





(C) 2013 Richard F. Ayrey. This is an open access article distributed under the terms of the Creative Commons Attribution License 3.0 (CC-BY), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
For reference, use of the paginated PDF or printed version of this article is recommended.
A new species of the vorhiesi group of Vaejovis C.L. Koch, 1836, Vaejovis brysoni sp. n., is described from the Santa Catalina Mountains in southern Arizona. Vaejovis deboerae Ayrey also inhabits this mountain range, making this the first documented case of two vorhiesi group species distributed on the same mountain. When compared to all other vorhiesi group species, Vaejovis brysoni sp. n. is distinct based on several combinations of morphological characters and morphometric ratios.
Madrean sky islands, speciation, taxonomy, Vaejovis vorhiesi group
For over 50 years, only four species of montane scorpions in the specious genus Vaejovis were known from the topographically complex states of Arizona, New Mexico and Sonora. That number has more than doubled over the past six years, with a total of 13 species now known (see
The systematics adhered to in this paper follow the classification established in
Acronyms of depositories — RFA, Richard F. Ayrey; MES, Michael E. Soleglad; USNM, United States National Museum, Smithsonian.
Material –In addition to the type material listed below for the new species, the following additional specimens were examined:
Vaejovis brysoni sp. n. USA: Arizona: Pima Co.: above Molino Basin on Catalina Highway near Seven Cataracts Vista, Santa Catalina Mountains. 32.35796°N, 110.72538°W, 1626 m. 16 March 2012. R.W. Bryson Jr. 1 ♂, 7 ♀ (RFA). Same locality. 5 April 2012. R.W. Bryson Jr. & D. Hartman 8 ♀ (RFA). Same locality. 18 August 2012. R. F. Ayrey & M. DeBoer-Ayrey. 8 ♀ (RFA).
Vaejovis cashi Graham, 2007. USA: Arizona: Cochise Co.: Cave Creek Canyon, Chiricahua Mountains. 2 August 2008. R. F. Ayrey & M. M. DeBoer-Ayrey 4 ♂, 4 ♀ (RFA). Same locality. 23 August 2011. R. F. Ayrey & M. M. DeBoer-Ayrey 3 ♂, 4 ♀ (RFA).
Vaejovis crumpi Ayrey et Soleglad, 2011. USA: Arizona: Yavapai Co.: by Lynx Lake, Prescott. 14 August 2008. R. F. Ayrey & M. M. DeBoer-Ayrey 3 ♂, 5 ♀ topotypes (RFA). Same locality. 14 September 2009. R. F. Ayrey & M. M. DeBoer-Ayrey 4 ♂, 4 ♀ (RFA). Same locality. 8 August 2010. R. F. Ayrey & M. M. DeBoer-Ayrey 3 ♂, 5 ♀ (RFA).
Vaejovis deboerae Ayrey, 2009. USA: Arizona: Pima Co.: Rose Canyon Campground, Santa Catalina Mountains. 28 August 2011. R. F. Ayrey & M. M. DeBoer-Ayrey 3 ♂, 5 ♀ (RFA). Same locality. 29 August 2011. R. F. Ayrey & M. M. DeBoer-Ayrey 4 ♂, 4 ♀ (RFA).
Vaejovis electrum Hughes, 2011. USA: Arizona: Graham Co.: Upper Arcadia Campground, Mount Graham. 17 July 2009. R. F. Ayrey & M. M. DeBoer-Ayrey 2 ♂, 6 ♀ (RFA). USA: Arizona: Graham Co.: 9415 feet asl, Mt Graham Hwy., Mt. Graham. 18 July 2009. R. F. Ayrey & M. M. DeBoer-Ayrey 1 ♂, 4 ♀ (RFA).
Vaejovis feti Graham, 2007. USA: New Mexico: Meadow Creek, Black Mountains. 6 July 1978. M. H. Muma 4 ♂, 3 ♀ (MES).
Vaejovis halli Ayrey, 2012. USA: Arizona: Gila Co.: Mount Ord. 11 September 2010. R. F. Ayrey & M. M. DeBoer-Ayrey 2 ♂, 6 ♀, paratypes (RFA). Same locality. 2 May 2011. R. F. Ayrey & M. M. DeBoer-Ayrey 3 ♂, 5 ♀, paratypes (RFA).
Vaejovis jonesi Stahnke, 1940. USA: Arizona: Coconino County: near Wupatki National Monument. 1 April 2011. R. F. Ayrey. 1 ♀ topotype (RFA).
Vaejovis lapidicola Stahnke, 1940. USA: Arizona: Coconino County: Red Sandstone Quarry, Flagstaff. 1 June 2011. R. F. Ayrey & M. M. DeBoer-Ayrey 1 ♂, 7 ♀ topotypes (RFA).
Vaejovis paysonensis Soleglad, 1973. USA: Arizona: Coconino County: Control Road, 25 miles East of Payson. 5 July 2011. R. F. Ayrey & M. M. DeBoer-Ayrey 1 ♂, 7 ♀ topotypes (RFA). Same locality. 6 July 2011. R. F. Ayrey & M. M. DeBoer-Ayrey 2 ♂, 6 ♀ topotypes (RFA).
Vaejovis tenuipalpus Sissom et al., 2012. USA: Arizona: Mojave Co.: Getz Peak, Hualapai Mountains. 9 August 2009. R. F. Ayrey & M. M. DeBoer-Ayrey 1 ♂, 7 ♀ paratopotypes (RFA).
Vaejovis vorhiesi Stahnke, 1940. USA: Arizona: Cochise Co.: Miller Canyon, Huachuca Mountains. 24 May 2011. R. F. Ayrey & M. M. DeBoer-Ayrey 1 ♂, 7 ♀ topotypes (RFA). Garden Canyon, Huachuca Mountains. 26 August 2011. R. F. Ayrey & M. M. DeBoer-Ayrey 4 ♂, 6 ♀ (RFA). Lutz Canyon, Huachuca Mountains. 27 March 2011. R. F. Ayrey & M. M. DeBoer-Ayrey 2 ♂, 2 ♀ (RFA).
urn:lsid:zoobank.org:act:80FC6074-1CD9-4DED-B155-2F7FE348495C
http://species-id.net/wiki/Vaejovis_brysoni
Figs 1–10, 12; Table 1Female holotype.USA: Arizona: Pima Co.: above Molino Basin on Catalina Highway near Seven Cataracts Vista, Santa Catalina Mountains. 32.35796°N, 110.72538°W, 1626 m. 16 March 2012. R.W. Bryson Jr. (RFA specimen number 632, deposited in USNM). Paratypes. Same locality as holotype. 16 March 2012. R.W. Bryson Jr. 1 ♂ (RFA specimen number 633) 2 ♀ (RFA specimen numbers 634 and 635). 17 August 2012. R. F. Ayrey. 1 ♀ (RFA specimen number 643).
The specific epithet is a patronym honoring our colleague Dr. Robert W. Bryson, Jr., the collector of the holotype.
Relatively small-bodied scorpion from the Seven Cataracts Overlook area of the Santa Catalina Mountains, southern Arizona (total body length of the female holotype is 27.50 mm). Color is light to medium brown, light brown to yellow on the legs, with underlying dark mottling on carapace and mesosoma. Metasoma is light brown with darker carinae.
Significant characters that distinguish Vaejovis brysoni sp. n. from other known species in the vorhiesi group are described below.
Vaejovis jonesi, Vaejovis lapidicola, Vaejovis paysonensis, Vaejovis crumpi, and Vaejovis bigelowi all possess 7 inner denticles (ID) on the chela movable finger, not 6 as in Vaejovis brysoni sp. n. The new species can be distinguished from Vaejovis halli by having significantly larger metasomal segment L/W ratios on I, II, and V (Table 1). Vaejovis brysoni can be distinguished from Vaejovis bandido by having larger metasomal segment I L/W ratios in addition to larger fixed finger L/chela L ratios. Vaejovis brysoni sp. n. can be distinguished from Vaejovis deboerae by having a smaller and less-developed subaculear tubercle. Vaejovis brysoni sp. n. also have shorter total body lengths and shorter carapace lengths. In addition, Vaejovis deboerae have larger telson vesicle L/W ratios. However, Vaejovis brysoni sp. n. have larger metasomal segment I L/W ratios and larger fixed finger L/chela L ratios. Vaejovis brysoni sp. n. also have fewer pectinal teeth than Vaejovis deboerae. Vaejovis brysoni sp. n. can be distinguished from Vaejovis vorhiesi by having larger metasomal segments L/W ratios on I, II, and III. However, Vaejovis vorhiesi have larger chela L/W ratios. Vaejovis brysoni sp. n. also have fewer pectinal teeth than Vaejovis vorhiesi. Vaejovis brysoni sp. n. can be distinguished from Vaejovis cashi by having longer total body lengths, and larger metasomal segment L/W ratios on segments I, II, and III. In addition, Vaejovis brysoni sp. n. exhibit larger fixed finger L/chela L ratios.Vaejovis brysoni sp. n. can be distinguished from Vaejovis feti by having larger metasomal segment I and II L/W ratios. Additionally, Vaejovis brysoni sp. n. also have larger fixed finger L/chela L ratios than Vaejovis feti. Vaejovis brysoni sp. n. can also be distinguished from Vaejovis feti by having a higher number of pectinal teeth. Vaejovis brysoni sp. n. can be distinguished from Vaejovis electrum by having larger metasomal segment L/W ratios on segments I, II, III, and V. In addition, Vaejovis brysoni sp. n. also have larger fixed finger L/chela L ratios than Vaejovis electrum. Finally, Vaejovis brysoni sp. n. can be distinguished from Vaejovis tenuipalpus by having smaller metasomal segment L/W ratios on segments II, III, and IV. Vaejovis tenuipalpus also have larger femur, patella, and chela L/W ratios.
Color of the holotype is light to medium brown, light brown to yellow on the legs, with underlying dark mottling on carapace and mesosoma. Metasoma is light brown with darker carinae. Metasomal segments are slightly wider than the vesicle. Small spinoid subaculear tubercle is present (Fig. 1). The pedipalp fixed finger has 5 to 6 ID denticles and movable finger has 6 ID denticles. Carapace:Anterior margin of the carapace is slightly emarginated, the posterior margin is straight. The carapace is moderately granular, with three lateral eyes present on each side. The median furrow is moderate and traverses the entire length of the carapace. The ratio of the location of the median eyes on the carapace (anterior edge/carapace length 0.73/3.75) = 0.19; carapace length/width at median eyes 3.75/2.35 = 1.60. The carapace is longer than metasomal segmentV.Mesosoma: Tergites are moderately granular with vestigial median carina on tergites I–VI. Tergite VII with weak median carina on anterior third and strong dorsal lateral and lateral supramedian granular carinae. Sternites I–V are finely granular and without carinae. Sternite V with weak granular ventral lateral carinae on middle 1/3. Presternites are smooth. Spiracles are ovoid with median side rotated 35 degrees from posterior sternite margin. Sternites with variable number of microsetae. Pectines:Pectinal tooth count is11/12. All pectinal teeth have exterodistal angling with a large sensorial area. Middle lamellae are 6/6. Fulcra are present. Each fulcra with 1–3 central setae.Metasoma: The carapace of the holotype female is longer than the fifth metasomal segment. Ratio of segment I length/width 0.93; of segment II length/width 1.03; of segment III length/width 1.18; of segment IV length/width 1.50; of segment V length/width 2.32. Segments I–IV: dorsolateral carinae are strong and granular to slightly dentate, with the distal denticle of I–IV enlarged and spinoid. Lateral supramedian carinae I–IV are strong and crenulate, with enlarged spinoid distal denticle. Lateral inframedian carinae are moderately granular on posterior 4/5 of segment I, 4/5 of II, 1/2 of III, and nearly obsolete on segment IV. Ventrolateral carinae on segment I, II, and III are moderate and granular; on IV moderate, granular and slightly serrate. Ventral submedian carinae are weak on segment I, weak to moderate on II, moderate, granular to slightly serrate on III and IV. The dorsal and lateral intercarinal spaces are very finely granular. Segment I–IV: ventral submedian setae count is 3/3. Segment V: dorsolateral carinae are moderate and slightly serrate on anterior 1/3. Lateromedian carinae are weak to moderate and granular on basal 3/5, and obsolete on distal 2/5. Ventrolateral and ventromedian carinae are strong and crenate to serrate. Intercarinal spaces are finely granular. Ventrolateral setae count 4/4. Telson:Smooth with four pairs of large setae on the ventral surface, three large setae are along both lateral edges of the vesicle with numerous smaller setae. A small spinoid subaculear tubercle is present. Chelicerae: The dorsal edge of movable cheliceral finger with two subdistal (sd) denticles. Ventral edge is smooth, with well developed serrula on distal half. Pedipalps: Trichobothrial pattern type C (
Variability of fixed finger ID denticle count was found. For Vaejovis brysoni sp. n., fixed finger ID denticle counts ranged from 5 (n=3) to 6 (n=5). Variation also existed for female Vaejovis brysoni sp. n. in the number of pectinal teeth 11/11 (n=2), 11/12 (n=3), 12/11 (n=1), 12/12 (n=2) with a mean of 11.5 for females, and 13/14 for the paratype male (n=1). In addition, there was variation in the number of middle lamellae 5/5 (n=1), 6/6 (n=5), 7/6 (n=1), 7/7 (n=1) and for the paratype male 8/9 (n=1). The right hemispermatophore was extracted from the paratype male. The right hemispermatophore is 3.10 mm in total length, and its lamina is 1.20 mm in length and 0.39 mm in width. The hemispermatophore is lightly sclerotized near the dorsal trough, and possesses a subtle distal crest on the inner distal aspect of the lamina. The lamellar hook is strong and widely bifurcated, and emanates from the dorsal trough. A medium, defined truncal flexure is visible on the external aspect of the trunk/lamina juncture. The male paratype also posseses an area of reduced pigmentation (white patch) on the posterior ¼ of the third sternal plate. (
(mm).Female holotype: total length 27.5; carapace length 3.75; mesosoma length 8.13; metasoma length 15.63. Metasoma: segment I length/width/depth 1.81/1.94/1.38; segment II length/width/depth 2.00/1.94/1.25; segment III length/width/depth 2.06/1.75/1.31; segment IV length/width/depth 2.63/1.75/1.25; segment V length/width/depth 3.63/1.56/1.25. Telson: length 3.50; vesicle length/width/depth 2.25/1.25/1.06; aculeus length 1.25. Pedipalps: total length 10.44; femur length/width 2.69/1.00; patella length/width 2.75/1.06; chela length 5.00; palm length/width/depth 2.44/1.25/1.13; movable finger length 2.69; fixed finger length 2.56. Male paratype: total length 21.1; carapace length 2.75; mesosoma length 5.25; metasoma length 13.1; Metasoma: segment I length/width/depth 1.44/1.81/1.25; segment II length/width/depth 1.56/1.88/1.13; segment III length/width/depth 1.75/1.63/1.19; segment IV length/width/depth 2.38/1.63/1.19; segment V length/width/depth 3.25/2.26/1.13. Telson: length 2.69; vesicle length/width/depth 1.81/1.00/1.00; aculeus length 0.88. Pedipalps: total length 9.5; femur length/width 2.44/0.81; patella length/width 2.63/0.88; chela length 4.44; palm length/width/depth 1.88/1.06/1.06; movable finger length 2.69; fixed finger length 2.56.
Vaejovis brysoni sp. n. is known only from the type locality above Molino Basin on the Catalina Highway near the Seven Cataracts Vista, Santa Catalina Mountains, Arizona, USA. The type localities of the 12 described species in the vorhiesi group from Arizona and western New Mexico are shown in Figure 13. Vaejovis brysoni sp. n. is widely allopatric with Vaejovis halli, Vaejovis vorhiesi, Vaejovis cashi, Vaejovis feti, and Vaejovis electrum (Fig. 13). Vaejovis brysoni sp. n. and Vaejovis deboerae both occur within the Santa Catalina Mountains, and their ranges may overlap, perhaps along the mid-elevation pine-oak woodlands between 1800–1900 m.
The type specimens were found at night using a UV flashlight alongside the Catalina Highway. This area lies within open oak woodland and the transition zone from drier desert grassland to pine-oak woodland (
Morphometrics (mm) of female Vaejovis brysoni sp.n. versus other Vaejovis vorhiesi group species. Data on pectinal teeth with * from (
| Vaejovis brysoni Ratio Comparisons | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Vaejovis spp. | Vaejovis brysoni (8) | Vaejovis bandido (5) | Vaejovis halli (3) | Vaejovis deboerae (3) | Vaejovis vorhiesi (3) | Vaejovis cashi (3) | Vaejovis feti (3) | Vaejovis electrum (3) | Vaejovis tenuipalpus (3) |
| Total length Carapace length Ca L/MetV L | 22.88–27.50 2.88–3.75 0.88–1.03 | 24.65–27.75 3.12–3.38 0.97–0.98 | 21.87–23.43 2.90–3.18 0.98–0.99 | 29.64–33.14 3.79–4.38 0.89–0.97 | 24.62–26.55 3.21–3.39 0.98–1.06 | 20.90–22.10 2.86–3.11 0.96–1.14 | 22.00–23.46 3.01–3.19 0.96–1.05 | 24.48–25.38 3.48–3.60 1.02–1.13 | 28.38–31.24 3.52–3.71 0.93–0.98 |
| Segment I length/width | 0.93–1.00 | 0.73–0.78 | 0.70–0.77 | 0.72–0.79 | 0.68–0.73 | 0.61–0.66 | 0.69–0.73 | 0.65–0.68 | 0.90–0.94 |
| Segment II length/width | 0.90–1.03 | 0.88–0.94 | 0.79–0.87 | 0.98–1.03 | 0.85–0.92 | 0.74–0.80 | 0.69–0.73 | 0.68–0.89 | 1.09–1.18 |
| Segment III length/width | 1.00–1.09 | 0.96–1.04 | 0.94–1.00 | 1.02–1.14 | 0.96–0.98 | 0.89–0.92 | 0.93–1.13 | 0.93–0.98 | 1.29–1.36 |
| Segment IV length/width | 1.34–1.61 | 1.35–1.44 | 1.27–1.50 | 1.48–1.60 | 1.39–1.62 | 1.28–1.39 | 1.35–1.60 | 1.33–1.46 | 1.74–1.83 |
| Segment V length/width | 2.15–2.82 | 2.02–2.16 | 1.79–2.11 | 2.10–2.32 | 2.08–2.22 | 2.05–2.15 | 2.07–2.24 | 1.81–1.91 | 2.49–2.52 |
| Telson VesicleLength/width | 1.60–2.06 | 1.59–1.69 | 1.68–1.85 | 2.22–2.43 | 1.63–1.72 | 1.56–1.71 | 1.62–1.87 | 1.45–1.62 | 1.60–1.77 |
| Femurlength/width | 2.87–3.22 | 2.86–3.07 | 2.83–3.33 | 2.74–2.90 | 2.87–3.22 | 2.74–3.02 | 2.89–3.41 | 2.78–3.19 | 3.81–3.83 |
| Patellalength/width | 2.47–3.29 | 2.83–3.21 | 2.89–3.63 | 2.91–3.16 | 3.12–3.19 | 2.86–3.03 | 2.53–2.66 | 2.86–2.94 | 3.72–3.85 |
| Chela length/width Ff L/Ca L Ff L/Ch L | 3.71–4.55 0.68–0.83 0.51–0.56 | 4.09–4.48 0.65–0.69 0.48–0.49 | 4.33–4.79 0.67–0.77 0.46–0.53 | 4.17–4.53 0.71–0.77 0.48–0.52 | 4.57–5.30 0.72–0.81 0.47–0.54 | 3.84–4.52 0.68–0.73 0.47–0.49 | 3.59–3.91 0.69–0.80 0.48–0.50 | 3.74–4.19 0.68–0.69 0.47–0.51 | 5.05–5.43 0.83–0.87 0.50–0.51 |
| Pectinal Teeth | 11–12 11.5(16) | 11–12 11.1(10) | 11–13 11.94(16) | 12–13 12.17(6) | 12–13 12.87(339)* | 10–12 10.98(337)* | 10 10.00(6) | 11–12 11.75(96)* | 11–12 11.15(26)** |

Vaejovis brysonisp. n., paratype female in natural habitat.

Vaejovis brysoni sp. n., paratype female dorsal and ventral views.

Vaejovis brysonisp. n., paratype female with first instar juveniles.

Vaejovis brysonisp. n., paratype male right hemispermatophore.
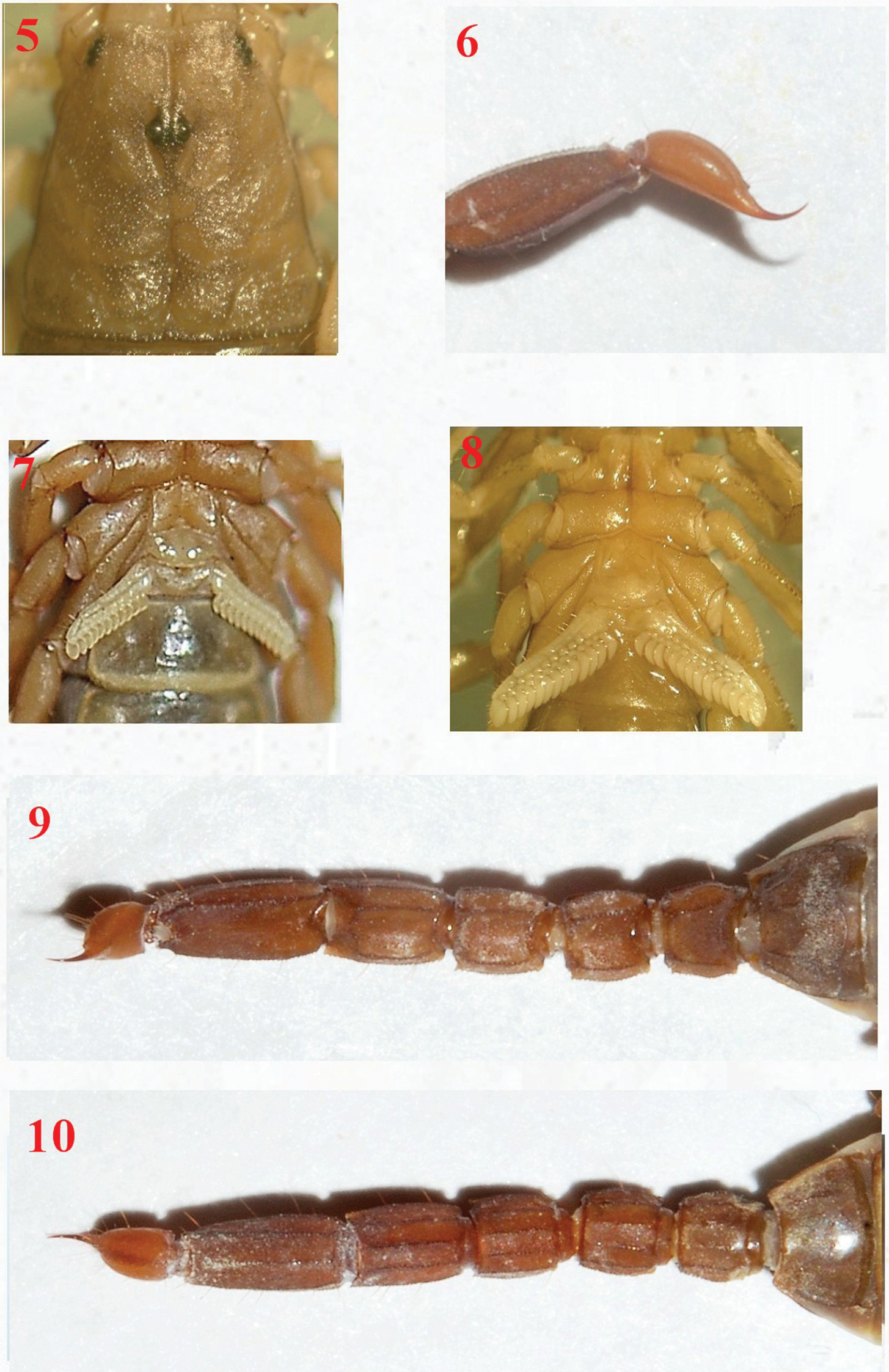
Vaejovis brysonisp. n., paratype female carapace 5; telson 6; pectines 7; paratype male pectines and sternites 8; paratype female metasoma dorsal 9; and ventral 10.

Vaejovis brysonisp. n.habitat.

Vaejovis brysonisp. n.trichobothrial pattern.
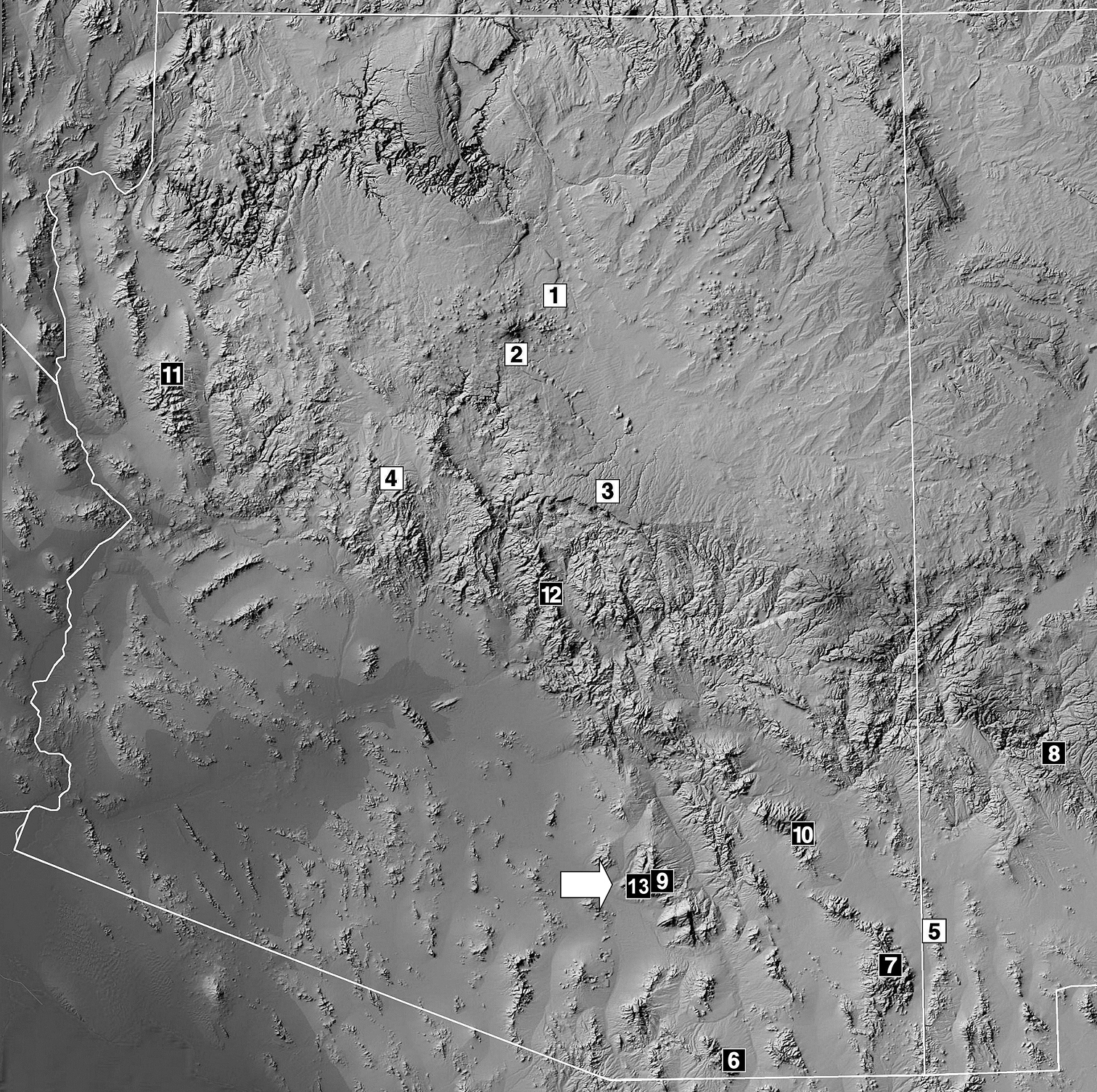
Map of Arizona and extreme western New Mexico showing the type localities of 13 of the 14 species in the Vaejovis vorhiesi group discussed in this paper, including the new species Vaejovis brysoni sp. n. The Vaejovis bandido type locality is south of Arizona in Sonora, Mexico. Localities are divided into those species exhibiting seven inner denticles (ID) on the chelal movable finger (white rectangles with black numbering) and those with primarily six ID denticles (black rectangles with white numbering). Seven IDs: 1 Vaejovis jonesi 2 Vaejovis lapidicola 3 Vaejovis paysonensis 4 Vaejovis crumpi and 5 Vaejovis bigelowi. Six IDs: 6 Vaejovis vorhiesi 7 Vaejovis cashi 8 Vaejovis feti 9 Vaejovis deboerae 10 Vaejovis electrum 11 Vaejovis tenuipalpus 12 Vaejovis halli and 13 Vaejovis brysoni sp. n.
We would like to thank Dr. Robert W. Bryson Jr. for originally discovering the scorpions, Dr. Matthew R. Graham for valuable comments on earlier versions of this manuscript and two anonymous reviewers.