Hieke; 8 Amara (Xenocelia) sikkimensis Andrewes 9 Amara (Harpaloamara) latithorax Baliani 10 Amara (Bradytus) chalciope (Bates) 11 Amara (Bradytus) dissimilis Tschitschérine 12 Amara (Pseudoamara) birmana Baliani; 13 Amara (Bradytus) elegantula Tschitschérine.

Chart showing the representation of Amara species in project-designated Core Areas (see Fig. 3) in the Gaoligong Shan region.

Chart illustrating the altitudinal ranges of Amara species represented in the Gaoligong Shan region.

Chart illustrating the co-occurence (syntopy) of Amara species in samples from the same habitats and at the same sites in the Gaoligong Shan region.
Although the Gaoligong Shan region is at the heart of one of the world’s biodiversity hotspots (
Broad geographical distribution patterns. The overall geographical ranges of the 13 species, superimposed on one another, are graphically approximated in Fig. 27. Among the geographical ranges of these species, three general range patterns are apparent. The first pattern includes seven species (Amara congrua, Amara davidi, Amara lucidissima, Amara pingshiangi, Amara shaanxiensis, Amara sikkimensis and Amara simplicidens), more or less broadly distributed in China and eastern Asia, but all of which have their western range limits at or near the Gaoligong Shan region. Two of these, Amara congrua and Amara simplicidens, have ranges that extend east and north to Japan and the Russian Far East, as well as across most of China east of the study area. Strangely, members of these two species were among the least frequently encountered Amara in the area during our study. The ranges of four species with eastern distributional ranges, Amara congrua, Amara simplicidens, Amara pingshiangi and Amara lucidissima, include Fujian Province in China. There are gaps in the known distributions of all of these species between the study area and Fujian, and inadequate sampling is likely the cause of these apparent gaps. For three species, Amara congrua, Amara lucidissima and Amara silvestrii, known geographical ranges also include Taiwan. Both Amara congrua and Amara lucidissima are also known from Fujian, right across the South China Sea from Taiwan, a disjunction of only about 250 to 400 km, depending on localities compared. The nearest Taiwanese localities for Amara silvestrii are more than 1400 km distant from the nearest mainland sites, in Shaanxi Province, and this may represent a real and significant range disjunction. The remaining species included among the class of eastern patterns, Amara shaanxiensis, has the most restricted known geographical distribution, recorded only from Yunnan and from Shaanxi Province, but it likely also occurs in intervening areas in along the southern edge of the Qinghai-Xizang Plateau in Sichuan and Gansu Provinces.
The second general geographical range pattern includes two species, Amara elegantula and Amara latithorax, with geographical ranges that include only the Himalayan region immediately to the west and the Gaoligong Shan region, where they reach their eastern distributional limit. Amara elegantula ranges as far west as eastern Nepal and adjacent parts of Xizang Autonomous Region (Tibet), and the range of Amara latithorax also extends west to Nepal, but apparently only on the southern slope of the Himalaya (i.e., it has not been recorded from the northern side of that range). The third general range pattern includes the remaining four species, Amara birmana, Amara chalciope, Amara dissimilis and Amara sikkimensis. The geographical distributions of each of these species include the Gaoligong Shan region and areas of varied distance to both the northwest and the northeast along the Himalayan range and the southern edge of the Qinghai-Xizang Plateau, respectively. Of these, Amara sikkimensis has the broadest distribution, extended from northen Pakistan in the west to Gansu Province in the east. The known range of Amara chalciope extends from Bhutan and Xizang Autonomous Region in the west to Sichuan in the east, with a significantly disjunct occurrence also in Fujian Province in southeastern China. Populations currently included under this name actually may represent a complex of species, a solid understanding of which will require additional study. The known range of Amara dissimilis extends from Xizhang in the northwest to Shaanxi in the northeast and actually includes parts of the Qinghai-Xizang Plateau itself in southern Qinghai Province. Finally, Amara birmana has the most restricted range of the species with this pattern, known only from northeastern India and northern Myanmar in the west to Sichuan in the east.
On first glance, the observed diversity of Amara species in the study area appears to be due simply to an overlap of faunal elements from both the east and the west. Certainly several of the species with broad distributions in large parts of eastern Asia (e.g., Amara congrua and Amara simplicidens) likely have their origins in regions to the northeast and so their occurrence in the study area is likely the results of range expansion into the region. However, even a casual look at the majority of range patterns illustrated in Fig. 27 tempts us to suggest that the Gaoligong Shan region, as a core part of the Hengduan Mountains System, may have served as an area of origin for at least part of the Amara fauna which now also occupies the Himalayan Ranges and southern edge of the Qinghai-Xizang Plateau to the west and east, respectively. The Hengduan Mountains date their origins to the late Mesozoic, whereas the uplift of the Himalayan Ranges and Qinghai-Xizang Plateau began later, in the early Cenozoic (
Regional geographical and altitudinal distribution patterns. Within the Gaoligong Shan study area, most of the species represented are broadly distributed, both geographically and altitudinally. This is not surprising given the shared preferences of their members for open and disturbed habitats. Such areas are abundant in the lowlands, most often associated with agriculture and human habitation. They are also abundant at all elevations within in the Gaoligong Shan itself, whether in naturally open alpine areas above tree line or in open areas along streams, on flood outwash flats or stabilized landslides, or in open sites created by humans, such as along road cuts, in forest clearcuts and previously burned areas and around all forms of human settlement and activity. What is perhaps unexpected is that a few of the species are quite restricted in their geographical and altitudinal ranges within the study area.
The chart in Fig. 28 summarizes the recorded regional distributions of the species with respect to our project-designated Core Areas; and the recorded altitudinal ranges for each species are shown in Fig. 29. Only Amara chalciope is recorded from all seven Core Areas, and this species also shows the broadest altitudinal range (from 1500 to just over 3600 m). Amara dissimilis is recorded from six of the Core Areas (not yet recorded from Core Area 5) and also has a broad altitudinal range (from 1515 to 3150 m). Two species, Amara silvestrii and Amara sikkimensis, are recorded from five of the Core Areas, including both the northernmost and southernmost, so they are likely to be found in the other Core Areas with additional sampling. Both species also occupy broad altitudinal ranges (from 1515 to 3000 m and from 1175 to 2800 m, respectively). Four species have been recorded from fewer Core Areas but nonetheless are likely to be found more widely in the region with additional sampling in gap areas in their ranges. Amara davidi has been recorded from three Core Areas (1, 6 and 7), Amara shaanxiensis from four (2, 5, 6 and 7), Amara pingshiangi from two (2 and 6) and Amara congrua from four (2, 3, 5 and 6). Among these, Amara shaanxiensis has the broadest recorded altitudinal range (from just over 1800 to 3000 m) and Amara davidi and Amara pingshiangi the narrowest (from 2020 to 2440 m and from 1515 to 2010 m, respectively). Amara congrua has a moderately broad recorded altitudinal range (from 892 to 2000 m), including the lowest elevation at which any Amara species has been recorded in the study area. The five species not yet mentioned all have geographical and, in some cases, also altitudinal ranges that are restricted to at least some extent. Among these, Amara lucidissima is the most widespread, both geographically (Core Areas 3–7) and altitudinally (from 1185 to 3140 m); and because it has been locally abundant wherever it has been found, the absence of records from the northern Core Areas (1 and 2) is significant. Similarly, Amara birmana has been recorded only from the southern half of the study area (Core Areas 4-7), at elevations ranging from 1590 to 3150 m, and this is likely a true measure of its actual distribution in the region. Two species, Amara elegantula and Amara latithorax, have been recorded only from the northern part of the study area (in Core Areas 1 and 2 and 1-3, respectively). However, these two differ markedly in their altitudinal ranges, with Amara latithorax recorded at elevations ranging from about 1200 to 2000 m and Amara elegantula found only at elevations ranging from 3350 to at least 4000 m. For Amara elegantula, a geographical range restricted to the north is expected, given its restricted high-elevation altitudinal range and the physiography of the study area, with virtually all contiguous areas above 3300 m found in the north. However, we find the restricted northern distribution of Amara latithorax surpising, given that its altitudinal range requirements seemingly could be met virtually anywhere along the length of the Gaoligong Shan. The remaining species, Amara simplicidens, has been recorded only from Core Area 6 in the southwestern part of the study area and only at relatively low elevations (from 1515 to 1740 m). This is somewhat surprising given that the overall geographical range of this species is so large, extended eastward all the way to Japan and the Russian Far East.
A comparison of recorded diversity for Amara species among the seven Core Areas (Fig. 28) shows that all areas are occupied by at least five species, with highest diversity in Core Area 6 (with 11 of the 13 species), second highest in Core Area 2 (with 9 species) and lowest recorded diversity in Core Area 4 (which is also by far the smallest Core Area). Core Areas 1 and 2 uniquely share one species, Amara elegantula, and Core Areas 1-3 together are uniquely occupied two species (Amara elegantula and Amara latithorax). Core areas 3-7 together are uniquely occupied by three species, Amara birmana, Amara lucidissima and Amara simplicidens; and Core Areas 4-7 together uniquely include two of these, Amara birmana and Amara simplicidens. Only Core Area 6 has a species, namely Amara simplicidens, not shared with any other Core Area. Consequently, but perhaps not surprisingly, the northern and southern extremes of the study area (Core Areas 1 and 2 in the north and 4-7 in the south) have the most distinctive Amara faunas, with Core Area 3 apparently representing a region of faunal transition.
With respect to the altitudinal distribution of the Amara fauna of the study area (Fig. 29), several points can be made. Highest species diversity is concentrated at about 2000 m in elevation, with 11 of the 13 species recorded from that elevation. Only Amara elegantula and Amara simplicidens do not occur at that elevation, the former only far above that level, the latter only below it. Three species, Amara congrua, Amara latithorax and Amara pingshiangi reach their upper altitudinal limit at about this level, and one, Amara davidi, reaches its lower limit. The altitudinal ranges of only four species (Amara congrua, Amara latithorax, Amara lucidissima and Amara sikkimensis) extend below about 1200 m, and those of only two species (Amara chalciope and Amara elegantula) include areas above 3200 m. It could be quite informative to monitor future changes in the altitudinal ranges of these species with respect to the baseline data recorded here as one measure of climate change.
Syntopy of species in the regional fauna. Because virtually all Amara species prefer habitats that are open and more or less disturbed, either by natural processes or through human activities, and because there is so much overlap in both the geographical and altitudinal ranges of species represented in the study area, it is not at all surprising to find that specimens of several different species can be found together at the same site in the same habitat (i.e., syntopic). This co-occurrence of species at sites in the region is summarized in Fig. 30. During fieldwork for this project, it was not uncommon to find several species, up to seven in at least one instance, together at the same time at the site. Four species, Amara birmana, Amara chalciope, Amara lucidissima and Amara sikkimensis, have been recorded syntopic with eight other species in one or more sites; three species, Amara dissimilis, Amara shaanxiensis and Amara silvestrii, have been found syntopic with seven other species; and Amara davidi, Amara latithorax and Amara pingshiangi have been recorded syntopic with four, five and three other species each, respectively. Members of these ten species are often found together, in various combinations, at different sites in the study area, although not one of them co-occurs with all the others. By contrast, three species have been found syntopic with just one other species. Amara congrua adults have been found together with those of Amara latithorax one time (in Shanga, Fugong County). Members of Amara simplicidens have been found just once with those of Amara lucidissima (near Datang Village, Tengchong County). Finally, adults of Amara elegantula have been found twice with those of Amara chalciope twice, both at sites around Heipu Yakou (Pass) on the Gongshan-Dulong Road at an elevation of 3365 m, where the altitudinal ranges of these species overlap narrowly. It is possible that Amara congrua and Amara simplicidens will be found co-occurring more broadly with the same species or with other species in the future because the geographical and/or altitudinal ranges of each are within the ranges of other species, whereas the restricted geographical and altitudinal ranges of Amara elegantula should preclude any but the already observed co-occurrence with Amara chalciope.
In the future, it will be interesting to compare and contrast both the broad and regional geographical distributions and the altitudinal and ecological range patterns seen in other carabid groups represented in the area with those found among the Amara species and reported here.
We thank our home institutions, the Museum für Naturkunde, Berlin, the California Academy of Sciences, San Francisco, Institute of Zoology, Beijing, and the Kunming Institute of Zoology, respectively, for ongoing support of our research programs. The Kunming Institutes of Botany and Zoology provided organization and logistical support for all our expeditions. The Yunnan Provincial Division of Forestry and Baoshan-Gaoligong and Nujiang Prefecture Forest Reserves granted access and collecting permits for our work in the reserves, and staffs of these institutions assisted with collecting and other efforts in these areas. Support for fieldwork for this project was provided through Grant No. DEB-0103795 from the National Science Foundation (Biotic Surveys and Inventories Program), Grant No. 30570213 from the National Science Foundation of China, grants from the National Geographic Society and the John D. and Catherine T. MacArthur Foundation, and private donations to the China Natural History Project at the California Academy of Sciences.
We owe special thanks to our many colleagues who assisted with the collecting effort over the ten-year duration of the fieldwork for this project and who, collectively, contributed many important specimens for this study. These included Bruce Bartholomew, Roberta L. Brett, Thomas S. Briggs, Peter W. Fritsch, David Garfield, Charles E. Griswold, Vincent F. Lee, Dong Lin, Paul E. Marek, Jeremy A. Miller, Pierre Paquin, Norman D. Penny, Darrel Ubick and Xinping Wang (U.S.A. team members); Weidong Ba, Keji Guo, Qingbai Hou, Peng Hu, Chunhua Li, Cailian Li, Rong Li, Xueyan Li, Xingcai Liang, Ye Liu, Zichao Liu, Chunlin Long, S. Z. Ma, Xuejun Meng, Guangxu Peng, Xianjin Peng, Yuehua San, Hongliang Shi, Xiaochun Shi, Guo Tang, Jun Wang, Meng Xie, J. Xiong, Xiang Xu, Hengmei Yan, Jialian Yang, Jiayu Yang, Song Yang, Jinfeng Zhang and Benxiang Zhu (Chinese team members); and Marilyn A. Dickson, David G. Long and Philip Thomas (of the Royal Botanical Garden, Edinburgh). Habitus images were taken by Victor G. Smith (CAS, Entomology), and Stan Blum (CAS, Informatics) and Lauren Scheinberg (CAS, Herpetology) provided assistance with generation of distribution maps from digital GPS data.