





(C) 2013 Amazonas Chagas-Júnior. This is an open access article distributed under the terms of the Creative Commons Attribution License 3.0 (CC-BY), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
For reference, use of the paginated PDF or printed version of this article is recommended.
Seven species of the genus Rhysida Wood, 1862 from Venezuela and one subspecies from Peru described by Manuel Angel González Sponga and Wolfgang Bücherl respectively, are revised. Rhysida caripensis González-Sponga, 2002, Rhysida neoespartana González-Sponga, 2002, Rhysida guayanica González-Sponga, 2002, Rhysida maritima González-Sponga, 2002, Rhysida monaguensis González-Sponga, 2002, Rhysida porlamarensis González-Sponga 2002, Rhysida sucupanensis González-Sponga, 2002 and Rhysida celeris andina Bücherl, 1953 are junior synonyms of Rhysida celeris (Humbert & Saussure, 1870), which is redescribed and illustrated for the first time. Its geographic distribution is updated and a map showing its distribution is presented.
Neotropic, Venezuela, Taxonomy, Chilopoda, Rhysida
Rhysida is the second largest genus of the Otostigminae with circa 40 species in the Neotropics, the Indo-Malayan region and west, central and east Africa (
In a paper on myriapods from Venezuela,
http://species-id.net/wiki/Rhysida_celeris
Figures 1–10Venezuela –Rhysida caripensis – Holotype (MIZA0015958, ex. MAGS 680a) and paratype (MIZA0015958, ex. MAGS 680b), Edo. Monagas, Caripe, Caripe (10°12'00"N, 63°30'00"W), ix.1991, Dora Padrón de García leg.; Rhysida neoespartana – Holotype (MIZA0015954, ex. MAGS 319a) and paratype (MIZA0015954, ex. MAGS 319b), Edo. Nueva Esparta, La Asunción, Marcano (11°02'00"N, 63°51'46"W), 1988–1989–1991, Dora Padrón de Garcia leg.; Rhysida guayanica – Holotype (MIZA0015961, ex. MAGS 738a) and paratype (MIZA0015961, ex. MAGS 738b), Edo. Bolívar, Los Pijiguaos, Cedeño (06°35'20"N, 66°45'12"W), 01.v.1992, José Manuel Ayala leg.; Rhysida maritima – Holotype (MIZA0015957, ex. MAGS 580a), Edo. Carabobo, Goaigoaza, Puerto Cabello (10°29'36"N, 68°02'48"W) in 19.iii.1990, and paratype (MIZA0015963, ex. MAGS 1002), Edo. Carabobo, San Esteban, Puerto Cabello, in 18.x.1998, both collected by A. R. Delgado de González & Manuel Ángel González Sponga; Rhysida monaguensis – Holotype (MIZA0016084, ex. MAGS 970a) and paratype (MIZA0016084, ex. MAGS 970b), Edo. Monagas, Uverito, Sotillo (08°42'10"N, 63°22'00"W), 30.iv.1998, Oswaldo Fuentes leg.; Rhysida porlamarensis – Holotype (MIZA0015953, ex. MAGS 315a and paratype MIZA0015953, ex. MAGS 315b), Edo. Nueva Esparta, Porlamar, Calle San Rafael, Porlamar (10°57'30"N, 63°51'00"W), 30.iii.1988, Carlos Contreras leg.; Rhysida sucupanensis – Holotype (MIZA0015908, ex. MAGS 63a) and paratype (MIZA0015908, ex. MAGS 63b), Edo. Delta Amaruco, Sucupana, Casacoima (08°41'N, 61°48'W), 16.ix.1987, Pedro Delgado leg.
Jamaica – BMNH – 1 specimen; Dominican Republic – ZMH – 4 specimens, Sánchez, 5.vii.1905, C. Gagzol; ZMH – 1 specimen, Sánchez, Bay V. Samaná, xii.1894, Bock; Haiti – ZMH – 4 specimens, Port au Prince, 29.v.1901, Dr. Rouch; Montserrat – BMNH – 1 specimen, B.W.I., ii.1932; Costa Rica – ZMH – 1 specimen, Puerto Simón, 19.xi.1899, R. Mull; Suriname – ZMH – 2 specimens, Paramaribo, 28.v.1910, C. Hellesl; Colombia – IAVH – 1 specimen, Vichada, Cumaribo, Cgto. Santa Rita, PNN El Tuparro, 14-16-II-2004, I. Quintero, E. Gonzalez; IAVH – 1 specimen, Vichada, Cumaribo, Selva de Mataven, 31-III-02-IV-2007, L.E. Franco; IAVH – 1 specimen, Vichada, Gaviotas, 31-VI-1995; IAVH – 1 specimen, Caquetá, Solano, PNN Chiribiquete, 24-26-II- 2000, M. Ospina & E. González; ICN-M.Ch-0005, 1 specimen, Amazonas, Leticia, Via Torame, X-2000, Sist. Animal; ICN-M.Ch-0035, 1 specimen, Sucre, San Onofre, Boca cerrada, canal del dique, 1-IV-2000, E. Ulloa; ICN-M.Ch-0037, 1 specimen, Sucre, Galeras, Vereda corozera, 14-I-1999, E. Hernández; ICN-M.Ch-0039, 1 specimen, Meta, Vereda Apiay, sector el bosque villa Lolé , 23-XII-2001, M. Rojas; ICN-M.Ch-0097, 1 specimen, Casanare, Aguazul, Vda. El Charte, Finca Namaste, 20-IX-1996, Estudiantes Biol UN; ICN-M.Ch-0100, 1 specimen, Tolima, San Luis, 13-VI-1992, A. Castillo; ICN-M.Ch-0136, 1 specimen, Meta, San Martin, 1–15 -IV-2011, W. Galvis; ICN-M.Ch-0138, 1 specimen, Vaupés, Est. Biol. Caparù, 2002–2004, J. Pinzón; Ecuador – QCAZ – 3 specimens, Francisco Orelanna, La Joya de Los Sachas, 3.vi.2006, J. Mideros; QCAZ – 1 specimen, Francisco Orelanna, PN Yasumi, M.J. Tamariz; QCAZ – 1 specimen, Napo, Tena, 19.ix.2004; Argentina – ZMUC – 11 specimens, Riacho del Oro, 17.06.1899, W. Sorensen; ZMUC – 2 specimens, 19.08.1895.
General body color light blue or olive green, sternites and legs light blue or yellowish; prefemur and femur of the ultimate legs light blue, sometimes tibiae and tarsi are pale. Antennae with 17 to 21 articles, first two articles, dorsal surface and ¾ of ventral surface of third articles glabrous. Cephalic plate smooth, without sutures or depressions; tooth plates wider than high, 4+4, 4+5 or 5+5 teeth. Tergites smooth; complete paramedian sutures present from tergites 3–5 to 19–20, margination from tergites 5–9 to 21. Posterior border of tergite 21 ending in an obtuse angle. A pair of spiracles at 7th leg -bearing segment. Sternites 2, 3 or 4 to 19 with anterior incomplete paramedian sutures, but without depressions. Coxopleuron not prolonged, very short process with two small apical spines. Legs 1 to 16 (or 17) or 2 to 18 (sometimes 17) with two tarsal spurs, 19 (or 20) with one and 21 without; ultimate legs long, prefemur without spines.
Body length from 40 to 70 mm. General body color light blue or olive green, sternites and legs light blue or yellowish; prefemur and femur of the ultimate legs light or dark blue, but sometimes tibia and tarsi are pale or light blue. Antennae with 17 to 21 articles, first two and dorsal surface and ¾ of the ventral surface of the third article are glabrous; antennae reaching the posterior margin of tergite 5. Cephalic plate smooth, with a median sulcus but without sutures or depressions (Fig. 1), wider than long. Anterior margin of cephalic plate right-angled, with four ocelli in each side, posterior margin slightly rounded (Fig. 1). Forcipular coxosternum without depressions (Fig. 2); tooth plates with 4+4, 4+5 or 5+5 teeth; the inner two teeth are closer to each other than to the external teeth (Fig. 3). Each tooth plate with a long seta in the center; the tooth plates with obtuse angled basal suture. Trochanteropreforal process well-developed, long and ending as a point, with three to five denticles laterally, these sometimes not visible (Fig. 4). Tergites smooth, wider than long. Tergite 1 without sutures, its anterior border overlapping the posterior border of the cephalic plate. Tergites 3–5 to 19–20 with complete paramedian sutures (Fig. 5). Tergites 5 to 21 marginate, usually from tergite 11 (Fig. 5), but sometimes also from tergite 16. The margination is clearly visible in the anterior part of the tergites; tergite 21 with a slight posterior depression and triangular posterior margin (Fig. 7). A pair of spiracles on the 7th leg -bearing segment. Sternites smooth, wider than long. Sternites 2 (4)-19 with short incomplete anterior sutures (Fig. 6). However, sometimes, they are overlapped by the posterior margin of the previous sternite and visible only at sternites 5–18. Sternite 21 longer than wide; converging caudad and with straight or slightly concave posterior margin (Fig. 8). Coxopleural pores of several sizes, and numerous; the pore-field occupies nearly all surface of coxopleuron except for its dorsal and posterior parts, which are free of pores, posterior part of coxopleuron with a longitudinal depression (Fig. 9); very short coxopleural process with two apical spines (Fig. 10); apical spines may be combined as 1+2 or 2+3. Leg 1 with one femoral spur and legs 1 and 2 with one tibial spurs; legs 1 to 16, 17 or from 2 to 18 with two tarsal spurs, 19 and 20 with one and 21 without. Sometimes legs 20 lack spurs. Pretarsi of all legs with two accessory spines. Ultimate legs long and slender (12 mm to 22 mm). Prefemur without spines.
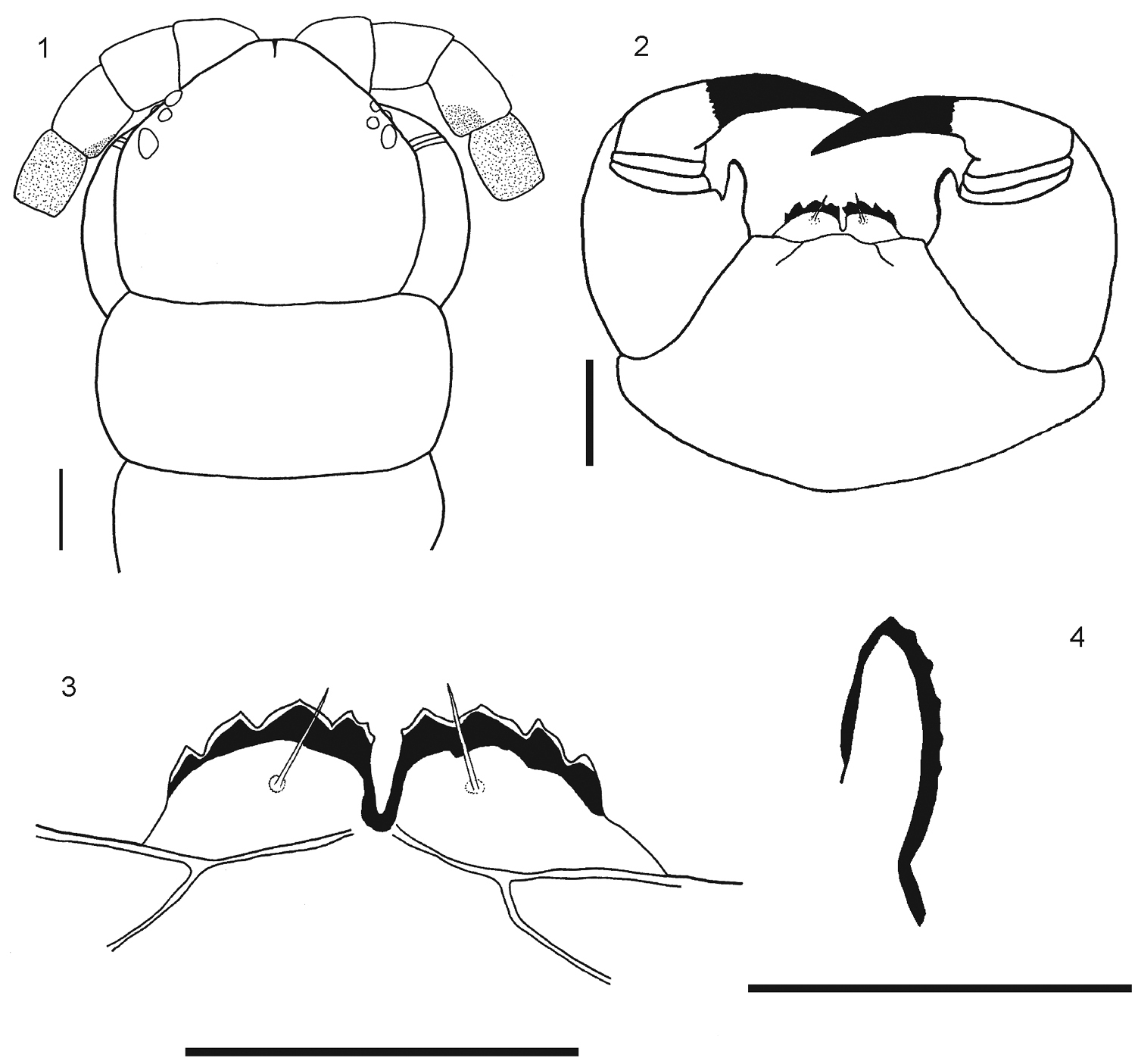
1 Rhysida celeris from Ecuador. Cephalic plate 2 Forcipular Coxosternum 3 Tooth plates 4 Forcipular trochanteroprefemur process. Scale bars 1 mm.
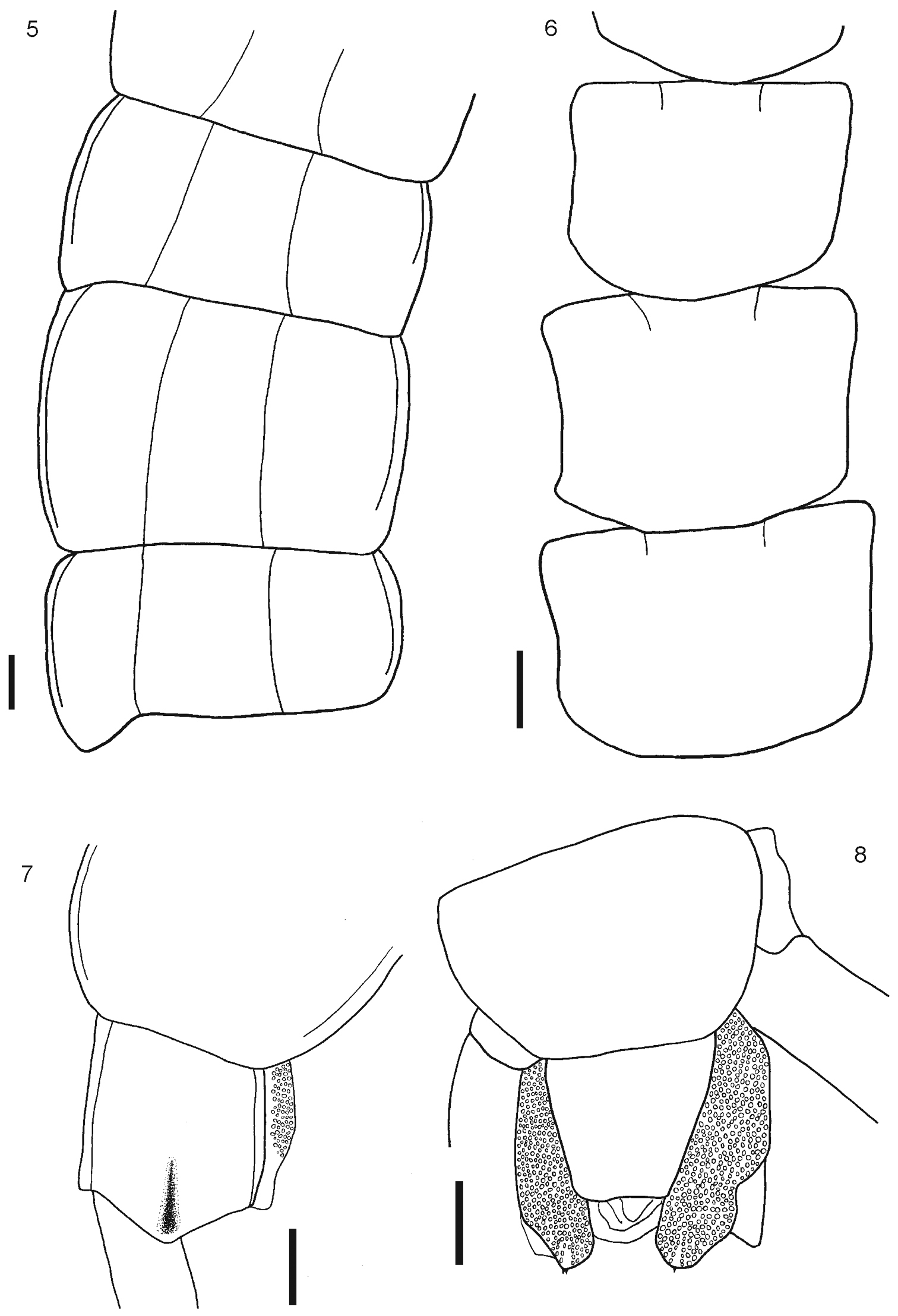
5 Tergites 11, 12 and 13 6 Sternites 4, 5 and 6 7 Tergite 21 8 Segment 21 showing sternite 21 and coxopleuron. Scale bars 1 mm.
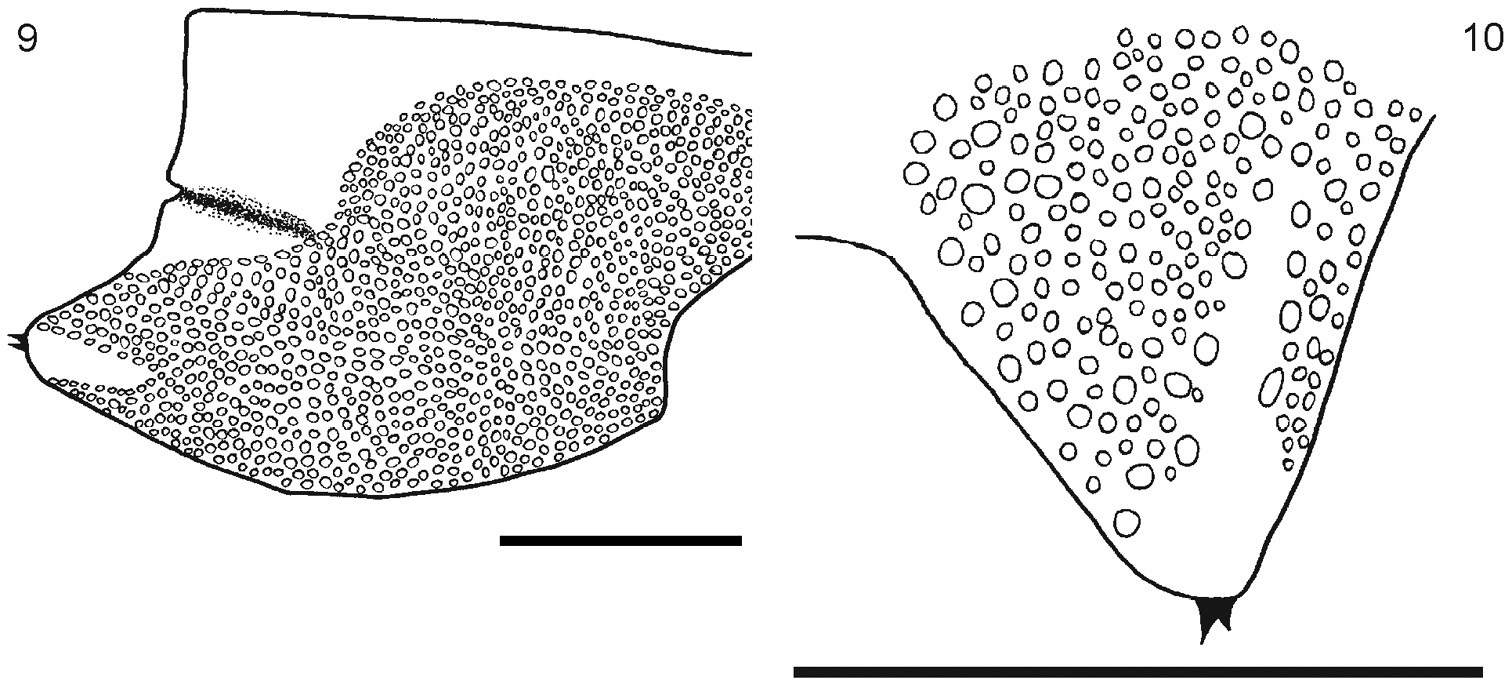
9 Segment 21 showing the coxopleuron 10 Detail of the terminal part of the coxopleuron showing the spines. Scale bar 1 mm.
Except for Rhysida maritima where the holotype and paratype were separated and given different numbers (e.g., holotype, MAGS 580a, and, paratype, MAGS 1002), the type specimens of all other species described by
The results of the review of the type-material described by
Characters of the species considered in this study. Information based on personal examination of types and original data (in parentheses) as published by
| Rhysida caripensis | Rhysida neoespartana | Rhysida guayanica | Rhysida maritima | Rhysida monaguensis | Rhysida porlamarensis | Rhysida socupanensis | Rhysida celeris andina | |
|---|---|---|---|---|---|---|---|---|
| Numbers of antenna articles | 19, (20) and 21 | (17), 19, 20, and 19+20 | 18, 19, 20 and (21) | 19, (20-21) and , 21 | 18, (19), 20 and 21 | 19, 20, 19+20 (17) | 20, (Not described) | (20) |
| Complete paramedian sutures on tergites | 3 to 20 (3 to 20) | 3 to 20 (2-3 to 20) | 3 to 20 (3 to 20) | 4 to 20 (4 to 20) | 7 to 20 (7 to 20) | 5 to 20 (5 to 20) | 3 to 20 (3 to 20) | (6 to ?) |
| Marginate tergites | 13 to 21 (16 to 21) | 16 to 21 (Posterior tergites) | 14 to 21 (Posterior tergites) | 16 to 21 (Posterior tergites) | 17 to 21 (Posterior tergites) | 18 to 21 (Posterior tergites) | 15 to 21 (16 to 21) | (16-19 to 21) |
| Incomplete sternite paramedian sutures | 2 to 18 (Without) | 1 to 18 (Without) | 2 to 18 (Without) | 4 to 17 (Without) | 3 to 17 (Without) | 3 to 18 (Without) | 2 to 18 (Without) | (1 to 20) |
| Tooth plates | 4+5 (5+5) | (4+4) 5+5 | 4+4, 4+5 and 5+5 (Not described) | 4+4 (4+4) | (3+3), 4+4, 4+5 and 5+5 | 4+4, 4+5 and 5+5 (Not described) | 5+5 (5+5) | (0+5 (damage)) |
| 2 spurs on tarsus 1 of legs | 1 to 17 and 18 (Not described) | 1 to 16, 17 and 18 (Not described) | 2 to 17 (Described only to leg 2) | 1 to 18 (Not described) | 1 to 18 (On all legs) | 1 to 18 (On all legs) | 1 to 18 (Not described) | (1 to 19) |
| 1 spur on tarsus 1 of legs | 18, 19 and 20 (Not described) | 17, 18, 19 and 20 (Not described) | 1, 19 and 20 (Described only to leg 1) | 19 and 20 (Not described) | 19 and 20 (Not described) | 19 and 20 (Not described) | 19 and 20 (Not described) | (20) |
| No tarsal spurs on legs | 21 (21) | 21 (21) | 21 (21) | 21 (21) | 21 (21) | 21 (21) | 21 (21) | (21) |
| Terminal spines on coxopleura | 2 (1) | 2 (2) | 2 (2) | 2 (2) | 1+2, (2) and 2+3 | 1+2 (2) | 2 (2) | (1) |
| Distribution | Venezuela | Venezuela | Venezuela | Venezuela | Venezuela | Venezuela | Venezuela | Peru |
His descriptions are based only on the holotypes, and he did not describe the variation of some characters, such as the number of antennal articles, the distribution of paramedian sutures and the margination of tergites and the number of teeth of forcipular tooth-plates. For example, complete paramedian sutures are present in all of the González-Sponga’s (2002) species, however, the distribution of sutures vary. The complete paramedian sutures on tergite 3 are not as evident as on tergite 4, but the sutures always end at tergite 20. The number of teeth in forcipular tooth-plates also vary from 3+3 to 5+5.
Two other important characters that were omitted or were poorly described by
Rhysida celeris was originally described from ‘Carolina’ (
Jamaica, Dominican Republic, Haiti, Nicaragua (
After reviewing the type material of the genus Rhysida described by
All taxa analyzed in this study are conspecific with Rhysida celeris which is redescribed and illustrated based on specimens from the Antilles, Central and South America. The characters used to determine Rhysida celeris and their variation are summarized in Table 2. Rhysida celeris is the most widespread species of the genus in the Neotropical Region and is found from the sea level up to1250 meters in the mountains. The species is herewith recorded from the Dominican Republic, Haiti, Montserrat, Suriname and Ecuador for the first time (Fig. 11). Morphologically, Rhysida celeris is close to Rhysida brasiliensis Kraepelin, 1903, but differs from the latter by having complete paramedian sutures on segments 3-19/20 (vs. incomplete and short ones on segments 4-19 in Rhysida brasiliensis;) shorter ultimate legs (18–22 mm vs. 20–32 mm in Rhysida brasiliensis). With present paper, the valid species of Rhysida in the Neotropical Region were reduced to seven, of which five indigenous (Rhysida celeris, Rhysida brasiliensis, Rhysida rubra Bücherl, 1939, Rhysida riograndensis Bucherl, 1939 and Rhysida chacona Verhoeff, 1944) and two introduced (Rhysida immarginata and Rhysida longipes). Both, Rhysida riograndensis, from southern Brazil, and Rhysida chacona, from Paraguay, resemble in many aspects Rhysida celeris, but the lateral tergal margins are absent in both species. As the presence of lateral margins is considered an important character in Neotropical Rhysida and since I have not seen yet the type-material of these two species, I prefer to keep them as a valid species.
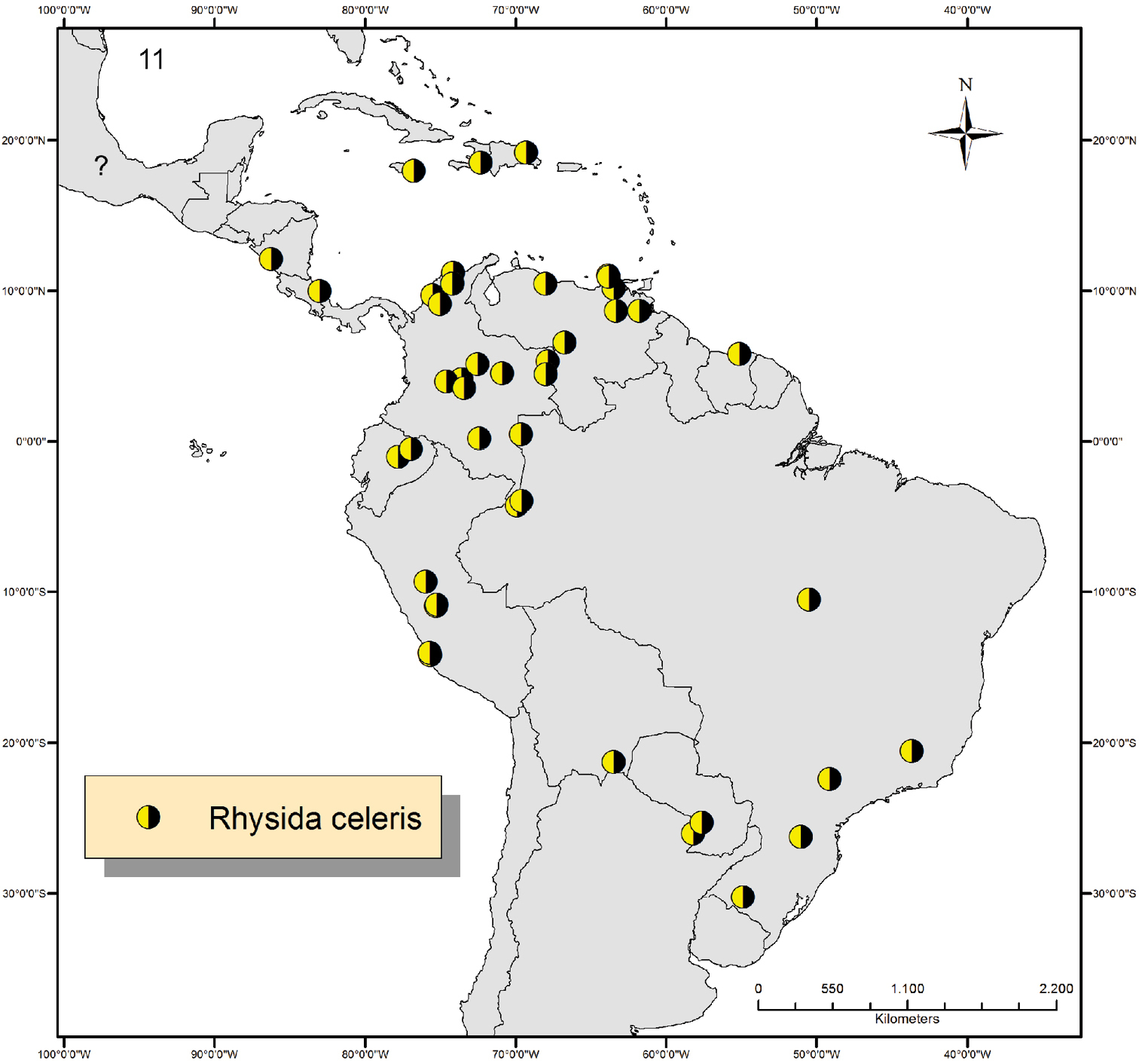
Map showing the distribution of Rhysida celeris in the Neotropical Region. Question mark indicates unspecified locality in Mexico.
Characters of Rhysida celeris showing the geographical variation.
| Rhysida celeris | Rhysida celeris | Rhysida celeris | Rhysida celeris | |
|---|---|---|---|---|
| Numbers of antennal articles | 17 to 21 | 18 to 21 | 17 to 21 | 19 to 20 |
| Complete paramedian sutures on tergites | 3 to 20 | 4 to 19 | 4 (5) to 19 | 3, 4 or 5 to 20 |
| Marginates tergites | 13 (14) to 21 | 5 to 21 | 10, 11, 12, 13 to 21 | 16 to 21 |
| Incomplete paramedian sutures on sternites | 3 (4) to 18 (19) | 4 to 19 | 4 to 19 | 2, 3 or 4 to 19 |
| Coxosternal Teeth | 4+4, 4+5 and 5+5 | 4+4, 4+5 and 5+5 | 4+4, 4+5 and 5+5 | 4+4, 4+5 and 5+5 |
| 2 spurs on tarsus 1 of legs | 1 to 16 (17 or 18) | 1 to 19 or 2 to 20 | 2 to 18 | 1 to 18 |
| 1 spur on tarsus 1 of legs | 19 and 20 | 1, 19 and 20 | 19 and 20 | 19 |
| No tarsal spurs on legs | 21 | 20 and 21 | 21 | 20 and 21 |
| Terminal spines on coxopleura | 1, 2 or 3 | 2 | 2 | 2 |
| Specimens studied | Venezuela | Colombia | Ecuador | Costa Rica, Jamaica, Dominican Republic, Haiti, Montserrat, Suriname, and Argentina |
I am very grateful to Hieronymus Dastych (ZMH), Osvaldo Villareal Manzanilla (MIZA), Henrik Enghoff (ZMUC), Janet Becalloni (BMNH), John Codington (NMNH), Claudia Alejandra Medina (IAVH), Álvaro Barragán and Mauricio Vega (QCAZ) for their kind support during my visits in the respective institutions and myriapod collections under their care. I also thank Elisa Chaparro Aguirre, Sebastian Galvis Jimenez, Hernan Dario Triana Pulido and Luis Edier Franco Galeano who have helped me to find and organize the centipede collections of ICN and IAVH. Both, Elisa Chaparro Aguirre and John Lewis have significantly improved the English of the manuscript. I also owe my sincere thanks to the two referees and the editor for their valuable comments that improved considerably the paper. This research was supported by Fundação de Amparo à Pesquisa do Estado de São Paulo (process 2010/10388-8).