





Citation: Mercado-Salas NF, Suárez-Morales E (2014) On Kiefer’s American Eucyclops (Copepoda, Eucyclopinae): redescriptions and comments on the historical records of E. delachauxi, E. prionophorus, E. bondi and E. leptacanthus. ZooKeys 402: 1–41. doi: 10.3897/zookeys.402.6112
The freshwater copepod genus Eucyclops contains many supposedly cosmopolitan species whose taxonomic status is still under discussion; some of them represent species complexes. The problem is not exclusive to these widespread species; there are several American Eucyclops needing a taxonomic re-evaluation. Based on the examination of Friedrich Kiefer’s collection in Karlsruhe, Germany, the type specimens of four American species of Eucyclops (E. delachauxi (Kiefer, 1926), E. prionophorus Kiefer, 1931, E. bondi Kiefer, 1934, E. leptacanthus Kiefer, 1956) were re-examined and redescribed using upgraded descriptive standards. Kiefer’s translated descriptions and unpublished original drawings of these species are also presented. Characters like the ornamentation of the antennal basis, ornamentation of intercoxal sclerites of the swimming legs 1–4, length of basipodal seta of leg 1, ornamentation of caudal rami, the presence of aesthetascs and modified setae on the antennules in male, and the structure of the male sixth leg are compared herein to aid a more accurate separation of these American species. A revision of the American records of these species confirms that some are likely to refer to undescribed species. Overall, the diversity of the American Eucyclops appears to be underestimated and certainly deserves further study.
Freshwater copepods, Eucyclopinae, morphology, systematics, taxonomy
The freshwater genus Eucyclops Claus, 1893 is currently known to contain more than 108 species and subspecies (
The taxonomic problems within this taxon started with the incomplete description of the type species of the genus: Eucyclops serrulatus (Fisher, 1851) from Russia (
In a project to explore the species diversity of the genus Eucyclops in Mexico, the type material of some of these species was examined. There are four species described by Kiefer, which have been recorded in Mexico: Eucyclops delachauxi (Kiefer, 1925), Eucyclops prionophorus Kiefer, 1931, Eucyclops bondi Kiefer, 1934 and Eucyclops leptacanthus Kiefer, 1956 (
In order to provide an upgraded morphological redescription of Eucyclops delachauxi, Eucyclops prionophorus, Eucyclops bondi, and Eucyclops leptacanthus, we examined the type material of Kiefer’s collection deposited at the Staatliches Museum für Naturkunde Karlsruhe (Germany). Drawings were made at 1000× with a Zeiss Axioskop 2 plus compound microscope equipped with a camera lucida. Mapping of rows of spinules and setules on the antennal basis and on the coxopodite and intercoxal slerite of P4 followed
For each of these four species we present first the complete translation from German to English of Kiefer’s description, followed by an upgraded description based on our personal observations on Kiefer’s material. Characters or structures not observed but previously published by other researchers are included in the descriptions with its reference. Figures mentioned in the translated text correspond to the numbers of the figures in the original descriptions published by
Figs 1–7
Cyclops delachauxi (Kiefer, 1925)
Cyclops delachauxi (Kiefer, 1926)
Eucyclops (Eucyclops) delachauxi Kiefer, 1929
Cyclops Delachauxi Kiefer, 1925
Cyclops Delachauxi Kiefer, 1926
Eucyclops (Eucyclops) Delachauxi, Kiefer, 1929
Eucyclops Delachauxi Kiefer, 1943
Eucyclops delachauxi Lindberg, 1955, 1957
Kiefer’s description.
a) The female: the general appearance as Cyclops serrulatus. Fifth segment of cephalothorax with lateral hair-setae. Last abdominal segment longer than the previous one. Caudal rami parallel, relatively short, about four times as long as wide; outer edge with serra, this is formed by a small number of (4–10) rather long, slender spinules. Inner edge naked (Fig. 1). Of the four setae on the end (apical), only the two median setae are strongly developed, the longest is approximately twice the length of the other, with fine plumage, almost the entire length is homogeneous, the innermost apical hair-like seta is as long as or slightly longer than the outermost seta, more similar to a spine. First antenna twelve–segmented, reaching only a little above of the posterior margin of the first segment of cephalothorax; the last three segments with a narrow hyaline membrane; the seta of the last member originates in the middle of the edge. The branches of all swimming legs with three segments. The terminal segment of the endopod of fourth leg is, usually, exactly twice as long as wide and its two setae on the inner margin and single seta on the outer margin are formed normally. Of the two apical spines, the innermost is strongly curved outside and it is longer than the segment, the outermost is just as long as the segment (Fig. 4). The rudimentary leg is a monomial plate; the inner edge is slightly distended, of the three elements the medial is significantly longer than the other two, which are approximately equal in length, the inner spine is, at its insertion, about twice as wide as one of the two setae (Fig. 2). The seminal receptacle was not correctly identified in the preserved animals (Fig. 1). Total length about 950 µm.
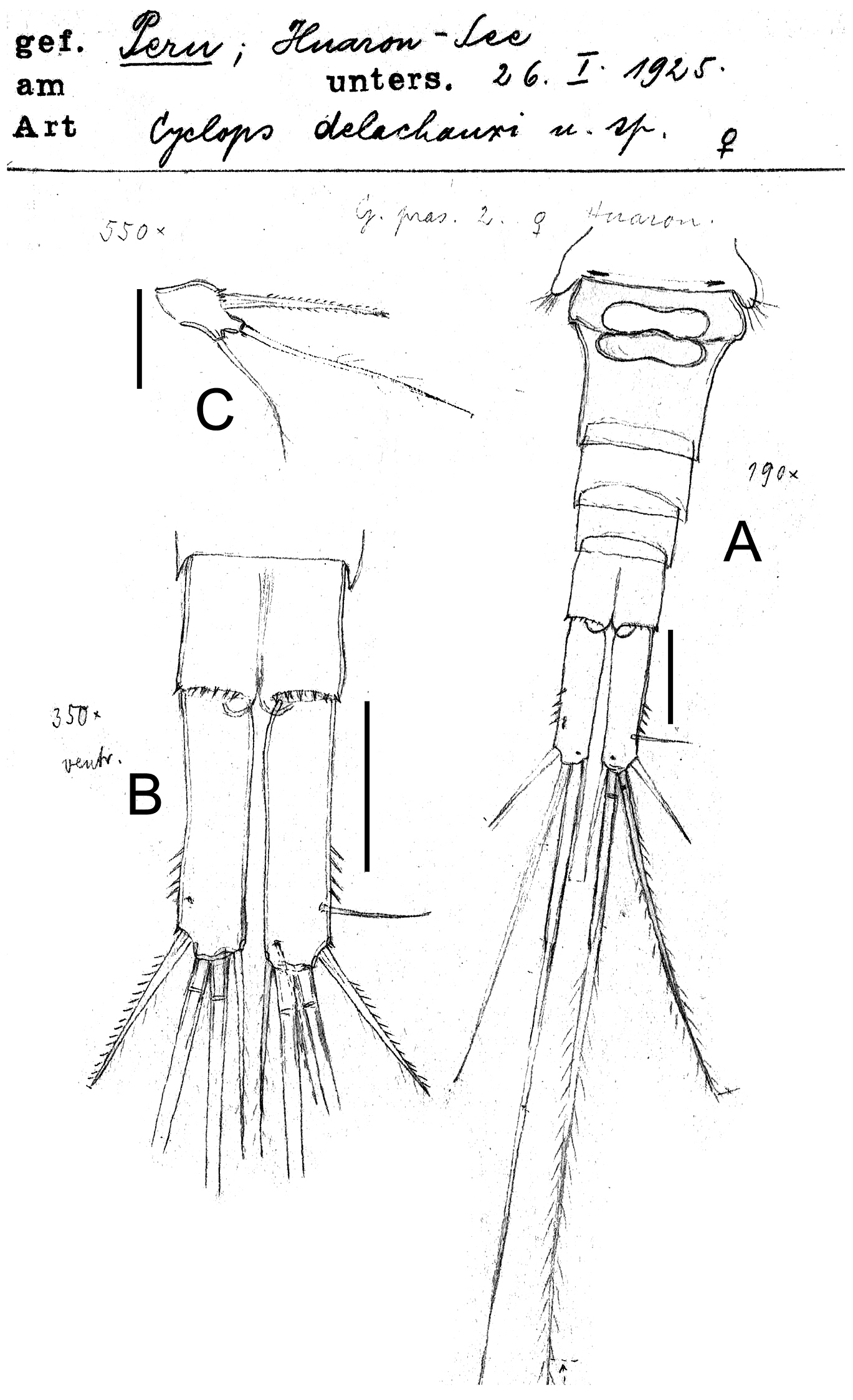
Eucyclops delachauxi (Kiefer, 1925). Original drawings of F. Kiefer. Female Holotype from Huaron, Peru. A Urosome, ventral view B Caudal rami C P5. Scale bars: A–B = 50 µm, C = 20 µm.
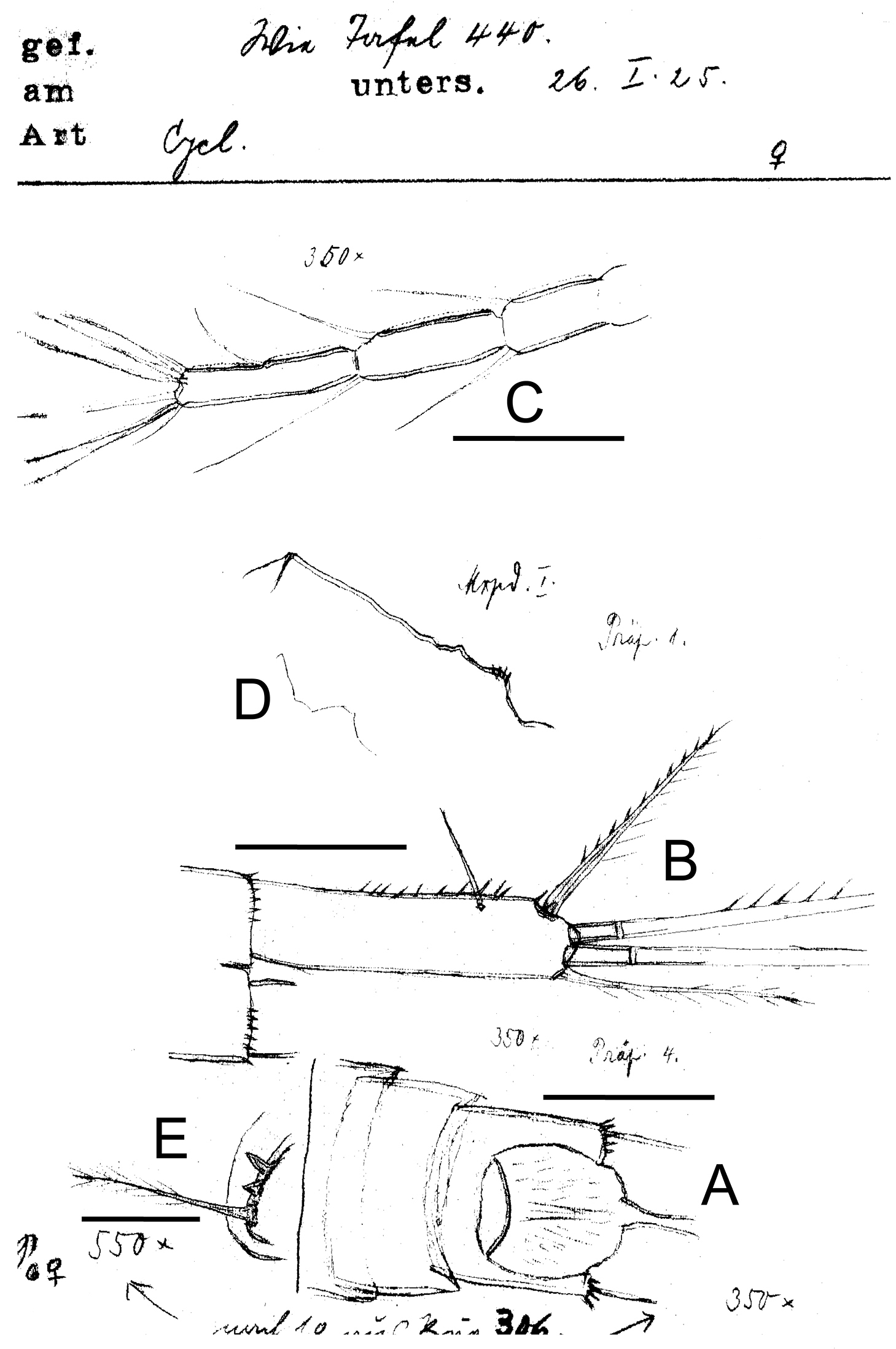
Eucyclops delachauxi (Kiefer, 1925). Original drawings of F. Kiefer. Female Holotype from Huaron, Peru. A Anal somite, dorsal view B Caudal rami C Antennule, segments 10–12 D Praecoxa of maxilla E P6. Scale bars: A–D = 50 µm, E = 20 µm.
b) The male: slender and slightly smaller than the female. Last abdominal segment also noticeably longer than the previous one, its posterior margin provided on ventral side with only very few spinules, as well as in the female. Caudal rami parallel, also four times as long as wide. The serra is missing on the outer edge. Ratios and plumage of the two middle apical setae as in the female. Of the two short terminal setae in the male the inner seems to be always longer than the outer (Fig. 3). The final segment of the endopod of the fourth pair of legs with its setae and apical spines as in the female (Fig. 4); the fifth leg as well. The shape of the genital valve reinforcement is best seen in the figure (Fig. 5).
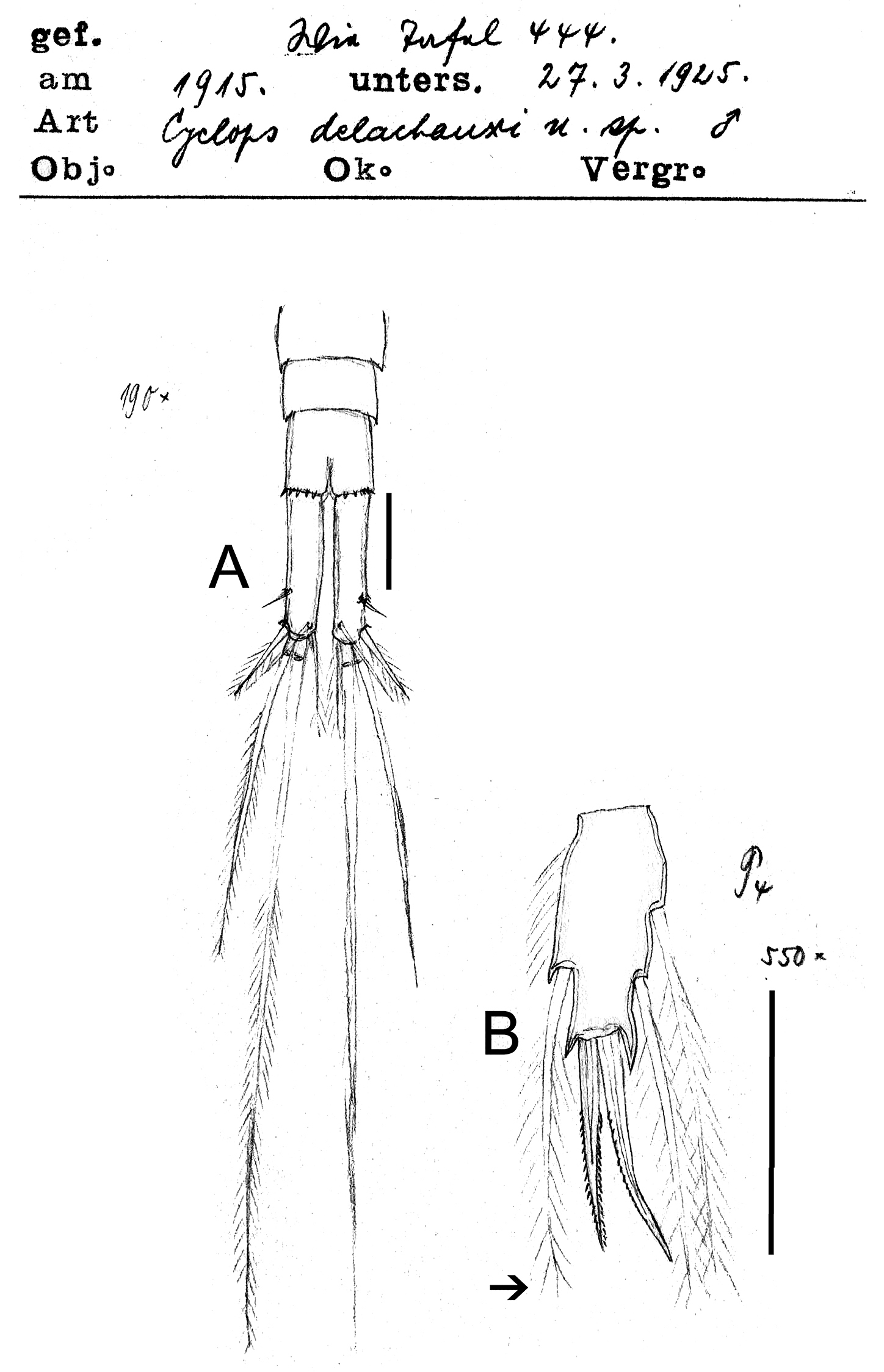
Eucyclops delachauxi (Kiefer, 1925). Original drawings of F. Kiefer. Paratype from Huaron, Peru. A Last urosomites and furca B Enp3P4. Scale bars: A–B = 50 µm.
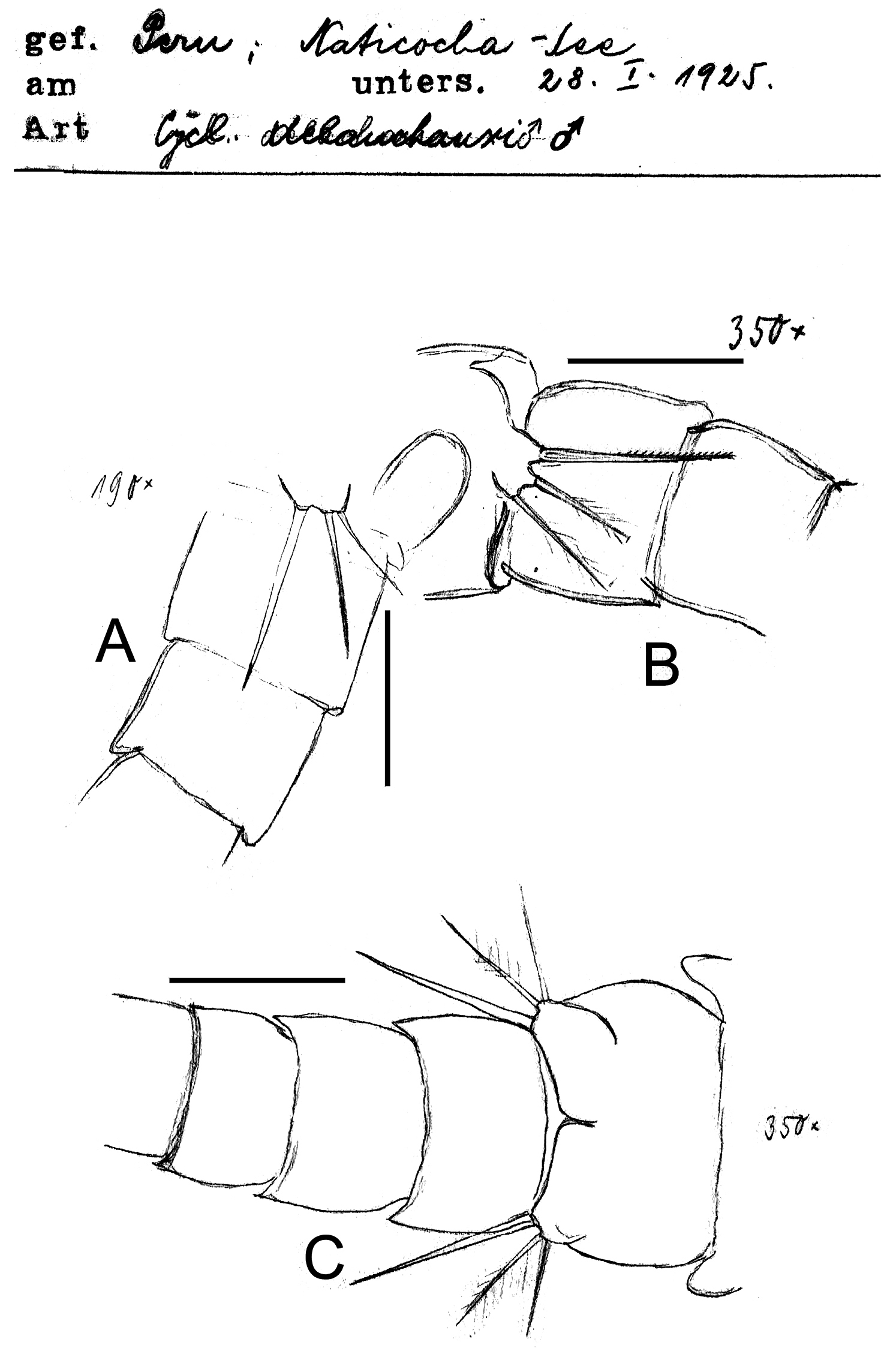
Eucyclops delachauxi (Kiefer, 1925). Original drawings of F. Kiefer. Male Paratype from Naticocha, Peru. A–C P6 and urosomites, ventral. Scale bars: A–C = 50 µm.
Eucyclops delachauxi (Kiefer, 1925). Author’s drawings. Female Holotype from Huaron, Peru. A Urosome, ventral view B Caudal ramus C Antennule D Antenna E Mandible (part). Scale bars: A = 100 µm; B–C = 50 µm; D–E = 20 µm.
This Cyclops, of the numerous “serrulatus-like” that I know cannot be identified as one, it comes from two closely located lakes in the Andes of Peru (Huaron and Naticocha, 5140 m high) and has been collected by Ing. E. Godet in 1915. It is named after Dr. Th. Delachaux, Neuenburg, which I am indebted for the provision of his Cyclopoida material. The above description must be regarded as provisional. A more detail, equipped with longer pictures in the description of the Cyclops-forms from the mentioned lakes will be published in Archives of Hydrobiology.
Description based on Kiefer’s material.
Material examined. Holotype. Adult ♀ From Huaron, central highlands of Peru, specimen dissected (slide reference numbers SMNK00248, SMNK00249, SMNK00250). Additional material (adult ♂) from Lake Naticocha, Peru (slide reference number SMNK00253). Both Lake Huaron and Naticocha 5140 m high, samples collected by Ing. E. Godet in 1915. Deposited at the Staatliches Museum für Naturkunde Karlsruhe, Germany.
Female: Average length excluding caudal setae 950 µm. Five-segmented urosome (Figs 1A; 5A), relatively elongated; posterior margin of anal somite with one row of strong spinules. Genital double-somite symmetrical. Seminal receptacle typical of serrulatus-group, with rounded lateral arms. Genital double-somite 1.3 times as long as wide. Anal somite with a group of spinules at each side of anal opening, anal operculum rounded (see Fig. 2A). Length/width ratio of caudal ramus = 3.5–4.4; inner margin of caudal ramus naked. Serra with strong spinules covering 19.4–43% of outer margin, spinules about the same size (Figs 2B, 5B). Dorsal seta (VII) long: 0.6 times the length of caudal ramus and 0.9-1.0 times as long as outermost caudal seta (III). Ratio of innermost caudal seta (VI)/outermost caudal seta (III) = 1.1. Lateral caudal seta (II) inserted at 63–72% of caudal rami. All terminal caudal setae plumose.
Antennule (Figs 2C, 5C): 12-segmented. Armament per segment as follows (s = seta, ae = aesthetasc, sp = spine): 1(5s), 2(3s), 3(2s), 4(5s), 5(2s), 6 (1s+1sp), 7(0s), 8(3s), 9(2s), 10(2s), 11(2s), 12(8s). Numbers in bold face indicate segments with incomplete ornamentation.
Antenna (Fig. 5D): Basis (2s + Exp), and 3-segmented Enp (1s, 8s and 6s). Basis ornamentation as follows (sensu
Labrum, Maxillule, and Maxilliped not observable in the slides.
Mandible (Fig. 5E): with 6 tooth on gnathobase. Innermost margin with 1 spinulose seta.
Maxilla (Fig. 2D): Precoxa with row of small spinules on dorsal surface.
P1-P4: Endopod and exopods of all swimming legs 3-segmented. Armature formula as in Table 1.
Setation formula of the swimming legs in the types (females and males) of the four Eucyclops species here studied; (spines in Roman numerals, setae in Arabic numerals). (–) represents structures not observed on the type material.
| Species | Coxa | Basis | Exp | Enp | |
|---|---|---|---|---|---|
| Eucyclops delachauxi | P1 P2 P3 P4 |
0-1 - 0-1 - |
1-I - 1-0 - |
I-1; I-1; III-5 -, -, - I-1; I-1; IV-5 -, -, - |
0-1; 0-21-I-4 -, -, - 0-1; 0-2; 1-I-4 -, -, 1-II-2 |
| Eucyclops prionophorus | P1 P2 P3 P4 |
0-1 - 0-1 0-1 |
1-I - 1-0 1-0 |
I-1, I-1, III-5 -, -, - I-1, I-1, IV-5 I-1, I-1, III-5 |
0-1, 0-2 1-I-4 -, -, - 0-1, 0-2, 1-I-4 0-1, 0-2, 1-II-2 |
| Eucyclops bondi | P1 P2 P3 P4 |
0-1 0-1 0-1 0-1 |
1-I 1-0 1-0 1-0 |
I-1, I-1, III-5 I-1, I-1, IV-5 I-1, I-1, IV-5 I-1, I-1, III-5 |
0-1, 0-2, 1-I-4 0-1, 0-2, 1-I-4 0-1, 0-2, 1-I-4 0-1, 0-2, 1-II-2 |
| Eucyclops leptacanthus | P1 P2 P3 P4 |
0-1 0-1 0-1 - |
1-I 1-0 1-0 - |
I-1, I-1, III-5 I-1, I-1, IV-5 I-1, I-1, IV-5 -, -, - |
0-1, 0-2, 1-I-4 0-1, 0-2, 1-I-4 0-1, 0-2, 1-I-4 -, -, 1-II-2 |
Leg 1 (Fig. 6A): Group of small hairs present in each side on anterior surface of intercoxal sclerite, distal margin with 2 rounded chitinized projections. Basipodal spine reaching middle length of Enp3, 0.8 times as long as Enp.
Eucyclops delachauxi (Kiefer, 1925). Author’s drawings. Female Holotype from Huaron, Peru. A P1 B P2 C–D P3. Scale bars: A–D = 50 µm.
Leg 2 and 3: General shape as in Fig. 6B–D.
Leg 4 (Fig. 3B): Intercoxal sclerite with short hairs in rows I, II, III. Coxopodite with row A, B (3-4), C+D (21), E (5), F and H (from
Leg 5 (Fig. 1C): One free segment subrectangular, 1.4–1.8 times as long as wide, bearing 1 inner spine and 2 setae; medial seta about 1.7 times longer than outer seta. Inner spine 1.2–1.6 times longer than outer seta and 0.5–0.8 times as long as median seta. Inner spine 2.0–2.2 times as long as segment length.
Leg 6 (Fig. 2E): Represented by small, flat plate with 1 slender and long seta and 2 small spines. Outer seta notably long, 12 times longer than medial spine and 6 times longer than inner spine.
Male: Urosome 6-segmented, posterior margin of urosomites serrated (Figs 4, 7A). Caudal ramus rectangular, 2.8–4.0 times as long as wide; inner margin of caudal ramus naked. Dorsal seta (VII) 0.6 times as long as caudal ramus, and 1.1 times as long as outermost caudal seta (III). Length ratio of innermost caudal seta (VI)/outermost caudal seta (III) = 1.2. Lateral caudal seta (II) inserted at 70% of caudal rami length. All terminal caudal setae plumose.
Eucyclops delachauxi (Kiefer, 1925). Author’s drawings. Male Paratype from Huaron, Peru. A Urosome, ventral view B Antennule. Scale bars: A–B = 50 µm.
Antennule (Fig. 7B): ornamentation per segment incomplete to described in details.
Leg 5 (Fig. 7A): segment subrectangular in shape, 1.5 times longer than wide, bearing 1 inner spine and 2 setae; medial seta about 1.4 times longer than outer seta. Inner spine 0.8 times longer than outer seta and 0.6 times as long as medial seta.
Leg 6 (Figs 4, 7A): Represented by one small, flat plate placed near lateral margin of genital somite with 1 strong, long inner spine and 2 unequal setae. Inner spine reaching medial length of fourth urosomite. Inner spine about 1.3–1.8 times longer than medial seta and about 1.4–2.0 times longer than outer seta.
Remarks. In the publication posterior to the description of Eucyclops delachauxi made by
Figs 8–13
Eucyclops (s. str.) prionophorus Kiefer, 1931
Eucyclops prionophorus, Yeatman, in: Edmonson 1959, Smith and Fernando 1977, 1978, Harris 1978)
Eucyclops prionophorus,
Kiefer’s description. The female: Caudal rami slender, four times as long as wide, row of spinules on the outer margin, distal spinules are longer and proximal spinules are slender (Fig. 2). First antenna twelve-segmented, slightly shorter than the cephalothorax, with a narrow hyaline membrane along the margin of last three segments. Spine formula of exopods of four swimming legs is 3443. Fifth leg with one segment, bearing 3 elements, of which the inner spine at its insertion is twice as wide as both other setae. Seminal receptacle is similar to that in Eucyclops serrulatus. Length excluding caudal setae 0.94 mm.
The male: Spine of genital plate (P6) extremely long, longer than the genital segment, with two short, slender, plumose setae. Body length excluding caudal setae 0.8 mm.
Distribution. North America, close to New Heaven.
Description based on Kiefer’s material.
Material examined. Holotype. Adult ♀ collected 05.05.1929 from New Haven, USA. Specimen dissected (slide number SMNK01508). Additional material from San Bernardino, Paraguay (slides numbers SMNK03103, SMNK03104). Staatliches Museum für Naturkunde Karlsruhe, Germany.
Female: (Unless otherwise stated the character states are same in the holotype and in the Paraguay specimen) Average length excluding caudal setae 940 µm. Urosome 5-segmented (Fig. 11A), relatively elongated; urosomal fringes smooth or slightly serrated, posterior margin of anal somite with 1 row of relatively long spinules. Genital double-somite symmetrical (Fig. 9A). Seminal receptacle typical of the serrulatus-group, with rounded lateral arms on posterior margin. Genital double-somite slightly wider than long (about 1.1 times). Anal somite with one row of hair-like spinules in each side of anal opening, anal operculum slightly rounded (Figs 10C, 11B). Length/width ratio of caudal ramus = 4.0–4.5; inner margin of caudal ramus naked in specimens from New Haven and with groups of small spinules in specimens from San Bernardino (see Fig. 11A). Serra on outer margin with strong spinules covering 65–68% of lateral margin, spinules increasing in size distally (Figs 8A, 10C, 11B). Dorsal seta (VII) short, 0.4–0.6 times the length of caudal ramus, and 0.5–0.6 times as long as outermost caudal seta (III). Length ratio of innermost caudal seta (VI)/outermost caudal seta (III) =1.0–1.1. Lateral caudal seta (II) inserted at 75–77% of caudal ramus. All terminal caudal setae plumose.
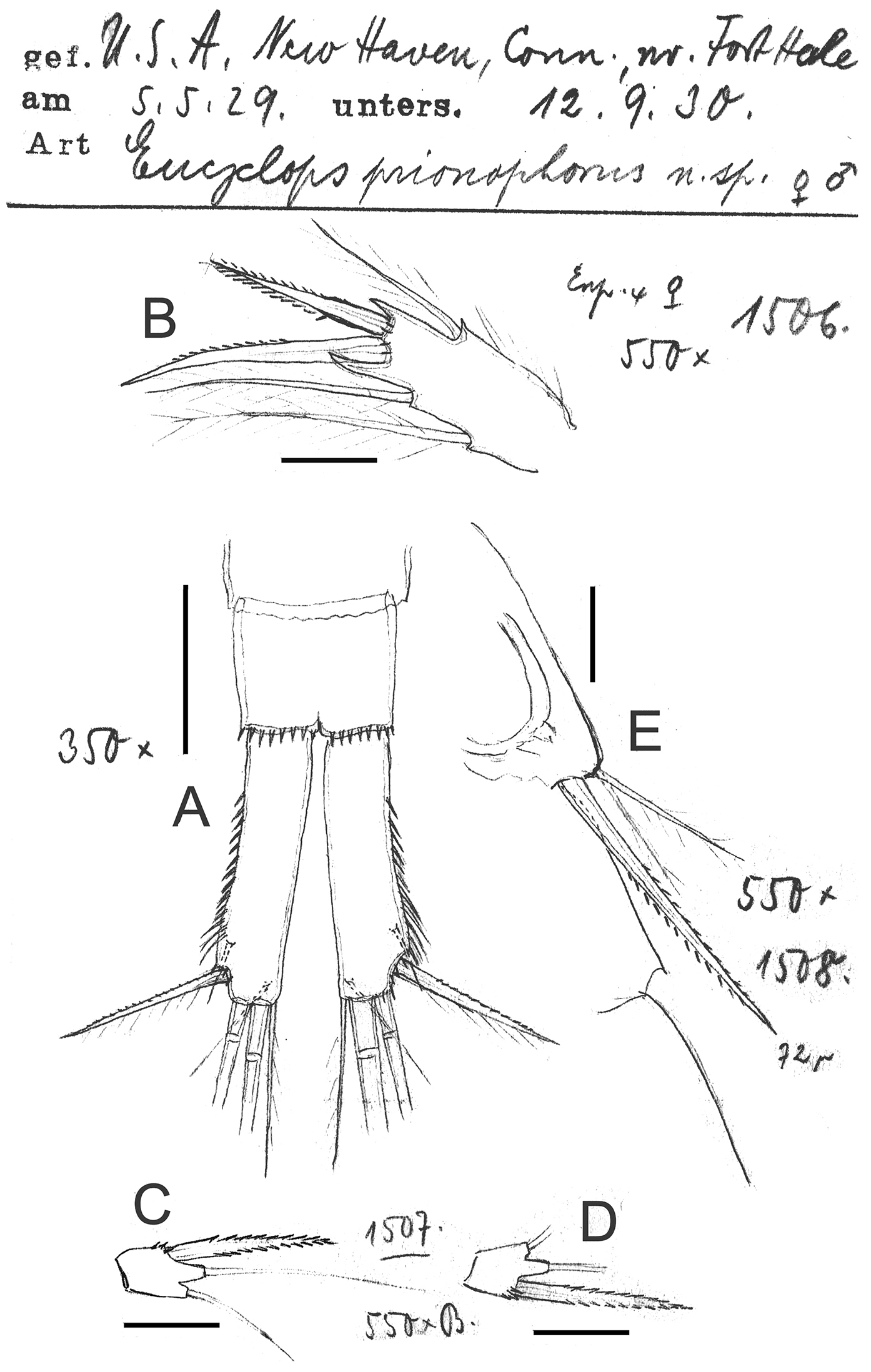
Eucyclops prionophorus Kiefer, 1931. Original drawings of F. Kiefer. A Female Holotype B female Paratype A, C–E from New Haven, U.S.A. A Caudal rami B Enp3P4 C–D P5 E P6. Scale bars: A = 50 µm; B–E = 20 µm.
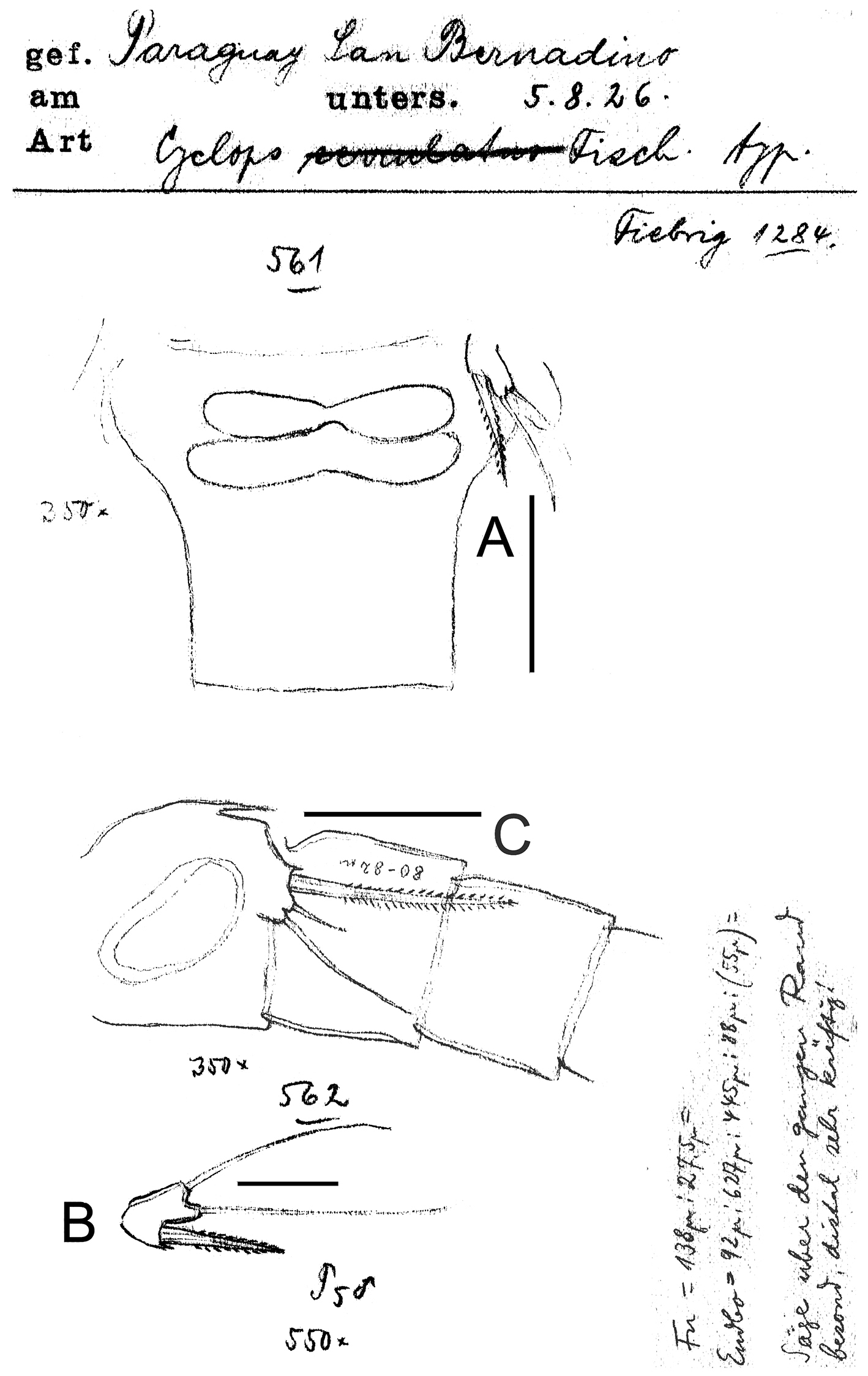
Eucyclops prionophorus Kiefer, 1931. Original drawings of F. Kiefer. Female A Male B–C from San Bernardino, Paraguay. A Genital double-somite and P5 B P5 C P6. Scale bars: A, C = 50 µm, B = 20 µm.
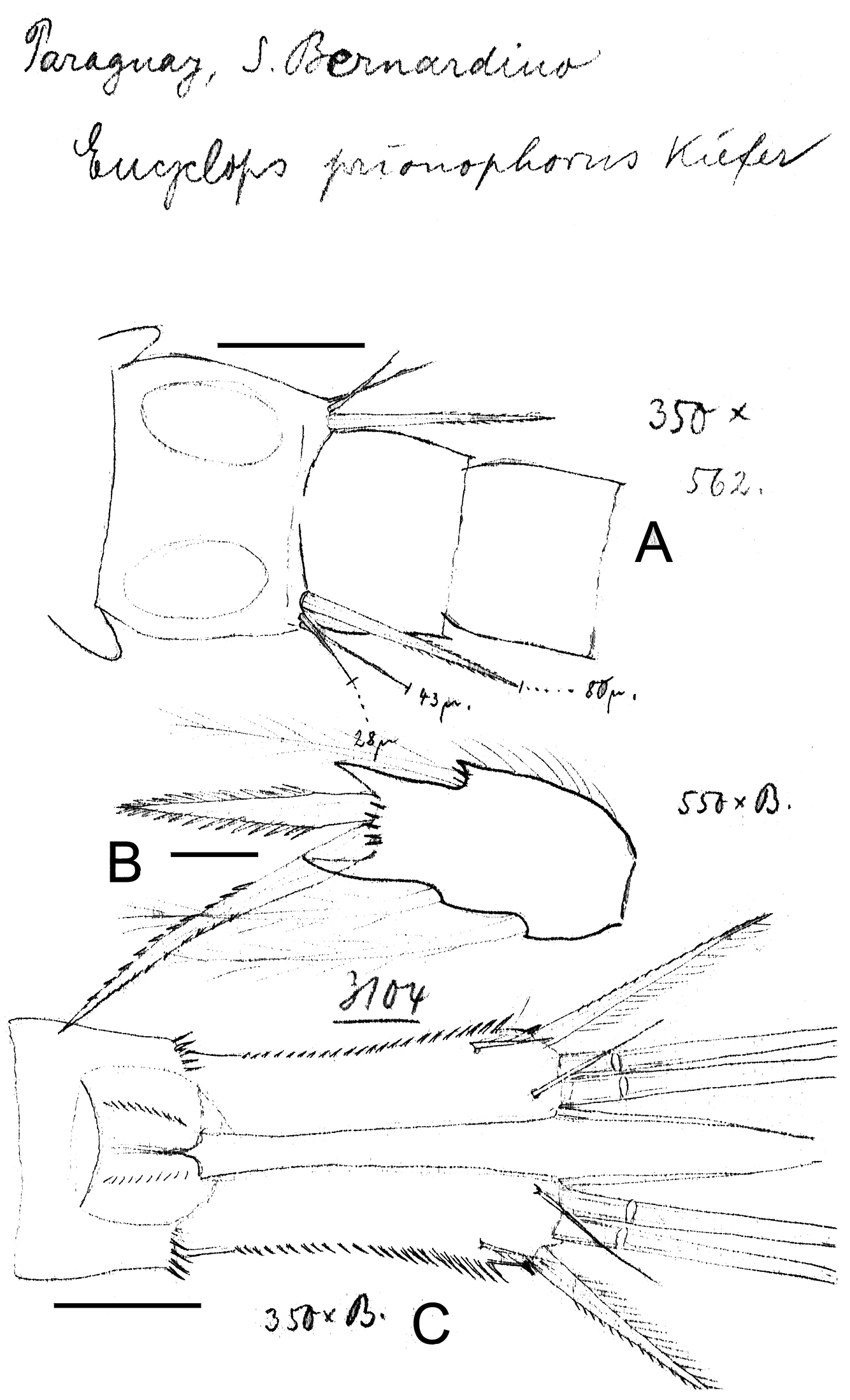
Eucyclops prionophorus Kiefer, 1931. Original drawings of F. Kiefer. Female B–C Male A from San Bernardino, Paraguay. A Genital somite and P6 B Enp3P4 C Anal somite and caudal rami. Scale bars: A–C = 50 µm.
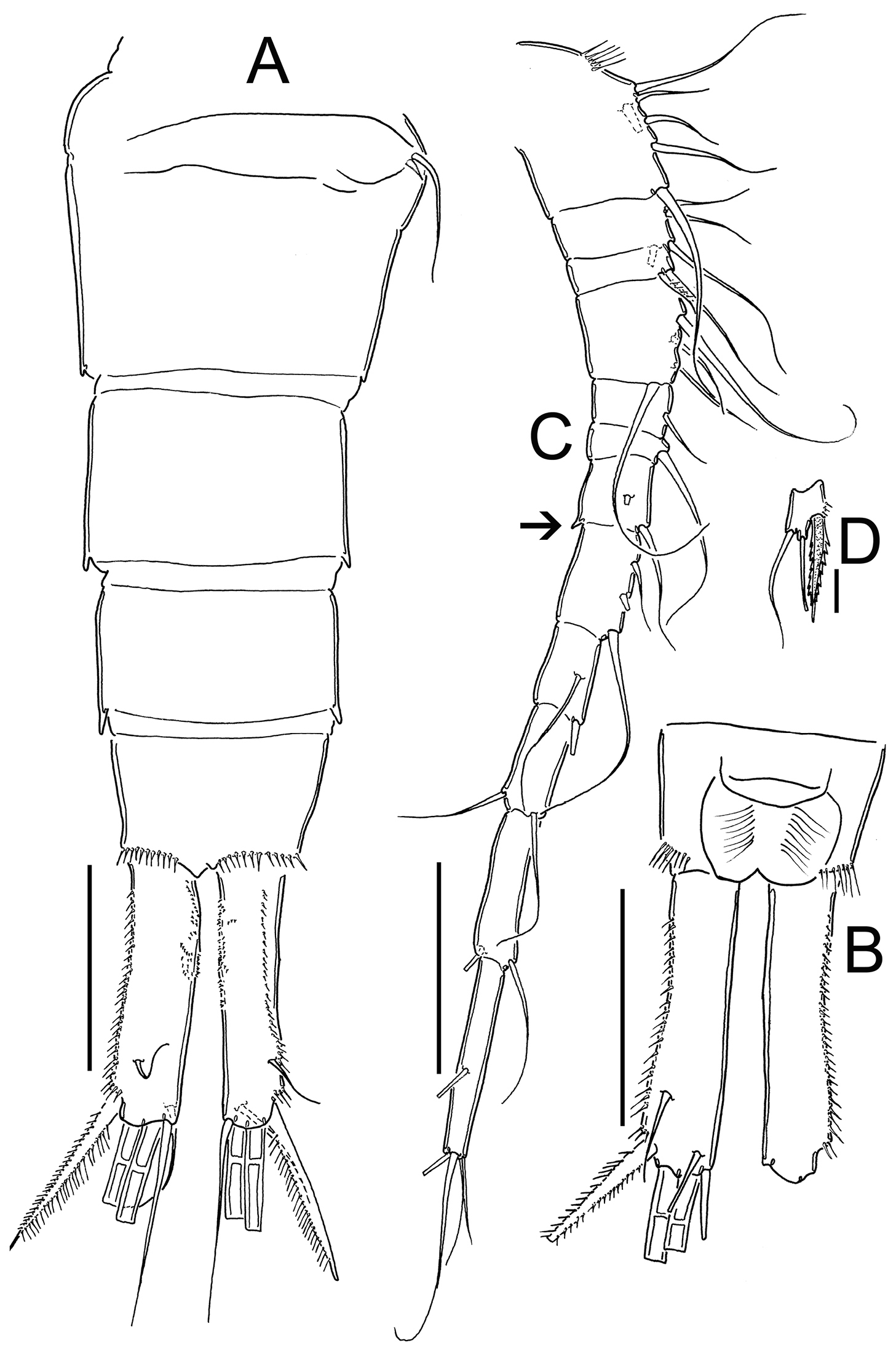
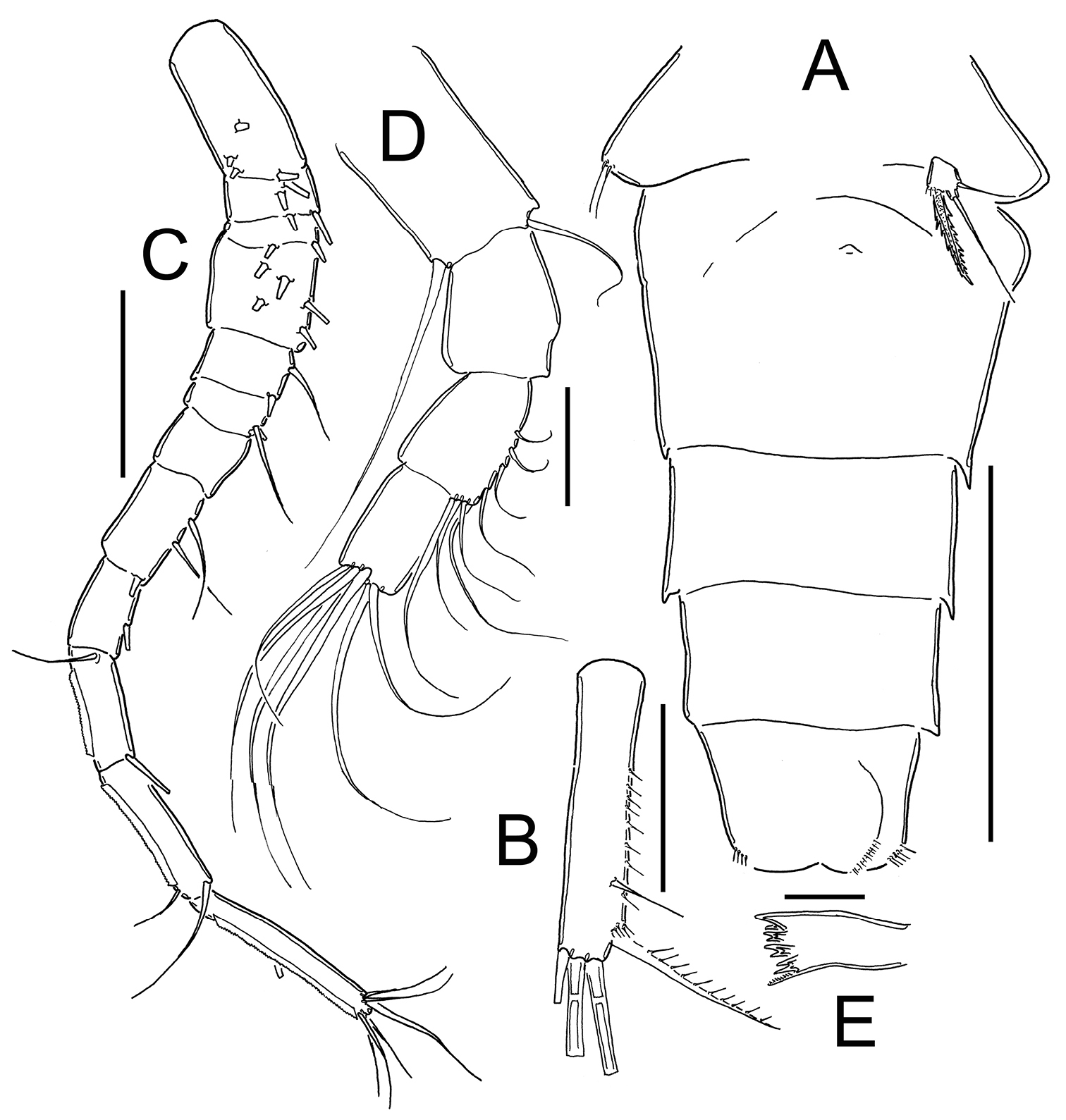
Eucyclops prionophorus Kiefer, 1931. Authors’ drawings. Female from San Bernardino, Paraguay. A Urosome, dorsal view B Anal somite and caudal rami C Antennule D P5. Scale bars: A–C = 100 µm, D = 20 µm.
Antennule (Fig. 11C): 12-segmented. Armament per segment as follows (s = seta, ae = aesthetasc, sp = spine): 1(6s), 2(4s), 3(2s), 4(4s), 5(1s), 6 (1s), 7(2s, with small projection on inner margin, arrowed in Fig. 11C), 8(3s), 9(2s), 10(2s), 11(2s), 12(5s). Numbers in bold indicate segments with incomplete ornamentation.
Antenna, mouthparts and Leg 2: not observable in slides.
P1-P4: Endopods and exopods of all swimming legs 3-segmented. Armature formula of all swimming legs as in Table 1.
Leg 1 (Fig. 12A): New Haven: Intercoxal sclerite without ornamentation and with 2 rounded chitinized projections. Coxa with strong biserially setulated inner coxal seta. Basipodal spine not reaching middle length of Enp 3; basipodal spine 0.6 times as long as total length of Enp. Third segment of Enp 1.5 times as long as wide, apical spine of Enp3 1.4 times longer than segment, apical most seta of Enp3 1.2 times longer than apical spine. Spines of Exp slightly elongated. San Bernardino: intercoxal sclerite not available. Basipodal spine reaching beyond middle length of Enp3; basipodal spine 0.75 times as long as total length of Enp. Third segment of Enp 1.2 times as long as wide, apical spine of Enp3 1.2 times longer than segment; apicalmost seta of Enp3 1.6 times longer than apical spine.
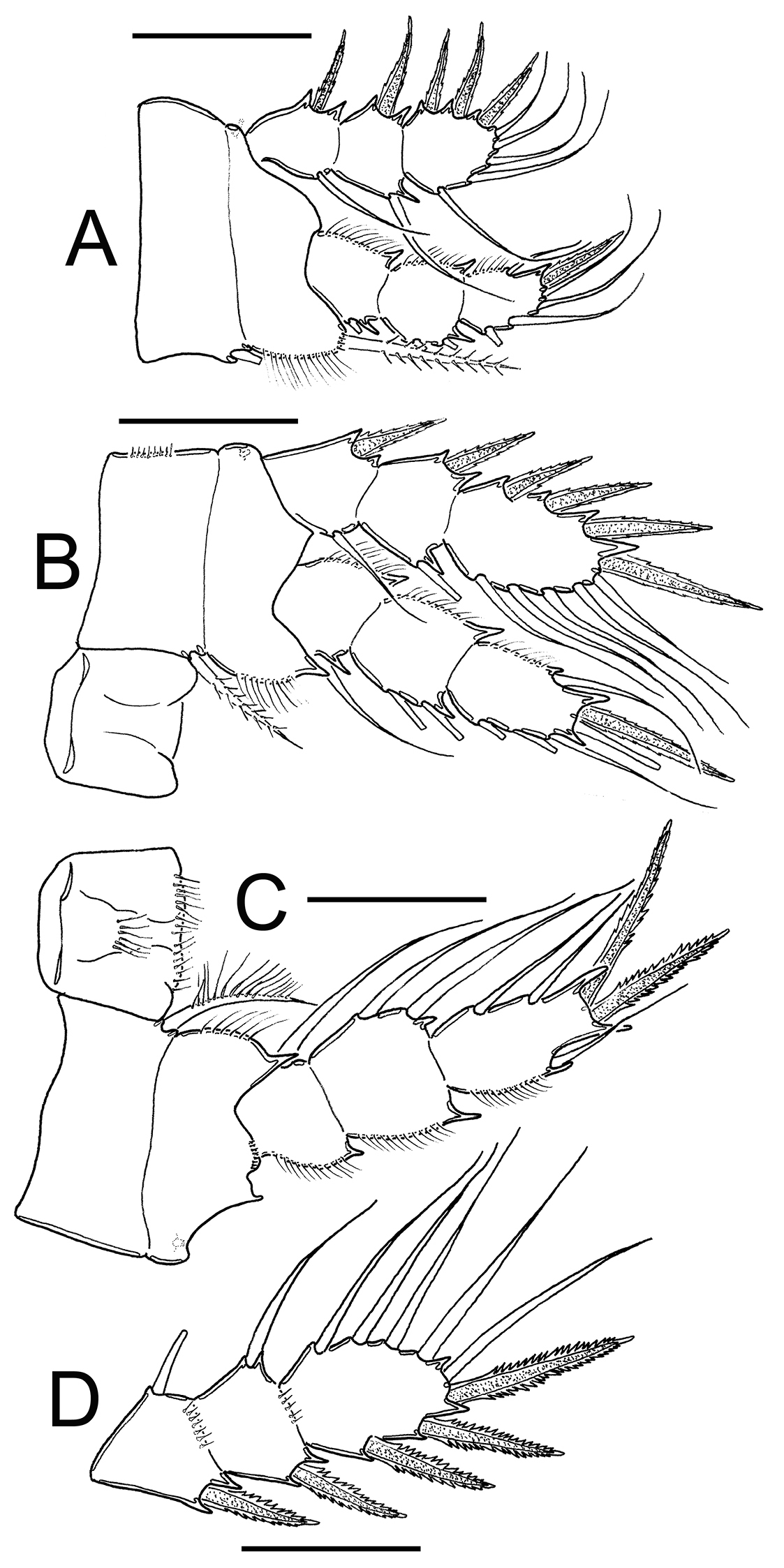
Eucyclops prionophorus Kiefer, 1931. Authors’ drawings. Female from San Bernardino, Paraguay. A P1 B P3 C P4 D Exp P4. Scale bars: A–D = 50 µm.
Leg 3 (Fig. 12B): No ornamentation observed on intercoxal sclerite, distal margin with 2 rounded projections. Coxa with strong, biserially setulated inner coxal seta. Coxa with row of tiny spinules along outer margin. Enp3 1.7–1.8 times as long as wide, apical spine on Enp3 1.2 times as long as segment, Exp3 1.5–1.7 times as long as wide, apicalmost spine of Exp3 1.1 times as long as segment.
Leg 4 (Fig. 12C–D): Intercoxal sclerite with rows I, II and III. Row I with 7 long spinules in each side and a small gap between. Row II with 6 spinules on middle margin. Row III divided in 3 sections, first one with 3 short spinules, middle section with 2 short spinules and third section with 2–3 short spinules (all observed in San Bernardino’s specimens). Caudal coxal surface with spinule formula: A, B (4), C+D (6) (sensu
Leg 5 (Figs 8C–D, 9A, 11D): Free segment subrectangular, 1.4–1.8 times longer than wide, bearing 1 inner spine and 2 setae; median seta longer than outer seta (1.0–1.7 times) and 1.3–1.6 times times longer than inner spine. Inner spine 1.7–2.0 times as long as segment.
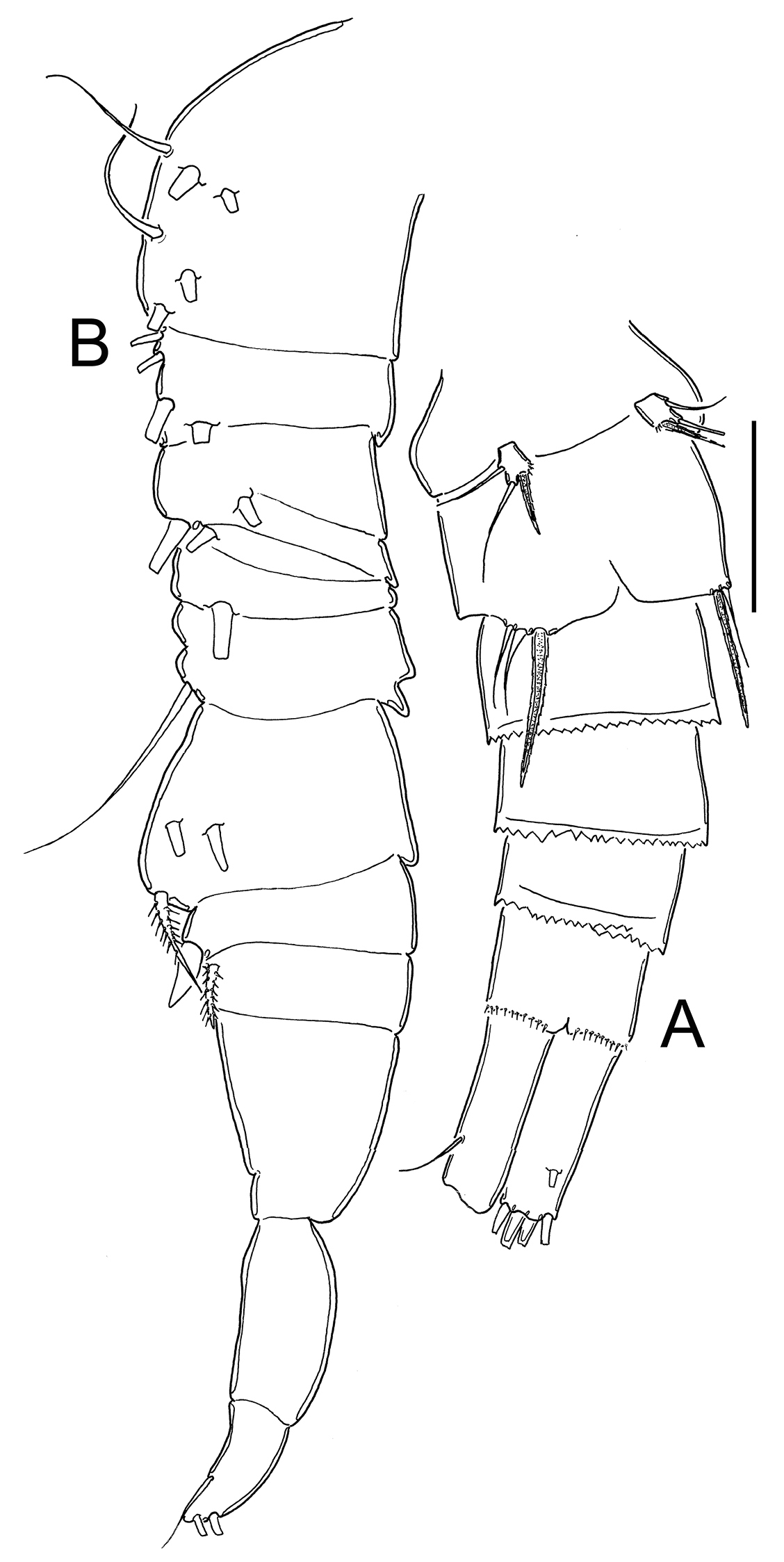
Male: Average length excluding caudal setae 800 µm. Urosome 6-segmented, posterior margin of urosomites smooth. Caudal ramus 3.5 times as long as wide, inner margin naked. with a group of spinules present at insertion of lateral seta. Ratio of innermost caudal seta (VI)/outermost caudal seta (III) = 1.6. All terminal caudal setae plumose.
Antennule: 1-segmented (Fig. 13B), armament per segment as follows (s = seta, modified seta = ms, ae = aesthetasc, sp = spine): 1(6s + 2ms + 1ae); 2(42 + 1ms); 3(2s+3ms +1ae); 4(1s); 5(1s); 6(0); 7(1s); 8(2s); 9(1s +1sp); 10(0); 11(0); 12(0); 13(2s); 14 (4s).

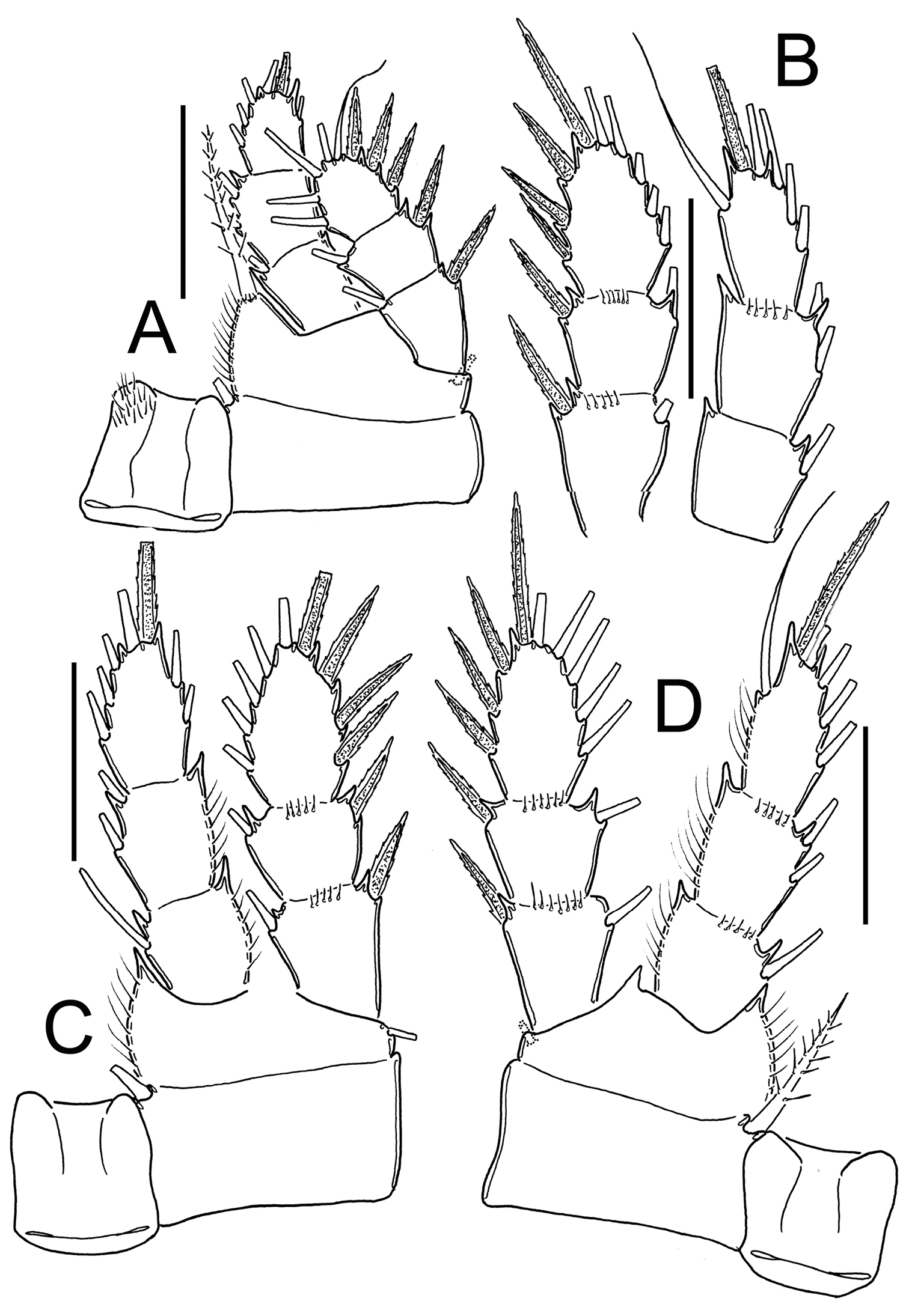
Eucyclops prionophorus Kiefer, 1931. Authors’ drawings. Male from San Bernardino, Paraguay. A Urosome, ventral view B Antennule C P1 D P3 E P4. Scale bars: A = 100 µm, B–E = 50 µm.
Leg 5: Free segment subrectangular, 1.4 times longer than wide, bearing 1 inner spine and 2 setae; median seta longer than outer seta (about 1.3 times).
Inner spine 0.6 times longer than outer seta and 0.4 times as long as median seta.
Leg 6 (Figs 8E, 9C, 10A, 13A): Represented by small, low plate near lateral margin of genital somite, armed with 1 strong and long inner spine and 2 unequal setae. Inner spine not reaching half length of fourth urosomite. Inner spine about 1.6–2.5 times longer than median seta and 1.2–1.8 times longer than outer seta.
Remarks. Since its description in 1931 from a material collected in the USA, Eucyclops prionophorus has been recorded from various habitats in the Americas including the Laurentian Great Lakes in Canada and the USA, the Chihuahuan Desert in Mexico, and water bodies in savannahs and deciduous forests in Argentina, Paraguay and Uruguay (
Figs 14–17
Eucyclops (s. str) Bondi Kiefer, 1934
Eucyclops (s. str.) Bondi Kiefer, 1936
Eucyclops bondi, Smith and Fernando 1980;
Kiefer’s description.
Female: General aspect as the American Eucyclops prionophorus. Caudal rami 3.5 times longer than wide, with rami slightly divergent. Inner margin of caudal ramus naked, outer margin strongly ornamented with strong spinules. Proximal spines small but distal spines long (Fig. 4). Innermost apical seta longer than outermost seta; two middle setae show strongly heteronomous plumage.
Antennule only slightly longer than cephalothorax, bearing 12 segments, last three segments with a narrow hyaline membrane along margin.
Swimming legs normally segmented, with spines and setae. Third endopod of P4 two times longer than wide, inner spine longer than segment and even 1.5 times longer than the outer spine. The connecting plate of this leg is hairy on the free margin. Rudimentary leg (P5) with a slender inner spine, inner spine is at its insertion about twice as wide as one of the both setae. Seminal receptacle without special characteristics. Total length of animals, excluding apical setae of caudal ramus, 720–800 µm.
Male: total length, excluding apical setae of caudal ramus, 580–600 µm. As a main characteristic the reinforcement of genital somite (P6) should be considered. Of the three elements, the inner spine only measures 22–23 µm, clearly differing from the similar North American species Eucyclops prionophorus, the outermost plumose seta is longer than the spine, the median seta is as long as the spine. In the male of Eucyclops prionophorus the inner spine is more than three times longer, 71 µm.
Description based on Kiefer’s material.
Material examined. Holotype. Adult ♀ collected 16.02.1933 from Trou Caiman, Haiti, specimen dissected (slide reference numbers SMNK02079, SMNK02080). Additional material from Laguna Rincon, Haiti (slide reference numbers SMNK02393, SMNK02394). Staatliches Museum für Naturkunde Karlsruhe, Germany.
Female: Body length excluding caudal setae 720–800 µm. Prosome expanded at first and second somite, symmetrical in dorsal view. Urosome 5-segmented (Figs 14C, 16A–B), relatively elongate. Urosomal fringes strongly serrated, posterior margin of anal somite with row of long spinules. Genital double-somite symmetrical. Seminal receptacle typical of serrulatus-group, with rounded lateral arms on posterior margin. Genital double-somite 1.1 times as long as wide. Anal somite with hair-like spinules in anal opening, anal operculum slightly rounded (Fig. 16A). Length/width ratio of caudal ramus = 3.5. Inner margin of caudal ramus naked, outer margin partially covered (53–57%) by strong spinules which increase in size distally (Figs 14C, 16B). Dorsal seta (VII) long: 0.8 times of caudal ramus length, and 1.26–1.4 times as long as outermost caudal seta (III). Length ratio of innermost caudal seta (VI)/outermost caudal seta (III) = 1.07–1.25. Lateral caudal seta (II) inserted at 77–80% of caudal rami. All terminal caudal setae plumose.
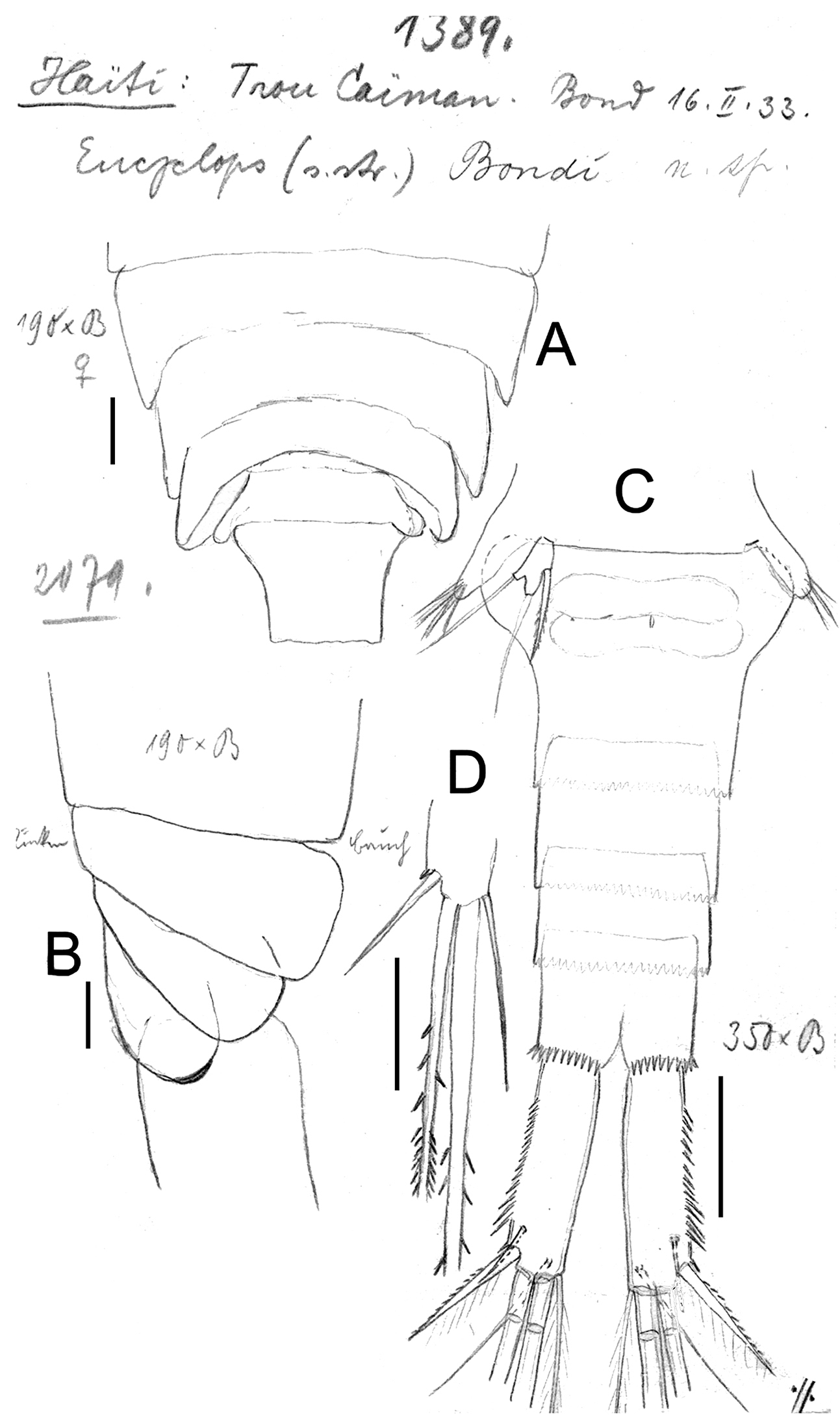
Eucyclops bondi Kiefer, 1934. Original drawings of F. Kiefer. Female Holotype from Trou Caiman, Haiti. A Prosome 2–5 and genital somite, dorsal view B Prosome 2–5, lateral view C Urosome, ventral view D Caudal setae of CR. Scale bars: A–D = 100 µm.
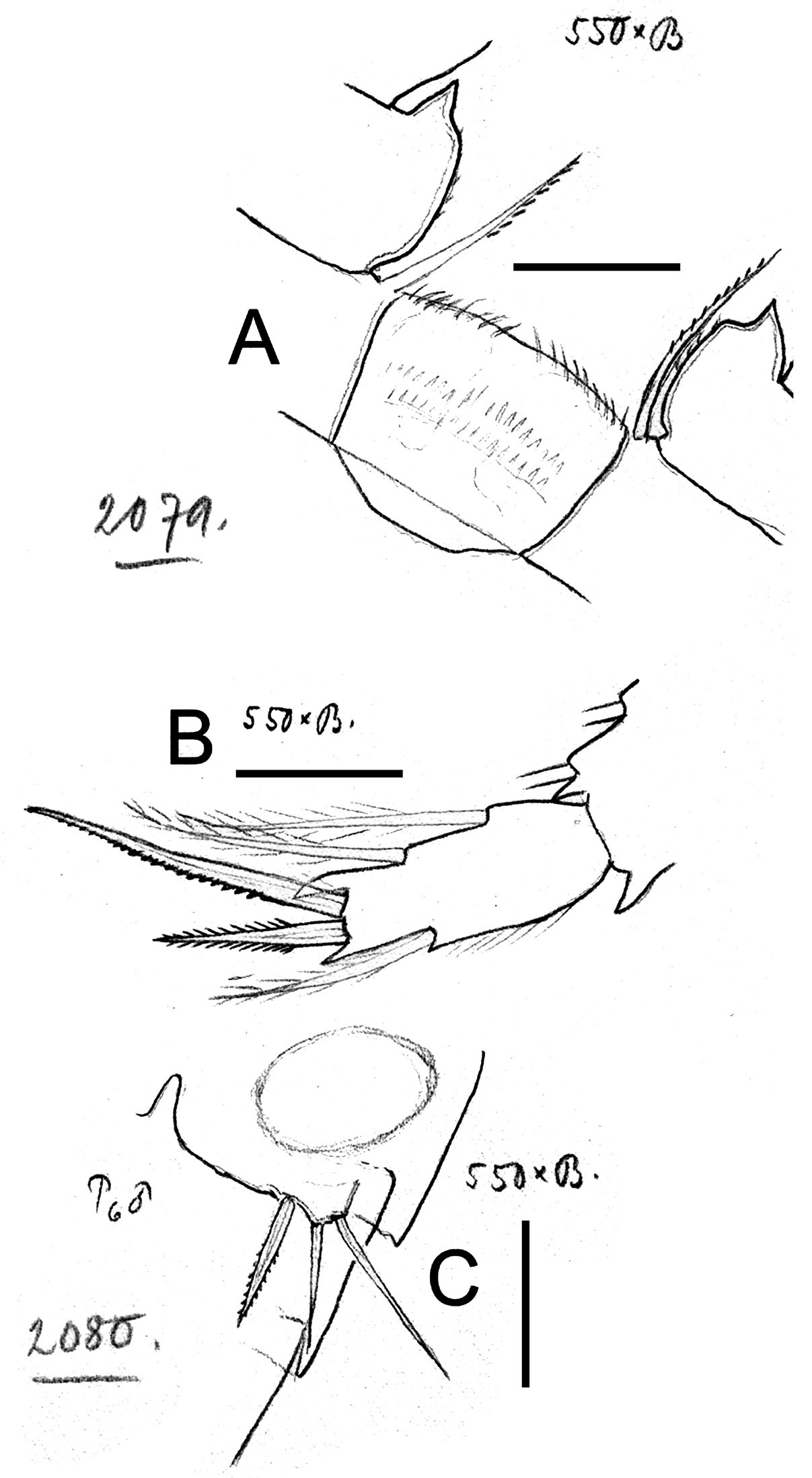
Eucyclops bondi Kiefer, 1934. Original drawings of F. Kiefer. Female Holotype A–B and male Paratype C from Trou Caiman, Haiti. A Intercoxal sclerite and coxal spines P4 B Enp3P4 C P6. Scale bars: A–C = 50 µm.
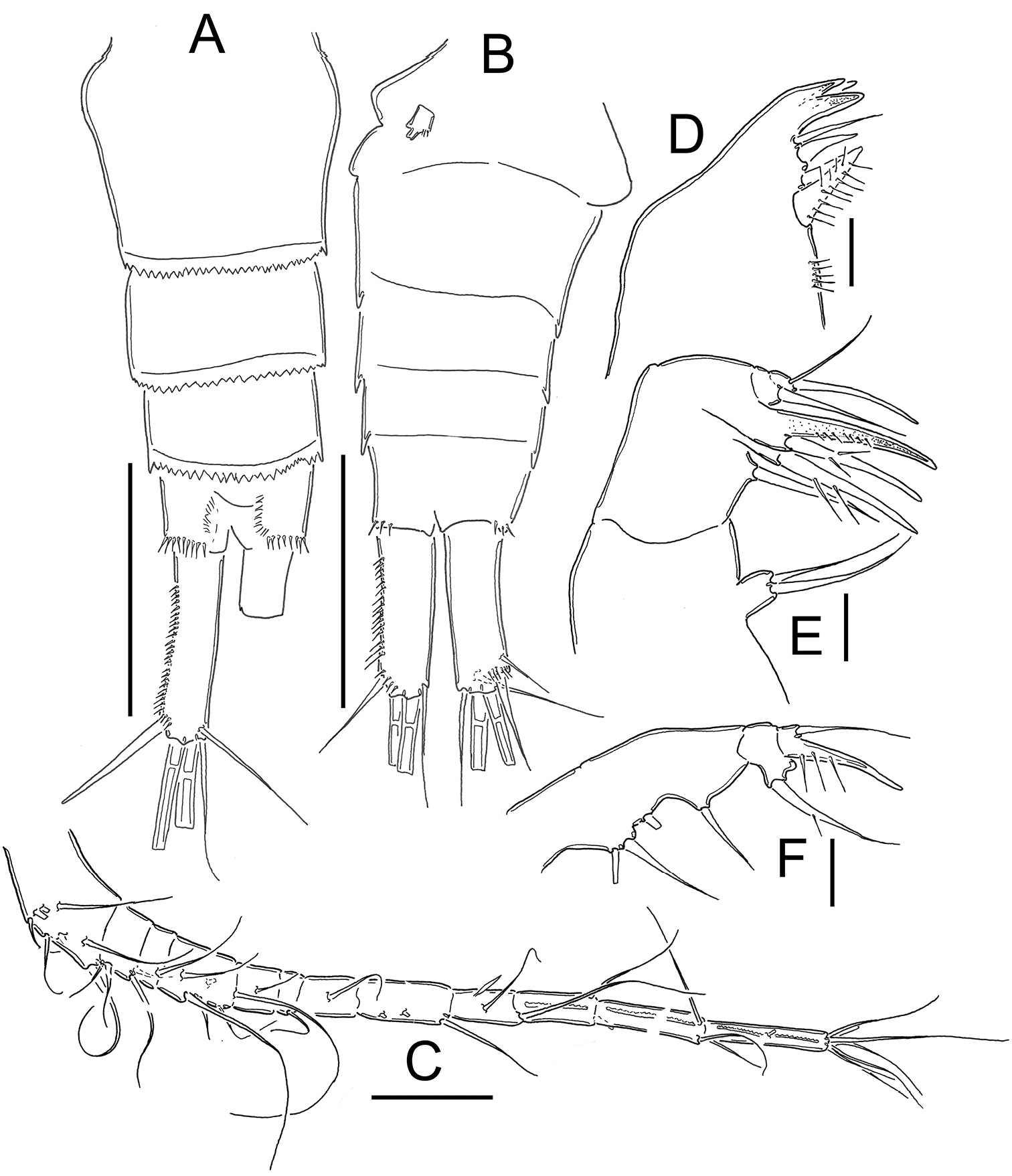
Eucyclops bondi Kiefer, 1934. Authors’ drawings. Female Holotype from Trou Caiman, Haiti. A Urosome, dorsal view B Urosome, ventral view C Antennule D Maxillule E Maxilla F Maxilliped. Scale bars: A–B = 100 µm; C = 50 µm; D–F = 20 µm.
Antennule (Fig. 16C): 12-segmented, reaching second prosomite; three distal segments with narrow hyaline membrane. Armament per segment as follows (s = seta, ae = aesthetasc, sp = spine): 1(8s), 2(4s), 3(1s), 4(6s), 5(1s), 6 (1s+1sp), 7(1s), 8(3s), 9(2s+1ae), 10(2s), 11(3s), 12(8s). Numbers in bold indicate segments with incomplete ornamentation. Aesthetasc of ninth segment short, reaching posterior margin of segment.
Antenna, Labrum and Mandible: not observable in slides.
Maxillule (Fig. 16D): precoxal arthrite with naked surface, with 3 strong chitinized distal claws. Spiniform seta on frontal side and palp not observed.
Maxilla (Fig. 16E): praecoxa and coxa partially fused. Praecoxa with 2 armed setae on endite. Coxal surface naked, bearing 1 biserially plumose seta. Distal endite of coxopodite well developed, with 2 apical setae, 1 strong and furnished with spinules and the other noticeably thicker and longer. Basal claw of basis with proximal row of spinules and 1 chitinized armed seta. Endopod 2-segmented, first segment with 1 seta, second with 2 setae.
Maxilliped (Fig. 16F): syncoxa naked, bearing 3 setae. Maxillipedal basis with 1 seta and no observable additional ornamentation. Endopod 2-segmented: Enp1 with 1 long, strong seta, Enp2 with 2 setae, proximal 1 chitinized and fused with segment, apical seta normal.
P1–P4: Endopod and exopods of all swimming legs three-segmented. Armature formula of all swimming legs as in Table 1.
Leg 1 (Fig. 17A): Coxa with strong, biserially setulated inner coxal seta. Basipodal spine long, reaching apical margin of Enp3; basipodal spine as long as endopod. Third endopodal segment 1.5 times as long as wide, apical spine of Enp3 0.9 times longer than length of Enp3, apicalmost seta of Enp3 1.4 times longer than apical spine. Spines of all exopodal segments elongate.
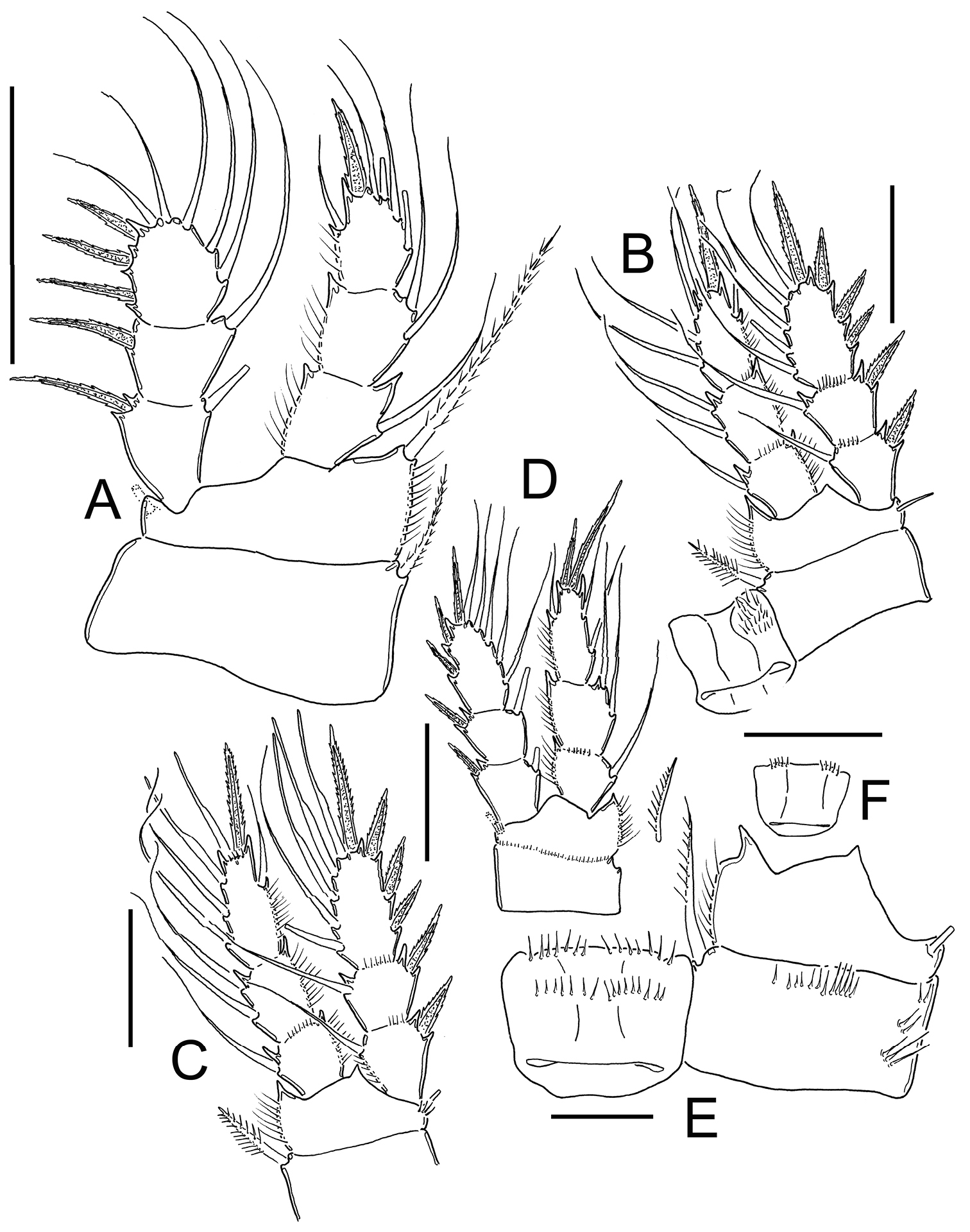
Eucyclops bondi Kiefer, 1934. Authors’ drawings. Female Holotype from Trou Caiman, Haiti. A P1 B P2 C P3 D P4 E P4, caudal surface F Intercoxal sclerite P4, frontal surface. Scale bars: A–D, F = 50 µm, E = 20 µm.
Leg 2 (Fig. 17B): Single group of small spinules in each side on anterior surface of intercoxal sclerite. Distal margin of intercoxal sclerite with 2 round chitinized projections. Coxa with strong, biserially setulated inner coxal seta. Enp3 1.9 times as long as wide, apical spine 1.2 times longer than length of Enp3, apicalmost seta of Enp3 1.4 times longer than apical spine.
Leg 3 (Fig. 17C): Coxa with strong biserially setulated coxal seta. Enp3 2.2 times longer than wide, apical spine as long as Enp3, apicalmost seta of Enp3 as long as apical spine. Enp3 and Exp3 with modified setae on.
Leg 4 (Figs 15A–B, 17D–F): Intercoxal sclerite with rows I, II and III. Row I with 7 strong spinules in each side and small gap in between. Row II with 16–18 spinules, row III with 14 strong spinules. Caudal coxal surface with spinule formula as: A, B (3), C+D (12), E (2), F, G (2), H, I. Inner spine of coxa with heteronomous setulation: basally with long hair-like setules, distally with spine-like setules; lateral edge of coxal spine naked. Enp3P4 length/width ratio 2.5; length ratio inner/outer spines of Enp3P4 = 1.6–1.8; length ratio inner spine in Enp3P4/Enp3P4 = 1.3; length ratio outer spines in Enp3P4/Enp3P4 = 0.7. Lateral seta in Enp3P4 inserted at 71% of segment length. Enp3 and Exp3 with normal setae.
Leg 5 (Fig. 14C): Free segment subrectangular, 2 times longer than wide, with 1 inner spine and 2 setae; median seta as long as outer seta (1:1) but about 1.3 times longer than inner spine. Inner spine 1.6 times as long as segment.
Male: Body length excluding caudal setae 580–600 µm. Urosome 6-segmented, posterior margins of urosomites smooth. Caudal rami 2.6 times longer than wide; medial margin of caudal ramus naked, strong spines at insertion point of lateral caudal seta (II).
Leg 6 (Fig. 15C): Represented by small, flat plate near lateral margin of genital double somite with 1 strong short inner spine and 2 unequal setae. Inner spine not reaching posterior margin of third urosomite. Inner spine about 0.7 times as long as outer seta and as long as median seta.
Remarks. After Kiefer’s description (
Figs 18–21
Eucyclops leptacanthus,
Eucyclops cf. leptacanthus, Reid 1993
Eucyclops leptacanthus,
Kiefer’s description.
Total length of the single female found 0.76 mm, excluding caudal setae. The caudal ramus exactly four times as long as wide (82.5µ: 20.5µ). Position and shape of the two rami can be seen in Fig. 25. Inner margin is naked, outer margin provided with a “serra”, extending along almost the entire length of ramus, proximal spinules very small, very long spinules at the height of insertion of lateral seta, practically setiform. Measurements of setae of caudal rami from the innermost to outermost: 80µ, 356µ, 198µ, 60µ, length of dorsal seta about 45–50 µ; the ratio, based on the outermost terminal seta, is therefore 1.33: 5.93: 3.30: 1 (0.8). The two longest setae are quite weak and heteronomously plumose (Fig. 25).
Twelve-segmented antennules, margin of last three segments with a narrow hyaline membrane, margin of membrane finely denticulated (Fig. 26). Segmentation and setation of the swimming legs as usual, the final segment of the Enp of fourth leg with a length/width: 44.5µ: 20.5µ = 2.17. Inner apical spine about 55µ-56µ, notably longer than the length of segment and significantly longer than the outer spine which measures 35µ (Fig. 28). Segment of rudimentary leg elongated and bearing small and slender inner spine (Fig. 29). The seminal receptacle was not perceptible.
The male has not yet been found.
Systematic position. in the complex genus Eucyclops, where many forms exist that are similar to the single female specimen found here, a new species can only be established when the new form is distinguished by unique characteristics. The specimen of Eucyclops herein described possesses a well-developed serra on the caudal rami, a rudimentary leg with a slender inner spine, long apical spines on the last segment of endopod of fourth leg, as well as a finely denticulate hyaline membrane on the three last segments of the antennule, a combination of characters that I have not seen in any of the known Eucyclops species. For this reason I considered it as a new species and named as Eucyclops leptacanthus because of its slender spines on the fifth and fourth legs.
In the sample 10e I did find another Eucyclops-female. The hyaline membrane on the last segments of the antennules is equally finely denticulate; the serra on the caudal rami is not particularly remarkable, the inner spine of rudimentary leg is thicker, and the apical spine of the last segment of the endopodite of the fourth leg is wider than that in the above described species. This specimen must remain undetermined because of the few characters that could be seen. The same holds true in the two Eucyclops males from sample 11.
Description based on Kiefer’s material.
Material examined. Holotype. Adult ♀collected 03.11.1952 from Lake Orinoco, Barrancas, Venezuela (slides reference numbers SMNK05409, SMNK05410). Staatliches Museum für Naturkunde Karlsruhe, Germany.
Female: Body length of holotype, excluding caudal setae, 760 µm. Urosome 5-segmented (Fig. 20A): relatively elongate; urosomal fringes smooth or weakly serrated. Genital double-somite symmetrical. Seminal receptacle typical of serrulatus-group, with rounded, lateral arms in posterior margin. Genital double somite about 1.3 times longer than wide. Length/width ratio of caudal ramus = 3.9–4.0; inner margin of caudal ramus naked; outer margin with strong spinules covering 63–64% of segment length, spinules distally increase in size (Figs 18A, 20A). Dorsal seta (VII) short, 0.5 times of length of caudal ramus, and 0.7 times as long as outermost caudal seta (III). Ratio of innermost caudal seta (VI)/outermost caudal seta (III) = 1.3–1.4. Lateral caudal seta (II) inserted at 74% of caudal rami. All terminal caudal setae plumose.
Antennule (Figs 19A, 20B–C): 12-segmented. Armament per segment as follows (s = seta, ae = aesthetasc, sp = spine): 1(7s), 2(3s), 3(1s), 4(6s), 5(1s), 6 (1s+1sp), 7(2s), 8(3s), 9(2s), 10(2s), 11(3s), 12(7s). Numbers in boldface indicate segments with incomplete ornamentation.
Antenna (Fig. 20D): Basis (2s + Exp), 3-segmented End (1s, 8s and 7s). Only row N17 was observable on basis of the holotype.
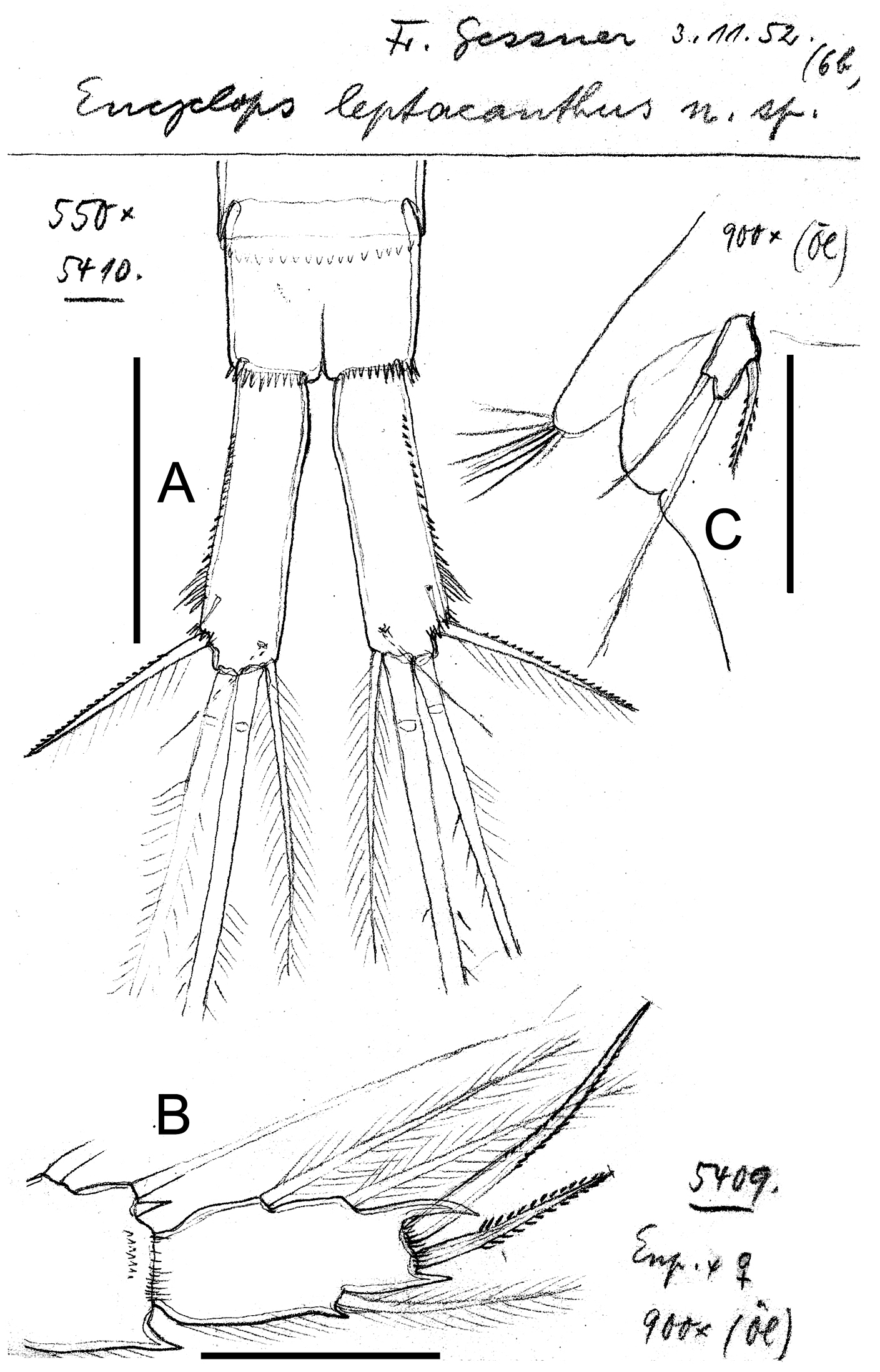
Eucyclops leptacanthus Kiefer, 1956. Original drawings of F. Kiefer. Female Holotype (two preparations same specimen) from Lake Orinoco, Venezuela. A Caudal rami B Enp3P4 C P5. Scale bars: A = 100 µm; B–C = 50 µm.
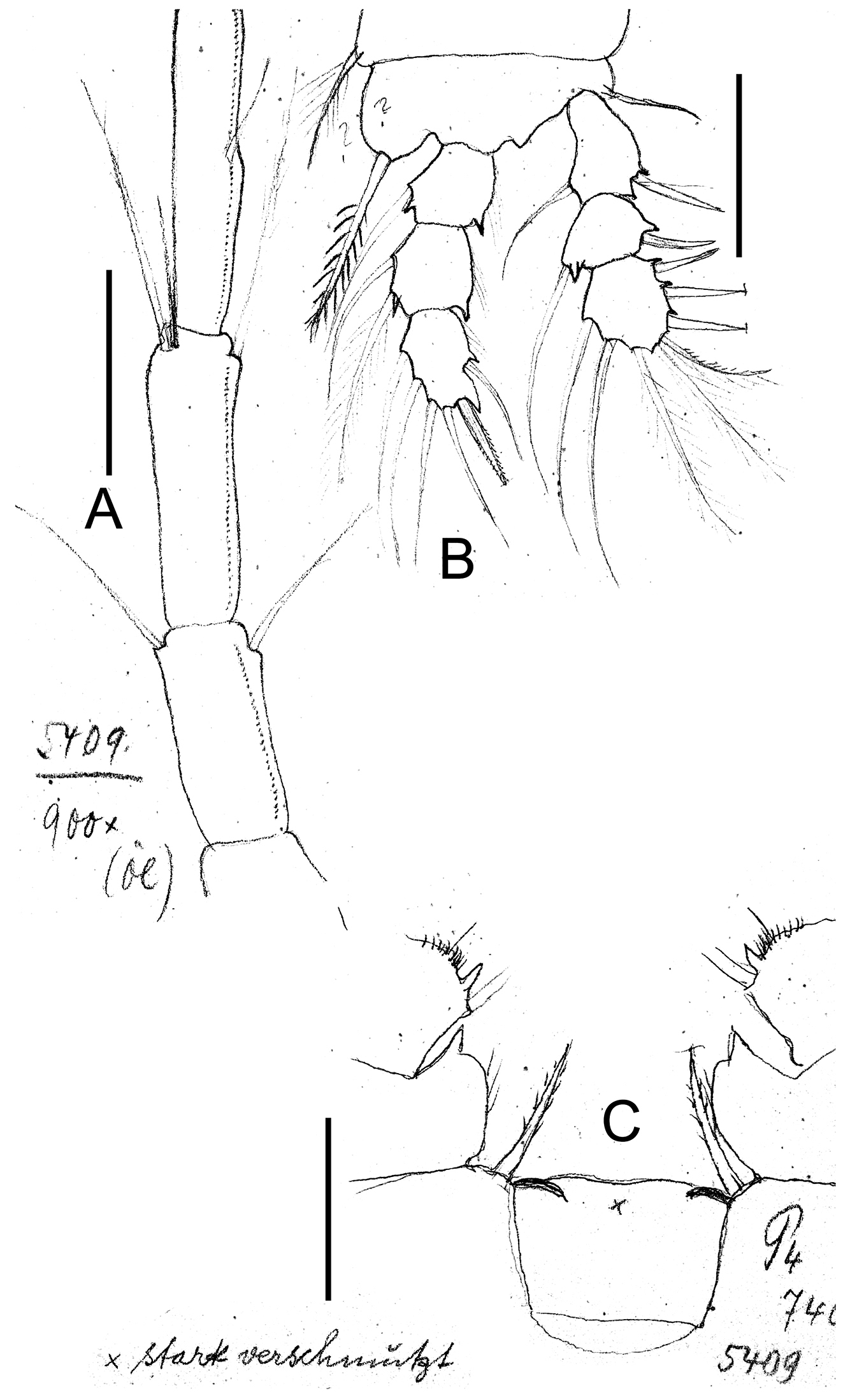
Eucyclops leptacanthus Kiefer, 1956. Original drawings of F. Kiefer. Holotype (two preparations same specimen) from Lake Orinoco, Venezuela. A Last segments of antennule B P1 C Intercoxal sclerite P4. Scale bars: A–C = 50 µm.
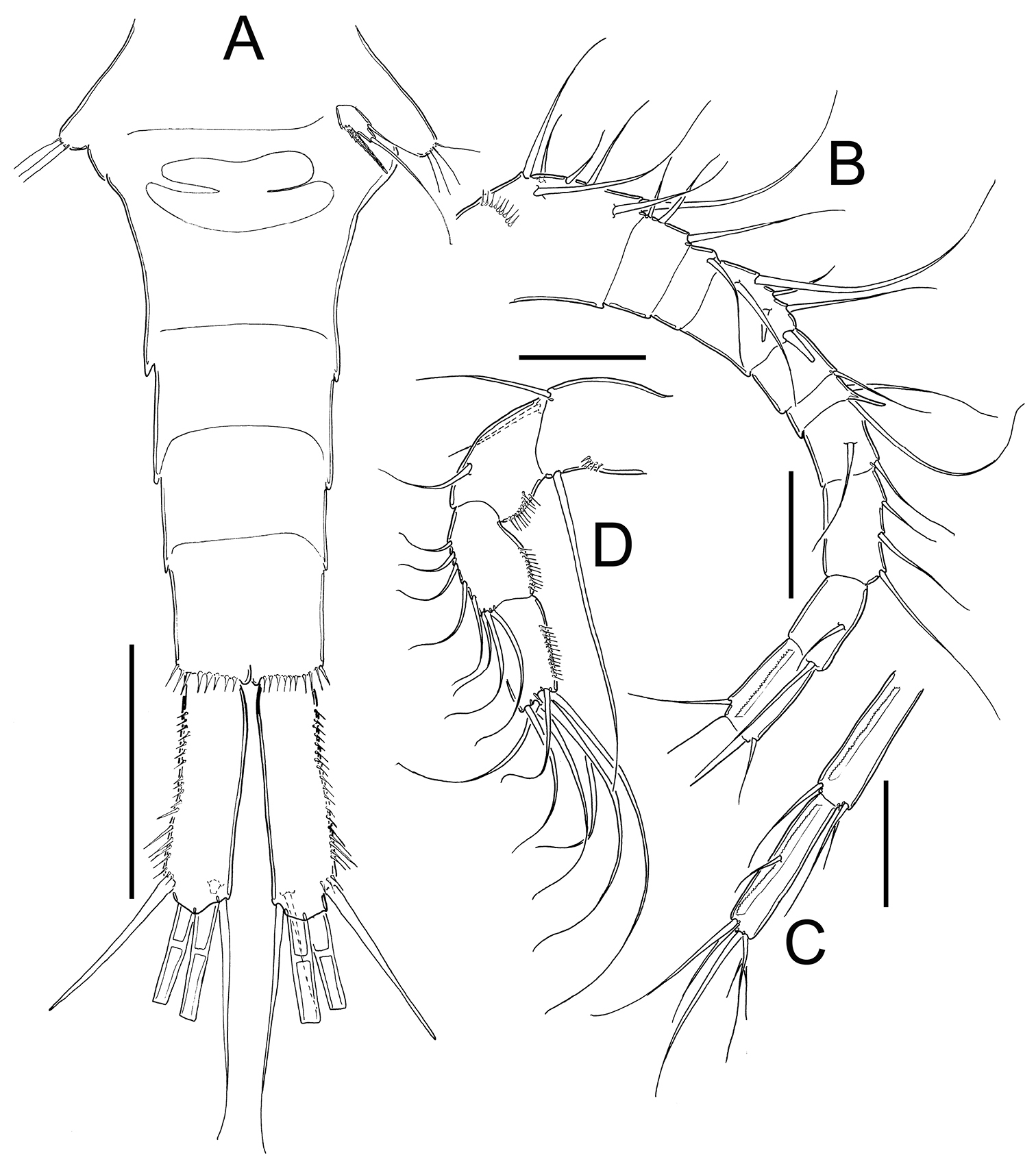
Eucyclops leptacanthus Kiefer, 1956. Authors’ drawings. Female Holotype from Lake Orinoco, Venezuela. A Urosome, ventral view B Antennule, segments 1–10 C Antennule, segments 11–12 D Antenna. Scale bars: A = 100 µm, B–D = 50 µm.
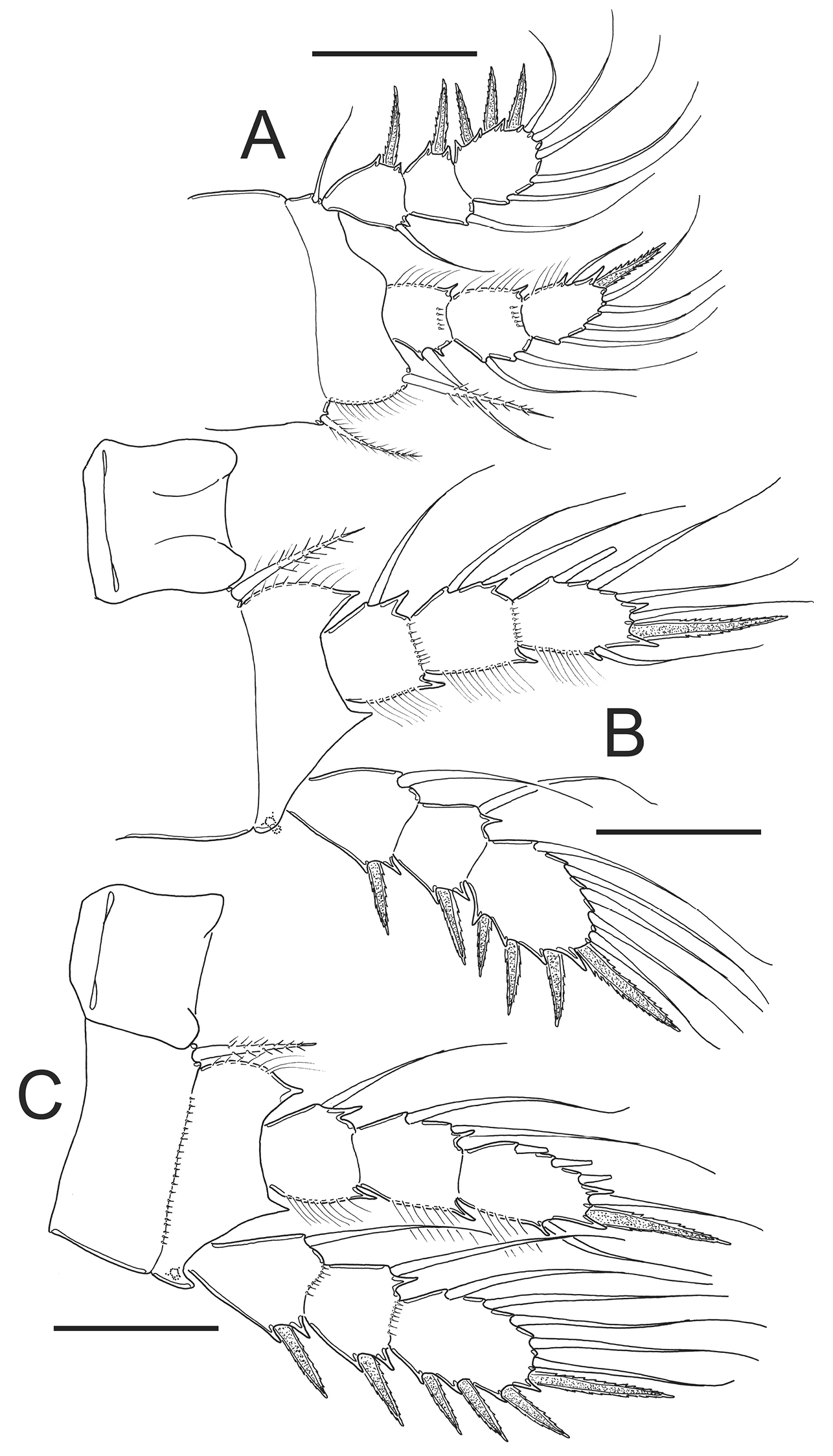
Eucyclops leptacanthus Kiefer, 1956. Authors’ drawings. Female Holotype from Lake Orinoco, Venezuela. A P1 B P2 C P3. Scale bars: A–C = 50 µm.
Mouthparts: not observable in the slide.
P1–P4: Endopods and exopods of all swimming legs three-segmented. Armature formula of all swimming legs as in Table 1.
Leg 1 (Figs 19B, 21A): Coxa with strong, biserially setulated inner coxal seta. Basipodal spine not reaching middle of Enp3; and 0.7 times as long as endopodal ramus. Enp3 1.4–1.7 times as long as wide, apical spine of Enp3 as long as segment Enp3 (1:1), apicalmost seta of Enp3 1.5–1.8 times longer than apical spine.
Leg 2 (Fig. 21B): No ornamentation observed on intercoxal sclerite, distal margin with 2 rounded projections. Coxa with strong, biserially setulated inner coxal seta. Enp3 1.6 times longer than wide, apical spine on Enp3 1.3 times as long as segment, Exp3 2.2 times as long as wide, apical spines of Exp3 0.9 times as long as segment. No modified setae were observed.
Leg 3 (Fig. 21C): No cuticular ornamentation was observed on intercoxal sclerite, distal margin with 2 rounded projections. Coxa with strong, biserially setulated inner coxal seta. Enp3 2.0 times longer than wide, apical spine on Enp3 1.1 times as long as segment, apical seta of Enp3 1.2 times as long as apical spine. Exp3 1.9 times as long as wide, apical spine of Exp3 as long as segment (1:1). No modified setae observed.
Leg. 4 (Figs 18B, 19C): Intercoxal sclerite could not be clearly observed. Inner coxal spine with heteronomous setulation: basally with long hairs yet distally with spinules; lateral edge of inner coxal spine with 2 apical spine-like setules, proximal surface naked. Length/width ratio Enp3P4 = 2.3; length ratio inner/outer spines of Enp3P4 =1.5; length ratio inner spine of Enp3P4/Enp3P4 = 1.3; length ratio outer spine of Enp3P4/Enp3P4 = 0.8-0.9. Lateral seta of Enp3P4 inserted at 67-70% of total length of segment. No modified setae were observed.
Leg 5 (Figs 18C, 20A): Free segment subrectangular, 1.8 times as long as wide bearing 1 inner spine and 2 setae; median seta longer than outer seta (about 1.8 times) and inner spine (about 2.6 times). Inner spine noticeably slender, 1.3 times as long as segment.
Remarks. Eucyclops leptacanthus is another species described by
Among the 108 species and subspecies currently known in the genus Eucyclops, 28 are distributed in the Americas, most of the records in the continent are from surveys in the Eastern United States, Mexico, Argentina and Brazil (
We gratefully acknowledge the support and confidence from Dr. Hans Walter Mittmann, Staatliches Museum Für Naturkunde, Karlsruhe, Germany in allowing us to examine the type material of the species included in the present work. He also granted permission to include the unpublished original drawings by F. Kiefer in this work. We also acknowledge the help, time and patience of Jörg Pertzel and Gerald Islebe in translating the original descriptions of the species from German to English. This work is part of the first author’s (NFM-S) Doctoral Thesis developed at El Colegio de la Frontera Sur (ECOSUR). This contribution was supported by CONACyT project 133404-Investigación Científica Básica 2009. The comments and suggestions of two anonymous reviewers and the ZooKeys editor of Copepoda are deeply appreciated.