





(C) 2013 Arkady A. Schileyko. This is an open access article distributed under the terms of the Creative Commons Attribution License 3.0 (CC-BY), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
For reference, use of the paginated PDF or printed version of this article is recommended.
A new species of the centipede genus Newportia, Newportia stoevi sp. n., is described from Rio Encantado Cave, Puerto Rico. It differs from all congeners by having sternites distinctly margined laterally and ultimate legs bearing 4 spinous processes on both prefemur and femur, and 2 on tibia. The value of some terms used in the taxonomy of the genus have been analyzed and an amended identification key to the species of Newportia is provided.
Newportia, Newportiinae, new species, identification key, external characters
The genus Newportia Gervais, 1847 is still poorly known. It is especially so with regards to the Puerto Rican fauna where only two species have hitherto been registered.
Herewith, I describe a new species of Newportia recently collected in Puerto Rico by Dr Petar Beron from the National Museum of Natural History, Sofia (NMNHS). The specimen was assigned to Newportia and tentatively identified as a new species by Dr Pavel Stoev, curator of Myriapoda at the NMNHS who committed it for further study to me. This specimen differs from all congeners, in the first place, by important traits of the ultimate legs (which are normally developed and have no traces of regeneration) and unusually developed lateral margination of sternites.
The identification key to the species of Newportia (
urn:lsid:zoobank.org:act:AE5E2F31-F3F2-45EB-9AD6-24EDA31D1D66
http://species-id.net/wiki/Newportia_stoevi
Figs 1–10Puerto Rico, Florida Co., Rio Encantado Cave, 1 (sub?)adult, 29.07.2009, leg. P. Beron (NMNHS).
Puerto Rico, Florida Co., Rio Encantado Cave.
named after my friend and colleague Dr Pavel Stoev who drew my attention to this new species.
Tergite 1 with rounded anterior transverse suture and incomplete paramedian sutures. Sternites distinctly margined laterally. Ultimate legs: prefemur with 4, femurwith 3 small spinous processes medially and 1 ventrally; tibia with 2 small spinous processes medially. Tarsus 1 large and clavate (bulbous), clearly differing from the much thinner tarsus 2; the latter consisting of 19–20 articles.
Length of body ca 17 mm, length of ultimate legs about 9 mm. Color (in ethanol): entire animal uniformly light-yellow with cephalic plate and forcipular segment slightly darker (Fig. 1). Body sparsely pilose; sternites and legs less setose than tergites.
Antennae composed of 17 articles (Fig. 2), reaching rear edge of tergite 5 when folded backwards; 2.5 basal antennal articles covered by a few long setae, subsequent articles densely pilose. Basal antennal articles somewhatflattened dorso-ventrally.
Head: cephalic plate visibly longer than wide, with rounded corners and very short paramedian sutures at posterior margin.
Second maxillae: as in all other Newportia species but dorsal spur on article 2 of the telopodite not recognisable. Pretarsus without spurs, with well-developed dorsal brush. The angle between the longitudinal axes of pretarsus and article 3 of telopodite slightly more than 100° (Fig. 3), which is quite unusual condition in Scolopendromorpha.
Forcipular segment: coxosternite without any visible sutures (including thechitin-lines). Anterior margin of coxosternite evidently convex (Fig. 3), divided by a median diastema into two low additionally sclerotised lobes; each lobe bearing a longseta. Trochanteroprefemoral process absent. Tarsungula normal.
Tergites: anterior margin of tergite 1 covered by the cephalic plate; tergite 1with a rounded anterior transverse suture and paramedian sutures stretching from the transverse suture to the posterior tergal margin. Tergite 3 with a very characteristic thin oblique sutures bordering the anterior corners of tergite. Tergites 2-22 with complete paramedian sutures, tergites 3-21(22)withlateral longitudinal sutures (Fig. 4). Tergite 23 lacking sutures, its posterior margin convex. Tergite margination virtually absent, only tergite 23 distinctly margined laterally.Tergite 23 much wider rather than long and nearly rectangular in shape; its lateral sides slightly rounded (Fig. 4). All tergites without medial keel; pretergites also missing.
Sternites: trapeziform, 2-22 with incomplete (equally shortened from both sides) but with a well expressed median longitudinal sulcus. Sternites 2-21 with definite and complete lateral margination (Fig. 5) through lateral longitudinal sutures (see Remark 2); endosternites absent. Sternite 23 trapeziform, with a few very short (spur-like) setae on lateral sides (Fig. 6), with a straight posterior margin.
Legs: prefemur, femur and tibia with a few large setae (Fig. 5); tarsi with more numerous setae of various length and size. Tibia of legs 1–20 with a lateral spur; both, ventral tibial spur and tarsal spur absent. Tarsi of legs 1–21 (Fig. 5) without distinct division between tarsus 1 and 2; pretarsi long, thin and sharply pointed.Pretarsi of legs 1-22 with two thin and long (as long as 1/2 of pretarsus)accessory spines.
Coxopleuron (Figs 6, 7):nearly completely pierced with coxal pores of various size – only coxopleural process and a narrow area bordering posterior margin of coxopleuron remaining poreless. Coxopleural process (Figs 6, 7)as long as ultimate sternite, conical, without additional spines. Coxopleural surface without setae.Posterior margin of pleuron of ultimate leg-bearing segment forming avery obtuse angle.
Ultimate legs (Fig. 8): slender, ca 9 mm long, width of prefemur ca 0.5 mm.Prefemur triangular in cross-section, with a standard row of 4 ventral spinous processes (Fig. 7), some spurs (strong, spine-like setae of various length) dorso-laterally and more numerous similar spurs dorso-medially (Fig. 4). All four prefemoral ventral spinous processes are of the same size, apically curved and ending in a pointed harpoon-like tip, which is accompanied by a long seta. Femurcylindrical, with 3 small spinous processes medially (Fig. 9) and 1 ventrally in the middle of femur (Fig. 10). Tibia cylindrical, with 2 small spinous processes medially: one close to its base and another at mid length (Figs 9, 10). Both femoral and tibial spinous processes are accompanied by a single long ventral seta. Tibia practically as long as prefemur or femur. Tarsus well divided into tarsus 1 and tarsus 2 (Fig. 8), former as long as 1/2 of tibia. Tarsus 1 (Figs 8–10) is enlarged and clavate (bulbous); tarsus 2 thin, consisting of 19 (or 20) articles (Fig. 8). In a few places annulation of tarsus 2 is somewhat vague; for example, the very long ultimate article seems to consist of two articles, which are not well divided. Ultimate legs without pretarsus.
The species is hitherto known only from its type locality.
Being -250 m deep and 16 910 m long Rio Encantado is the deepest and the longest cave system in Puerto Rico. This system lies in the Tertiary limestone area which stretches along the northern coast of the island (
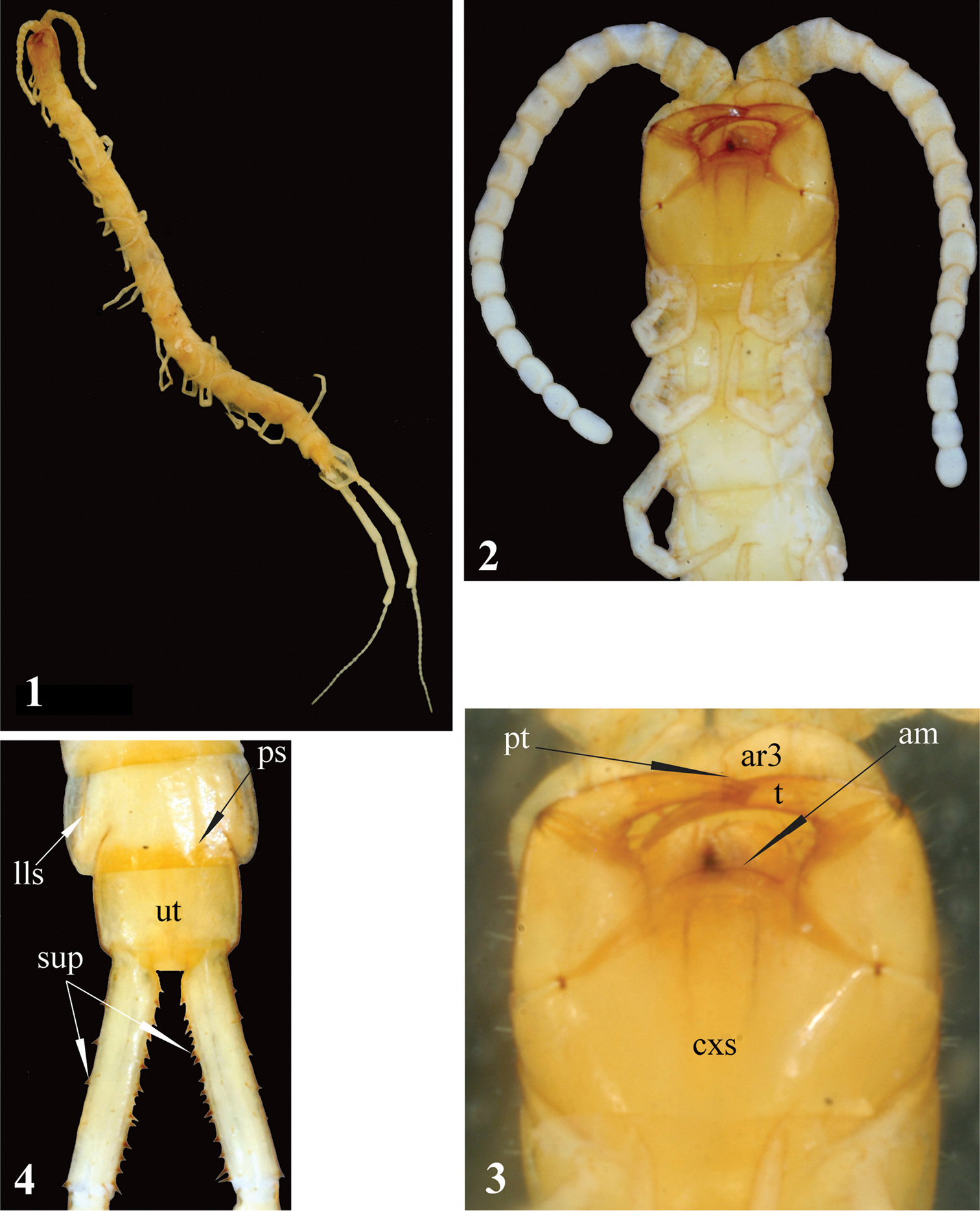
Newportia stoevi, sp. n. 1 Habitus 2 Head and anterior segments, ventral view 3 Forcipular segment, ventral view 4 Tergites 22 and 23 and prefemora of ultimate legs, dorsal view; (pt) – pretarsus of second maxilla, (ar3) – article 3 of telopodite of second maxilla, (cxs) – forcipular coxosternite, (am) – anterior margin of coxosternite, (t) – tarsungulum, (ps) – paramedian sutures, (lls) – lateral longitudinal sutures, (ut) – tergite of ultimate leg-bearing segment, (sup) – spurs of ultimate prefemur.
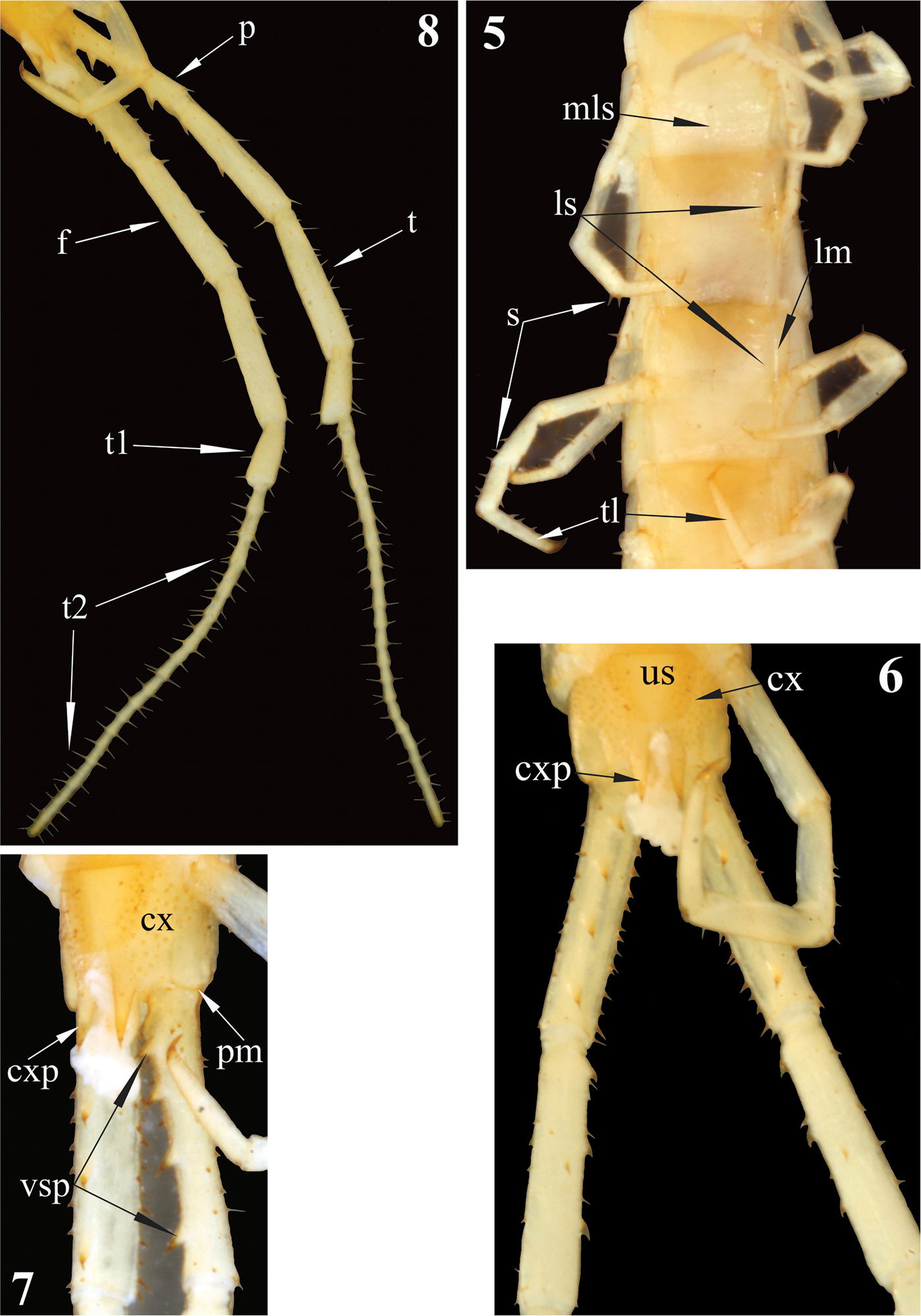
Newportia stoevi, sp. n. 5 Segments and midbody legs, ventral view 6 Posterior body end, ventral view 7 Left side of ultimate leg-bearing segment and prefemora of ultimate legs, ventro-lateral view 8 Ultimate legs, ventro-lateral view; (mls) – median longitudinal sulcus, (ls) – lateral sutures, (lm) – lateral margination, (s) – setae, (tl) – monoarticulated tarsus of locomotory leg, (us) – sternite of ultimate leg-bearing segment, (cx) – coxopleuron, (cxp) – coxopleural process, (pm) – posterior margin of pleuron of ultimate leg-bearing segment, (vsp) – ventral spinous processes of ultimate prefemur, (p) – prefemur, (f) – femur, (t) – tibia, (t1) – tarsus 1, (t2) – tarsus 2.
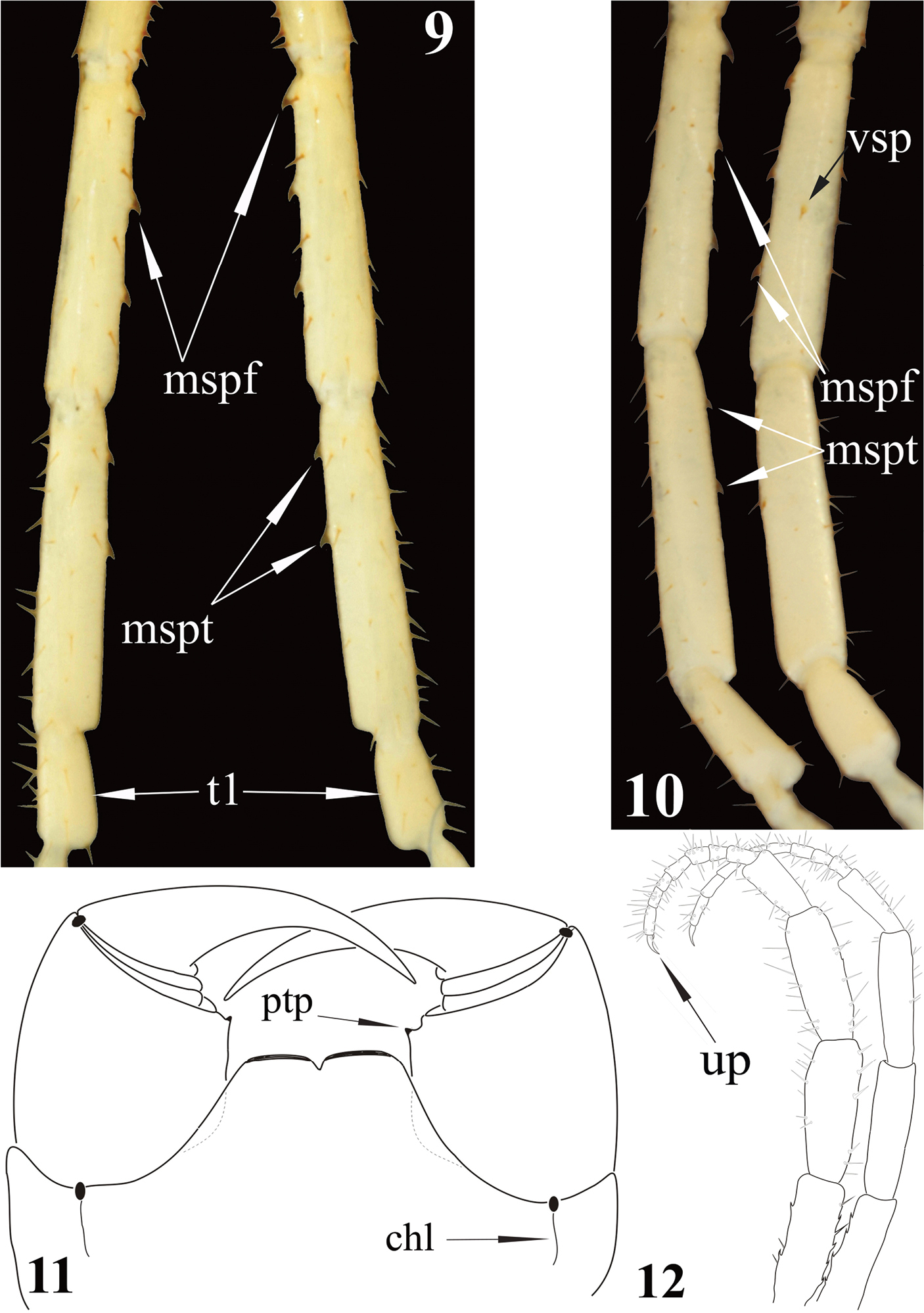
Newportia stoevi, sp. n. 9 Femora, tibiae and tarsi 1 of ultimate legs, dorsal view 10 Femora, tibiae and tarsi 1 of ultimate legs, ventral view; Newportia divergens Chamberlin, 1922 11 Forcipular segment, ventral view (after
1 This species is morphologically close to Newportia heteropoda Chamberlin, 1918 from which it can be readily distinguished by the following traits of the ultimate pair of legs: number of articles of tarsus 2 (19 vs. 9 in Newportia heteropoda); presence of 4 (vs. 3 in Newportia heteropoda) spinous processes on femur; presence of 2 (vs. 0 in Newportia heteropoda) such processes on tibia.
2
3 Some groups of scolopendromorphs (the majority of Scolopendrinae, Otostigminae, Scolopocryptopinae and Plutoniumidae) have a well-developed, strongly sclerotized disto-medial projection of the forcipular trochanteroprefemur. Formerly, I used the term “forcipular median tooth” for it, but
4 As for the vague annulation of some articles of ultimate tarsus 2 in Newportia stoevi, I should mention that there are a few other species of Newportia in which this trait is observed, for example Newportia albana Chamberlin, 1957 and Newportia diagramma Chamberlin, 1921 (see REMARKS to Newportia albana and Figure 5c of Newportia diagramma in
5 In some species of Newportia legs have one tarsal spur and two (lateral and ventral) tibial spurs, other species have one (lateral) tibial spur only (as Newportia stoevi) and in Newportia phoretha Chamberlin, 1950 spurs are entirely lacking (see p. 290 in
One of the main problems for identification of scolopendromorph centipedes is the high number of new species, described in the last decades that are still remaining outside the contemporary identification keys. I suggest that every description of new species in large genera (like Newportia) to be accompanied by the respective update of the available identification key. In cases where the genus includes just a few species, the identification key should be completely re-written.
The most recent key to the species of Newportia was provided by
| 1 | Tarsus 2 of ultimate legs clearly divided into distinct articles | 2 |
| – | Tarsus 2 of ultimate legs undivided | 31 |
| 2 | Ultimate leg with a well-developed (claw-shaped) pretarsus which is as long as, or longer than half of the ultimate article of tarsus 2 (Fig. 12) | Newportia unguifer |
| – | Ultimate leg without a well-developed pretarsus | 3 |
| 3 | Tergite 1 without an anterior transverse suture | Newportia sargenti |
| – | Tergite 1 with an anterior transverse suture | 4 |
| 4 | Tergite 1 with a rounded anterior transverse suture and, generally, with paramedian sutures which do not form a “W” just behind the anterior transverse suture; in a few species these sutures are absent or extremely short (Fig. 13) | 5 |
| – | Tergite 1 with an anterior transverse suture in the form of a very obtuse angle and with paramedian sutures forked anteriorly, thus forming a “W” just behind the anterior transverse suture (Fig. 14) | 21 |
| 5 | Some pairs of legs, usually 2(4)-(19)20, with tibial spurs | 6 |
| – | Tibial spurs missing on all legs | Newportia phoretha |
| 6 | Femur of ultimate legs without spinous processes | 7 |
| – | Femur of ultimate legs with spinous processes | 10 |
| 7 | Tergite 1 with rudimentary paramedian sutures (Fig. 13) or sutures completely lacking | Newportia pusilla |
| – | Paramedian sutures of tergite 1 half-complete or complete, sometimes shortly interrupted in the middle | 8 |
| 8 | Coxopleural process extremely short; tergite 1 with poorly developed paramedian sutures (Fig. 15) which cross the anterior transverse suture | Newportia diagramma |
| – | Coxopleural process normal (Fig. 7); tergite 1 with well-developed paramedian sutures stretching between anterior transverse suture and posterior tergal margin | 9 |
| 9 | Tarsus 2 of ultimate legs composed of 19–25 articles | Newportia aureana |
| – | Tarsus 2 of ultimate legs composed of 6–7 articles | Newportia longitarsis tropicalis |
| 10 | Tibiae 2-20 with lateral and ventral spurs | 11 |
| – | Tibiae 2-20 with a lateral spur only | 12 |
| 11 | Femur of ultimate legs with 1(-2) ventral spinous process(es) | Newportia cubana |
| – | Femur of ultimate legs with 2-3 medial spinous processes | Newportia longitarsis virginensis |
| 12 | Tarsus 2 of ultimate legs composed of 4 articles; tarsus 1 almost as long as the tibia | Newportia dentata |
| – | Tarsus 2 of ultimate legs composed of 7-26 articles; tarsus 1 quite shorter than the tibia | 13 |
| 13 | Tarsus 2 of ultimate legs composed of 26 articles; cephalic plate without paramedian sutures | Newportia leptotarsis |
| – | Tarsus 2 of ultimate legs composed of 7–20 articles; cephalic plate often with incomplete paramedian sutures | 14 |
| 14 | Anterior ends of the half-complete paramedian sutures of tergite 1 very shortly bifurcate behind the anterior transverse suture (Fig. 16) | Newportia oligopla |
| – | Paramedian sutures of tergite 1 from absent to complete, never bifurcated anteriorly | 15 |
| 15 | Tarsus of ultimate legs uniformly divided, without distinction into tarsus 1 and tarsus 2 | Newportia adisi |
| – | Tarsus of ultimate legs distinctly divided into tarsus 1 and tarsus 2 | 16 |
| 16 | Femur of ultimate legs with 3-4 spinous processes | 17 |
| – | Femur of ultimate legs with 1-2 spinous processes | 18 |
| 17 | Femur of ultimate legs with 3 spinous processes, tibia without spinous processes and tarsus 2 composed of 9 articles | Newportia heteropoda |
| – | Femur of ultimate legs with 4 spinous processes, tibia with 2 spinous processes (Figs 9, 10) tarsus 2 of 19 articles (Fig. 8) | Newportia stoevi sp. n. |
| 18 | Tergite 1 with complete paramedian sutures which cross the anterior transverse suture | Newportia longitarsis longitarsis |
| – | Tergite 1 with incomplete paramedian sutures of various length, from half-complete (Fig. 17) to rudimentary (Fig. 13), running between the posterior tergal margin and the anterior transverse suture; rarely without any trace of paramedian sutures | 19 |
| 19 | Cephalic plate with a thin transverse suture which crosses the short paramedian sutures close to the posterior margin of the cephalic plate | Newportia longitarsis sylvae |
| – | Cephalic plate without any transverse suture | 20 |
| 20 | Prefemur of ultimate legs with 3 large ventral spinous processes | Newportia longitarsis guadeloupensis |
| – | Prefemur of ultimate legs with 4 large ventral spinous processes | Newportia longitarsis stechowi |
| 21 | Tarsus 2 of ultimate legs composed of 39–40 articles | Newportia sabina |
| – | Tarsus 2 of ultimate legs composed of less than 30 articles | 22 |
| 22 | Each leg with a tarsal spur; tibia of ultimate legs longer than femur | 23 |
| – | Legs without tarsal spurs; tibia of ultimate legs shorter or as long as femur | 26 |
| 23 | Femur of ultimate legs with 2 ventral spinous processes | Newportia morela |
| – | Femur of ultimate legs with 3 ventral spinous processes | 24 |
| 24 | Outer branches of forked paramedian sutures of tergite 1 extending in front of the anterior transverse suture up to the anterior border of this tergite | Newportia spinipes + Newportia azteca |
| – | Forked paramedian sutures of tergite 1 ending up in the anterior transverse suture | 25 |
| 25 | Tarsus 2 of ultimate legs consists of 11–12 articles, prefemur laterally with strong setae | Newportia atoyaca |
| – | Tarsus 2 of ultimate legs consists of 5–8 articles, prefemur laterally with small spines | Newportia oriena |
| 26 | Tarsus of ultimate legs composed of uniform articles (Fig. 18) | 27 |
| – | Tarsus 1 and tarsus 2 of ultimate legs with different shapes (Fig. 19) | 29 |
| 27 | Tergite 1 with paramedian sutures in front of the anterior transverse suture (Fig. 14); tibia of ultimate legs cylindrical, tarsus composed of 7–9 articles (Fig. 18) | Newportia ignorata |
| – | Tergite 1 without paramedian sutures in front of the anterior transverse suture; tibia of ultimate legs distinctly claviform distally (Fig. 20), tarsus composed of ca. 15 articles (Newportia weyrauchi) | 28 |
| 28 | Prefemur of ultimate legs with 4 ventral spinous processes | Newportia weyrauchi weyrauchi |
| – | Prefemur of ultimate legs with 3 ventral spinous processes | Newportia weyrauchi thibaudi |
| 29 | Outer branches of the forked paramedian sutures of tergite 1 crossing the anterior transverse suture (Fig. 14) and ending onto the tergal anterior margin | 30 |
| – | Forked paramedian sutures of tergite 1 ending in the anterior transverse suture | Newportia monticola |
| 30 | Femur of ultimate legs medially with one basal and one distal spinous processes, ventrally without them. Sternites 2-12(15) each with a median longitudinal sulcus | Newportia fuhrmanni |
| – | Femur of ultimate legs medially with one basal spinous process, ventrally with 1-2 such processes. Sternites 2-19 each with a median longitudinal sulcus | Newportia simoni |
| 31 | Ultimate legs with a well-developed claw-shaped pretarsus (which is as long as the poorly distinct ultimate article of tarsus 2) | Newportia amazonica |
| – | Ultimate legs normally without claw-shaped pretarsus (if a small ultimate claw is present, then it is less than half as long as the poorly distinct ultimate article of tarsus 2) | 32 |
| 32 | Tibia of ultimate legs with 3 ventral spinous processes; medial spinous processes of prefemur almost as large as the ventral ones | Newportia mexicana |
| – | Tibia of ultimate legs without spinous processes, medial spinous processes of prefemur (when present) considerably smaller than the large ventral ones | 33 |
| 33 | Tergite 1 with rounded anterior transverse suture and with or without paramedian sutures | 34 |
| – | Tergite 1 with anterior transverse suture angulated caudad to midline and giving rise to short longitudinal suture, which bifurcate caudally (fig. 1 in |
Newportia troglobia |
| 34 | Tergite 1 without paramedian sutures between its posterior margin and the anterior transverse suture, rarely with very short tracks just behind the anterior transverse suture (Fig. 13) | 35 |
| – | Tergite 1 with complete (more rarely half-complete) paramedian sutures | 37 |
| 35 | Paramedian sutures of cephalic plate very short. Forcipular coxosternite without median suture. Ultimate sternite without median longitudinal sulcus. Femur of ultimate legs with a row of 2-3 spinous processes | 36 |
| – | Paramedian sutures of cephalic plate almost reaching its middle. Forcipular coxosternite with a well-developed median suture. Ultimate sternite with a clear median longitudinal sulcus or depression. Femur of ultimate legs without or with a single very small spinous process | Newportia maxima |
| 36 | Tarsus of ultimate legs uniform, without distinct division into tarsus 1 and 2. Forcipular trochanteroprefemur without process | Newportia lasia |
| – | Tarsus of ultimate legs distinctly divided into a shorter tarsus 1 and a longer tarsus 2 (Fig. 21). Forcipular trochanteroprefemur with a process | Newportia patavina |
| 37 | Paramedian sutures of tergite 1 not bifurcate | 38 |
| – | Paramedian sutures of tergite 1 bifurcate | Newportia pelaezi |
| 38 | Anterior transverse suture of tergite 1 interrupted between the paramedian sutures (Fig. 22) | Newportia divergens |
| – | Tergite 1 with a complete anterior transverse suture (Figs 15, 17) | 39 |
| 39 | Tarsus of ultimate legs uniform | Newportia brevipes |
| – | Tarsus 1 of ultimate legs abruptly differing from the tarsus 2 | 40 |
| 40 | Cephalic plate with a transverse suture crossing the paramedian sutures near the posterior margin (Newportia ernsti) | 41 |
| – | Cephalic plate without transverse suture (Fig. 13) | 42 |
| 41 | Prefemur of ultimate legs with 6 (rarely 7) ventral spinous processes. Transverse suture of cephalic plate very distinct | Newportia ernsti ernsti |
| – | Prefemur of ultimate legs with 5 ventral spinous processes. Cephalic plate: median part of transverse suture between the paramedian sutures often poorly visible | Newportia ernsti fossulata |
| 42 | Paramedian sutures of tergite 1 ending up in the anterior transverse suture; tergite 2 with complete paramedian sutures | Newportia bielawskii |
| – | Paramedian sutures of tergite 1 complete, crossing the anterior transverse suture; tergite 2 with short paramedian sutures or sutures totally missing | 43 |
| 43 | Tergite 2 without paramedian sutures (these begin from tergite 5-6 onwards); four basal articles of tarsus 2 of ultimate legs definitely separated from each other (fig. 9 in |
Newportia albana |
| – | Tergite 2 with shortened paramedian sutures (Fig. 22); all articles of tarsus 2 of ultimate legs not well separated | 44 |
| 44 | All legs with a tarsal spur and both lateral and ventral tibial spurs | Newportia stolli |
| – | All legs with a lateral tibial spur only | Newportia paraensis |
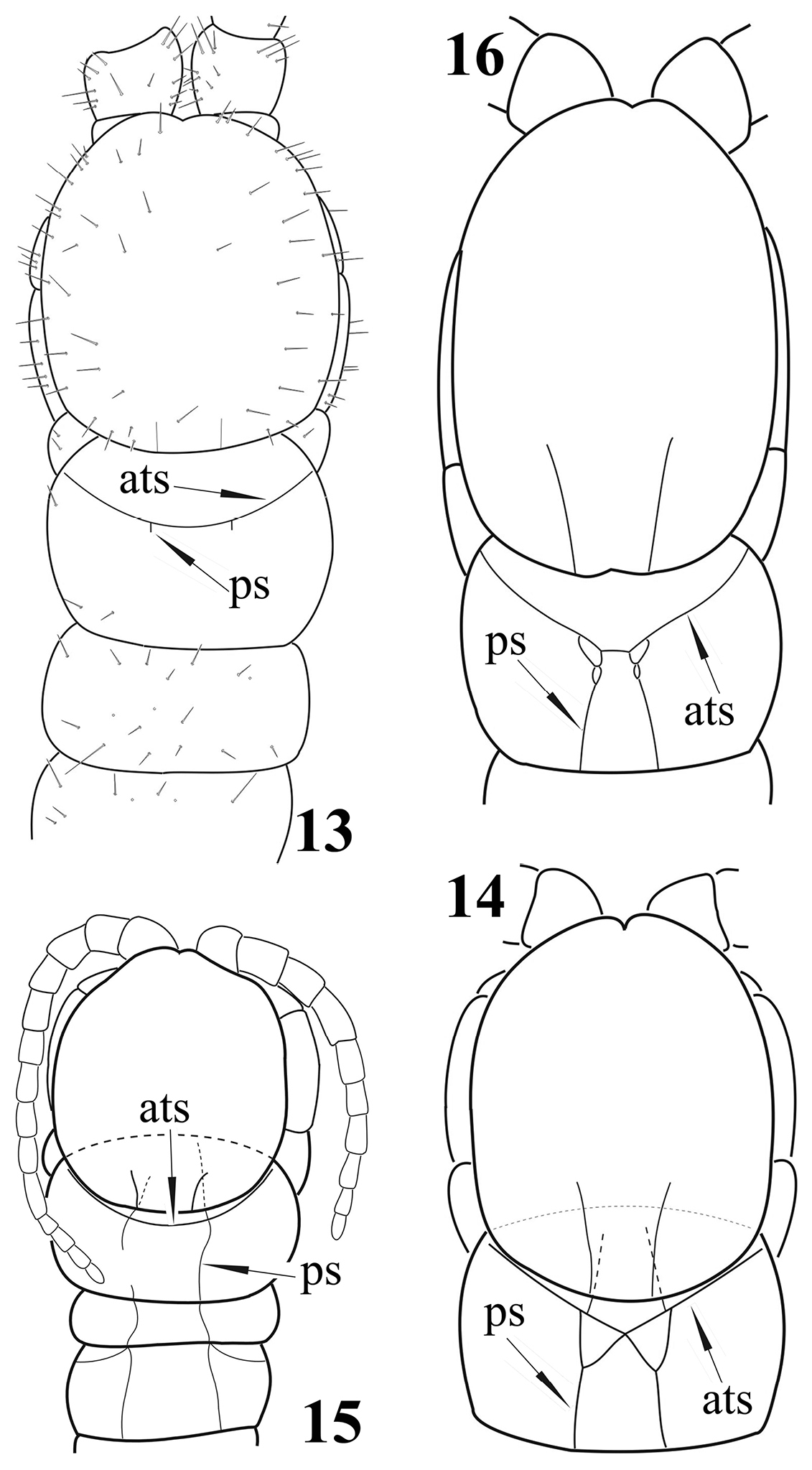
Newportia sp. 13 Cephalic plate and tergites 1-3, dorsal view (after
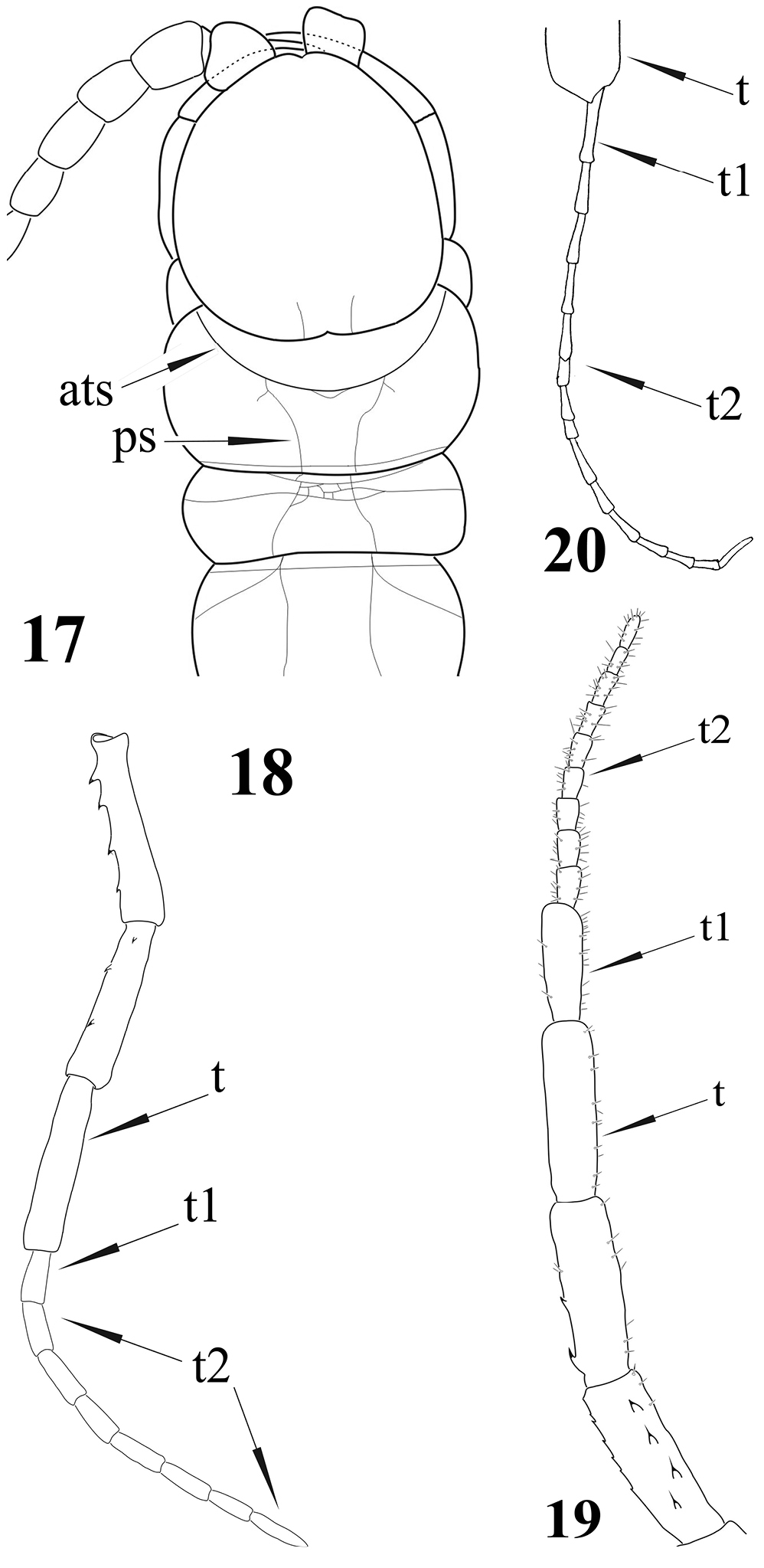
Newportia adisi Schileyko & Minelli, 1998 17 Cephalic plate and tergites 1-3, dorsal view (after
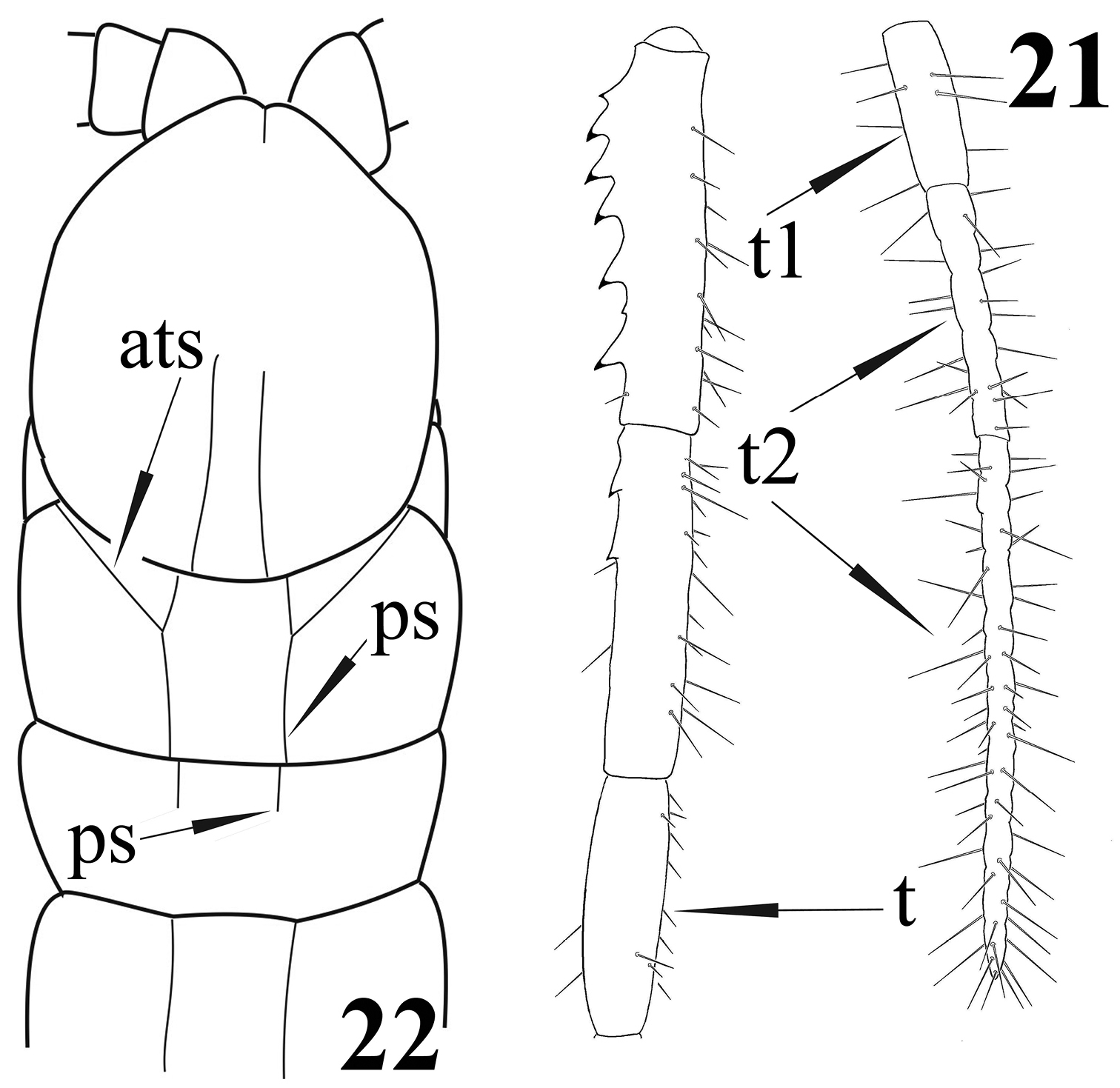
Newportia patavina Schileyko & Minelli, 1998 21 Right ultimate leg, medially (after
I am very grateful to Prof. Dr. Alessandro Minelli, Dr. Amazonas Chagas-Jr. and Dr. John Lewis for their valuable comments and constructive criticism that helped me improve the quality of the paper. My sincerest thanks also to my father, Prof. Dr. Anatoly A. Schileyko (Moscow Institute of Problems of Evolutionary Morphology and Ecology of Animals), for correcting the English of the earlier version of the manuscript. I would also like to thank Dr. Pavel Stoev for his invaluable support during the preparation of the paper. Open access to this paper was supported by the Encyclopedia of Life (EOL) Open Access Support Project (EOASP).