





(C) 2013 Azadeh Ghafarian. This is an open access article distributed under the terms of the Creative Commons Attribution License 3.0 (CC-BY), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
For reference, use of the paginated PDF or printed version of this article is recommended.
This paper report on a new species of mites of the genus Myrmozercon associated with ant in Iran – Myrmozercon cyrusi Ghafarian and Joharchi sp. n. was collected associated of the Monomorium sp. in Kenevist Rural District in the Central District of Mashhad County, Khorasan Razavi Province, Iran. This new species is described and illustrations provided. Myrmozercon ovatum Karawajew, 1909 is suspected to be a junior synonym of Myrmozercon brevipes Berlese, 1902 and host-specificity and host range of Myrmozercon are also reviewed.
Laelapidae, Myrmozercon, ants, taxonomy, Iran, myrmecophiles, nest
Many species Laelapidae have been reported from ants or their nests. The myrmecophiles genus Myrmozercon includes about 22 described species from Europe, Australia, Africa, Middle East, Transcaucasia, North America and Central Asia (
All species are associated with ants, except for one intercepted at quarantine on plant material (
Laelapidae associated with ants were collected mainly in Khorasan Razavi Province over a period of two years. Mites were removed from ants’ nests by hand picking or by extraction from ant nesting material using Tullgren funnels. Mites were cleared in Nesbitt’s solution and mounted in Hoyer’s medium. The nomenclature used for the dorsal idiosomal chaetotaxy is that of
http://species-id.net/wiki/Myrmozercon
The diagnosis of Myrmozercon used here is based on that of
urn:lsid:zoobank.org:act:5DD70D4C-1312-49FA-A881-C663EB04E98E
http://species-id.net/wiki/Myrmozercon_cyrusi
Figures 1–13Holotype, female, Kenevist Rural District in the Central District of Mashhad County, Khorasan Razavi Province, Iran, 36.97'N, 59.68'E, alt. 945 m, 25 April 2012, A. Ghafarian coll., in nest of Monomorium sp. (in YIAU). Paratypes, four females, same data as holotype (in JAZM and ANIC).
Figures 1–13. Dorsal idiosoma (Fig. 1). Length 522–534. Dorsal shield length 488–500, width 420–436 (n = 5). Shield posteriorly truncate, not covering entire idiosoma, leaving a curved strip of unprotected skin posterior to setae J5, shield without distinct reticulate ornamentation over whole surface; with 33 pairs of setae, 21 podonotal (z2 absent), 12 opisthonotal (Z4, S5 absent) and Z5 in soft skin posterior to shield, almost all setae except j1 and J4 slightly barbed in apical third or less, with club-like tip (Fig. 2), opisthonotal setae very long, reaching well past base of next posterior setae, dorsal shield setae increasing in length from anterior to posterior (j1 25-27, J1 54-59, J2 67-69, J3 79-82), without unpaired and asymmetrical seta, setae on shield uniform in length and thickness except j1 (25-27) and J4 (20-25) very fine and minute. A pair of very fine and minute setae in R series on the lateral soft skin but appear on ventral view. Shield with eight pairs of minute pores and lyrifissures including a pair of lyrifissures situated near z1, other pores inconspicuous.
Ventral idiosoma (Fig. 3). Tritosternum with short broad base (10-11 × 15-17 wide) fused to sternal shield, bifurcated at a short distance above suture, laciniae 37-40 in length, with smooth edges, strap-like and broad at base; pre-sternal shields fused with sternal shield. Sternal shield (length 248-255) narrowest between coxae II (104-108) widest between coxae II and III (218-22), with biconvex anterior margin and extending beyond level of st1, lateral margins thickened and posterolateral corners fused with endopodal shield; posterior margin concave; shield bearing three pairs of smooth pointed setae (st1 30-37, st2 40-45, st3 50-51) and two pairs of lyrifissures, one pair between setae st1 and st2 and the other between st2 and st3; surface with indistinct reticulate ornamentation. Seta st4 absent, metasternal pores also on extent of sternal shield but metasternal plates apparently absent. Genito-ventral shield wide, strongly tapering posteriorly, 320–346 long, 168–174 maximum width. Surface of shield smooth with longitudinal markings in anterior half; with one pair of simple setae st5 (35–37). Anal shield triangular, its anterior without lineate ornamentation, cribrum small, anal pores indistinct, bearing short post-anal seta 15-17 long, and a pair of para-anal setae 37–42 long. Opisthogastric skin with long, narrow metapodal plates (40–44 × 8–10 wide) and eight pairs of setae, almost all setae slightly barbed in apical third or less, each arising on small sclerotised platelet (Figs. 4, 5), (Jv1 47-50, Jv2 35-37, Jv5 64-73, Zv1 37-42, Zv2 45-50, Zv3 55-63, Zv4 45-50, Zv5 67-75). Peritreme very short (35-40), extending to posterior level of coxae Ш. Peritrematal shields absent, post-stigmatal section conspicuous, with one pair of pore.
Gnathosoma. Hypostomal groove with nine rows of denticles, 10 to 15 very fine denticles per row (Fig. 6). Hypostome with three pairs of setae, internal posterior hypostomal setae h3 longest, palp coxal setae absent; surface of hypostome ornamented with transverse and curved lines. Palp chaetotaxy: trochanter 1, femur 5, genu 5, tibia 12; all palp setae pointed, palp tarsal claw two-tined, dorsodistal edge of palp femur without swelling. Epistome triangular, smooth, with pointed apex (Fig. 7). Chelicera hyaline, fixed digit of chelicera reduced, with four minute terminal denticles, pilus dentilis, dorsal lyrifissure present (Fig. 8); movable digit weakly sclerotised, distally curved, with one small subterminal tooth and one stronger terminal tooth, cheliceral seta absent, arthrodial corona with hyaline flap without filaments (Fig. 8). Corniculi long, weakly sclerotised.
Legs: Legs II and III short (258–268, 268–272), I and IV longer (288-298). Chaetotaxy: Leg I: coxa 0 0/1 0/1 0, trochanter 1 0/1 1/1 1 (pd thick), femur 1 2/1 2/1 1 (ad1 and pd1 long, ventral setae all thick, Fig. 9), genu 1 2/1 2/1 1 (ad1 and pd1 long with club-like tip, ventral setae all thick, Fig. 9), tibia 1 2/1 2/1 1 (ad1 and pd1 long with club-like tip, ventral setae all thick, Fig. 9). Leg II: coxa 0 0/1 0/1 0, trochanter 1 0/1 0/2 1, femur 1 2/1 2/1 1 (ad1 and pd1 thick, al and pl long, Fig. 10), genu 1 2/1 2/1 1 (ad1 thick, pd1 long with club-like tip, al and pl long, Fig. 10), tibia 1 1/1 2/1 1 (al and pl long, Fig. 10). Leg III: coxa 0 0/1 0/1 0, trochanter 1 0/1 0/1 1 (al thick), femur 1 2/1 1/1 1 (ad1 thick, al and pl long, Fig. 11), genu 1 2/1 2/1 1 (ad1 and pd1 thick, ad1 in two paratypes club-like tip, al and pl long, Fig. 11), tibia 1 1/1 2/1 1 (al and pl long, Fig. 11). Leg IV: coxa 0 0/1 0/0 0, trochanter 1 1/2 0/1 0 (ad thick), femur 1 2/1 1/1 0 (al long Fig. 12), genu 1 2/1 2/1 1 (ad1 and pd1 thick, ad1 in two paratypes club-like tip, al and pl long, Fig. 12), tibia 1 1/1 2/1 1 (al and pl long, Fig. 12). Tarsi I-IV with 16 setae, pre-tarsi with membranous ambulacrum, claws absent.
Genital structures: Insemination ducts opening on posterior margin of coxa III; sacculus an irregular, dark coloured mass behind coxae IV, ducts entering sacculus via a pair of circularopenings (Fig. 13).
The species is named in memory of Cyrus the Great (Old Persian: Kuruš; c. 600 BC or 576 BC–530 BC) was the first Achaemenian Emperor of Persia, as the “father of the Iranian nation”, who issued a decree on his aims and policies, later hailed as his charter of the rights of nations.
Myrmozercon cyrusi differs from all other species in the genus by its very short peritreme, palp coxal setae absent, genua I-IV with similar chaetotaxy (1 2/1 2/1 1) and trochanter of palp with only one ventral seta.
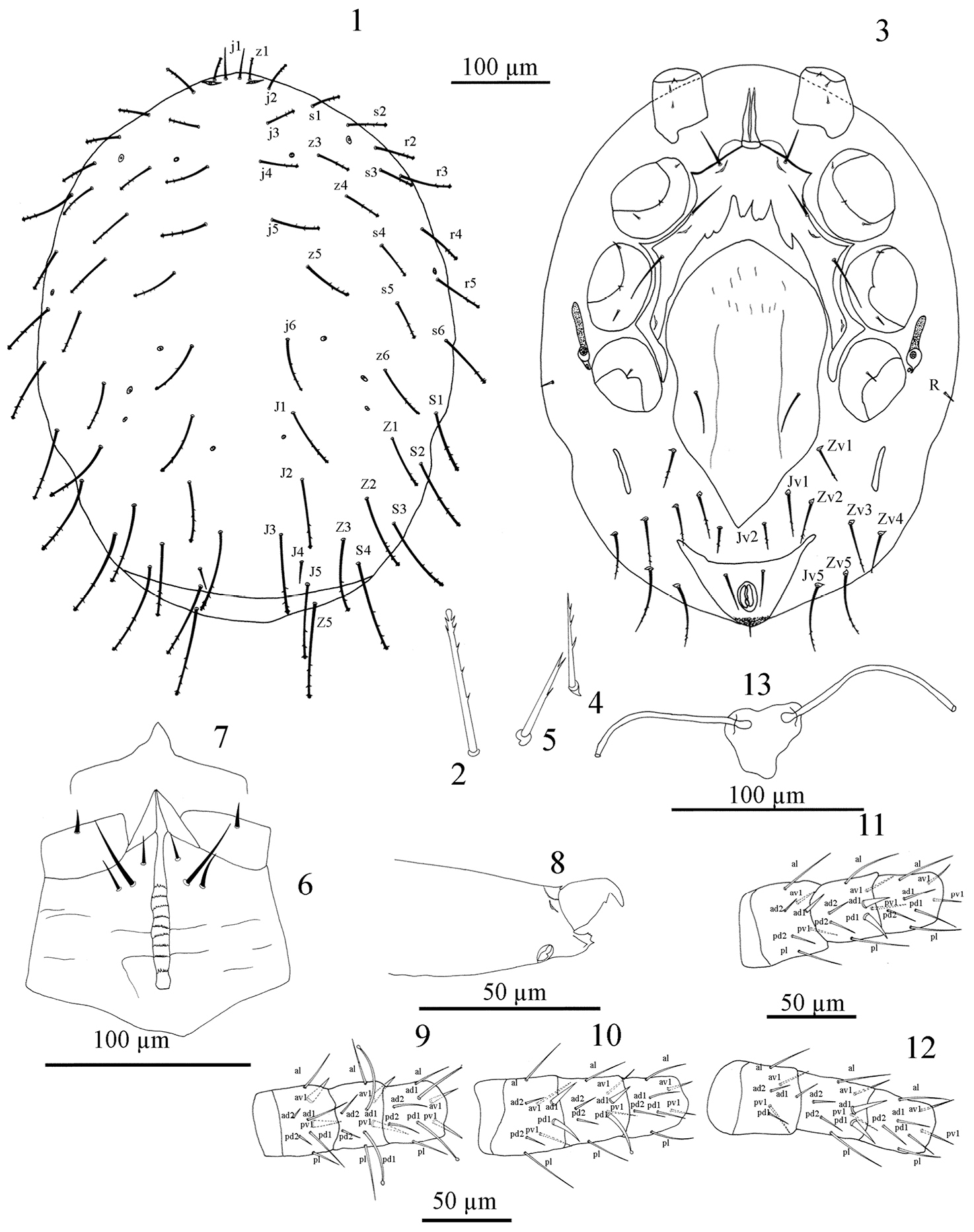
Myrmozercon cyrusi Ghafarian and Joharchi sp. n., female. 1 Dorsal shield 2 Dorsal seta enlarged (J5) (not to scale) 3 Ventral idiosoma 4–5 Opisthogastric setae enlarged (not to scale) 6 Hypostome 7 Epistome 8 Chelicera 9 femur, genu and tibia I, dorsal aspect 10 femur, genu and tibia II, dorsal aspect 11 femur, genu and tibia III, dorsal aspect 12 femur, genu and tibia IV, dorsal aspect 13 Insemination structures.
Only eleven species of Myrmozercon have been described from the Palaearctic Region (Myrmozercon acuminatus (Berlese, 1903) on Messor capitatus (Latereille, 1798) from Italy; Myrmozercon antennophoroides (Berlese, 1904) on Camponotus aethiops (Latereille, 1798) from Italy; Myrmozercon brachiatus (Berlese, 1903) on Messor capitatus from Italy; Myrmozercon brevipes Berlese, 1902 on Tapinoma erraticum (Latereille, 1798) from Italy; Myrmozercon clarus (Hunter and Hunter, 1963) on Crematogaster clara Mayr from Georgia; Myrmozercon diplogenius (Berlese, 1903) on Camponotus aethiops from Italy; Myrmozercon flexuosa (Michael, 1891) on Camponotus herculeanus (L., 1758); Myrmozercon Karajensis Joharchi et al., 2011 on Camponotus sp. from Iran; Myrmozercon liguricus Vitzthum, 1930 on Crematogaster scutellaris (Olivier, 1792) from Germany; Myrmozercon ovatum Karawajew, 1909 one from a worker Myrmecocystus emeryi Karawajew, 1909, but mostly on workers of Tapinoma erraticum nigerrimum from Turkmenistan; Myrmozercon tauricus Trach & Khaustov, 2011 on Crematogaster schmidti (Mayr, 1853) from Ukraine). Three subfamilies and seven genera of ants have been reported as hosts from the world: Formicinae, Camponotus, Cataglyphis, Polyrhachis; Dolochoderine, Iridomyrmex, Tapinoma; Myrmecinae, Crematogaster, Messor. Myrmozercon cyrusi has been collected in association with Monomorium sp. and this is the first record of ant host.
According topublications, Myrmozercon ovatum Karawajew, 1909 shares many compelling characters with Myrmozercon brevipes Berlese, 1902 especially form of genital shield, short peritreme, short legs, dorsal shield highly hypertrichous and collecting on same host, but we have not had the opportunity to examine type specimens of these two species therefore we consider the Myrmozercon ovatum to be a suspected synonym of Myrmozercon brevipes.
The biology of Myrmozercon species has not been studied yet. However, instability in morphology, the edentate chelicerae and short peritremes might suggest that Myrmozeron is parasitic on its ant hosts, and not simply a commensal in its host’s nests, but this has not been established experimentally.
We are grateful to Dr. Steven O. Shatuck (CSIRO Entomology, Canberra, Australia) for help with identifying the host ant of the mite. Special thanks go to Dr. Bruce Halliday (CSIRO Entomology, Canberra, Australia) for constructive comments during preparation of the manuscript. We are very grateful to the reviewers for their very useful comments.