





Citation: Yoshida T, Hirowatari T (2014) A revision of Japanese species of the genus Psammoecus Latreille (Coleoptera, Silvanidae). ZooKeys 403: 15–45. doi: 10.3897/zookeys.403.7145
Japanese species of the genus Psammoecus Latreille, 1829 are taxonomically revised. Four new species, P. scitus sp. n. (misidentified with P. quadrimaculatus), P. labyrinthicus sp. n., P. boreas sp. n. and P. omotoensis sp. n. are described. Psammoecus bipunctatus (Fabricius, 1792), P. trimaculatus Motschulsky, 1858 (misidentified with P. triguttatus), P. simoni Grouvelle, 1892, P. fasciatus Reitter, 1874 and P. triguttatus are redescribed. Another described species whose distribution in Japan is questionable. P. quadrimaculatus is also redescribed. Lectotype and paralectotype of P. fasciatus and P. triguttatus are designated.
Taxonomy, Cucujoidea, Japan, four new species, misidentification, redescription, taxonomic key
The family Silvanidae Kirby 1837 (Coleoptera, Cucujoidea) includes two subfamilies and about 58 genera, approximately 500 species (
The genus Psammoecus Latreille, 1829 (Brontinae, Telephanini) includes about 80 described species and is the second largest genus in the Silvanidae (
The species of Psammoecus are found in plant detritus and they are sometimes attracted to light (
As stated above, taxonomy of this genus involves many problems and it has a potential to provide important pests and alien species. Hence, the authors here review the classification of Japanese Psammoecus by comparison of morphological characters and seek to resolve the taxonomic problems.
External characters were observed under a stereoscopic microscope (Olympus SZX10). Genital structures were placed on a cavity slide glass with 50% glycerol solution and observed with an optical microscope (Nikon Eclipse E400). The genitalia slide was prepared in the following steps: the removed abdomen was placed in a 200 µl PCR tube filled with 10% solution of potassium hydroxide (KOH) and kept in heated water for about seven minutes. After rinsing in 70% ethanol solution, the abdomen was dissected by cutting its side using fine insect pins. The genitalia were removed and transferred to a cavity slide glass with 50% glycerol solution for observation. After the observation, the genitalia and the abdomen were mounted in Euparal on cover glasses each glued to a piece of cardboard, and pinned with the specimens.
Photographs were taken with digital camera (Canon EOS 7D), and composite images were produced using automontage software Combine ZM. These images were retouched using Photoshop 6.0 (Adobe Systems Inc.)
Technical terms of genital structures follow
Depositories of the examined specimens are in the Ehime University Museum, Matsuyama (EUMJ), the Natural History Museum, London (BMNH), the Osaka Museum of Natural History, Osaka (OMNH), the Systematic Entomology, Hokkaido University, Sapporo (SEHU), Isamu Tanaka Collection, Nishinomiya (ITC), Yukihiko Hirano Collection, Odawara (YHC) and the Entomological Laboratory, Kyushu University, Fukuoka (ELKU).
http://zoobank.org/EEAFC47C-A1E6-4E6F-B4CF-0F099AE6686F
http://species-id.net/wiki/Psammoecus_scitus
Japanese name: Yotsumon-semaru-hiratamushi
Figs 1A, 3 and 13A–CThis species is similar to Psammoecus hiranoi Yoshida & Hirowatari, 2013, especially in darker specimens. However they can be distinguished by the ventral shape of the head. The temples of this species are immediately narrowed behind eyes, while those of Psammoecus hiranoi are widened behind the eyes and gradually narrowed toward the anterior margins.
Body length. 2.65–3.26 mm (n=48).
Coloration (Fig. 1A). Head and pronotum yellowish-brown to reddish-brown. Elytra black with yellowish-brown maculae; horizontal band at anterior 1/4, sometimes enlarged toward bases of elytra, round maculae at posterior 1/4, connected to lateral margins. Antennae yellowish-brown basally, 6th antennomere darker, 7th to 10th blackish-brown, 11th (apex) very bright.

Habitus of Psammoecus spp. A Psammoecus scitus sp. n., holotype B Psammoecus bipunctatus (Fabricius, 1792) C Psammoecus trimaculatus Motschulsky, 1858 D Psammoecus triguttatus Reitter, 1874, lectotype E Psammoecus labyrinthicus sp. n., holotype. Scale: 1.0 mm.
Head (Fig. 3A, B, C). Broad, HW/HL 1.31–2.00; IE/HL 0.87–1.30. Temples immediately narrowed behind eyes, slightly incised at bases. Dorsal surface with comparatively coarse punctuation, ventral surface with very sparse punctures. Antennae 1.40–1.43 mm long, slender; covered relatively sparsely with long erect pubescence on each antennomere; approximate ratio of antennomeres of holotype as follows: 2.5: 1.1: 1.2: 1.3: 1.3: 1.3: 1.1: 1.2: 1.2: 1.0: 2.0 (Fig. 3A).
Pronotum (Figs 3B, C). Subquadrate, PW/PL 1.14–1.32. Punctuation on dorsal surface comparatively sparse, punctuation on ventral surface sparser than on dorsal surface. Pubescence composed of many short setae, and long setae on teeth on lateral margins and anterior and posterior angles. Anterior angles with a few very small protrusions, lateral margins with several small teeth, these teeth variable, slightly longer on lateral margins around anterior and posterior angles, and each posterior angle with very small teeth.
Elytra (Fig. 3E). Elongate-oval, EW/BL 0.38–0.51. Rows of punctures narrower than interstices. Pubescence composed of many semi-long medium length setae, and some long erect setae in a row around lateral margins, longer toward anterior portion.
9th abdominal sternite (Fig. 13A). Strut very long, narrow, cut at anterior 3/8, diverging gradually at posterior 1/8, branches comparatively small. Lateral sclerites elongate with small sclerites attached around posterior apical portion.
Aedeagus (Fig. 13B, C). Parameres broad, depressed at base, narrowed on inner margins in posterior half, incised around apex, punctuated very sparsely, a few long setae around apex, several short setae on lateral margins and posterior half of inner margins. Phallobase diverging in anterior half, each branch twisted at anterior 1/4. Penis relatively elongate, curved dorsally before apex.
Holotype: male, Kûra, Ishigaki Island, Okinawa Prefecture, Japan, 22–IX–2012, T. Yoshida leg. (EUMJ). Paratypes: [Kagoshima Pref.] 1 ex., Honcha, Amami-Ôshima Island, 23–VII–1962, N. Ohbayashi leg. (EUMJ); 1 ex., Nadakawa, Amami-Ôshima Island, 16–VII–1962, N. Ohbayashi leg. (EUMJ); 1 ex., Kamiya, Amami-Ôshima Island, 4–XI–1984, M. Tomokuni leg. (EUMJ). [Okinawa Pref.] 1 ex., Hiji, Kunigami Village, Okinawa Island, 15–20–VI–1994, K. Okada leg. (EUMJ); 1 male & 1 female, Takazato, Kunigami Village, Okinawa Island, 18–22–VI–2003, I. Tanaka leg. (ITC); 2 exs., Inamine, Nago City, Okinawa Island, 26–IX–2012, T. Yoshida leg. (ELKU); 1 ex., Shokubutsuen, Miyako Island, 3–III–1999, T. Mizoguchi leg. (EUMJ); 1 female, Kûra, Ishigaki Island, 22–IX–2012, T. Yoshida leg. (ELKU); 4 exs., Ôura Dam, Ishigaki Island, 7–X–2013, R. Itô leg. (ELKU); 2 exs., Mt. Yarabu-dake, Ishigaki Island, 26–III–2000, T. Kurihara leg. (EUMJ); 7 exs., same locality, 16–VI–2002, T. Watanabe leg. (ELKU); 5 exs., same locality, 8–X–2013, R. Itô leg. (ELKU); 1 male, same locality, 8–III–2006, I. Tanaka leg. (ITC); 1 female, Kabira, Ishigaki Island, 19–IV–2010, I. Tanaka leg. (ITC); 2 exs., Takeda Rindô, Ishigaki Island, 14–III–1999, T. Mizoguchi leg. (EUMJ); 2 exs., same locality, 16–VI–2002, T. Watanabe leg. (ELKU); 2 exs., same locality, 22–IX–2012, T. Yoshida leg. (ELKU); 6 exs., same locality, 23–IX–2012, T. Yoshida leg. (ELKU); 3 exs., Funaura, Iriomote Island, 18–III–2008, S. Yamamoto leg. (ELKU); 1 ex., Urauchi-gawa, Iriomote Island, 2–VII–1994, K. Okada leg. (EUMJ); 3 exs., Komi, Iriomote Island, 20–III–2008, S. Yamamoto leg. (ELKU); 1 ex., Ôtomi Rindô, Iriomote Island, 13–III–1999, T. Mizoguchi leg. (EUMJ); 1 ex., Mt. Inbi-dake, Yonaguni Island, 31–III–2009, S. Yamamoto leg. (ELKU).
JAPAN: Amami-Ôshima, Okinawa, Miyako, Ishigaki, Iriomote and Yonaguni Islands.
This species is found in piled dead leaves such as Japanese pampas grass Miscanthus sinensis Andersson, Banana Musa sp. and some other kinds of broad-leaved tree. This species is most common in the Nansei Islands.
The specific name means ‘pretty’ and ‘beautiful’. This new species has vivid coloration.
Psammoecus scitus sp. n. has been misidentified as Psammoecus quadrimaculatus Reitter, 1874 for a long time. Psammoecus quadrimaculatus was originally described from ‘Japonia’ (=Japan) by
http://species-id.net/wiki/Psammoecus_bipunctatus
[Japanese name: Futamon-semaru-hiratamushi]
Figs 1B, 4 and 13D–FThis species is similar to Psammoecus trimaculatus and Psammoecus boreas sp. n., but can be distinguished from the former by the shorter teeth on the lateral margins of the pronotum and from the latter by the shorter body length and the shorter antennae.
Body length. 2.25–2.83 mm (n=9).
Coloration (Fig. 1B). Head dark brown, gradually darker toward anterior. Pronotum yellowish-brown to reddish-brown. Elytra yellowish-brown with round dark maculae on posterior 1/3, elytral suture dark in posterior half. Elytra of lighter colored specimens with reduced dark maculae and dark areas of elytral suture. Antennae yellowish-brown basally, 9th, 10th and posterior end of 8th antennomeres blackish-brown, 11th (apex) yellowish-brown.
Head (Fig. 4A, B, C). Triangular, HW/HL 1.55–2.06; IE/HL 1.03–1.33. Dorsal surface with strong dense punctures, temples and posterior ventral surface also with irregularly dense punctures. Temples well enlarged, immediately narrowing thereafter. Eyes prominent, relatively small. Antennae 1.09–1.20 mm, comparatively thick, short; covered with medium length erect pubescence on each antennomere; approximate ratio of one of examined specimens as follows: 2.6: 1.1: 1.2: 1.1: 1.2: 1.2: 1.1: 1.0: 1.1: 1.0: 1.8 (Fig. 4A).
Pronotum (Fig. 4B, C). Roundly subquadrate, PW/PL 1.16–1.24. Dorsal surface with strong moderately dense punctuation, denser than that on ventral surface; some punctures on ventral surface in rows. Pubescence composed of many short setae on dorsal surface and several medium length setae on each lateral margin, distance between medium length setae approximately regular. Each anterior angle with several very small teeth, each lateral margin with four similarly sized small teeth, the distance between them irregular and variable; each posterior angle with a small tooth, similar in size to those on lateral margins.
Elytra (Fig. 4E). Elongate, EW/BL 0.36–0.47. Rows of punctures almost same width as interstices. Pubescence composed of numerous comparatively short setae, without long setae.
9th abdominal sternite (Fig. 13D). Strut Y-shaped, cut at anterior 1/3, diverging in posterior 1/4, branches relatively wide, ends of each branch pointed and curved inwards. Lateral sclerites elongate, membranous.
Aedeagus (Figs 13E and F). Parameres club-shaped, wide and thick at bases, each apical portion with a long seta, stick-shaped portions with sparse punctuation and several short setae, lateral portions and inner margins of bases with successions of dense punctures, inner margin of bases with numerous short setae. Phallobase enlarged toward posterior margin from posterior 1/3, anterior margin roundly incised, anterior portion extended. Penis flat, wide, narrowed gradually toward apex, apical portion pointed, apical portion of dorsal part with sparse punctures.
JAPAN: [Hokkaido Pref.] 1 male, Takkobu-numa Lake, Kushiro City, 1–IX–2006, T. Yoshida leg. (ELKU); 1 ex., Iwabokki, Kushiro City, 25–VIII–1990, M. Sakai leg. (EUMJ); 6 exs., same locality, 26–VIII–1990, M. Sakai leg. (EUMJ); 1 ex., same locality, 29–VIII–1990, M. Sakai leg. (EUMJ).
JAPAN: Hokkaido; Africa; Europe; Russia; Turkestan.
This species is known to inhabit marshland.
The habitat and distribution of this species are unusual in the genus Psammoecus. This species inhabits marshland and is distributed in comparatively high latitudes where species richness of Psammoecus is poor. Coloration is variable. In the past, four aberrations and two varieties (e.g.
http://species-id.net/wiki/Psammoecus_trimaculatus
Japanese name: Mitsumon-semaru-hiratamushi
Figs 1C, 5 and 13G–IThis species is closely similar to Psammoecus triguttatus and Psammoecus labyrinthicus sp. n. but it can be distinguished from the former by the larger basal parameres and the broader distance between the posterior margin of the phallobase and the deepest point of incision of the anterior margin of the phallobase, and from the latter by the larger bases of parameres and the wider protuberances on the inner margins of the branches of the anterior phallobase. The rows of punctures on the elytra of Psammoecus trimaculatus (Fig. 5E) are comparatively narrower than in these two closely similar species. However, their external characters, especially their coloration, are variable. These features may cause misidentification.
Body length. 2.32–2.96 mm (n=38).
9th abdominal sternite (Fig. 13G). Strut Y-shaped, elongate, cut at anterior 3/8, diverging in posterior 1/5, branches relatively wide, narrowed gradually toward apex, ends of each branch curved inward. Lateral sclerites elongate, membranous.
Aedeagus (Fig. 13H, I). Parameres hatchet-shaped, inner margin around bases pointed, punctuated sparsely on stick-shaped portions, densely on anterior portions, a long seta at each apex, numerous short setae on stick-shaped portions and around inner margins of anterior parts. Posterior margin of phallobase rounded, phallobase roundly hollowed, its anterior branches markedly protruding inward. Penis gradually narrowed toward apex, posterior 1/9 flattened, punctuated sparsely on posterior 1/9.
India: [Andhra Pradesh State] 1 ex., Hyderabad, 18–X–1969, T. Ishihara leg. (EUMJ). JAPAN: [Hokkaido Pref.] 1 male, Suehiro, Asahikawa City, 18–IX–2013, T. Yoshida leg. (ELKU); 1 male, Aizankei, Kamikawa Town, 2–IX–1977, A. Oda leg. (EUMJ); 1 ex., Sôunkyo, Kamikawa Town, 4–IX–1977, A. Oda leg. (EUMJ); 1 ex., Chûbisei, Memuro Town, 25–VIII–1995, S. Hisamatsu leg. (EUMJ); 2 exs., Fushimi Marsh, Memuro Town, 25–VIII–1995, S. Hisamatsu leg. (EUMJ). [Aomori Pref.] 1 male, Namioka Ôaza Yoshinoda, Aomori City, 16–IX–2012, K. Ikeuchi leg. (ELKU). [Gifu Pref.] 1 male, Tentoku, Mizunami City, 31–VII–2009, K. Itô leg. (ELKU). [Hyogo Pref.] 1 male, Mt. Mikusa-yama, Inagawa Town, 27–VI–1991, K. Ikeuchi leg. (ELKU). [Wakayama Pref.] 2 males, 3 females & 7 exs., Biwadani, Kinokawa City, 25–IV–2012, T. Yoshida leg. (ELKU). [Kagawa Pref.] 1 male, Yoshima Island, Sakaide City, 10–12–IX–1973, S. Kinoshita leg. (EUMJ). [Tokushima Pref.] 2 males, 2 females & 4 exs., Kamojima-chô, Yoshinogawa City, 4–III–2012, T. Yoshida leg. (ELKU). [Kochi Pref.] 1 male, Godaisan-Park, Kochi City, 21–V–1983, K. Ishida leg. (EUMJ). [Fukuoka Pref.] 1 male, Kitano-chô, Kurume City, 1–VII–2013, T. Yoshida leg. (ELKU). [Kagoshima Pref.] 1 male, Kotoku-gawa, Amami-Ôshima Island, 14–III–1988, M. Satô leg. (EUMJ). [Okinawa Pref.] 1 male, Yona, Kunigami Village, Okinawa Island, 17–VII–1965, Y. Hori leg. (EUMJ); 1 male, same locality, 19–X–1987, M. Sakai leg. (EUMJ); 1 male & 1 ex., Inaba, Iriomote Island, 9–VIII–1962, M. Satô & Y. Arita leg. (EUMJ).
JAPAN: Hokkaido, Honshu, Shikoku, Kyushu, Amami-Ôshima, Okinawa, and Iriomote Islands. (Madagascar recorded by
This species was redescribed by Pal (1980) with a description of the larva. It is distributed worldwide except for Europe and North America and common at least in Japan and India (
http://species-id.net/wiki/Psammoecus_triguttatus
Japanese name: Nise-mitsumon-semaru-hiratamushi
Figs 1D, 6 and 13J–LThis species is closely similar to Psammoecus trimaculatus and Psammoecus labyrinthicus sp. n. Morphological differences among these species were stated in diagnosis of Psammoecus trimaculatus.
Description. Body length. 2.32–2.93 mm (n=23).
Coloration (Fig. 1D). Head and pronotum yellowish-brown to reddish-brown. Elytra yellowish-brown with variable dark maculae: round ones at half, oval ones on the posterior half to posterior 1/4 of elytral suture, sometimes connected with maculae at half, darkened around end of elytra. Antennae reddish-brown basally, blackish-brown from 8th to 10th antennomeres, 11th (apex) comparatively bright.
Head (Fig. 6A, B, C). Triangular, HW/HL 1.46–1.90; IE/HL 0.92–1.20. Temples slightly expanded behind eyes, narrowed toward posterior. Eyes large, prominent. Dorsal surface with relatively dense punctuation. Antennae 1.06–1.38 mm, moderately long; covered with considerable medium length or long semi-erect setae and short pubescence on each antennomere; approximate ratio of lectotype as follows: 2.7: 1.0: 1.1: 1.2: 1.3: 1.3: 1.3: 1.1: 1.1: 1.1: 2.0 (Fig. 6A).
Pronotum (Fig. 6B, C). Broad, PW/PL 1.31–1.50. Dorsal surface with relatively dense punctuation. Pubescence composed of relatively fine short or medium length setae, a long seta on each tooth on lateral margins and anterior and posterior angles. Each anterior angle with four or five small teeth; each lateral margin with four teeth, tooth I small, tooth II longer than tooth I, teeth III and IV relatively narrow and of almost identical size, longer than tooth II, tooth V smallest or absent; each posterior angle with a few very small protuberances of variable shape.
Elytra (Fig. 6E). Elongate-oval, EW/BL 0.39–0.45. Rows of punctures narrower than interstices. Pubescence composed of many semi-erect setae of medium length; several long setae in a row around anterolateral margins.
9th abdominal sternite (Fig. 13J). Strut Y-shaped, cut at anterior 2/5. Lateral sclerites curved inward, relatively elongate.
Aedeagus (Fig. 13K, L). Parameres club-shaped, wide around bases, posterior inner margins of wide portions a little prominent, punctuation on narrow portions relatively sparse, a little denser on wide portions, posterior half of inner margins of wide portions with many setae, narrow portions with several sparse setae, apex with a long seta. Posterior half of phallobase wider towards posterior margin, distance between posterior margin and deepest point of incision of margin of upper layer narrow, protuberances around anterior 1/4 pointed inwards, posterior margin of lower layer widely deeply incised. Penis relatively flat and thin, narrowed toward apex, punctuation around apex denser toward apex.
Lectotype: male, Nagasaki, Nagasaki Prefecture, Japan, 1869, G. Lewis leg. (BMNH). Paralectotype: 1 female, same locality, 1869, G. Lewis leg. (BMNH). (here designated)
JAPAM: [Tokyo Pref.] 1 male, Hatagaya, Shibuya-ku, 30–VII–1956, K. Tanaka leg. (EUMJ). [Gifu Pref.] 1 male, Tentoku, Toki-chô, Mizunami City, 6–VIII–2010, K. Itô leg. (ELKU); 1 male, Konokure, Toki-chô, Mizunami City, 16–VII–2011, K. Itô leg. (ELKU). [Kagoshima Pref.] 1 male, 4 females and 3exs., Sata Cape, Minamiôsumi Town, 6–VII–1968, K. Suga leg. (HUSE); 1 male, Nakanoshima Island, Toshima Village, 9–VII–1969, M. Satô leg. (EUMJ); 1 male, same locality, 26–VI–1965, S. Fukuda leg. (HUSE). [Okinawa Pref.] 1 male, Aha beach, Kunigami Village, Okinawa Island, 23–I–2008, Y. Hirano leg. (YHC); 2 males and 2 exs., Motobu Town, Okinawa Island, 19–VIII–2008, T. Yoshida leg. (ELKU); 1 male, Fusato, Nanjô City, Okinawa Island, 5–VI–1970, M. Takagi leg. (EUMJ); 1 male, Mt. Omoto-san, Ishigaki Island, 1–VII–1965, Y. Hori leg. (EUMJ); 1 male, Ushuku-no-mori, Iriomote Island, 26–VI–1965, Y. Hori leg. (EUMJ).
JAPAN: Honshu, Kyushu, Nakanoshima (Tokara Islands), Okinawa, Ishigaki and Iriomote Islands.
Psammoecus sp. 4 illustrated by
Syntypes of this species consist of two specimens, one male and one female. The male specimen is designated as a lectotype, and the female specimen is designated as a paralectotype.
http://zoobank.org/D11F2212-7A76-46A8-951C-B3FCC1543BCC
http://species-id.net/wiki/Psammoecus_labyrinthicus
[Japanese name: Hachijô-mitsumon-semaru-hiratamushi]
Figs 1E, 7 and 14A–CThis species is closely similar to Psammoecus trimaculatus and Psammoecus triguttatus. However, it can be distinguished by the male genital structure, especially the shape of the parameres, and the comparatively dense punctuation of the dorsal pronotum.
Body length: 2.66–3.38 mm (n=25).
Coloration (Fig. 1E). Head and pronotum reddish-brown. Elytra yellowish-brown with dark maculae; round ones at half, oblong ones on the posterior half to posterior 1/4 along elytral suture, darkened around end of elytra: these maculae sometimes connected each other. Antennae reddish-brown basally, 7th antennomere darkened, from 8th to 10th blackish-brown, 11th (apex) comparatively bright.
Head (Fig. 7A, B, C). Rounded-triangular, HW/HL 1.25–2.00; IE/HL 0.75–1.27. Temples slightly expanded behind eyes, gradually narrowed. Eyes large, prominent. Dorsal surface with dense and strong punctuation. Antennae 1.35–1.56 mm, moderately long; covered with medium length semi-erect pubescence and some long erect setae on each antennomere; approximate ratio of holotype as follows: 2.4: 1.0: 1.0: 1.2: 1.3: 1.1: 1.1: 1.1: 1.1: 1.1: 1.9 (Fig. 7A).
Pronotum (Fig. 7B, C). Roundly subquadrate, PW/PL 1.25–1.53. Relatively strong and dense punctuation on dorsal surface. Pubescence composed of short setae, a long seta on each tooth on lateral margins and anterior and posterior angles. Each anterior angle with several small teeth, each lateral margin with four short teeth; tooth I small, tooth II longer than tooth I, teeth III and IV almost the same size, longer than tooth II, teeth II, III and IV broadened at base, each posterior angle with a very small tooth, the shape of these teeth variable.
Elytra (Fig. 7E). Elongate-oval, EW/BL 0.41–0.47. Rows of punctures narrower than interstices. Pubescence composed of numerous medium length semi-erect setae and some long setae in a row around lateral margins.
9th abdominal sternite (Fig. 14A). Strut relatively short, cut at anterior 1/3. Lateral sclerites fused with strut, curved inwards.
Aedeagus (Fig. 14B, C). Parameres stout, club-shaped, punctuated on anterior half of inner margins, anterolateral portions well punctuated, anterior half of inner margins with many setae of variable size, apex with a long seta. Phallobase broadened toward posterior margin, posterior margin widely incised, protuberances around 1/3 of inner margins of branches, especially right protuberance thin. Penis flat, narrowed toward apex, apex pointed, punctuated around anterior 1/8, apex densely punctuated.
Holotype: male, Hachijô Island, Hachijô Town, Tokyo Prefecture, Japan, 21–VII–1957, S. Hisamatsu leg. (EUMJ). Paratypes: [Tokyo Pref.] 3 females, same data as holotype. (EUMJ). [Mie Pref.] 4 males, Ujitachi-chô, Ise City, 29–VIII–2009, N. Narukawa leg. (ELKU). [Nagasaki Pref.] 1 male and 11 exs., Ta, Toyotama-chô, Tsushima Island, 7–VII–2013, T. Yoshida leg. (ELKU). [Kagoshima Pref.] 2 males and 3 exs., Takarajima Island, Toshima Village, 2–VII–1960, M. Satô leg. (EUMJ); 1 ex., same locality, 3–VII–1960, M. Satô leg. (EUMJ); 1 ex., same locality, 1–VI–1962, M. Satô leg. (EUMJ).
JAPAN: Honshu, Hachijô (Izu Islands), Tsushima, Takarajima Islands (Tokara Islands).
The first author (Yoshida) collected many individuals of this new species from dead leaves of evergreen trees in Tsushima Island, Nagasaki Prefecture (Fig. 15A).
The specific name means ‘labyrinthine’. Identification of Psammoecus trimaculatus and closely similar species is very difficult. The addition of this new species to this group of closely similar species made their identification more difficult like the labyrinth.
Psammoecus trimaculatus is closely related to this species, however the former has been often collected from dead leaves of monocotyledon plant. These two species may have each distinct ecological trait.
http://zoobank.org/1E58DE65-FC6D-4E40-B55E-A952B311A454
http://species-id.net/wiki/Psammoecus_boreas
Japanese name: Arame-semaru-hiratamushi
Figs 2A, 8 and 14D–FThis species is similar to Psammoecus trimaculatus and other species closely similar to Psammoecus trimaculatus.
Body length. 2.74–3.27 mm (n=19).
Coloration (Fig. 2A). Head and pronotum yellowish-brown. Elytra yellowish-brown with dark brown maculae at half, oval ones at center of each elytron and dark oblique bands toward posterior portion, sometimes connected at elytral suture, forming a V-shaped band. Elytra of lighter color specimens with reduced maculae, oval ones and bands separated. Antennae yellowish-brown basally, posterior ends of 8th to 10th antennomeres darkened, or all antennomeres yellowish-brown in lighter colored specimens.

Habitus of Psammoecus spp. A Psammoecus boreas sp. n., holotype B Psammoecus omotoensis sp. n., holotype C Psammoecus simoni Grouvelle, 1892 D Psammoecus fasciatus Reitter, 1874, lectotype E Psammoecus hiranoi Yoshida & Hirowatari, 2013 F Psammoecus quadrimaculatus Reitter, 1874, holotype. Scale: 1.0 mm.
Head (Fig. 8A, B, C). Broad, HW/HL 1.35–1.65; IE/HL 0.96–1.13. Temples narrowed around each base. Eyes small, moderately rounded. Dorsal surface with moderately dense punctuation, ventral surface punctuated sparsely. Antennae 1.54–1.71 mm, thin, very long; covered with considerable long semi-erect pubescence on each antennomere; approximate ratio of holotype as follows: 2.6: 1.0: 1.1: 1.2: 1.2: 1.3: 1.2: 1.0: 1.0: 1.1: 1.7 (Fig. 8A).
Pronotum (Fig. 8B, C). Subquadrate, PW/PL 1.18–1.30. Dorsolateral portions lightly impressed. Dorsal surface with relatively dense punctuation, no punctures around posterior margin, comparatively sparse punctuation on ventral surface. Pubescence composed of many short setae and fine long setae on teeth on lateral margins and anterior and posterior angles. Each anterior angle with a distinct group of a few very small teeth, each lateral margin with four small teeth of almost the same size, each posterior angle with a small tooth, almost the same size as those on lateral margins.
Elytra (Fig. 8E). Oval, EW/BL 0.32–0.45. Rows of punctures wider than interstices. Pubescence composed of many short setae, no long setae.
9th abdominal sternite (Fig. 14D). Strut Y-shaped, cut deeply at anterior 1/5, diverging for posterior 1/4. Lateral sclerites elongate.
Aedeagus (Fig. 14E, F). Parameres cone-shaped with almost even sparse punctuation, sparser on bases, a few long setae around apical portions, a few short setae distributed sparsely. Phallobase tube-like, consisting of two layers, anterior margin rounded, dorsal surface around anterior margin thin, protuberances of upper layer directed towards anterior portion, small protuberances at beginning of divergence of upper layer. Penis stout, punctuated on posterior 1/9, rather coarsely on ventral surface.
Holotype: male, Yoshin, Tanzawa, Kanagawa Prefecture, Japan, 26–V–1989, Y. Hirano leg. (EUMJ). Paratypes: [Hokkaido Pref.] 2 exs., Chûbisei, Memuro Town, 25–VIII–1995, S. Hisamatsu leg. (EUMJ); 1 ex., Mt. Sapporo-dake, Sapporo City, 5–VIII–1970, S. Kinoshita leg. (EUMJ). [Niigata Pref.] 1 ex., Mikuni Touge, 1–VII–1967, K. Baba leg. (HUSE). [Kanagawa Pref.] 1 ex., Yoshin, Tanzawa, 26–V–1989, Y. Hirano leg. (YHC). [Nagano Pref.] 1 ex., Tokugo Touge, 29–VII–1955, T. Nakane leg. (HUSE); 5 exs., Ôbora, Ueda City, 24–VII–2013, T. Yoshida leg. (ELKU). [Oita Pref.] 6 ex., Mt. Sobo-san, 7–VI–2009, S. Yamamoto leg. (ELKU).
JAPAN: Hokkaido, Honshu, Kyushu.
The specific name is from the god of the north wind of ancient Greek mythology. Most Psammoecus species are distributed in tropical or subtropical zones, however, this new species is exceptionally distributed in Hokkaido or on mountains of high altitude located in Honshu and Kyushu.
Psammoecus sp. 3 illustrated by
http://zoobank.org/E64E6D2D-B71B-40FC-83FA-551947595432
http://species-id.net/wiki/Psammoecus_omotoensis
Japanese name: Higenaga-semaru-hiratamushi
Figs 2B, 9 and 14G–IThis species is similar to Psammoecus trimaculatus and other species closely similar to Psammoecus trimaculatus. However, it can be distinguished from them by the male genital structure, especially the parameres which are fused to the phallobase. The distinguishing external characters of this species are tooth IV of the lateral margins of pronotum extended in a posterolateral direction and the long antennae, especially the 1st antennomere.
Body length. 2.71–2.77 mm (n=2).
Coloration (Fig. 2B). Head and pronotum yellowish-brown to reddish-brown. Elytra yellowish-brown with dark maculae, round ones at half of each elytron, black ones on posterior half of elytral suture narrower toward posterior elytral suture, a thin short black band between these maculae. Antennae yellowish-brown, almost unicolorous, 11th (apical) antennomere comparatively bright.
Head (Fig. 9A, B, C). Transverse, HW/HL 1.59–1.63; IE/HL 1.07. Temples well enlarged behind eyes, narrowed at bases. Eyes small, moderately rounded. Frequent pubescence and very sparse punctuation on ventral surface, moderately dense punctuation on dorsal surface. Antennae 1.44–1.49 mm long; covered with frequent medium length semi-erect fine setae and some long erect setae on each antennomere; approximate ratio of holotype as follows: 2.6: 1.0: 1.2: 1.4: 1.4: 1.4: 1.3: 1.1: 1.2: 1.0: 1.9 (Fig. 9A).
Pronotum (Fig. 9B, C). Transverse, PW/PL 1.30–1.35. Punctuation on dorsal surface coarse. Pubescence composed of short fine setae, a long seta on each tooth on lateral margins and anterior and posterior angles. Each anterior angle with several very small prominences and two small teeth, each lateral margin with four teeth; tooth I comparatively small, teeth II and III almost the same size, longer than tooth I, tooth IV extended in posterolateral direction, longer than tooth II and III, each posterior angle with a very small protuberance.
Elytra (Fig. 9E). Oval, EW/BL 0.37–0.44. Around half of lateral margins relatively expanded. Rows of punctures wider than interstices. Pubescence composed of medium length semi-erect setae and a few long setae around anterolateral margins.
9th abdominal sternite (Fig. 14G) Strut Y-shaped, anterior 2/7 slightly thickened, cut from anterior 2/7, diverging in posterior 1/3, branches long, ends of each branch curved inwards. Lateral sclerites elongate, membranous.
Aedeagus (Figs 14H and I) Parameres fused with phallobase, incised shallowly at inner and lateral margins of bases, relatively stout, gradually curved inwards, three long setae around posterior 1/3, some short setae on posterior half, well punctuated on anterior half of inner margins. Phallobase broad and somewhat flat, anterior margin widely but shallowly incised, lateral margins slightly depressed around anterior 1/4. Penis wide, thinner toward apex, narrowed around apex, punctuated around apex, densely punctuated at apex.
Holotype: male, Mt. Omoto-san, Ishigaki Island, Okinawa Prefecture, Japan, 23–III–2000, T. Kurihara leg. (EUMJ). Paratype: [Okinawa Pref.] 1 male, same data as holotype, T. Kurihara leg. (EUMJ).
JAPAN: Ishigaki Island.
The specific name is derived from the type locality of the new species, Mt. omoto-san, Ishigaki Island, Okinawa Prefecture.
http://species-id.net/wiki/Psammoecus_simoni
Japanese name: Hababiro-semaru-hiratamushi
Figs 2C, 10 and 14J–LThis species is distinguished from other Japanese Psammoecus species by the black elytra with yellow maculae and the posterior teeth on lateral margins of pronotum longer than those of anterior margins.
Body length. 2.24–2.56 mm (n=8).
9th abdominal sternite (Fig. 14J). Strut comparatively short, cut at anterior 2/5, diverging gradually, branches comparatively long and wide. Lateral sclerites elongate, slightly curved inwards, apical sclerites pointed.
Aedeagus (Fig. 14K, L). Parameres fused with phallobase, wide, short, posterior and lateral portions punctuated sparsely, densely punctuated triangular areas in anterior portions, a few long setae and short setae on posterior half, several short folds around beginning of divergence of parameres, a few lines on upper densely punctuated areas. Phallobase stout, broad in lateral aspect, anterior margin straight, a few subparallel lines on posterior dorsal portion. Penis comparatively broad, flattened around apex, apical margin rounded, punctuated sparsely on apex.
JAPAN: [Okinawa Pref.] 1 male, 4 females & 2 exs., Inamine, Nago City, Okinawa Island, 26–IX–2012, T. Yoshida leg. (ELKU); 1 ex., Mt. Yarabu-dake, Ishigaki Island, 16–VI–2002, T. Watanabe leg. (ELKU).
JAPAN: Okinawa, Hateruma and Ishigaki Islands; Madagascar; India; Sri Lanka; Malaysia; Indonesia; Philippines.
This species is found in dead leaves to which fungi are attached and occurs sympatrically with other Silvanid species such as Psammoecus trimaculatus, Psammoecus scitus sp. n., Cryptamorpha desjardinsi (Guérin-Méneville, 1844) and Monanus concinnulus (Walker, 1858) (Fig. 15B).
Pal (1980) and
http://species-id.net/wiki/Psammoecus_fasciatus
Japanese name: Kuroobi-semaru-hiratamushi
Figs 2D, 11 and 14M–OThis species is closely similar to Psammoecus hiranoi. It can be distinguished by the longer parameres, the shallower incision of the phallobase on the anterior 1/4 and the shorter teeth on the lateral margins of the pronotum.
Body length. 2.75–3.50 mm (n=49).
Coloration (Fig. 2D). Head and pronotum yellowish-brown to reddish-brown. Elytra with a variable horizontal dark band at middle, this band in lighter color specimens thinner towards elytral suture, lateral margins, apical portion and posterior elytral suture dark. Antennae brown or reddish-brown basally, 7th and 8th antennomeres darker, 9th and 10th almost black, 11th (apex) lighter than basal ones. Antennae of some specimens almost unicolorous.
Head (Fig. 11A, B, C). Broad, HW/HL 1.40–2.08; IE/HL 0.94–1.38. Temples enlarged behind eyes, narrowed gradually toward posterior margin. Eyes large, slightly prominent. Ventral surface with rough irregular punctuation, sparser punctuation on dorsal surface. Antennae 1.38–1.57 mm, 2nd antennomere short; covered with thick medium length pubescence and some long erect pubescence on each antennomere; approximate ratio of lectotype as follows: 2.5: 1.0: 1.3: 1.3: 1.3: 1.3: 1.3: 1.3: 1.3: 1.2: 1.8 (Fig. 11A).
Pronotum (Fig. 11B, C). Broad, PW/PL 0.69–1.52. Punctuation on dorsal surface very dense, but no punctures around posterior margin, punctuation on ventral surface sparser than dorsal surface, some of them in rows especially on posterior half. Pubescence composed of frequent short setae on dorsal surface and a long seta on each tooth on lateral margins and at anterior and posterior angles. Each anterior angle with a distinct group of a few very small teeth, each lateral margin with four teeth; tooth I comparatively small, teeth II, III and IV almost the same moderate size, each posterior angle with a comparatively small tooth.
Elytra (Fig. 11E). Elongate-oval, EW/BL 0.41–0.47. Punctures wider than interstices. Pubescence composed of semi-erect medium length setae and some long erect setae in a row around lateral margins, longer toward anterior portion.
9th abdominal sternite (Fig. 14M). Strut cut at around half, diverging widely around the posterior base, ends of each branch curved inwards. Lateral sclerites triangular, flat.
Aedeagus (Figs 14N and O). Parameres simple stick-shaped, elongate, slightly curved inwards, punctuated sparsely except base, a few semi-long setae around apical portion. Phallobase diverging gradually around anterior 1/4, a little bulging along inner margins of each branch, lateral margins broadening. Penis also elongate, punctuated on posterior 1/6, more densely on ventral portion, especially densely on apical portion.
Lectotype: male, Mt. Maya-san, Kobe City, Hyogo Prefecture, Japan, 1871, G. Lewis leg. (BMNH). Paralectotype: 1 female, same locality, 1871, G. Lewis leg. (BMNH). (here designated)
Russia: [Primorsky Krai] 1 ex., Chuguevka Village, 27–VI–1999, Y. Notsu leg. (EUMJ). JAPAN: [Niigata Pref.] 1 male & 1 ex., Renge-Onsen, Itoigawa City, 14–VIII–2003, I. Tanaka leg. (ITC). [Tokyo Pref.] 7 exs., Mikura Island, Mikurajima Village, 11–VI–1977, T. Nakane leg. (HUSE). [Nagano Pref.] 1 ex., Togakushi-Kôgen, Nagano City, 5–6–VII–2008, K. Mizuno leg. (KMC); 2 ex., Ôbora, Ueda City, 24–VII–2013, T. Yoshida leg. (ELKU). [Kyoto Pref.] 1 ex., Kifune, Kyoto City, 6–IV–1988, I. Tanaka leg. (ITC). [Hyogo Pref.] 1 ex., Itoi, Asago City, 26–IX–2004, I. Tanaka leg. (ITC); 5 exs., Sakanotani, Shisô City, 19–VII–2009, K. Itô leg. (KMC); 1 ex., Onzui, Shisô City, 19–VII–2009, K. Utô leg. (KMC); 1 ex., Mt. Yuzuruha-san, Nantan Town, Awaji Island, 18–X–2003, I. Tanaka leg. (ITC). [Nara Pref.] 1 ex., Mt. Kasuga-yama, Nara City, 5–V–1985, I. Tanaka leg. (ITC); 2 exs., same locality, 5–V–2012, K. Ikeuchi leg. (ELKU); 1 ex., same locality, 31–VII–2012, K. Matsuda leg. (ELKU). [Tokushima Pref.] 1 ex., Mt. Shibakoya-yama, Kamiyama Town, 31–VII–1975, M. Yoshida leg. (EUMJ). [Ehime Pref.] 1 ex., Nishidani, Yanadani Village, 15–16–VII–1994, K. Aita leg. (EUMJ). [Kochi Pref.] 1 ex., Muroto Cape, Muroto City, 7–VI–1959, S. Hisamatsu leg. (EUMJ). [Nagasaki Pref.] 1 ex., Mt. Tatera-san, Tsushima Island 16–IX–1995, N. Narukawa leg. (ELKU); 3 exs., Midake, 12–X–1977, Y. Notsu leg. (EUMJ). [Kagoshima Pref.] 1 male & 4 exs., Kirishima, 29–VII–1971, T. Nakane leg. (HUSE); 2 exs., Shiratani-unsuikyô, Yakushima Island, 15–VIII–2005, J. Ogawa leg. (EUMJ); 1 male & 1 ex., Onoaida Hodô, Yakushima Island, 14–15–IX–2002, N. Ohbayashi leg. (EUMJ); 2 exs., Kosugidani, Yakushima Island, 2–V–1984, K. Mizuno leg. (OMNH); 4 exs., Anbô, Yakushima Island, 16–VII–1989, N. Narukawa leg. (ELKU); 5 exs., Nogi, Nishino-omote, Tanegashima Island, 10–VII–1974, S. Hisamatsu leg. (EUMJ).
JAPAN: Hokkaido, Honshu, Shikoku, Kyushu, Mikura (Izu Islands), Awaji, Yakushima and Tanegashima Islands; Burma; Korea; Russia.
The first author (Yoshida) collected this species from various kinds of dead leaves from evergreen trees, deciduous trees and bamboo.
The syntypes of this species consist of three specimens: two of them belong to this species, but the remaining one represents Psammoecus trimaculatus or Psammoecus triguttatus. We designate a male specimen as lectotype, and a female specimen as paralectotype. The type specimens were collected from Mt. Maya-san, Kobe City, Hyogo.
http://species-id.net/wiki/Psammoecus_hiranoi
Japanese name: Herimon-semaru-hiratamushi
Fig. 2EThis species is closely similar to Psammoecus fasciatus and morphological differences between these two species were stated in diagnosis of Psammoecus fasciatus.
The following specimens were found after description of this species:
JAPAN: [Okinawa Pref.] 1 male & 4 exs., Mt. Yonaha-dake, Kunigami Village, Okinawa Island, 3–IV–1974, T. Kinoshita leg. (EUMJ); 10 exs., Ôkuni-rindô, Kuigami Village, Okinawa Island, 4–XI–2013, T. Yoshida leg. (ELKU).
http://species-id.net/wiki/Psammoecus_quadrimaculatus
Japanese name: Ruisu-yotsumon-semaru-hiratamushi
Figs 2F, 12 and 14P–RThis species is similar to Psammoecus trimaculatus, Psammoecus triguttatus and Psammoecus labyrinthicus sp. n., but can be distinguished from Psammoecus trimaculatus and Psammoecus triguttatus by the wide triangular basal portion of the parameres and the apically narrow portion of the penis, and from Psammoecus labyrinthicus sp. n. by the longer parameres, the apically narrow portion of the penis and the shape of the phallobase.
Body length. 2.50 mm (n=1).
Coloration (Fig. 2F). Head reddish-brown, pronotum somewhat light reddish-brown. Elytra blackish-brown with four large yellowish-brown maculae; macula around anterior 1/4 of each elytron almost quadrate, macula on posterior half longer than wide. Antennae yellowish-brown basally, 6th to 10th antennomeres black, 6th slightly brighter, 11th (apex) bright.
Head (Figs 12A, B and C). Rounded-triangular, HW/HL 1.68; IE/HL 1.09. Temples slightly expanded behind eyes, narrowed at base. Eyes large, prominent, diameter about half of length of head. Punctuation of dorsal surface moderately dense, on ventral surface sparse, and absent on center portion of ventral surface. Antennae 1.44 mm; covered with medium length pubescence and some relatively long erect setae on each antennomere; approximate ratio of holotype as follows: 2.4: 1.0: 1.0: 1.1: 1.1: 1.3: 1.0: 1.1: 1.2: 1.1: 2.2 (Fig. 12A).
Pronotum (Fig. 12B, C). Roundly subquadrate, PW/PL 1.34. Punctuation on dorsal surface relatively strong and moderately sparse. Pubescence composed of medium length setae, a long seta on each tooth on lateral margins and anterior angles, a relatively long seta on each posterior angle. Each anterior angle with several small teeth, each lateral margin with four short teeth; tooth I small, tooth II longer than tooth I, teeth III and IV almost same size, longer than tooth II, teeth II, III and IV relatively widened around base, each posterior angle with a few very small teeth.
Elytra (Fig. 12E). Elongate-oval, EW/BL 0.46. Rows of punctures wider than interstices. Pubescence composed of many medium length semi-erect setae, long erect setae in a row on lateral margins.
9th abdominal sternite (Fig. 14P). Strut cut at anterior 1/3, diverging deeply around posterior 1/3. Lateral sclerites rhomboid, comparatively large, curved inwardly.
Aedeagus (Fig. 14Q, R). Parameres club-shaped; narrow portions relatively broad, punctuated sparsely, with several sparse setae, apex with a long seta; wide portions punctuated densely on posterior half of inner margins and anterolateral portions of outer margins, posterior half of inner margins with many setae. Phallobase consisting of two layers, posterior margin incised roundly, distance between posterior margin and deepest point of incision of margin of upper layer narrow, anterior margin of lower layer relatively narrowly incised, protuberances around anterior 1/4 narrow, projecting inwards, posterior margin of lower layer broadly incised. Penis relatively elongate and flat, with relatively dense punctuation on posterior 1/8.
Holotype: male, Nagasaki, Nagasaki Prefecture, Japan, 1869, G. Lewis leg. (BMNH).
JAPAN: Nagasaki?.
Type specimen was mounted with a label reading ‘Nagasaki | 1869 | ? imported in Rice –’. We have not been able to find any specimen of this species from Japan other than holotype. Hence, occurrence of this species in Japan seems to be questionable.
| 1 | Teeth of lateral margins of pronotum relatively short and of identical size | 2 |
| – | Teeth of lateral margins of pronotum relatively long. Posterior teeth longer than those of anterior margins | 6 |
| 2 | 1st antennomere longer than combined length of 2nd and 3rd antennomeres. Dorsolateral portions of pronotum impressed lightly | Psammoecus boreas sp. n. |
| – | 1st antennomere almost as long as or shorter than combined length of 2nd and 3rd antennomeres. Dorsolateral portions of pronotum with no impressions | 3 |
| 3 | No long seta on anterolateral margins of elytra. Body oblong | Psammoecus bipunctatus (Fabricius, 1792) |
| – | Some long setae on anterolateral margins of elytra | 4 |
| 4 | Distance between teeth of lateral margins of pronotum irregular | Psammoecus scitus sp. n. |
| – | Distance between teeth of lateral margins of pronotum regular | 5 |
| 5 | Parameres long. Incision of anterior margin shallow | Psammoecus fasciatus Reitter, 1874 |
| – | Parameres short. Incision of anterior margin deep | Psammoecus hiranoi Yoshida & Hirowatari, 2013 |
| 6 | Black elytra with yellow maculae | Psammoecus simoni Grouvelle, 1892 |
| – | Yellow elytra with black maculae or no maculae | 7 |
| 7 | Parameres fused with phallobase. Tooth IV of lateral margins of pronotum extended in a posterolateral direction. Antennae, especially 1st antennomere, long | Psammoecus omotoensis sp. n. |
| – | Parameres and phallobase divided. Tooth IV of lateral margins of pronotum extended in a roughly lateral direction. Antennae moderately long | 8 |
| 8 | Basal parameres large. Distance between posterior margin of phallobase and deepest point of incision of anterior margin broad. Rows of punctures on elytra comparatively narrow | Psammoecus trimaculatus Motschulsky, 1858 |
| – | Basal parameres comparatively narrow. Rows of punctures on elytra comparatively wide | 9 |
| 9 | Distance between posterior margin of phallobase and deepest point of incision of anterior margin broad. Wide basal portion of parameres square. Punctuation of pronotal disk moderately dense | Psammoecus labyrinthicus sp. n. |
| – | Distance between posterior margin of phallobase and deepest point of incision of anterior margin narrow. Wide basal portion of parameres triangular. Punctuation of pronotal disk comparatively dense | Psammoecus triguttatus Reitter, 1874 |

Psammoecus scitus sp. n., holotype, male. A Left antenna B head and pronotum of dorsal view C head and pronotum of ventral view D metaventrite E right elytron with rows of punctures and pubescence. Scale: 0.5 mm.

Psammoecus bipunctatus (Fabricius, 1792), male specimen which genital structures were illustrated in this paper. A Left antenna B head and pronotum of dorsal view C head and pronotum of ventral view D metaventrite E right elytron with rows of punctures and pubescence. Scale: 0.5 mm.

Psammoecus trimaculatus Motschulsky, 1858, male specimen which genital structures were illustrated in this paper. A Left antenna B head and pronotum of dorsal view C head and pronotum of ventral view D metaventrite E right elytron with rows of punctures and pubescence. Scale: 0.5 mm.

Psammoecus triguttatus Reitter, 1874, lectotype, male. A Left antenna B head and pronotum of dorsal view C head and pronotum of ventral view D metaventrite E right elytron with rows of punctures and pubescence. Scale: 0.5 mm.

Psammoecus labyrinthicus sp. n., holotype, male. A Left antenna B head and pronotum of dorsal view C head and pronotum of ventral view D metaventrite E right elytron with rows of punctures and pubescence. Scale: 0.5 mm.

Psammoecus boreas sp. n., holotype, male. A Left antenna B head and pronotum of dorsal view C head and pronotum of ventral view D metaventrite E right elytron with rows of punctures and pubescence. Scale: 0.5 mm.

Psammoecus omotoensis sp. n., holotype, male. A Right antenna B head and pronotum of dorsal view C head and pronotum of ventral view D metaventrite E left elytron with rows of punctures and pubescence. Scale: 0.5 mm.

Psammoecus simoni Grouvelle, 1892, male specimen which genital structures were illustrated in this paper (B–E) and another male specimen (A). A Left antenna B head and pronotum of dorsal view C head and pronotum of ventral view D metaventrite E right elytron with rows of punctures and pubescence. Scale: 0.5 mm.

Psammoecus fasciatus Reitter, 1874, lectotype, male. A Left antenna B head and pronotum of dorsal view C head and pronotum of ventral view D metaventrite E right elytron with rows of punctures and pubescence. Scale: 0.5 mm.

Psammoecus quadrimaculatus Reitter, 1874, lectotype, male. A Left antenna B head and pronotum of dorsal view C head and pronotum of ventral view D metaventrite E right elytron with rows of punctures and pubescence. Scale: 0.5 mm.
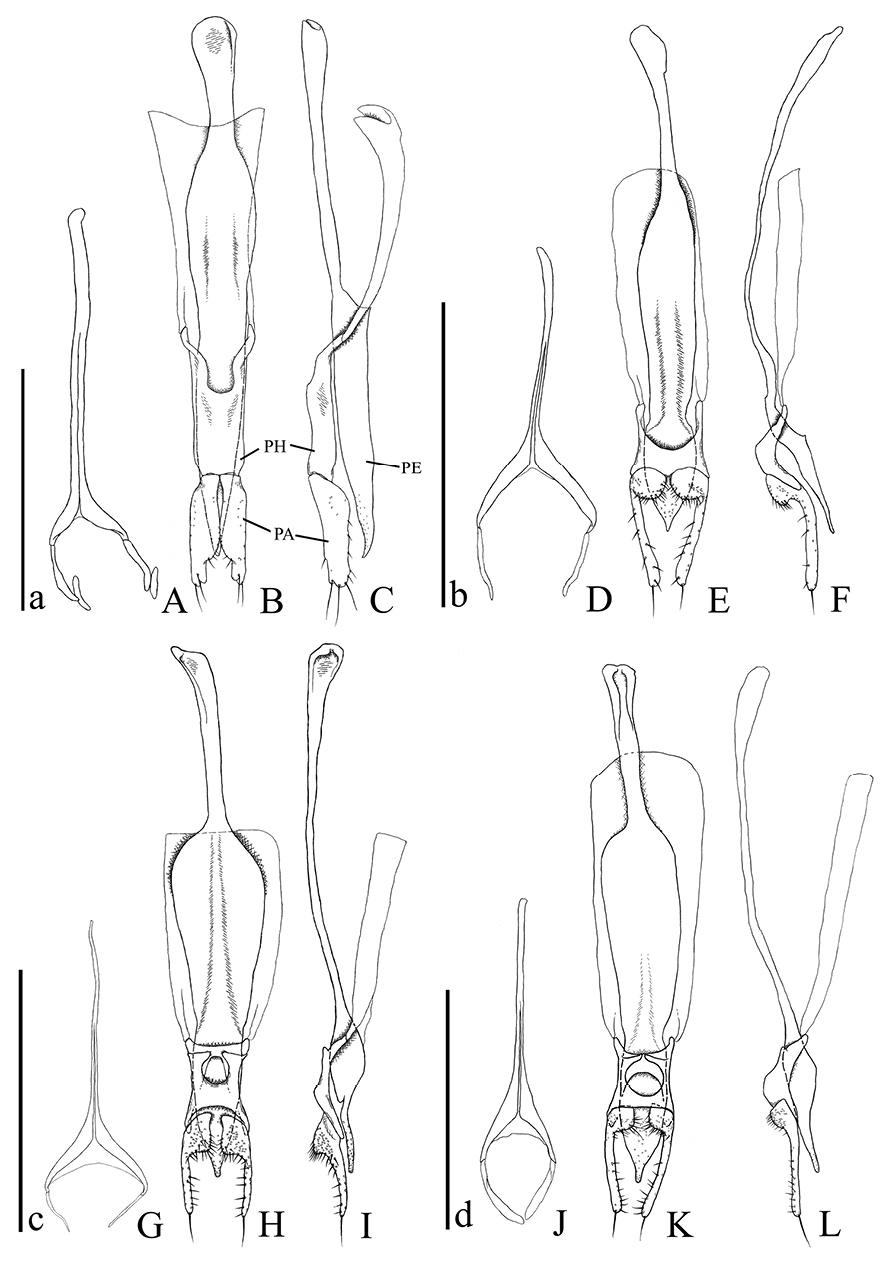
Male genital organs of Psammoecus spp. A–C Psammoecus scitus sp. n., holotype D–F Psammoecus bipunctatus (Fabricius, 1792) G–I Psammoecus trimaculatus Motschulsky, 1858 J–L Psammoecus triguttatus Reitter, 1874, lectotype. A, D, G and J 9th abdominal sternite B, C, E, F, H, I, K and L aedeagus in dorsal (B, E, H and K) and lateral (C, F, I and L). Abbreviations: PH—phallobase; PA—parameres; PE—penis. Scale: 0.5 mm; a for A–C; b for D–F; c for G–I; d for J–L.

Male genital organs of Psammoecus spp. A–C Psammoecus labyrinthicus sp. n., holotype D–F Psammoecus boreas sp. n., holotype G–I Psammoecus omotoensis sp. n., holotype J–L Psammoecus simoni Grouvelle, 1892 M–O Psammoecus fasciatus Reitter, 1874, lectotype P–R Psammoecus quadrimaculatus Reitter, 1874, holotype. A, D, G, J, M and P 9th abdominal sternite; B, C, E, F, H, I, K, L, N, O, Q and R aedeagus in dorsal (B, E, H, K, N and Q) and lateral (C, F, I, L, O and R). Scale: 0.5 mm; a for A–C; b for D–F; c for G–I; d for J–L; e for M–O; f for P–R.

The habitat of Psammoecus labyrinthicus sp. n. (A) and Psammoecus simoni Grouvelle, 1892 (B) A Ta, Toyotama-chô, Tsushima Island, Nagasaki Prefecture, Japan B Inamine, Nago City, Okinawa Island, Okinawa Prefecture, Japan.
We wish to express our cordial thanks to Prof. Minoru Ishii, Assoc. Prof. Norio Hirai and the members of the Entomological Laboratory (Osaka Prefecture University, Sakai) for their valuable advice and helpful support. Sir Anthony Galsworthy (BMNH) kindly corrected the language of the manuscript. We also thank the following entomologists for their kind advice and cooperation for our study and/or generous loan of numerous materials: Mr. Yukihiko Hirano (Odawara City, Kanagawa), Mr. Isamu Tanaka (Nishinomiya City, Hyogo), Prof. Masahiro Sakai and Assoc. Prof. Hiroyuki Yoshitomi (EUMJ), Dr. Roger Booth and Dr. Beulah Garner (BMNH), Dr. Masahiro Ôhara (SEHU), Mr. Shigehiko Shiyake (OMNH), Dr. Kiyoshi Matsuda and Mr. Ken Ikeuchi (Osaka Prefecture University, Sakai), Mr. Kôzô Mizuno (Uji City, Kyoto), Mr. Noriyuki Narukawa (Suzuka City, Mie), Dr. Jun-ichi Aoki (Minato-ku, Tokyo), Dr. Michael Karner (Senckenbergisches Naturforschendes Institut, Frankfurt am Main), Mr. Kyôhei Watanabe (Kanagawa Prefectural Museum of Natural History, Odawara), Mr. Yoshihiro Sawada (House Doctor, Ibaraki), Dr. Munetoshi Maruyama (Kyushu University Museum, Fukuoka) and the members of the Entomological Laboratory (ELKU).
This study was contributed from ELKU (Ser. 7, No. 7)