





(C) 2013 Nesrine Akkari. This is an open access article distributed under the terms of the Creative Commons Attribution License 3.0 (CC-BY), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
For reference, use of the paginated PDF or printed version of this article is recommended.
Citation: Akkari N, Cheung DK-B, Enghoff H, Stoev P (2013) Revolving SEM images visualising 3D taxonomic characters: application to six species of the millipede genus Ommatoiulus Latzel, 1884, with description of seven new species and an interactive key to the Tunisian members of the genus (Diplopoda, Julida, Julidae). ZooKeys 328: 5–45. doi: 10.3897/zookeys.328.5763
A novel illustration technique based on scanning electron microscopy is used for the first time to enhance taxonomic descriptions. The male genitalia (gonopods) of six species of millipedes are used for construction of interactive imaging models. Each model is a compilation of a number of SEM images taken consecutively while rotating the SEM stage 360°, which allows the structure in question to be seen from all angles of view in one plane. Seven new species of the genus Ommatoiulus collected in Tunisia are described: O. chambiensis, O. crassinigripes, O. kefi, O. khroumiriensis, O. xerophilus, O. xenos, and O. zaghouani spp. n. Size differences between syntopic adult males of O. chambiensis and O. xerophilus spp. n. from Châambi Mountain are illustrated using scatter diagrams. A similar diagram is used to illustrate size differences in O. crassinigripes, O. khroumiriensis spp. n. and O. punicus (Brölemann, 1894). In addition to morphological differences, the latter three species display allopatric distribution and different habitat preferences. A dichotomous interactive key with a high visual impact and an intuitive user interface is presented to serve identification of the 12 Ommatoiulus species so far known from Tunisia. Updates on the North African Ommatoiulus fauna in general are presented.
Taxonomy, millipedes, Diplopoda, Ommatoiulus, new species, North Africa, interactive SEM images, interactive key
Description of new species is just one among many tasks of
taxonomists (
Enhancing and modernizing taxonomy constitutes one of the
main challenges of this century, and several pilot
projects and initiatives have been taken in this respect
(see
It is natural that the word “imaging” appears in two of
Costello et al.’s 14 action points, because nowhere is the
saying “a picture is worth a thousand words” more true
than in taxonomic descriptions. Early taxonomic works
often included excellent drawings. In the course of time,
drawings have been supplemented with photographs,
microphotographs, SEM micrographs, multi-focus images, and
(most recently) images produced by confocal laser scanning
microscopy (cLSM), optical projection tomography (OPT),
magnetic resonance imaging (MRI) and micro-Computed
Tomography scan (e.g.,
To demonstrate a new technique for visualization of
taxonomic characters described in detail in (
This work is part of an ongoing project of revising the
tribe Schizophyllini
(
The male copulatory organs, or gonopods, are of prime
importance for characterising millipede species and higher
taxa. There are exceptions, where different species have
identical or almost identical gonopods, e.g., several
genera of Juliformia, such as Nepalmatoiulus
Mauriès, 1983 (
As useful as gonopod illustrations are for taxonomic
descriptions, they can sometimes be grossly misleading and
result in misidentification and production of synonymic
names, the Achilles’ heel of descriptive taxonomy. There
are several examples of this in the literature about
millipedes, but most striking is perhaps the case
presented by
Useful taxonomic characters in Ommatoiulus are almost exclusively derived from the gonopods. Differences between species are often subtle, and the pronouncedly “3D” nature of the gonopods makes recognition of the differences difficult. In many older papers dealing with Ommatoiulus taxonomy, authors have dissected the different parts of the gonopods and have illustrated them separately which has led not only to “angle-of-view” problems, but also to difficulties of relating the various gonopod components spatially to each other. By applying the novel imaging technique we have overcome these problems.
Ommatoiulus
is the dominant genus of julid millipedes in North Africa
and the Iberian Peninsula. A total of 70 species have
been described so far, and many more remain to be
recognised and named. For example,
North African species of the genus were examined in
considerable detail by several authors, especially
Despite these contributions, the North African Ommatoiulus
fauna is far from being thoroughly assessed, nor is its
taxonomy close to being fully revised. Without doubt,
numerous new species still await discovery, and several
taxonomic questions still remain unsolved, such as the
correct placement of the highly deviating species Ommatoiulus
lapidarius
(Lucas, 1846), type species of the subgenus Apareiulus
Brölemann, 1897 (e.g.,
Most specimens were hand collected during spring 2008 by
N.A. and P.S. Supplementary material was obtained from
museum collections. All studied specimens are preserved in
70% alcohol. Measurements were made using a Leica Wild M10
microscope equipped with an ocular micrometre. Photographs
were taken using Visionary Digital’s BK Plus Lab with a
Canon EOS 7D. For scanning electron microscopy, parts of
the specimens were cleaned with ultrasound, transferred to
96% ethanol then to acetone, air-dried, mounted on
adhesive electrical tape attached to aluminium stubs,
coated with platinum/palladium and studied in a JEOL
JSM-6335F scanning electron microscope. Photographs were
processed with a Leica Application Suite program and final
stacking made with Zerene Stacker 1.04. The rotatable
images were constructed from 18 SEM images taken at 20
degrees intervals starting from the mesal view and
continuing until all 360 degrees were captured by rotating
the SEM stage. The images were processed using Adobe
Lightroom 4.3 by adjusting the black, highlight and white
levels to achieve a uniform background and contrast. Each
image was then cropped to ensure a smooth transition
between each frame during rotation. The images were
imported into Adobe Flash CS5, where each image was made
into a single frame and the series combined to form a
rotating animation. The animation controls (moving from
one frame to the next) were mapped to the mouse cursor
using Actionscript 3.0. The html version available online
was compiled using Magic 360. The interactive key was
developed in Adobe Flash CS5.5 using Actionscript 2.0 to
handle screen transitions and image swapping. Plates were
assembled using Adobe Indesign CS 5.5. Respective image
libraries of the interactive rSEM have been deposited in
MorphBank. More details on the method of creation of the
interactive models can be found in
AR apodous rings
H vertical midbody diameter (height)
L body length
MNHN Muséum National d’histoire Naturelle, Paris
MSNB Museo Civico di Storia Naturale ‘Enrico Caffi’, Bergamo, Italy
NMNHS National Museum of Natural History, Sofia
PR podous rings
RO vertical rows of ocelli
T telson
ZMUC Natural History Museum of Denmark (Zoological Museum), University of Copenhagen
Ageneral characterisation of Ommatoiulus
was given by
Of the ca. 70 species of Ommatoiulus currently recognized, the following occur in Tunisia:
Ommatoiulus chambiensis Akkari & Enghoff, sp. n.
Ommatoiulus crassinigripes Akkari & Enghoff, sp. n.
Ommatoiulus fuscounilineatus (Lucas, 1846)
Ommatoiulus kefi Akkari & Enghoff, sp. n.
Ommatoiulus khroumiriensis Akkari & Enghoff, sp. n.
Ommatoiulus malleatus Akkari & Voigtländer, 2007
Ommatoiulus punicus (Brölemann, 1894)
Ommatoiulus sempervirilis Akkari & Enghoff, 2011
Ommatoiulus seurati (Brolemann, 1925)
Ommatoiulus xenos Akkari & Enghoff, sp. n.
Ommatoiulus xerophilus Akkari & Enghoff, sp. n.
Ommatoiulus zaghouani Akkari & Enghoff, sp. n.
http://zoobank.org/C6950942-CCF3-43CA-96FF-75F9166168ED
http://species-id.net/wiki/Ommatoiulus_chambiensis
Figs 1–6Holotype: ♂, W Tunisia, Kasserine Governorate, Châambi National Park, surroundings of the park´s guest house, 35°10.139'N, 8°40.486'E, alt. 950–1000m, scarce trees, Pinus halepensis, under stones, 7.3.2008, P. Stoev & N. Akkari leg. (ZMUC). Paratypes: 17 ♂♂, 31 ♀♀, 1 immature, same data as holotype (ZMUC); 2 ♂♂, 2 ♀♀, same data as holotype (NMNHS); 1 ♂, 2 ♀♀, W Tunisia, Kasserine Governorate, Châambi National Park, 35°11. 901'N, 8°39.505'E, alt. 1291m, Quercus ilex, Pinus halepensis, slope, under stones and in leaf litter, 9.3.2008, P. Stoev & N. Akkari leg. (ZMUC).
Most similar to Ommatoiulus xerophilus sp. n., but easily distinguished by the shape of promerite and the presence of a distal notch and a small pointed process on solenomerite.
Named after the type locality. Châambi Mountain is the highest mountain range in Tunisia, reaching 1550 m at peak Châambi.
Males: L: 17–23 mm, H: 1.6–2 mm, 46–49 PR+1-2 AR+T; females: L: 18.5–32.2 mm, H: 2.4–3.6 mm, 44–50 PR+1-2 AR+T. General colour brownish with a black sputter, dorsally darker, with a black mid-dorsal line. Head dark brown to black with yellow spots in the occipital area, uniformly black frontally, with yellow spots at antennal level and labrum, the latter yellow and brighter; antennae dark brown. Prozonites covered with yellowish-brown spots on a blackish background, also laterally, interrupted by big black spots at ozopore level, dorsally black with a narrow transverse row of yellow spots anteriorly; metazonites glossy pale to whitish; legs light brown to yellowish. Telson: anal valves black, with a yellow sputter, preanal ring black, somewhat paler on the caudal projection; subanal scale light brown. Prozonites with fine oblique striae; metazonites with regular striation, laterally narrower; suture complete, strongly curving at ozopore level; ozopores small, rounded, situated on metazonites, at about their diameter from the suture. Anal valves with numerous submarginal and marginal setae, ca. 2 setae on the surface; subanal scale rounded and setose; preanal ring protruding in a caudal projection with ca. 3+3 setae on the tip and a small hyaline process.
Male sexual characters. Mandibular stipites expanded in well-rounded posterior-ventral lobes, first pair of legs hook-shaped, remaining legs with postfemoral and tibial pads.
Gonopods. Promerite (P) bent anteriad (Figs 1, 2), proximally subrectangular, gradually narrowing in its distal third (Figs 1, 2); lateral margin with a moderately deep rounded incision (i). In a posterior view showing a distal process expanded in a subtriangular shape, bearing an apical blunt process (ap) and a lateral broad process pointing basad (lp); mesal ridge (M) apically broadened and delimiting a serrated lateral lamellar process; distal process laterally bearing a strong serrated ridge (se) marking a thickness on the posterior surface; telopodite (T) remnant ovoid located in the middle of the proximal part.
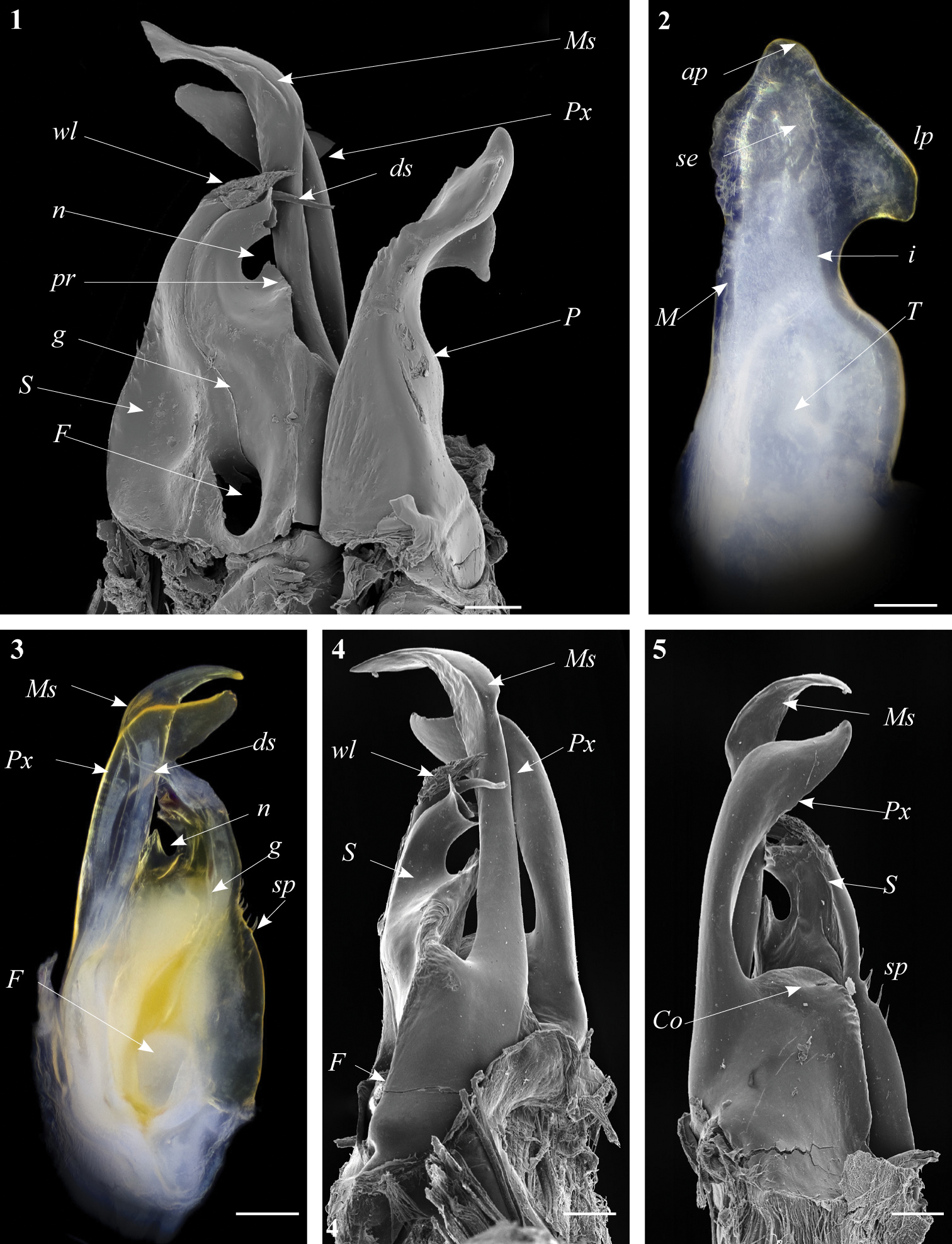
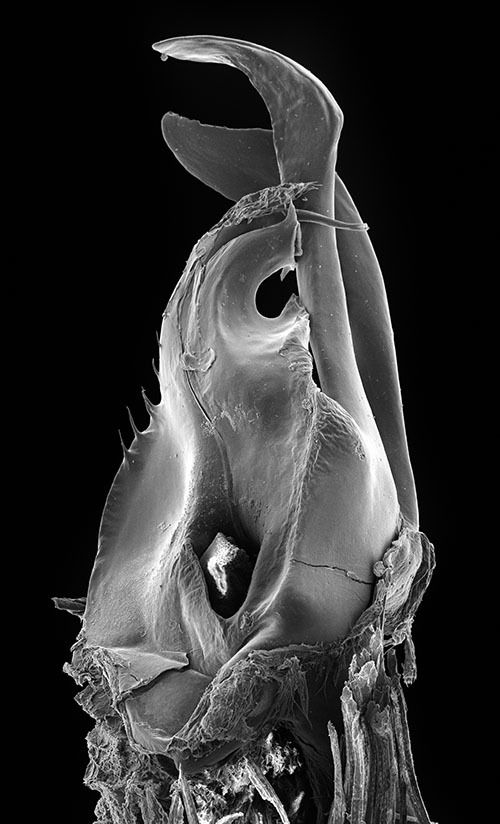
Ommatoiulus chambiensis sp. n. paratype, gonopod structures. 1 Right gonopod, mesal view 2 Left promerite, posterior view 3 Left posterior gonopod, mesal view 4 Right posterior gonopod, anterior view 5 Right posterior gonopod, posterior view. Abbreviations: ap apical process, ds distal process of the solenomerite, F fovea, g seminal groove, i incision on lateral margin of the promerite, lp lateral apical process of promerite, M mesal ridge, Ms mesomerite, P promerite, pr triangular process of the solenomerite, Px paracoxite, S solenomerite, se serrations, sp spikes, T remnant of telopodite, wl wrinkled lamella. Scale bar: 0.1 mm.
Posterior gonopod (Figs 3–6): Mesomerite (Ms) longer than the other processes of the gonopods, distally curved mesad and narrowing into an apical process folded and tapering toward the apex. Solenomerite (S) broad, slightly narrowing at mid-length, proximally with several strong spikes (sp) on the posterior margin; anterior margin with a big, serrated process (pr) pointing distad, separated from the apical part by a rounded notch (n); apical part with an anterior marginally furrowed lamella (Figs 1, 4–6), and a setose wrinkled protruding lamella (wl) covering a protruding slender process (ds) (Figs 1, 3–6) housing the distal part and the opening of the seminal groove (g), the latter running from the fovea (F) at the base of the solenomerite up to process ds. Paracoxite (Px) broad and curved, emerging from a well-rounded coxite (Co), distal third broad then gradually narrowing in a rounded apex pointing mesad (Figs 3–6).
Known only from the type locality, Châambi Mountain, Arid Bioclimatic zone, central Tunisia.
Mixed forest with Quercus ilex and Pinus halepensis, under stones and in leaf litter.
http://zoobank.org/82B068D7-2ABD-4771-9899-2F2E2C51B9CA
http://species-id.net/wiki/Ommatoiulus_crassinigripes
Figs 7–13Holotype: ♂, W Tunisia, Kasserine Governorate, Châambi National Park, Châambi peak and its surroundings, 35°12.285'N, 8°40.653'E, alt. 1500–1540m, Quercus ilex, Pinus halepensis, under stones and leaf litter, 9.3.2008, P. Stoev & N. Akkari leg. (ZMUC). Paratypes: 4 ♂♂, 2 ♀♀, same data as holotype (ZMUC); 1 ♂, 1 ♀, same data as holotype (NMNHS); 1 ♂, 3 ♀♀, 3 juveniles, CW Tunisia, El Kef, 21.4.1983, Bianchi & Moretti leg. (MSNB); 3 ♂♂, 11 ♀♀, 1 intercalary male, 30 juveniles, CW Tunisia, Makthar, 9.3.1986, Bianchi & Moretti leg. (MSNB); 6 ♀♀, 2 subadults, CW Tunisia, 12 km S Thala, 10.3.1986, Bianchi & Moretti leg. (MSNB); 2 ♂♂, 12 ♀♀, 4 juveniles, C Tunisia, Kairouan Governorate, El Manara, on the road Kairouan-Sidi Bouzid, open and dry area, 35°14'N, 09°45'E, alt. 673m, 17.3.2005, N. Akkari leg. (ZMUC).
Gonopods resembling those of Ommatoiulus punicus and Ommatoiulus khroumiriensis sp. n., but differing by the shape of promerite, a much broader and strongly serrated paracoxite, a broader mesomerite bearing subapical serrations on the mesal margin, and the apical processes on solenomerite.
An adjunction of Latin words referring to the body size and leg colour, crassus meaning fat and nigripes, black leg.
Males: L: 24.8–30 mm, H: 2.7–3.6 mm, 45–53 PR+1–2 AR+T. Females: 30–34 mm, H: 3.4–3.9 mm, 45–47 PR+1–2 AR+T. General colour grey, with alternating pale grey and golden brown, darker laterally, with a thin black mid-dorsal line. Head grey, with black sputter frontally, labral zone reddish-brown, brighter at the margin, antennae dark brown. Prozonites pale grey, with big black spots at the level of ozopores and below, a dense black sputter; metazonites whitish anteriorly and golden brown posteriorly, legs black. Telson: anal valves dark grey, bordered with black, preanal ring golden brown, darker on the tip of the caudal projection; subanal scale yellowish. Prozonites with scattered oblique striae; metazonites with regular striation becoming dense laterally; ozopores small and rounded, appearing as brown rounded spots located on metazonites, situated at about their diameter from the suture; the latter complete, only slightly curving at the level of ozopores. Anal valves mostly glabrous at the surface but bearing several submarginal and shorter marginal setae; subanal scale triangular, blunt and setose; preanal ring protruding in a caudal projection, with ca. 2+2 setae and a small hyaline process on the tip.
Male sexual characters. Mandibular stipites expanded in well-rounded posterior-ventral lobes, first pair of legs hook-shaped, remaining legs with postfemoral and tibial pads.
Gonopods. Promerite (Fig. 7) broad, proximally almost rectilinear, bent 90 degrees at notch level; strongly narrowing in its distal third with a deep incision (i) on the lateral margin, latter almost rectilinear; mesal ridge (M) broad distally, protruding in a blunt process (mp) (Fig. 7); posterior surface irregular on the mesal side, bearing a number of strong setae aligned in front of the notch; distal process (ap) laterally broadened and rounded, showing a small mesal serrated process (ap1); remnant of telopodite (T) as a small bump located proximally.
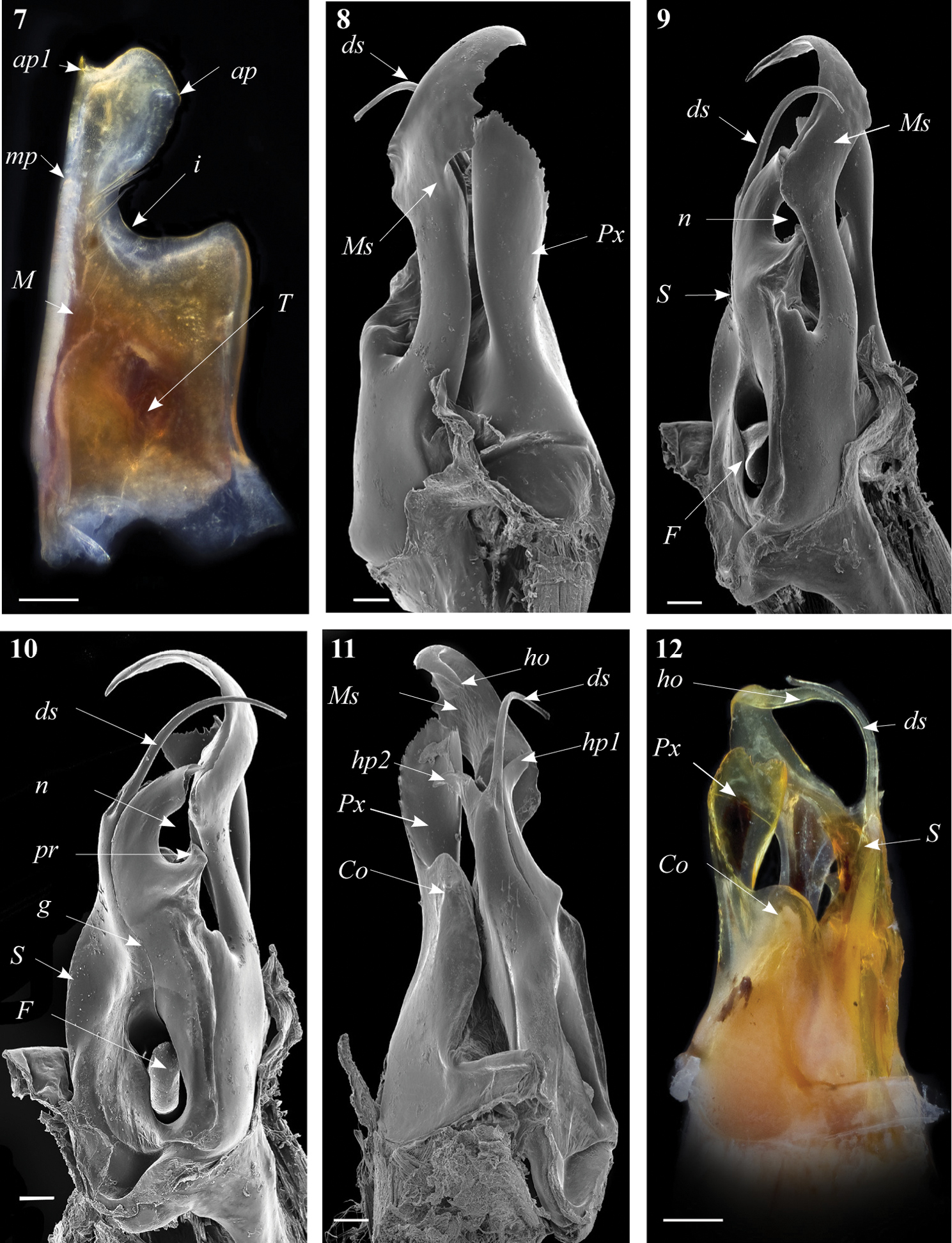
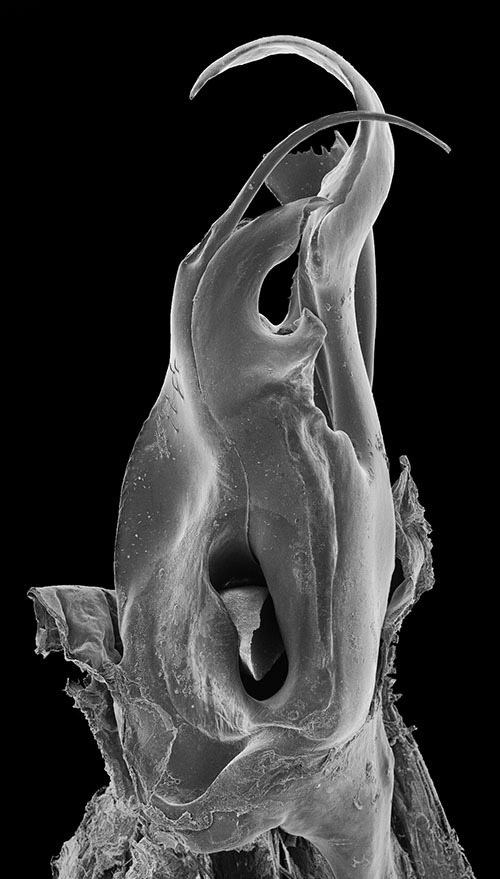
Ommatoiulus crassinigripes sp. n. paratype, gonopod structures. 7 Left promerite, posterior view 8 Right posterior gonopod, lateral view 9 Right posterior gonopod, anterior view 10 Right posterior gonopod, mesal view 11 Right posterior gonopod, posterior view 12 Right posterior gonopod latero-posterior view.Abbreviations: ap distal process of the promerite, ap1 apical mesal process, Co coxite, ds distal process of the solenomerite, F fovea, g seminal groove, ho hook-shaped process, hp1, hp1 distal processes of the solenomerite, i lateral margin incision of the promerite, M mesal ridge, Ms mesomerite, mp distal blunt process of the mesal ridge, n notch of the solenomerite, pr triangular process of the solenomerite, Px paracoxite, S solenomerite, T remnant of telepodite. Scale bar: 0.1 mm
Posterior gonopod (Figs 8–13). Mesomerite (Ms) long, sinuous, distal part asymmetrically enlarged mesolaterally, and showing in lateral view strong serrations at different levels on both margins (Figs 8, 9, 13), gradually narrowed apically in a hook-shaped process (ho) curved and tapering toward the apex (Figs 11, 12, 13). Solenomerite (S) broadest at the base, narrowing at mid-length and bearing a number of strong setae near the posterior margin (Figs 10, 12, 13); in mesal view showing at mid-length a triangular process (pr) pointing distad (Figs 10, 13), latter separated from the apical part by a deep rounded notch (n); apically bearing a long curved process (ds) pointing mesad, housing the distal part and the opening of the seminal groove (g) and emerging between a posterior and an anterior folded hyaline processes (hp1, hp2) (Figs 11–13). Seminal groove running from the fovea (F) at the base of the solenomerite up to process ds (Figs 10, 13). Paracoxite (Px) lamellar, broad and folded, emerging from a rounded coxite (Co), distally broadened, apical margin almost horizontal, and together with the posterior margin showing many strong, short serrations (Figs 8, 11–13).
Semi-arid and Arid bioclimatic zones in west central Tunisia, recorded from the governorates Kasserine, El Kef, Thala and Kairouan.
Dry and open habitats, to 1500 m in Châambi Mountain.
http://zoobank.org/93E41C97-AA12-45D5-BD86-F2B0ACF02465
http://species-id.net/wiki/Ommatoiulus_kefi
Figs 14–19Holotype: ♂, W Tunisia, El Kef Governorate, 13 km S El Kef, 22.iv.1981, Bianchi & Moretti leg. (MSNB).
Differing from all congeners by having a tri-lobate distal part of promerite and a bifurcated apical part of mesomerite, the latter divided in two short oppositely directed processes.
Named after El Kef city, the type locality of the species.
Male: L: 26 mm, H: 2.7 mm, 53 PR+1 AR+T. General colour alternating whitish and blackish with a thin black mid-dorsal line. Head brown, lighter on the frontal part, with yellowish spots at antennal level, labral zone yellowish, becoming brighter at the margin, antennae brownish. Prozonites pale grey, dorsally scarcely sputtered with black; metazonites anteriorly dark, with a blackish background and a line of light brown spots below ozopores; legs whitish. Telson: anal valves and preanal ring blackish, paler towards caudal projection, subanal scale yellowish. Prozonites with scattered oblique striae; metazonites with regular striation becoming dense laterally; suture complete and rectilinear; ozopores small, rounded and located in metazonites, well apart from the suture. Anal valves with numerous submarginal and marginal setae and ca. 1-2 setae on the surface; subanal scale triangular, blunt and setose; preanal ring protruding in a caudal projection, with ca. 3+3 setae and a small hyaline process on the tip.
Male sexual characters. Mandibular stipites expanded in rounded posterior-ventral lobes, first pair of legs hook-shaped, remaining legs with postfemoral and tibial pads.
Gonopods. Promerite (Fig. 14) in posterior view subrectangular, mesal ridge (M) fairly broad, distally narrowing and protruding into a pointed apical lobe (al); apical margin protruding in a curved median lobe (me) pointing laterad and a shorter broad lateral lobe (lb); the three apical lobes separated by two rounded incisions; lateral margin almost rectilinear. Remnant of telopodite (T) ovoid, located at mid-length of promerite.
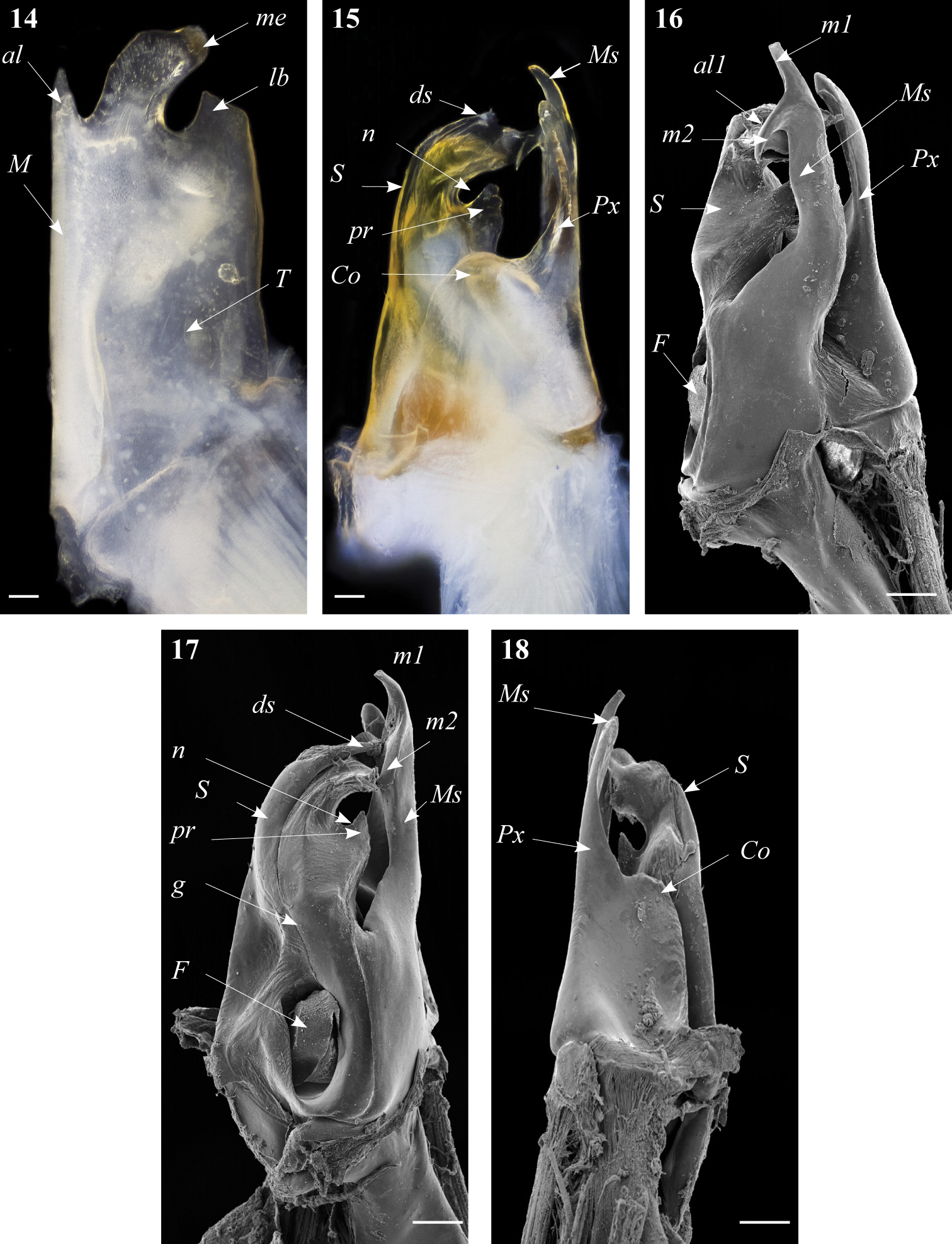
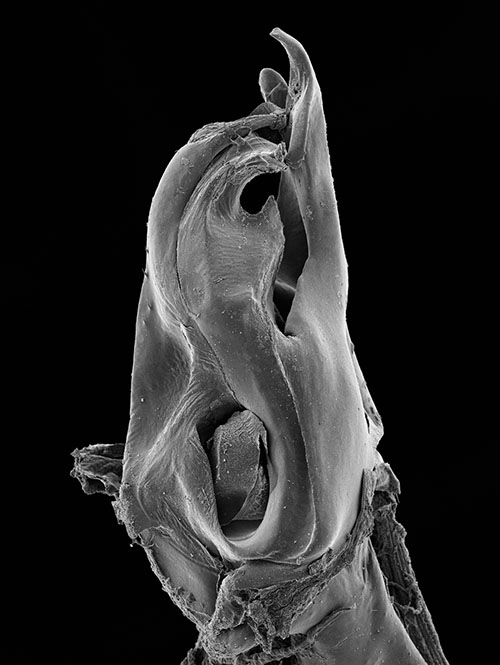
Ommatoiulus kefi sp. n. holotype, gonopod structures. 14 Left promerite, posterior view 15 Left posterior gonopod, posterior view 16 Right posterior gonopod, anterior view 17 Right posterior gonopod, mesal view 18 Right posterior gonopod, mesal view. Abbreviations: al apical lobe of the promerite, al1 apical folded lamella of the solenomerite, Co coxite, ds distal process of the solenomerite, F fovea, g seminal groove, lb lateral lobe, m1, m2 apical processes of the mesomerite, M mesal ridge, me median lobe, Ms mesomerite, n notch of the solenomerite, pr triangular process of the solenomerite, Px paracoxite, S solenomerite, T telopodite. Scale bar: 0.1 mm
Posterior gonopod (Figs 15–19): Mesomerite (Ms) broadest at the base, distally protruding in a uniformly broad process, apically splitting into two short and curved processes, pointing in opposite directions (m1, m2) (Figs 16, 17, 19); solenomerite (S) broad at the base, slightly narrowing at mid-length and showing a triangular process (pr) separated from the rest of the processes by a rounded (n) (Figs 17, 19), apical part of the solenomerite complex with a broad lamella (al1) extended latero-mesad, downturned and marginally furrowed (Fig. 16). Seminal groove (g) running from the fovea (F) at the base of solenomerite up to a slender and short conical process (ds) emerging on top of the median part of the apical lamella and pointing anteriad (Figs 15, 17). Paracoxite (Px) stout, with smooth margins, emerging from a broad rounded coxite (Co) (Figs 18, 19).
Semi-arid bioclimatic zone in western Tunisia; hitherto known only from the type locality near El Kef city.
We have examined three females (MSNB) collected from the same locality but could not assign them with certainty to Ommatoiulus kefi as they show a different colour pattern.
http://zoobank.org/05BAED95-725C-4395-92C3-6BF22A01CE18
http://species-id.net/wiki/Ommatoiulus_khroumiriensis
Figs 20–26Holotype: ♂, NW Tunisia, Jendouba Governorate, Aïn Draham, Col des Ruines, 1.11.2009, N. Akkari leg. (ZMUC). Paratypes: 2 ♂♂, 5 ♀♀, NW Tunisia, Jendouba Governorate, Aïn Draham, Col des Ruines, 1.11.2009, N. Akkari leg. (ZMUC); 2 ♂♂, 6 ♀♀, NW Tunisia, Jendouba Governorate, 7 km south Aïn Draham, les chênes, 22.3.1986, ZMUC expedition; 1 ♂, 1 ♀, 4 immatures, 5-18.3.1988, NW Tunisia, Jendouba Governorate, Aïn Draham area, ZMUC expedition; 1 ♂, 1 ♀, 1 intercalary male, NW Tunisia, Jendouba Governorate, Aïn Draham, 19.11.2003, forest with Quercus suber and Quercus faginea, under stones, N. Akkari leg. (NMNHS); 2 ♂♂, 5 ♀♀, 2 juveniles, NW Tunisia, Jendouba Governorate, Hammam Bourguiba, 36°45'N, 08°35'E, alt. 158m, mixed forest with Pinus pinaster and Quercus suber, under stones, 31.10.2009, N. Akkari leg. (ZMUC); 3 ♂♂, NW Tunisia, Jendouba Governorate, Aïn Draham, 36°47'N, 8°41'E, alt. 511m, 3.10.2005, N. Akkari leg. (ZMUC); 1 ♂, 2 ♀♀, NW Tunisia, Jendouba Governorate, Aïn Draham, 36°47'N, 8°41'E, 760m, Quercus suber-Erica forest, 11.3.2009, N. Akkari & H. Enghoff leg. (ZMUC); 3 ♂♂ Jendouba Governorate, route Aïn Draham- Fernana, 36°43'N, 8°40'E, Quercus suber-Erica forest, 9.3.2009, N. Akkari & H. Enghoff leg. (ZMUC); 1 ♂, 2 ♀♀, Jendouba Governorate, route Aïn Draham- Béni M’tir, 36°43'N, 8°42'E, Quercus suber-Erica forest, 10.3.2009, N. Akkari & H. Enghoff leg. (ZMUC).
Similar to Ommatoiulus punicus and Ommatoiulus crassinigripes sp. n. but readily distinguished by the shape of promerite having a deeper notch extended basad, much slenderer processes of posterior gonopods, and a more sinuous mesomerite devoid of conspicuous serrations.
The species name refers to the natural region of Khroumirie, NW Tunisia, to which the species seems confined.
Males: L: 26–27 mm, H: 2–2.8 mm, 43–48 PR+1–2 AR+T, females: L: 30–37 mm, H: 4–4.3, 44–48 PR+1 AR+T. General colour dark grey, alternating with brown-yellow laterally, and with a thin black mid-dorsal line. Head dark reddish-brown; occipital area blackish, with brown-reddish- spots; frontal part uniformly black, labral zone brown-reddish- to yellowish at margin, antennae brownish. Prozonites uniformly grey, with a pale narrow stripe anteriorly; metazonites darker, brown-greyish, densely sputtered with black, colour gradually vanishing on the sides, below ozopore level yellow-brownish; legs light brown. Telson: anal valves black, preanal ring blackish, caudal projection brown-reddish, subanal scale light brown to yellowish.
Prozonites with scattered oblique striae; metazonites densely striated; suture complete, curving at ozopore level; ozopores small, rounded and located on metazonites, situated at about their diameter from the suture. Anal valves with 4-5 setae on the surface, a submarginal row of 12-13 setae and numerous short marginal ones; subanal scale triangular and setose; preanal ring protruding in a caudal projection with ca. 3+3 setae and a small hyaline process on the tip.
Male sexual characters. Mandibular stipites expanded in rounded posterior-ventral lobes, first pair of legs hook-shaped, remaining legs with postfemoral and tibial pads.
Gonopods. Promerite (Fig. 20) strongly narrowed distally with a deep lateral incision (i) extending meso-basad, distal process (ap) broad, subtriangular, with two pointed edges, the tip of apical process with a small pointed lobe (ap2); mesal ridge (M) distally protruding in a blunt small cylindrical process (mp), posterior surface of promerite with a row of strong setae emerging at the level of the notch, in close proximity to the mesal ridge.
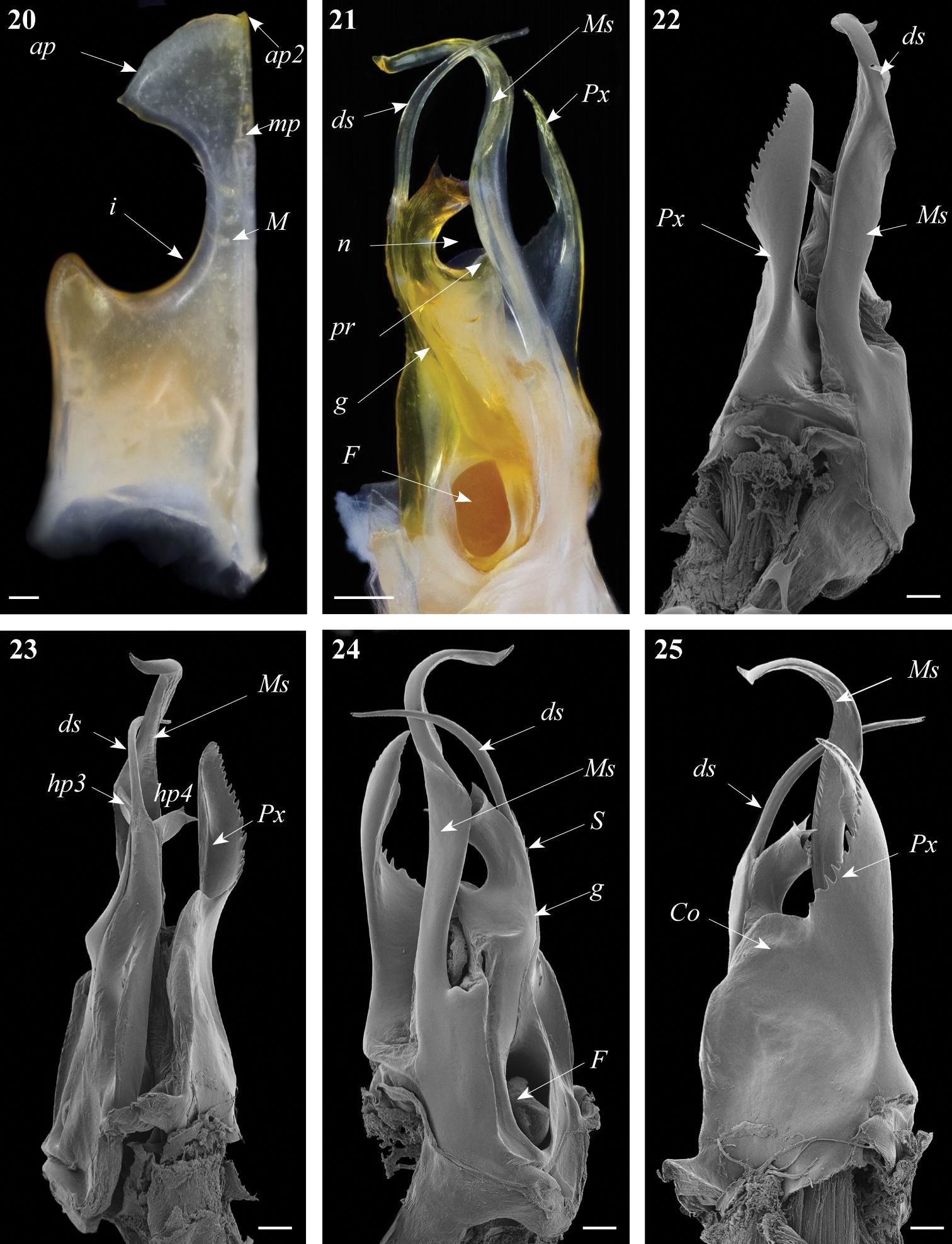
Ommatoiulus khroumiriensis sp. n. paratypes, gonopod structures. 20 Right promerite, posterior view 21 Right posterior gonopod, mesal view 22 Right posterior gonopod, lateral view 23 Right posterior gonopod, meso-posterior view 24 Right posterior gonopod, anterior-mesal view 25 Right posterior gonopod, anterior view. Abbreviations: ap distal process of the promerite, ap2 lobed process, Co coxite, ds distal process of the solenomerite, F fovea, g seminal groove, hp3, hp4 distal processes of the solenomerite, i lateral incision of the promerite, mp blunt mesal process, M mesal ridge, Ms mesomerite, n notch of the solenomerite, pr triangular subapical process of the solenomerite, Px paracoxite, S solenomerite. Scale bar: 0.1 mm.
Posterior gonopod (Figs 21–26). Mesomerite (Ms) large, longer than the other processes, uniformly broad, sinuous; distal third constricted to less than half breadth and apically protruding into a slender curved process, latter tapering and pointing mesad (Figs 21, 23, 24, 26). Solenomerite (S) broadest at the base, narrowing at mid-length, and bearing a number of strong setae near the posterior margin, distally with a broad, blunt triangular process (pr) separated from the apical part by a rounded notch (n), and with a long curved process (ds) protruding between two apical hyaline processes (hp3, hp4) and housing the apical part of seminal groove (g), the latter (g) running from the fovea (F) located at the base of the solenomerite (S) up to process ds.Paracoxite (Px) emerging from a depressed coxite (Co); Px curved, half as broad as in Ommatoiulus crassinigripes, gradually narrowing distad; lateral and apical margins, with a saw-like strongly jagged margin (Figs 22–26).

Humid bioclimatic zone in northwestern Tunisia; known from Aïn Draham, Fernana and Hammam Bourguiba in Khroumirie, Jendouba Governorate.
Mixed forests dominated by Quercus faginea and Quercus suber, or Pinus pinaster and Quercus suber.
http://zoobank.org/09D98AF5-9AD3-41E0-8B54-624E31B72480
http://species-id.net/wiki/Ommatoiulus_xenos
Figs 27–30Holotype: ♂, Tunisia (“Tunis”), 1861, J.P. Coindé leg. (MNHN). Paratypes: 5 ♀♀, Tunisia (“Tunis”), 1861, J.P. Coindé leg. (MNHN).
Resembling Ommatoiulus chambiensis and Ommatoiulus xerophilus spp. n. in size and general shape of gonopods, but distinguished by the shape of promerite, a much more slender mesomerite and shorter and stouter paracoxite.
The species name is a Greek noun meaning ‘stranger’, emphasising the fact that this species, found surprisingly in the collection of the MNHN shortly before completion of the manuscript, had remained unknown and out of the sight of a number of myriapodologists for more than 150 years.
Male: L: 20.5 mm, H: 1.85 mm, 47 PR+2 AR+T; females: L: 18.5–21 mm, H: 2.26–2.46 mm, 42–48 PR+2–3 AR+T. General colour faded, generally grey-greenish (very likely an artefact from the decomposition of the inserted label), somewhat lighter laterally. Head pale in the occipital and labral areas; antennae and legs darker. Prozonites with darker triangular spots laterally, latter situated along the ozopores line and forming two longitudinal dark bands, dorsally separated by a pale one; dorsum crossed by dark triangular spots and showing a thin black mid-dorsal line; metazonites mostly pale and glossy. Telson: anal valves and preanal ring dark, subanal scale pale.
Prozonites with fine striae; metazonites with regular striae, denser on the sides, suture complete, curving at the level of ozopores; ozopores small, rounded, situated on metazonites situated at about their diameter from the suture. Anal valves setose; preanal ring with 3-4 setae on each lateral side, protruding in a short caudal projection with 1-4/5 setae and a small hyaline process on the tip. Subanal scale blunt to rounded and setose.
Male sexual characters. Mandibular stipites expanded in rounded posterior-ventral lobes, first pair of legs hook-shaped, remaining legs with postfemoral and tibial pads.
Gonopods. Promerite (Fig. 27) gradually narrowed distally, lateral margin with a shallow incision (i); apical process of promerite with a rounded margin pointing laterad; mesal ridge (M) narrow, distally protruding in a pointed apex (mp) separated from the apical process by a small apical incision; remnant of telopodite not very conspicuous.
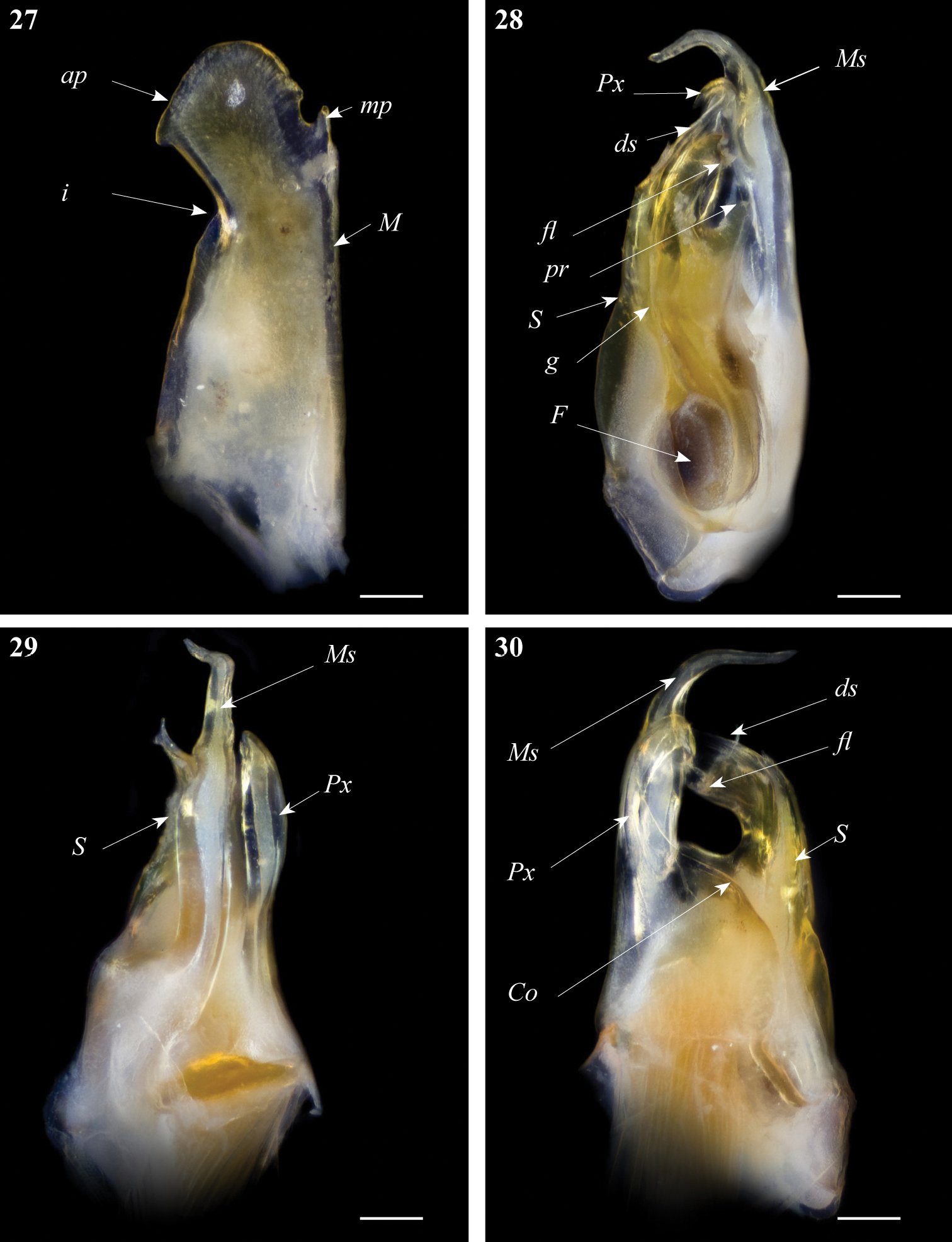
Ommatoiulus xenos sp. n. holotype, gonopod structures. 27 Right promerite, posterior view 28 Right posterior gonopod, mesal view 29 Right posterior gonopod, antero-lateral view 30 Right posterior gonopod, posterior view. Abbreviations: Co coxite, ds distal process of the solenomerite, F fovea, fl folded lamella, g seminal groove, i lateral incision of the promerite, M mesal ridge, mp distal process, Ms mesomerite, pr blunt process of the solenomerite, Px paracoxite, S solenomerite. Scale bar: 0.1 mm.
Posterior gonopod (Figs 28–30): Mesomerite (Ms) uniformly broad proximally, strongly narrowed in its distal third and bent posteriad (Figs 28, 29); solenomerite (S) broad, with scattered setae on posterior margin, narrowing at mid-length, and bearing a large blunt process (pr); solenomerite apically with a broad folded lamella (Fl) and a small wrinkled lamella laying on the top of a slender and slightly protruding process (ds) housing the distal part of the seminal groove (g); seminal groove running from the fovea (F) and opening at the apex of process ds (Fig. 28). Paracoxite (Px) stout, distally curved mesad and narrowed into a slender apex pointing basad emerging from a broad and rounded coxite (Co) (Fig. 30).
Exact locality unknown. The label mentions ‘Tunis’ which presumably refers to Tunisia in general.
Unknown.
The collector of this species, J.P. Coindé, who was a ‘zoologist-traveler’, made a collecting trip to Tunisia in 1861 during which he visited several localities throughout the country. Although we are certain that Ommatoiulus xenos sp. n., found by chance in an obscure jar among several unidentified myriapods from North Africa, labelled ‘Brolemann unidentified’, was collected in Tunisia, we couldn’t determine with certainty the locality where this species was collected 152 years ago.
http://zoobank.org/6C3A27BC-A781-4EF3-B2BD-2275C59126FF
http://species-id.net/wiki/Ommatoiulus_xerophilus
Figs 31–35Holotype: ♂, W Tunisia, Kasserine Governorate, Châambi National Park, surroundings of the park´s guest house, 35°10.139'N, 8°40.486'E, alt. 950–1000m, scarce trees, Pinus halepensis, Thuja, under stones, logs and in leaf litter, 8.3.2008, P. Stoev & N. Akkari leg. (ZMUC). Paratypes: 1 ♂, 2 ♀♀, W Tunisia, Kasserine Governorate, Châambi National Park, surroundings of the park's guest house, 35°10.139'N, 8°40.486'E, alt. 950m, scarce trees, Pinus halepensis, under stones, 7.3. 2008, N. Akkari & P. Stoev leg. (ZMUC); 2 ♂♂, 12 ♀♀, 8 subadult ♀♀, 8 juveniles, W Tunisia, El Kasserine Governorate, Châambi National Park, surroundings of the park’s guest house, 35°10. 139'N, 8°40.486'E, alt. 950–100m, scarce trees, Pinus halepensis, Thuja, under stones, logs and in leaf litter, 8.3.2008, P. Stoev & N. Akkari leg. (ZMUC); 1 ♂, 2 ♀♀, same data (NMNHS).
Resembling Ommatoiulus chambiensis in the structure of mesomerite, paracoxite, but well distinguished from the latter by the characteristic globular apex of promerite and the shape of solenomerite devoid of a rounded notch.
The species name is a Greek adjective referring to the affinity of the species for dry habitats.
Males: L: 15.7–15.9 mm, H: 1.54–1.65 mm, 44–46 PR+1–2 AR+T; females: L: 10.2–23.1 mm, H: 1.44–2.56 mm, 39–46 PR+1–4 AR+T. General colour black to brownish, light brown on the sides; dorsum pale yellow, crossed by thick black mid-dorsal spots. Head dark to blackish with yellow spots in the occipital area, frontal part uniformly black showing yellow spots at antennal level, labral zone yellowish, brighter at margin; antennae brownish. Prozonites blackish with light brown-yellowish spots becoming dominant laterally, just below the ozopore line; dorsally pale, with big, irregularly shaped black spots; metazonites predominantly grey-whitish and glossy, legs yellowish. Telson: anal valves dark brown-blackish, with dense yellow sputter, preanal ring blackish sputtered with yellow, dorsal side and caudal projection mostly pale, subanal scale yellowish.
Prozonites with fine striae; metazonites with regular striae, becoming denser laterally, suture complete, curving at the level of ozopores; latter small, rounded, situated on metazonites situated at about their diameter from the suture. Anal valves setose, with 6-7 setae on the surface and numerous submarginal and marginal setae; subanal scale rounded and setose; preanal ring with 1+1 setae on the sides, protruding in a caudal projection with 3+3 setae and a small hyaline process on the tip.
Male sexual characters. Mandibular stipites expanded in rounded posterior-ventral lobes, first pair of legs hook-shaped, remaining legs with postfemoral and tibial pads.
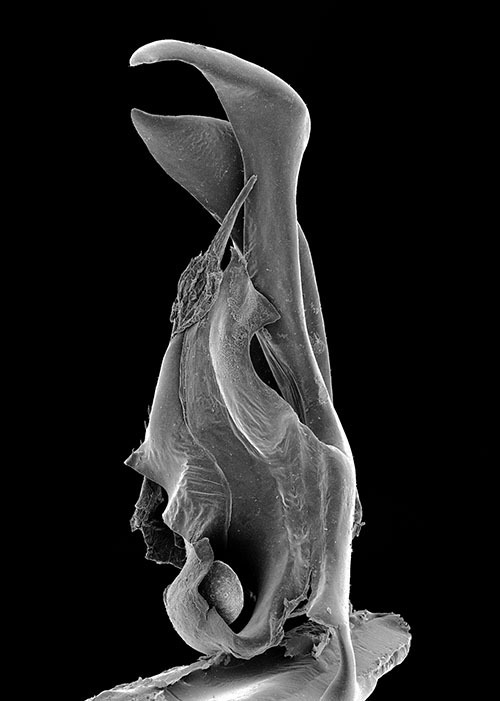
Gonopods. Promerite (P) (Figs 31, 32) bent anteriad, not very broad, with parallel margins, mesal ridge (M) broad, distally narrowing and truncate, bearing several serrations (se) and separated from the apical lobe (ap); by an incision; lateral margin mostly rectilinear, only slightly narrowing subapically at the level of the mesal incision; apical lobe globular, with rounded margin, curved laterad; posterior surface with a number of strong setae aligned distally; telopodite remnant inconspicuous.
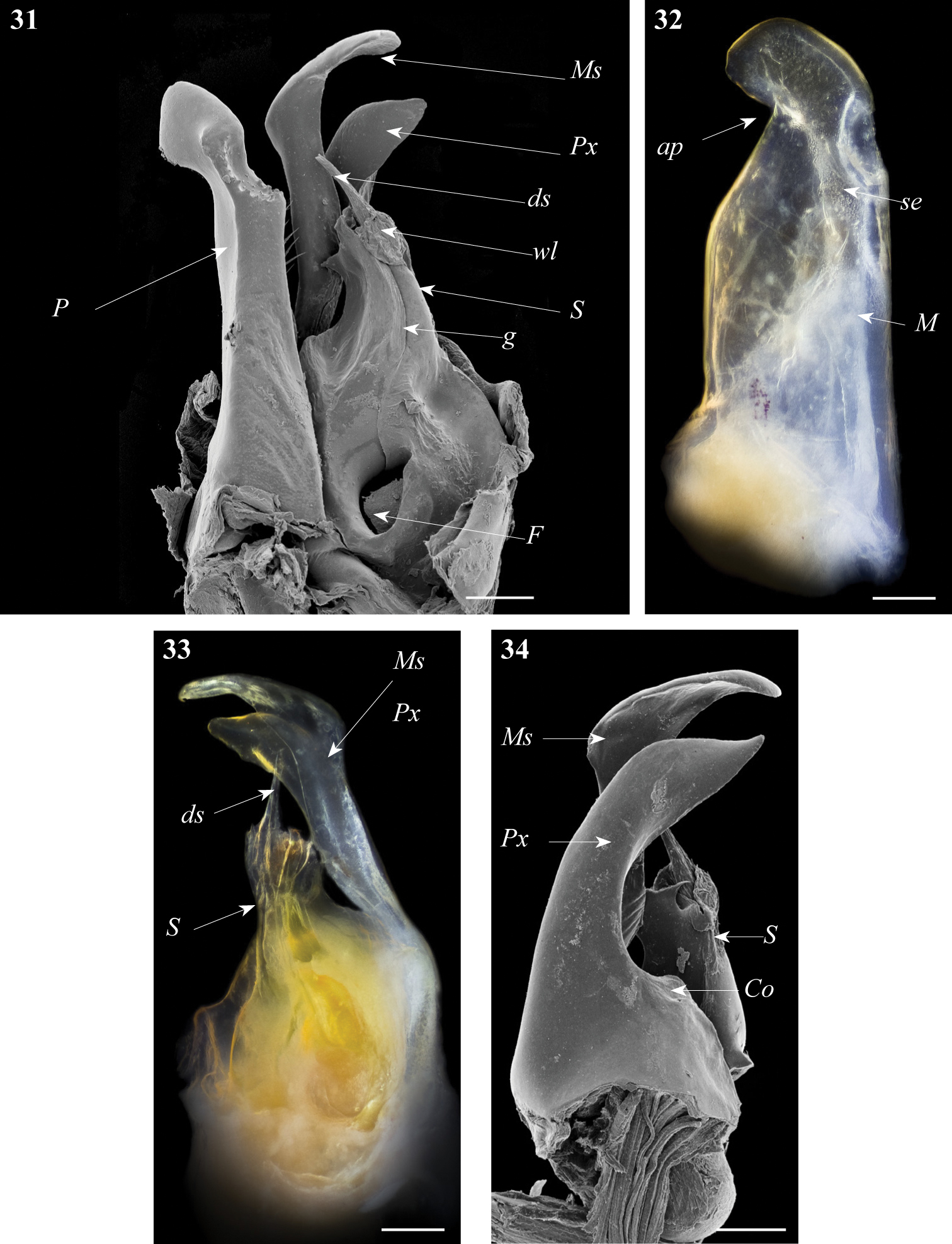
Ommatoiulus xerophilus sp. n. paratype, gonopod structures. 31 Left gonopod, mesal view 32 Rightpromerite, posterior view 33 Rightposterior gonopod, mesal view 34 Rightposterior gonopod, latero-posterior view. Abbreviations: ap distal process of the promerite, Co coxite, ds distal process of the solenomerite, F fovea, g seminal groove, M mesal ridge, Ms mesomerite, P promerite, Px paracoxite, S solenomerite, se serrations, wl wrinkled lamella. Scale bar: 0.1 mm.
Posterior gonopod (Figs 31, 33–35): Mesomerite (Ms) similar to Ommatoiulus chambiensis but broader, strongly truncated and distally bent mesad (Figs 33–35); solenomerite (S) broad, with scattered setae on posterior margin, strongly narrowing at mid-length, apical part with a wrinkled blunt lamella (wl) covering a slender and protruding process (ds) housing the distal part of the seminal groove (g) (Figs 31, 33, 35); the latter running from the fovea (F) (Figs 31, 33–35) up to process ds. Paracoxite (Px) broad, curved mesad, emerging from a depressed coxite (Co), distal third slightly enlarged, apically narrowing into a pointed apex, directed mesad (Figs 33–35).
Arid bioclimatic zone, central Tunisia; hitherto known only from the type locality, Châambi Mountain.
Open areas with scattered Pinus halepensis trees.
http://zoobank.org/F23E46BA-1E8C-4B6D-BF46-73D9A37E8E42
http://species-id.net/wiki/Ommatoiulus_zaghouani
Figs 36–39Holotype: ♂, NE Tunisia, Zaghouan Governorate, Jebel Zaghouan, 36°23'N, 10°06E, alt. 365m, Pinus forest, 13.3.2009, N. Akkari & H. Enghoff leg. (ZMUC).
Gonopods resembling those of Ommatoiulus seurati but distinguished by a broader distal part of the promerite, a subapical lateral projection on the mesomerite and a much shorter solenomerite.
Named after Jebel Zaghouan, the type locality.
Male: L: 28.5 mm, H: 2.56 mm, 49 PR+1 AR+T. General colour alternate dark and light golden brown; dorsum with a thin black axial line. Head dark to blackish, with brownish spots on the frontal part and on the mandibular stipites, labral zone and mouth parts pale, marginally bright yellow; antennae brownish. Prozonites dark to blackish, covered with yellowish spots; metazonites pale brown to whitish laterally and golden brown dorsally, legs yellowish. Telson: anal valves black, preanal ring blackish, caudal projection yellowish, subanal scale yellowish.
Prozonites with fine irregular striae; metazonites with regular striae, becoming dense laterally, suture complete, curving at the level of ozopores; ozopores small, rounded, situated in metazonites, situated at about their diameter from the suture. Anal valves setose, with 4-6 setae on the surface, ca 14 submarginal and numerous marginal setae; subanal scale rounded and setose; preanal ring with 2+2 setae on the sides, protruding in a caudal projection with (6-7)+(6-7) setae on the tip and bearing a small hyaline process.
Male sexual characters. Mandibular stipites expanded in rounded posterior-ventral lobes, first pair of legs hook-shaped, remaining legs with postfemoral and tibial pads.
Gonopods. Promerite (Fig. 36) proximally subrectangular, strongly narrowed distally by a deep incision (i) on the lateral margin; mesal ridge (M) broad, distally protruding in a serrated edge (se); apical part with a mesal triangular blunt process (rp) protruding mesodistad, and a lateral protruding process with two small apical bumps (bp1, bp2); posterior surface of promerite showing few scattered setae near the mesal margin; remnant of telopodite (T) large and ovoid, located at mid-length of the process.
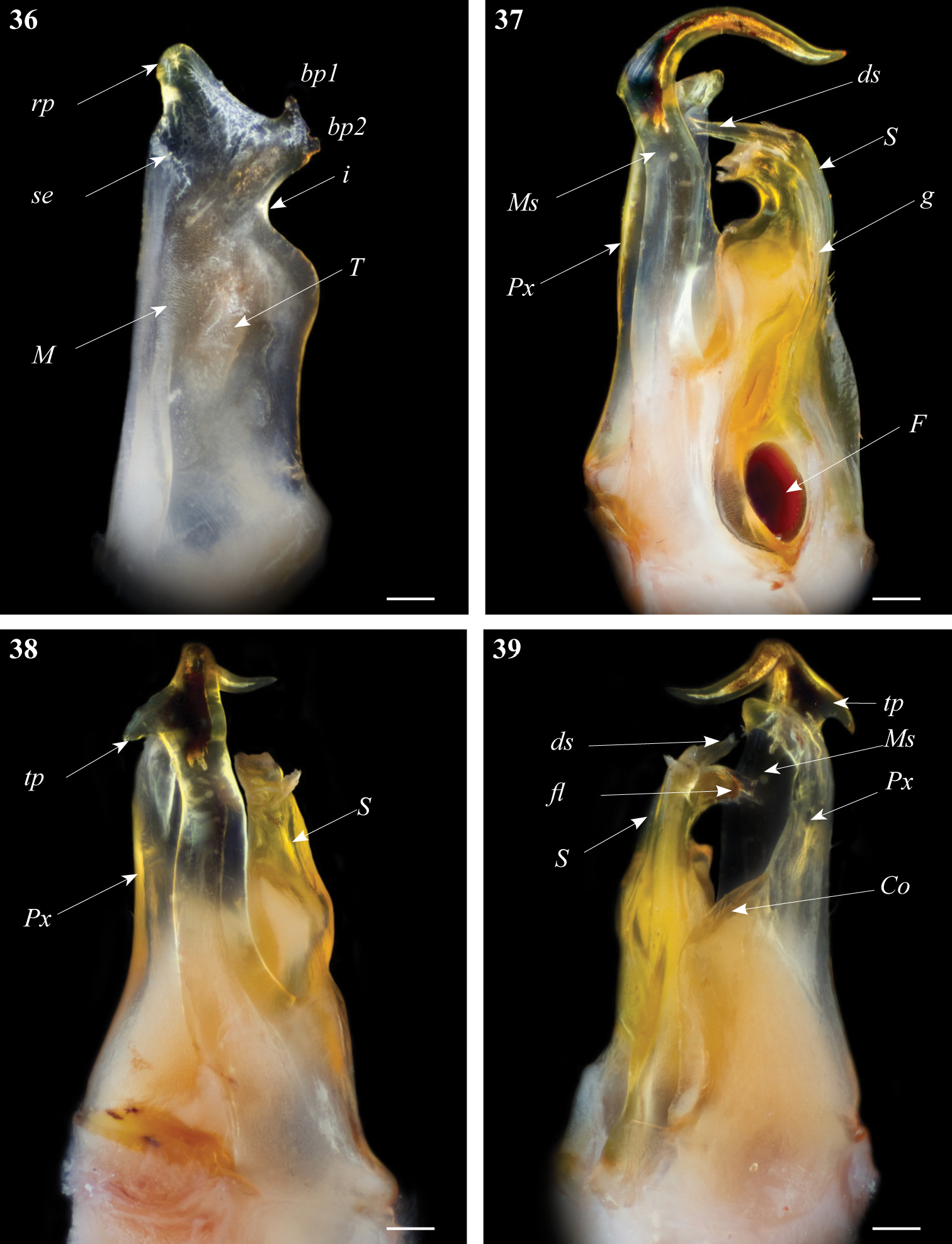
Ommatoiulus zaghouani sp. n. holotype, gonopod structures. 36 Left promerite, posterior view 37 Left posterior gonopod, mesal view 38 Left posterior gonopods, anterior view 39 Left posterior gonopods, postero-lateral view. Abbreviations: bp1, bp2: small bumps on the apical lateral process, Co coxite, ds distal process of the solenomerite, F fovea, fl folded lamella, g seminal groove, M mesal ridge, Ms mesomerite, i lateral incision on the promerite, Px paracoxite, rp apical mesal process of the promerite, S solenomerite, se serrated process, tp: triangular distal process. Scale bar: 0.1 mm.
Posterior gonopod (Figs 37–39): Mesomerite (Ms) large, and uniformly broad (Figs 37, 38) with a distal triangular pointed extension on the lateral margin (tp), distal third strongly curved mesoposteriad and narrowed in a long and slender apical process (Figs 37, 38, 39); solenomerite (S) broad, with scattered setae on posterior margin; anteriorly simply rounded devoid of processes; apically with a hyaline folded lamella (fl) and a slightly protruding process (ds) housing the distal part of the seminal groove (g); the latter running from the fovea (F) (Fig. 37) up to process ds. Paracoxite (Px) stout and curved apically slightly narrowing into a rounded apex directed mesad, coxite broad (Co) (Fig. 39).
Semi-arid bioclimatic zone in northeastern Tunisia; known only from Zaghouan Mountain.
Forest dominated by Pinus halepensis.
- We refute the presence of Ommatoiulus
diplurus appendiculatus
(Brolemann, 1925) in Algeria. This taxon is based on
females and juveniles only, and
Akkari and Enghoff (2012) already regarded the presence of Ommatoiulus diplurus (Attems, 1903) in North Africa uncertain. - The record of Ommatoiulus
aumalensis
(Brolemann, 1925) from Tunisia by
Akkari et al. (2009) was due to a misidentification and actually refers to Ommatoiulus fuscounilineatus (Lucas, 1846). - Ommatoiulus
sempervirilis
Akkari & Enghoff, 2011 was described from the
Tunisian islands Galita and Zembretta (
Akkari and Enghoff 2011 ). - Ommatoiulua chambiensis, Ommatoiulus crassinigripes, Ommatoiulus kefi, Ommatoiulus khroumiriensis, Ommatoiulus xerophilus, Ommatoiulus xenos and Ommatoiulus zaghouani spp. n., are described from northern and central Tunisia in the present paper bringing the overall number of Ommatoiulus species in Tunisia to 12, and to 28 in North Africa.
- We refute the presence of Ommatoiulus
lapidarius
(Lucas, 1846) in Libya. This record was based on the
synonymy of Julus
rimosus Karsch
1881 (see
Manfredi 1939 ,Schubart 1952 ) with Ommatoiulus lapidarius, but re-examination of the holotype of Julus rimosus (Museum für Naturkunde, Berlin) has shown that this is a valid species (Akkari 2013 ).
Three of the new Tunisian Ommatoiulus
species were found in Châambi Mountain, which is the
highest mountain range in Tunisia. The mountain is located
only a few kilometres from the Algerian border and in spite
of its arid character is known to harbour a number of
endemic species (cf.
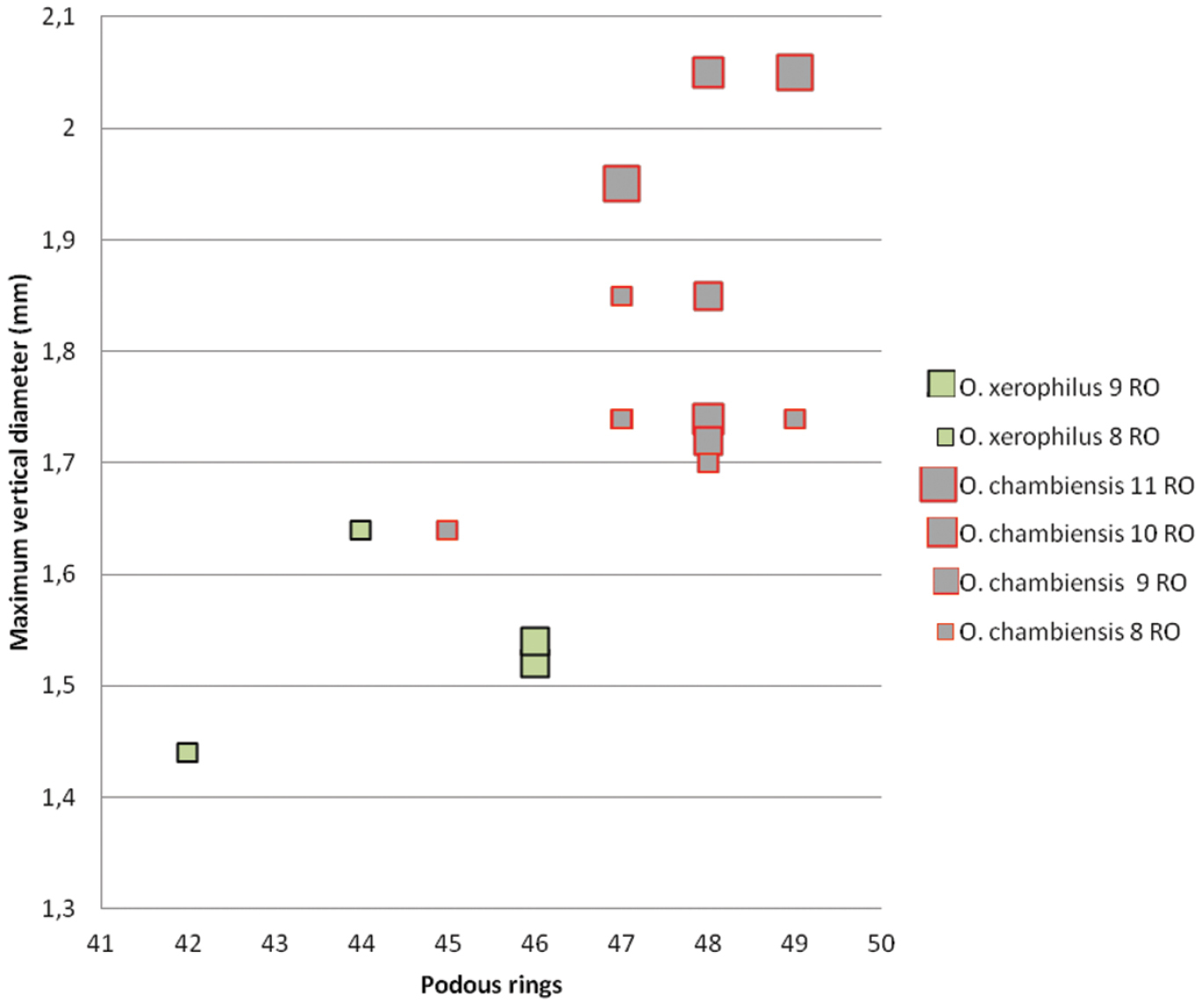
Ommatoiulus chambiensis and Ommatoiulus xerophilus spp. n.Scatter diagram illustrating size of adult males expressed as maximum vertical body diameter (mm) vs number of podous rings in different developmental stadia (8–11RO).
Ommatoiulus
crassinigripes is
the only schizophylline millipede collected at more than
1500 m elevation in Tunisia. However, it occurs also at
lower elevation in central Tunisia (El Kef, Thala, Makthar,
Kairouan) and is very unlikely to be an alticolous species
like Ommatoiulus
gravieri
(Brolemann, 1924), which is known from 3000–3200 m altitude
in Jebel Tachdirt of the High Atlas in Morrocco (
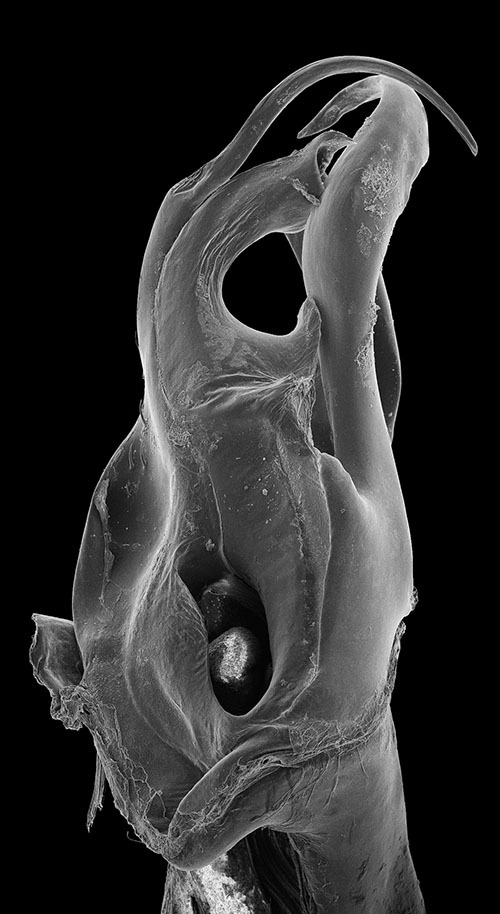
Ommatoiulus punicus, specimen from Jebel Rsas, NE Tunisia, right posterior gonopod. Interactive SEM image. [Morphbank # 831228–831245]
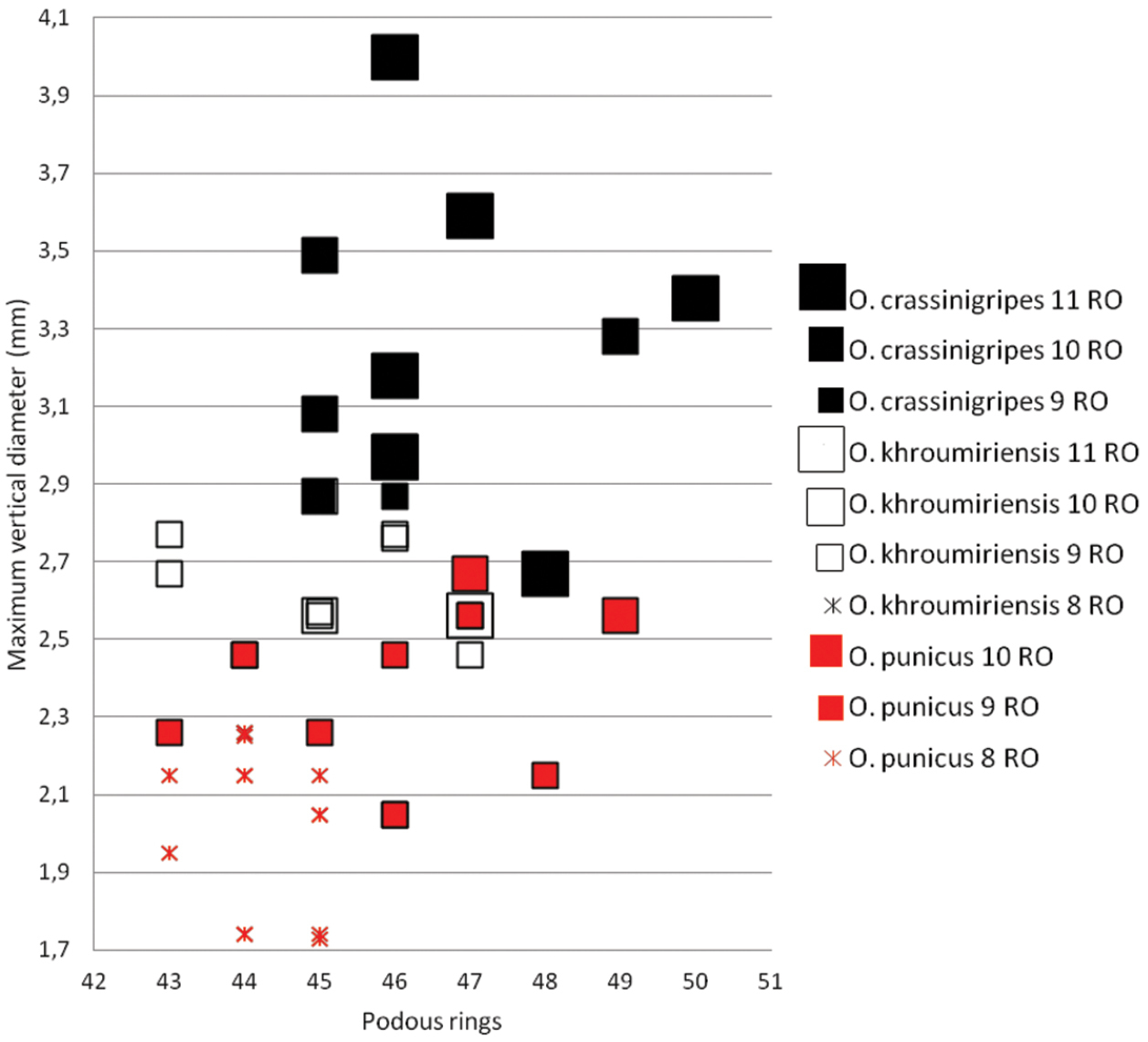
Ommatoiulus crassinigripes sp. n., Ommatoiulus khroumiriensis sp. n. and Ommatoiulus punicus. Scatter diagram illustrating size of adult males expressed as maximum vertical body diameter (mm) vs number of podous rings in different developmental stadia (8–11RO).
Comparison of main gonopod and peripheral structures of Ommatoiulus
punicus, Ommatoiulus
crassinigripes and
Ommatoiulus
khroumiriensis.
| Ommatoiulus punicus (Figs 41, 51–53) |
Ommatoiulus crassinigripes (Figs 7–13) |
Ommatoiulus khroumiriensis (Figs 20–26) |
|
|---|---|---|---|
| Gonopod structures | Similarities | ||
| Promerite | abruptly narrowing in the distal half-third | ||
| deep lateral incision | |||
| mesal ridge protruding in a distal process | |||
| Mesomerite | broad, distally narrowing and hook-shaped | ||
| Solenomerite | broad, narrowing at mid-length | ||
| subapical triangular process | |||
| apical folded lamella | |||
| long and curved seminal groove | |||
| Paracoxite | lamellar, folded, marginally serrated | ||
| Differences | |||
| Promerite | lateral margin well rounded | lateral margin angular | lateral margin reduced and pointed |
| distal process broad and triangular | distal process laterally rounded and bulgy | distal process slender, slightly expanded laterally | |
| Mesomerite | distally slightly broaden and bearing serrations on the lateral margin | distally broaden latero-mesad and bearing strong serrations on both margins | distally strongly constricted and narrowed |
| Solenomerite | |||
| Paracoxite | broad, apical margin rounded | broader, apical margin truncate | slender, apically tapering |
| Non gonopod structures (males) | |||
| Body vertical diameter (mm) | 1.7–2.7 | 2.7–3.6 | 2–2.8 |
| Podous rings | 43–48 | 45–53 | 43–48 |
| Apodous rings | 1–2 | 1–2 | 1–2 |
| Colour | ash-grey alternating with light brown, legs purple | pale grey alternating with golden brown, legs black | dark grey alternating with yellowish brown, legs yellowish brown |
Intercalary males are here recorded for the species Ommatoiulus
khroumiriensis and
Ommatoiulus
crassinigripes.
This is of little surprise considering that postembryonic
development involving periodomorphosis (regression of
secondary sexual characters followed by a return to a
morphologically copulatory stadium following an additional
moult) is considered as particularly prevalent for
schizophylline species (
Colour pattern and somatic characters cannot be used reliably to distinguish the Tunisian Ommatoiulus species, although we have provided information about these features above in order to point out fine differences between morphologically close or syntopic taxa. In the identification key (below) we prefer to include only gonopod characters.
The key is based on characters of the male gonopods.
| 1 | Promerite with parallel margins, apically regularly rounded, without lateral incision (Fig. 43); mesomerite distally expanded into a subrectangular apical plate, concave and mesally incised (Fig. 44) | Ommatoiulus sempervirilis |
| – | Promerite with at least one lateral or apical incision (Figs 2, 7, 14, 20, 32, 45, 47, 49, 51); mesomerite different | 2 |
| 2 | Mesomerite (Ms) apically expanded and bifurcated (Figs 16, 46) | 3 |
| – | Mesomerite (Ms) apically narrowed and simple (Figs 3, 9, 24, 33, 48, 50, 53) | 4 |
| 3 | Mesomerite (Ms) hammer-shaped (Fig. 46), bifurcated into long asymmetrical processes; promerite (Fig. 45) broad, with a complex distal process, apically bearing two blunt bumps, a pointed process and a lateral serrated lamella | Ommatoiulus malleatus |
| – | Mesomerite (Ms) horn-shaped (Fig. 16), bifurcated into short and curved processes; promerite (Fig. 14) apically with three lobes separated by deep incisions | Ommatoiulus kefi sp. n. |
| 4 | Promerite gradually narrowing distally, lateral incision shallow to moderately deep; mesal ridge sometimes protruding in a serrated subapical process (se) (Figs 2, 32, 47); paracoxite (Px) without conspicuous marginal serrations (Figs 5, 34, 48, 50) | 5 |
| – | Promerite strongly narrowing distally, lateral incision deep; mesal ridge protruding in a blunt subapical process (Figs 7, 20, 51); paracoxite (Px) broad, lamellar, folded, with strong marginal serrations (Figs 8, 22, 52) | 10 |
| 5 | Mesomerite (Ms) slender, sinuous and strongly curved distally; solenomerite (S) with a long distal process (ds) lodging the seminal groove (Fig. 48) | Ommatoiulus seurati |
| – | Mesomerite (Ms) broad, distally narrowing; solenomerite (S) with a shorter distal process lodging the seminal groove (Figs 3, 33, 50) | 6 |
| 6 | Mesomerite (Ms) with irregular margins, apically strongly narrowed and hook-shaped; solenomerite (S) with a short distal process (ds) (Figs 37, 50) | 7 |
| – | Mesomerite (Ms) with a regular mesal margin, distally gradually narrowed into a bent folded process; solenomerite (S) with a longer distal process (ds) (Figs 3, 33) | 8 |
| 7 | Mesomerite with a subapical serrated lateral process, a truncated mesal margin, apically strongly narrowed into a small hook-shaped process (Fig. 50); promerite with a shallow lateral incision and a rounded distal process (Fig. 49) | Ommatoiulus fuscounilineatus |
| – | Mesomerite with a subapical lateral process (Figs 37, 39), apically gradually narrowed in a long curved process (Fig. 39); promerite with a deeper lateral incision and a broad distal process with apical and lateral lobes (Fig. 36) | Ommatoiulus zaghouani sp. n. |
| 8 | Promerite straight, bearing a pointed distal process (Fig. 27); paracoxite stout and shorter (Fig. 30) | Ommatoiulus xenos sp. n. |
| – | Promerite strongly bent anteriad, bearing a subapical serrated process (Figs 1, 31); paracoxite broad and elongate (Figs 5, 34) | 9 |
| 9 | Solenomerite (S) with a rounded notch (n), a pointed process (pr) and apically a wrinkled protruding lamella (Fig. 5); promerite with a subtriangular apex (Fig. 2) | Ommatoiulus chambiensis sp. n. |
| – | Solenomerite (S) without such a process; wrinkled lamella not protruding (Fig. 33); promerite with a globular apex (Fig. 32) | Ommatoiulus xerophilus sp. n. |
| 10 | Lateral incision of promerite extending basad, distal process broad and triangular (Fig. 20); mesomerite (Ms) distally strongly narrowed, without subapical serrations on lateral margin, paracoxite (Px) slender (Fig. 22) | Ommatoiulus khroumiriensis sp. n. |
| – | Lateral incision of promerite not extending basad, distal process slenderer and more rounded (Figs 7, 51); mesomerite (Ms) with lateral subapical serrations, paracoxite (Px) broader (Figs 12, 53) | 11 |
| 11 | Apical process of promerite (ap) bulgy, with a lobed process (ap1) on tip (Fig. 7); mesomerite (Ms) with strong subapical mesal serrations (Fig. 8); paracoxite (Px) broadened distally with truncated apex (Fig. 12) | Ommatoiulus crassinigripes |
| – | Apical process of promerite slender, without a lobed process; mesomerite (Ms) without mesal serrations (Fig. 52); paracoxite (Px) distally narrowed, with rounded apex (Fig. 53) | Ommatoiulus punicus |
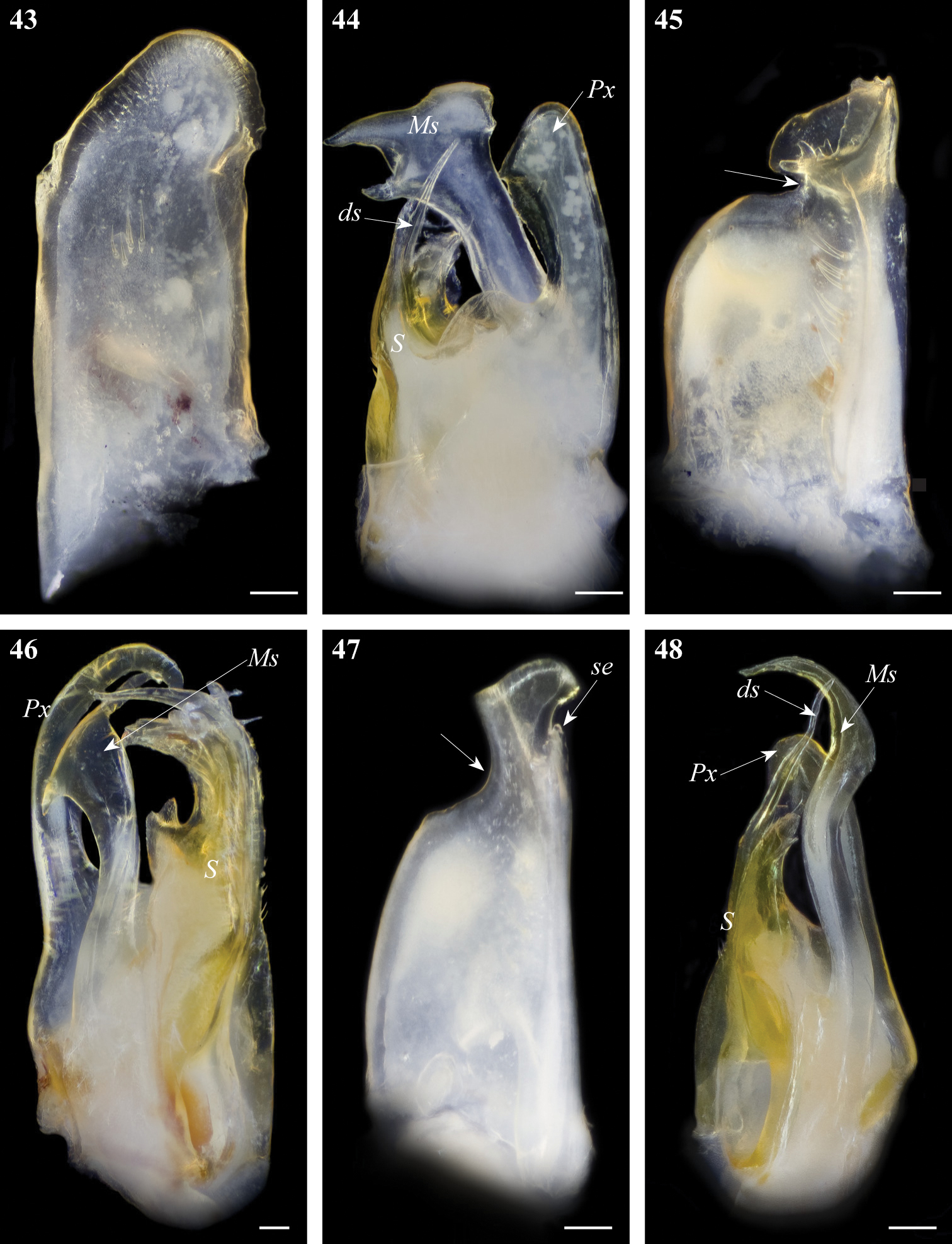
Gonopod structures of Ommatoiulussempervirilis, Ommatoiulus malleatus, Ommatoiulus seurati 43 Ommatoiulus sempervirilis left promerite, posterior view 44 Ommatoiulus sempervirilis, left posterior gonopod, meso-posterior view 45 Ommatoiulus malleatus, right promerite, anterior view 46 Ommatoiulus malleatus, left posterior, mesal view 47 Ommatoiulus seurati right promerite, posterior view 48 Ommatoiulus seurati, right posterior gonopod, antero-mesal view. Abbreviations: ds distal process of the solenomerite, Ms mesomerite, Px paracoxite, S solenomerite, se serrations. Arrow pointing to the lateral incision of the promerite. Scale bar 0.1 mm.
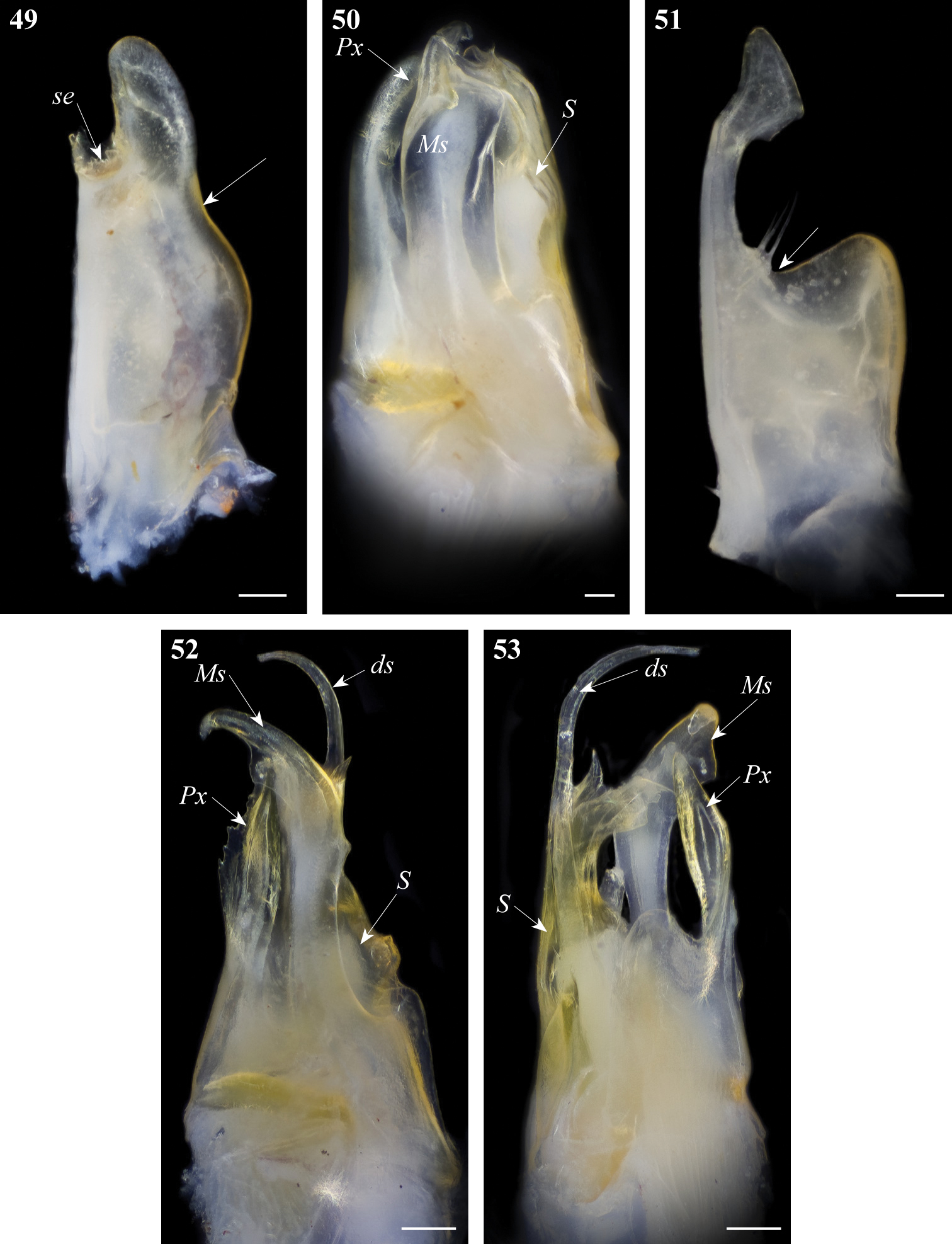
Gonopod structures of Ommatoiulus fuscounilineatus, Ommatoiulus punicus. 49 Ommatoiulus fuscounilineatus, left promerite, posterior view 50 Ommatoiulus fuscounilineatus, left posterior gonopod, anterior view 51 Ommatoiulus punicus, left promerite, posterior view 52 Ommatoiulus punicus, left posterior gonopod, lateral view 53 Ommatoiulus punicus, left posterior gonopod, posterior view. Abbreviations: ds distal process of the solenomerite, Ms mesomerite, Px paracoxite, S solenomerite, se serrations. Arrow pointing to the lateral incision of the promerite. Scale bar 0.1 mm.
We provide an interactive key in Flash (SWF) format to the Ommatoiulus species known from Tunisia. The key is dichotomous and based on gonopod characters. These are illustrated with line drawings, light microscopy photographs and SEMs, and for some of the species with rotatable SEM animations. A species list and species pages are included in the key to provide additional information on species diagnostic characters, distribution and habitats. An introductory section for the first-time user provides background information of importance for applying the key.
Adobe Flash Player (version 11.2 or higher) or a browser (e.g. Internet Explorer, Firefox and Chrome) with Flash Player plug-in is required to run the key.

|
This work was financed by a Carlsberg Foundation grant to N.A. and H.E. We are most grateful to Thomas Pape (ZMUC) for his overall support and comments on the manuscript. Nikolaj Scharff (ZMUC) provided advices about the SEM mounting technique and access to the photographing equipment. Jason Dunlop (Museum für Naturkunde, Berlin), Jean-Jacques Geoffroy (MNHN) and Rosana Pisoni (MSNB) kindly arranged loans under their care. The fieldwork in Tunisia in March 2008 was financed by the Field Museum Collection Fund. N.A. and P.S. are grateful to Petra Sierwald (Chicago, USA) for the logistic help. Specimens were collected in national parks and reserves with a permit from “La direction générale des forêts, ministère d’Agriculture” of Tunisia. NA and PS are thankful to Ridha Ouni and Malek Akkari for their assistance during the filed work. N.A.’s short visit to MNHN, March 2013, was financed by the EC FP7 Project SYNTHESYS. Many thanks are also due to Bob Mesibov (Queen Victoria Museum and Art Gallery, Tasmania) and two anonymous reviewers for their useful comments on the manuscript. The publication of this article was made possible with financial support from the EC FP7 project ViBRANT (www.vbrant.eu) to P.S.
Source image library of the interactive SEM image of Ommatoiulus chambiensis (doi: 10.3897/zookeys.328.5763.app1) File format: WinZip Image Archive (jpg).
Source image library of the interactive SEM image of Ommatoiulus crassinigripes (doi: 10.3897/zookeys.328.5763.app2) File format: WinZip Image Archive (jpg).
Source image library of the interactive SEM image of Ommatoiulus kefi (doi: 10.3897/zookeys.328.5763.app3) File format: WinZip Image Archive (jpg).
Source image library of the interactive SEM image of Ommatoiulus khroumiriensis (doi: 10.3897/zookeys.328.5763.app4) File format: WinZip Image Archive (jpg).
Source image library of the interactive SEM image of Ommatoiulus xerophilus (doi: 10.3897/zookeys.328.5763.app5) File format: WinZip Image Archive (jpg).
Source image library of the interactive SEM image of Ommatoiulus punicus (doi: 10.3897/zookeys.328.5763.app6) File format: WinZip Image Archive (jpg).
Source file library of the interractive key to genus Ommatoiulus in Tunisia (doi: 10.3897/zookeys.328.5763.app7) File format: WinZip Archive (zip).