





(C) 2013 Kambiz Minaei. This is an open access article distributed under the terms of the Creative Commons Attribution License 3.0 (CC-BY), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
For reference, use of the paginated PDF or printed version of this article is recommended.
Haplothrips herajius sp. n. is described from leaves and flowers of a species of Suaeda in the south of Fars Province, Iran. This is the second Iranian species of Haplothrips with the unusual character state of extra setae on the metanotum. Information on variation in color and structure of the new species is provided. The similarities and host plant associations of this new species and Haplothrips kermanensis are discussed, as both are phytophagous on species of Chenopodiaceae.
Haplothrips, new species, phytophagous, Suaeda sp.
Traditionally and still widely accepted, the known species of thrips are placed in a single order, the Thysanoptera, within which two suborders are recognized, the Terebrantia and Tubulifera (
Among 20 genera of Phlaeothripidae recorded from Iran so far (
The new species discussed below was collected by beating leaves and flowers of Suaeda sp. (Chenopodiaceae) onto a plastic tray. The specimens were removed with a fine brush into a collecting vial containing 90% ethyl alcohol. They were then mounted onto slides in Canada balsam using a form of the protocol given by
urn:lsid:zoobank.org:act:FFCC20A9-BDC2-4700-B252-C87F008BA82B
Holotype female, Iran, Fars Province, Mohr, Heraj village; Suaeda sp. (leaves), 31.iii.2012. (Mohsen Abdolahi); Paratypes: 58 females, 11 males taken with holotype; 14 females, 3 males, same place, Suaeda sp. (flowers), 21. ix. 2012.
Female macroptera. Body brown (paler in summer forms), all tarsi, fore tibiae in distal half, distal apex of mid and hind tibiae are yellow; antennal segments I–II brown but the color of remaining segments variable depending on collecting date (III–VI yellow, VII–VIII yellow-brownish in summer forms; III yellow, IV–VIII yellow-brownish, gradually darker brown in spring forms); fore wing pale except for basal area; major body setae as well as sub-basal wing setae pale but tergite setae and anal setae slightly shaded at base.
Antennae 8-segmented, segment III with two, IV with four sensoria, VII slightly constricted at base, VIII short and broad at base (Fig. 1). Head a little longer than wide with maxillary stylets 0.2–0.3 of head width apart, retracted anterior to post ocular setae; post ocular setae blunt or capitate, extending to posterior margin of eye (Fig. 2). Cheeks weakly rounded. Maxillary bridge well developed. Mouth cone rounded.
Pronotum transverse, without sculpture lines except close to posterior margin; notopleural sutures complete; five pairs of developed setae present: am, aa, ml, epim and pa, all blunt or capitate (Fig. 2); prosternum with paired basantra and ferna as well as a spinasternum, ferna broad (Fig. 3). Mesonotum transversely weakly reticulate, with no microtrichia, lateral setae well developed, weakly capitate (Fig. 4). Mesopresternum eroded medially (Fig. 3). Metanotum reticulate, with no microtrichia, median setae slender and acute, arise on posterior half of sclerite, with 2–4 small setae on anterior half (Fig. 4). Fore tarsal tooth conspicuous (Fig. 2). Fore wing constricted medially (Fig. 6), sub-basal setae S1, S2 and S3 blunt or capitate, their bases arranged in a triangle (Fig. 5), 2–7 duplicated cilia present (Fig. 6).
Pelta triangular, weakly reticulate (Fig. 7). Tergite II–VII with wing-retaining setae, anterior pair weaker than posterior one, these being weakest on tergite II; tergites II–VII with a few lines of sculpture and 3–5 discal setae lateral to two pairs of developed wing-retaining setae; marginal setae S1 and S2 on tergites VII–IX long and finely pointed, S2 on other tergites finely pointed but S1 usually blunt, rarely finely pointed and sometimes variable on different tergites, tending to be more pointed on posterior than anterior tergites. Tergite VII with two campaniform sensilla not close to each other, separated by at least 0.1 width of tergite, with four micro-setae laterally; tergite VIII campaniform sensilla further apart, more than two times as those on tergite VIII, three to four micro-setae between sensilla or sometimes in front of them (Fig. 8). Tube short, about twice as long as basal width (Fig. 9); anal setae usually longer than tube.
Measurements.(holotype female, in microns). Body distended length 1845. Head, length 190; median width 180; postocular setae 35. Pronotum, length 35; width 68; major setae am 34, aa 33, ml 26, epim 51, pa 43. Fore wing length 700; sub-basal wing setae 41, 50, 68. Tergite IX setae S1 95, S2 85. Tube length 108; basal width 58. Antennal segments III–VIII length 38, 47, 43, 41, 33, 21.
Male macroptera. Color and structure similar to female. Sternites with no pore plates; tergite IX setae S2 short and stout (Fig. 10). Pseudovirga spoon shaped at apex (Fig. 11).
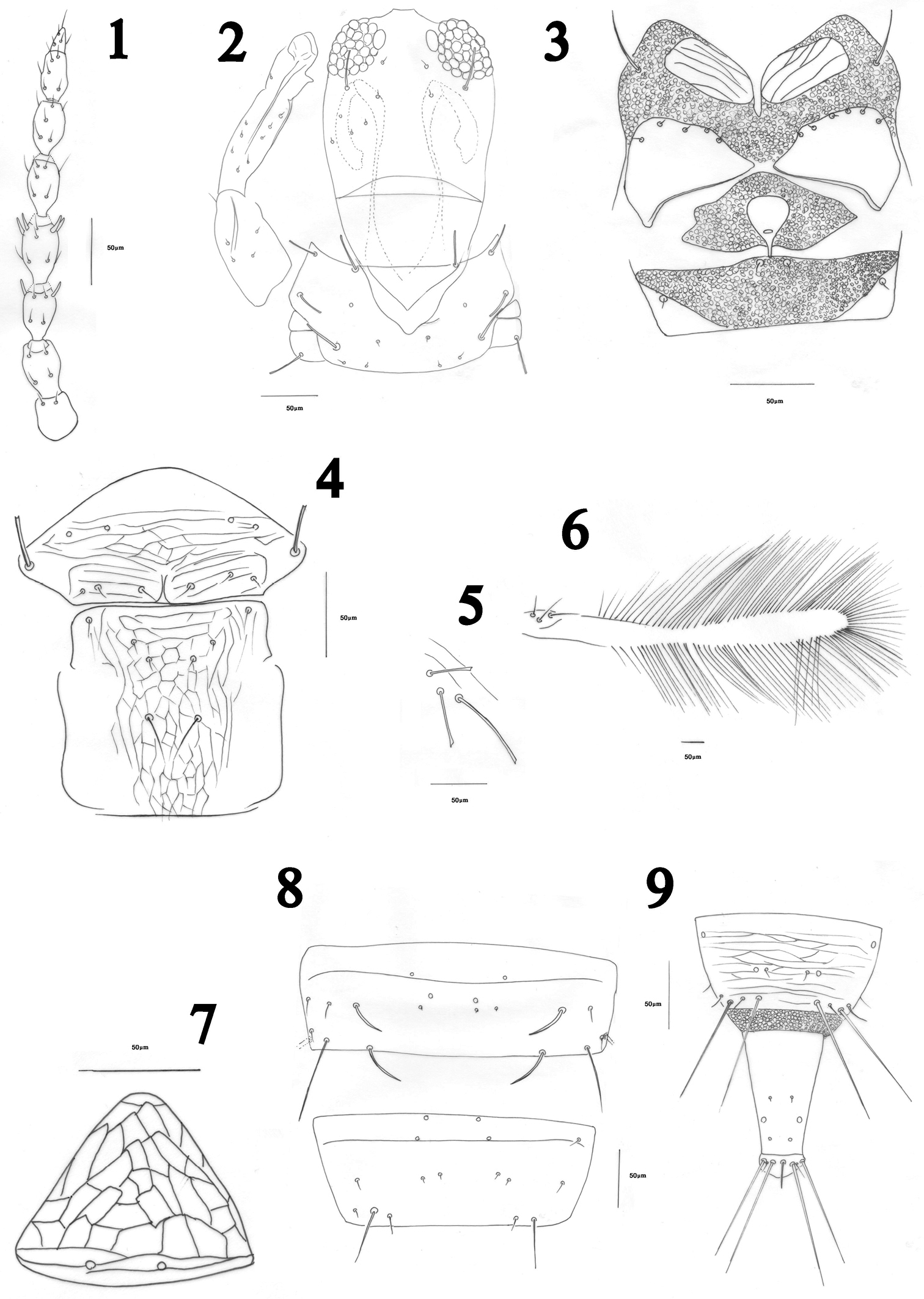
Haplothrips herajius sp. n. Female. 1 Antenna 2 Head and pronotum 3 Prosternum and mesopresternum 4 Mesonotum and metanotum 5 Sub basal wing setae 6 Forewing 7 Pelta 8 Tergites VII-VIII 9 Tergite IX and tube.
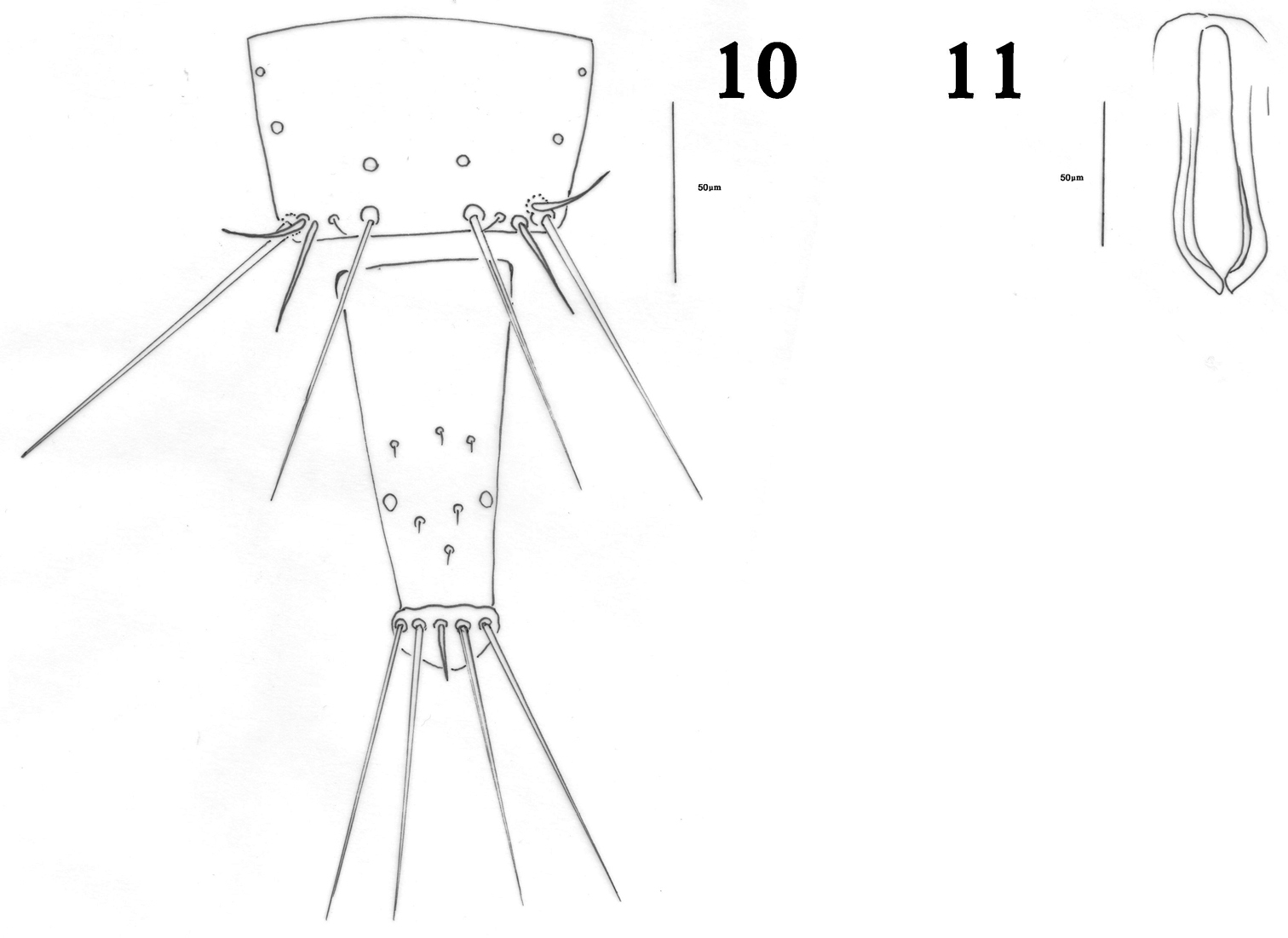
Haplothrips herajius sp. n. Male 10 Tergite IX and tube 11 Pseudovirga.
The reticulation on the mesonotum and metanotum of Haplothrips herajius (Fig. 4) is unique among Iranian Haplothrips as well as for most other Haplothrips species. In other species of Haplothrips recorded from Iran, this reticulation is weakly developed or absent. The new species is very close to Haplothrips kermanensis. Both species have extra setae on metanotum (Figs 4, 13) that are not seen in other Iranian species of Haplothrips. Moreover, in both species the basal wing setae are arranged in a triangle (Figs 5, 12) (this arrangement in Haplothrips kermanensis was not reported by
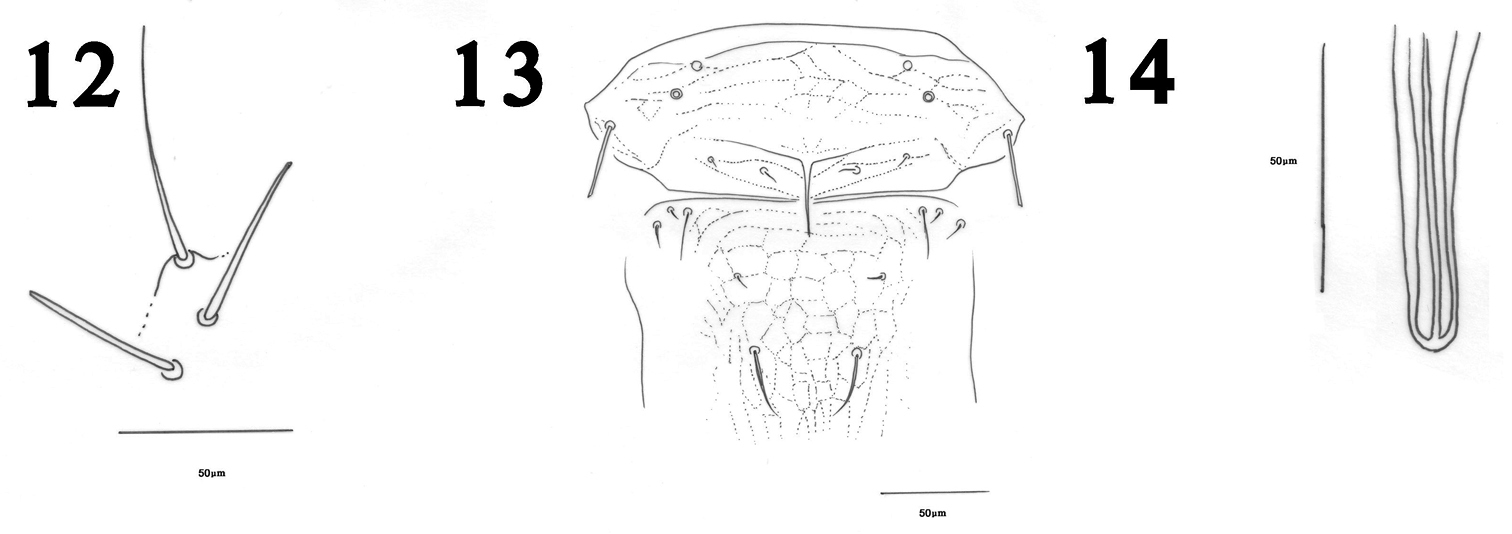
Haplothrips kermanensis.Female 12 Sub basal wing setae 13 Mesonotum and metanotum. Male 14 Pseudovirga.
Color of body and antennal segments varies among specimens, being paler in summer specimens compared with specimens collected in early spring. The fore tarsal tooth is conspicuous, but variable from small to large among male specimens. Maxillary stylets are retracted to postocular setae but rarely are low in the head and not reaching the postocular setae. Moreover, in a few specimens, the pronotal am setae are not developed.
Heraj is a village of Mohr city in the south of Fars Province, south of Iran which is located 300 km south of Shiraz, the capital of Fars Province.
The presence on two separate chenopod species of two Haplothrips species that share unusual character states, as discussed above, is interesting. The large number of collected specimens of Haplothrips herajius on both leaves (early spring) and flowers (late summer) of Suaeda sp. suggest that the new species is phytophagous, and apparently this plant species is a specific host for the new species in that area. Recently
Considering that most species of Chenopodiaceae bloom in summer months, the specimens studied here were collected at two seasons: in early spring on leaves and in late summer on flowers. The color of body and antennae differs between specimens collected at these two seasons as mentioned above. This difference is remarkable because the effect of temperature on body color during development was not noticed in any species of Haplothrips so far. However, in onion thrips, Thrips tabaci, a well-known pest of thripid family, experiments showed low temperatures during pupal development induce dark adult body color (
We are much indebted to Mr Mohsen Abdolahi and Ms Fatemeh Mohammadzadeh, BSc students of Plant Protection Department, Shiraz University, for collecting the materials of the new species described in this paperand sketching the illustrations respectively. Dr Ahmad Reza Khosravi, Department of Biology, Shiraz University, identified the plant on which the thrips were collected. The manuscript was improved through the advice and criticisms kindly provided by Dr Alice Wells (Australian Biological Resources Study, Canberra, Australia) and two anonymous reviewers. Open access to this paper was supported by the Encyclopedia of Life (EOL) Open Access Support Project (EOASP).