





(C) 2013 Guo-Liang Xu. This is an open access article distributed under the terms of the Creative Commons Attribution License 3.0 (CC-BY), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
For reference, use of the paginated PDF or printed version of this article is recommended.
The genus Acrocrytus is reported from Chinese mainland for the first time, with description of two new species Acrocyrtus zhujiensis sp. n. and Acrocyrtus finis sp. n. from Zhejiang Province, East China. They can be separated from other species of this genus by colour pattern, unscaled appendages (antennae, legs and ventral tube), interocular chaetae, labial basal chaetae, bothriotrichal complex chaetae on Abd. II–IV, microchaeta a2 on Abd. II, im on Abd. III and C1p on Abd. IV. Illustrations and a table showing main differences with closest Acrocyrtus species are provided.
A. zhujiensissp. n. , A. finissp. n. , chaetotaxy, China
Acrocyrtus was established by
Acrocyrtus is characterized by the presence of conical pointed dental tubercles, rounded and finely striated scales on body and ventral side of furcula, 8+8 ommatidia (G and H smaller), 4-segmented antennae and apical bulb absent on Ant. IV, bidentate mucro with or without accessory spinelet. It is widely distributed in Southeast Asia, such as Singapore, Malay and Indonesia. So far, more than 26 species of the genus Acrocyrtus were described all over the world (
The specimens were cleared in lactic acid, mounted under a coverslip in Marc André II solution, observed using Leica DM2500 and Nikon 80i microscopes. The photographs were taken with Nikon SM1000 microscope using a mounted Nikon DS-Fi1 camera and enhanced with Photoshop CS2. Length data were measured with NIS-Elements Documentation (Nikon). Dorsal cephalic chaetae were designated after
Abbreviations.Th. -thoracic segment; Abd. -abdominal segment; Ant. -antennal segment; ms -specialized S-microchaeta(e); S-chaeta(e) -specialized chaeta(e) (including ms); mac -macrochaeta(e); mic -microchaeta(e).
urn:lsid:zoobank.org:act:D8F04C9C-6364-44CF-AA8E-B5B86580D831
http://species-id.net/wiki/Acrocyrtus_zhujiensis
Figs 1–27, Table 1♀ on slide, Shaoxin City, Zhuji Country, Dongbaihu, Zhejiang Province, CHINA, 29°34.18'N, 120°24.06'E, 3.X.2009, collection number S4014, collected by Zhi-Xiang Pan & Chen-Chong Si, deposited in Taizhou University.
6 ♀ and 1 ♂ on slide and 10 in alcohol, same data as holotype. 4 paratypes (2 ♀ on slide and 2 in alcohol) deposited in School of Life Sciences, Nanjing University and others in School of Life Sciences, Taizhou University, China.
Named after the type locality.
Body length up to 0.93 mm.
Colour pattern. Ground colour pale yellow, with a pair of dark patches present on lateral Abd. III. Violet pigment distributed on antennae and gradually darker towards tip. Eye patches dark (Fig. 1). Scales hyaline, oval to circular (Fig. 2), present on head, body tergites, ventral side of furcula; antennae, ventral tube and legs unscaled.
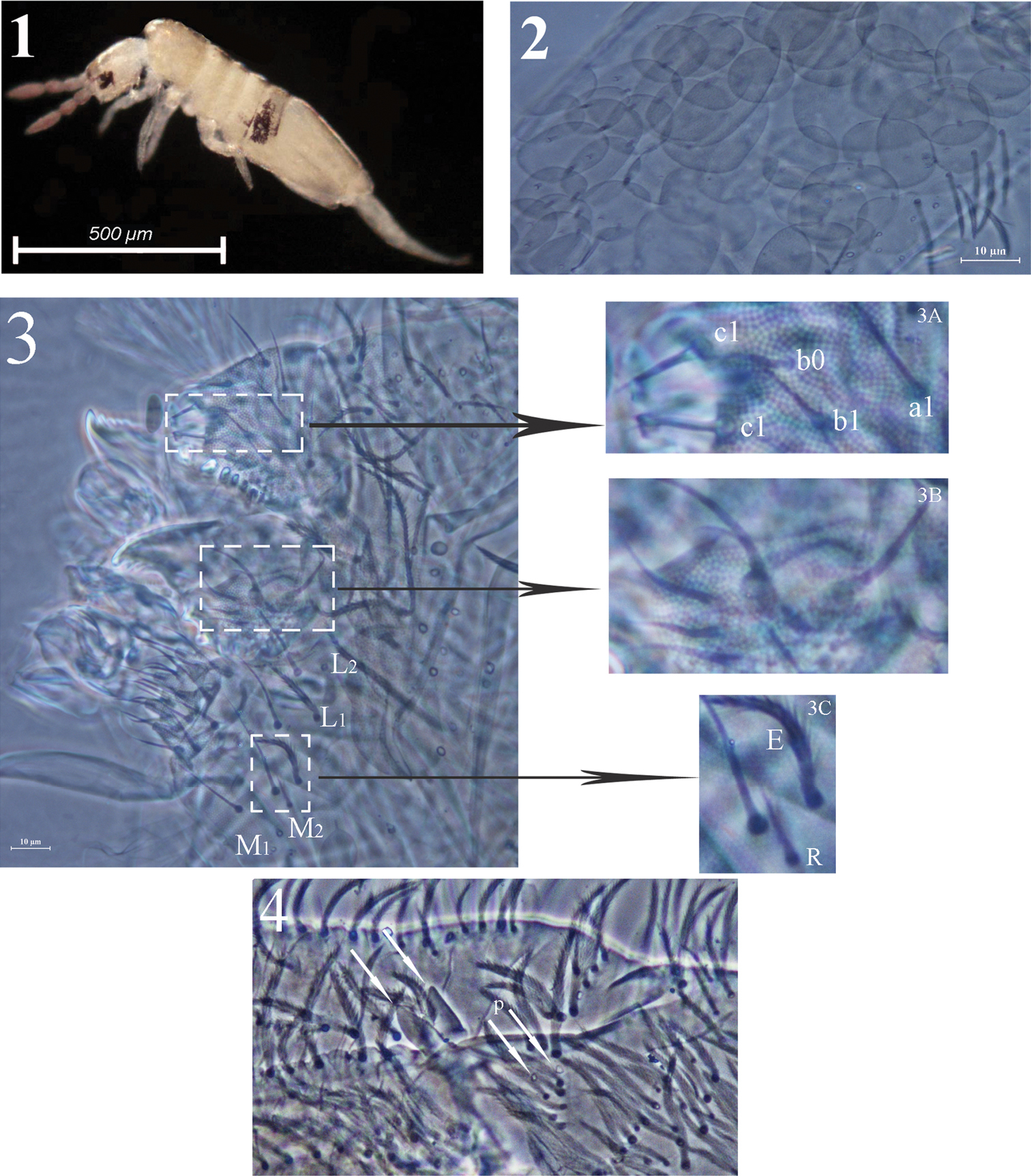
Acrocrytus zhujiensis sp. n. 1 colour pattern, lateral view 2 body scale 3 labium and labrum (3A labral intrusion 3B maxillary outer lobe 3C labial basal chaetae R and E) 4 apical manubrium and basal dentes.
Head. Ommatidia 8+8, G and H smaller than others. Interocular chaetae as p, r, t, q, s, v; chaeta s smooth, chaetae r and v transformed to scales, chaetae p, t and q ciliate (Fig. 5). Antennae 1.5–2.4 times as long as cephalic diagonal. Antennal segmental ratio as I:II:III:IV = 1:1.3–1.4:1.2–1.9:1.9–3.1. Ant. I with 3 dorsal and 3 ventral basal spiny mic (Fig. 6). Ant. II with 4 basal tiny spiny mic, 1 distal rod-like and 12–15 normal S-chaetae (Fig. 7). Ant. III organ with 2 rod-like S-chaetae (Fig. 8). Ant. IV without apical bulb. Anterior part of head with many long, ciliate chaetae but not claviform (Fig. 5). Prelabral and labral chaetae as 4/5, 5, 4, prelabrals ciliate and others smooth, labral intrusion V-shaped, chaetae of c-row thicker than those in other rows; labral margin with 4 conical papillae (Fig. 3). Clypeal chaetae as 3-1-4, without scales between them (Fig. 9). Cervical chaetae as 16 spiny chaetae, lateral two slightly longer than others (Fig. 10). Subapical chaeta of maxillary outer lobe subequal to apical one, 3 smooth sublobal hairs on sublobal plate. Labial palp with 5 papillae as A–E, respectively with 0, 5, 0, 4, 4 guard chaetae; lateral process (l.p.) of labial palp straight, thick and blunt with tip not reaching apex of papilla E. Chaetotaxy of labial base as M1M2REL1L2, all ciliate, chaeta R shorter than others (Fig. 11). Chaetal row along labial groove with 3 ciliate chaetae, and other postlabial chaetae ciliate (Fig. 12). Mandible with 4+5 (left+right) teeth (Fig. 3).
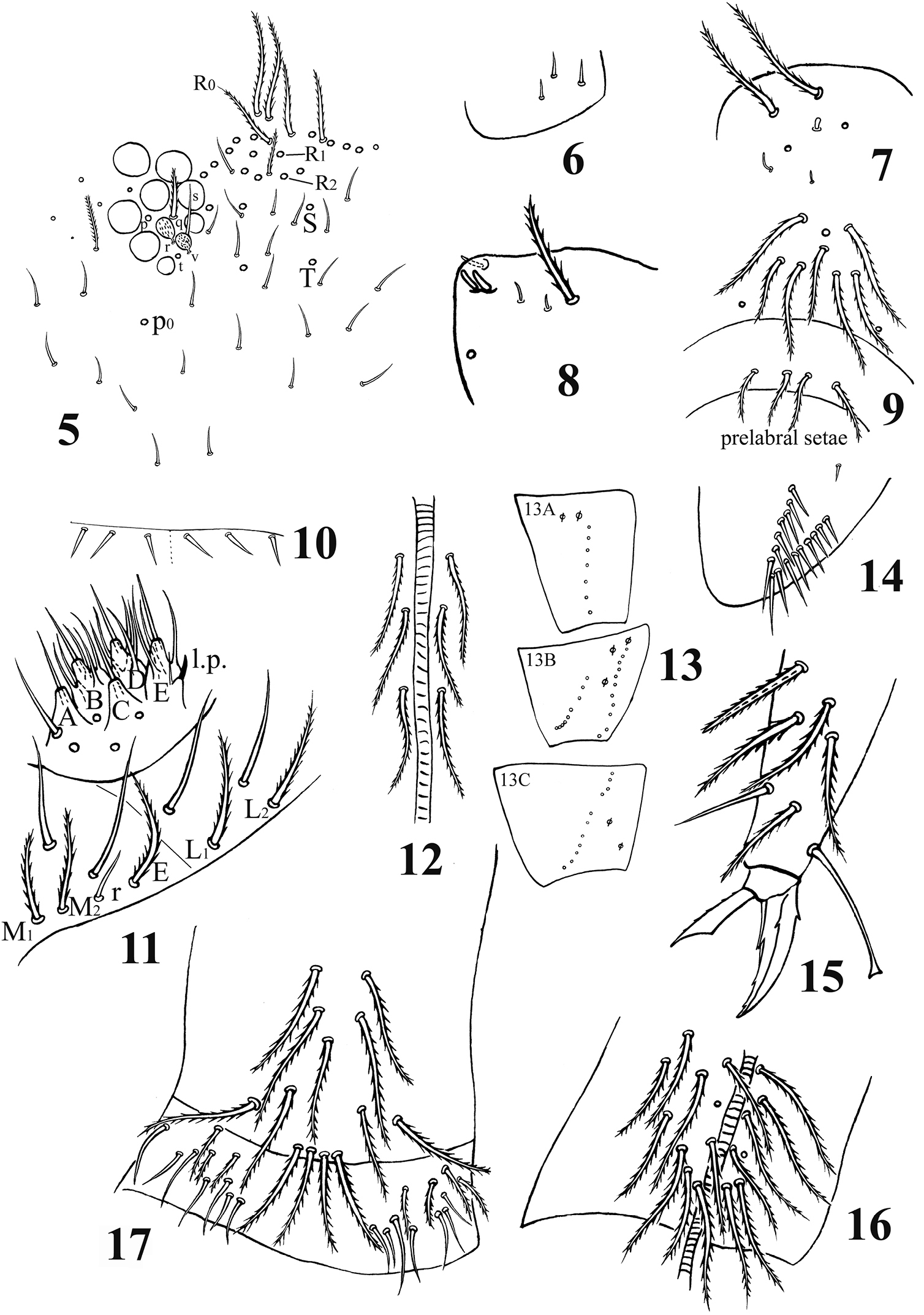
Acrocrytus zhujiensis sp. n. 5 head cheatotaxy 6 basal Ant. I 7 three kinds S-chaetae on Ant. II 8 Ant. III organ 9 clypeal chaetae 10 cervical chaetae 11 labial base and labial palp 12 cephalic groove 13 coxal macrochaetae (13A fore legs 13B mid legs 13C hind legs) 14 trochanteral organ 15 hind claw 16 anterior side of ventral tube 17 posterior side and lateral flap of ventral tube.
Leg. Coxae: I, with 7 ciliate mac and 2 pseudopores; II, with 7–8 ciliate mac in the anterior row, 8–11 ciliate mac in the posterior row and 3 pseudopores; III, with 9–11 ciliate mac and 2 pseudopores (Fig. 13). Trochanteral organ with 12–17 smooth spines (1–2 inner) (Fig. 14). Unguis with 4 inner teeth (paired ones at 1/3, middle one at 2/3 and apical one at 3/4 distance from base), 2 lateral teeth (at 1/4 distance from base) and 1 outer tooth (at 1/5 distance from base). Unguiculus slender and truncate with outer edge serrate. Tenent hair clavate, subequal to inner edge of unguis in length, and slightly longer than unguiculus. A distal smooth chaeta on tibiotarsus III subequal to unguiculus in length (Fig. 15).
Ventral tube. Anterior face with 14+14 ciliate chaetae; posterior face without smooth chaeta (Fig. 16); lateral flap with 6–8 smooth and 2–4 ciliate chaetae (Fig. 17).
Furcula. Manubrial plaque with 2–3 inner, 4–6 outer ciliate chaetae and 2 pseudopores. Dental tubercles conically pointed (Figs 4, 18). Ventral terminal manubrium with 2+2 ciliate chaetae (Fig. 19). Distal smooth part of dentes 2.1–2.5 times as long as mucro. Mucro bidentate, mucronal basal spine reaching subapical tooth with an accessory spinelet (Fig. 20).
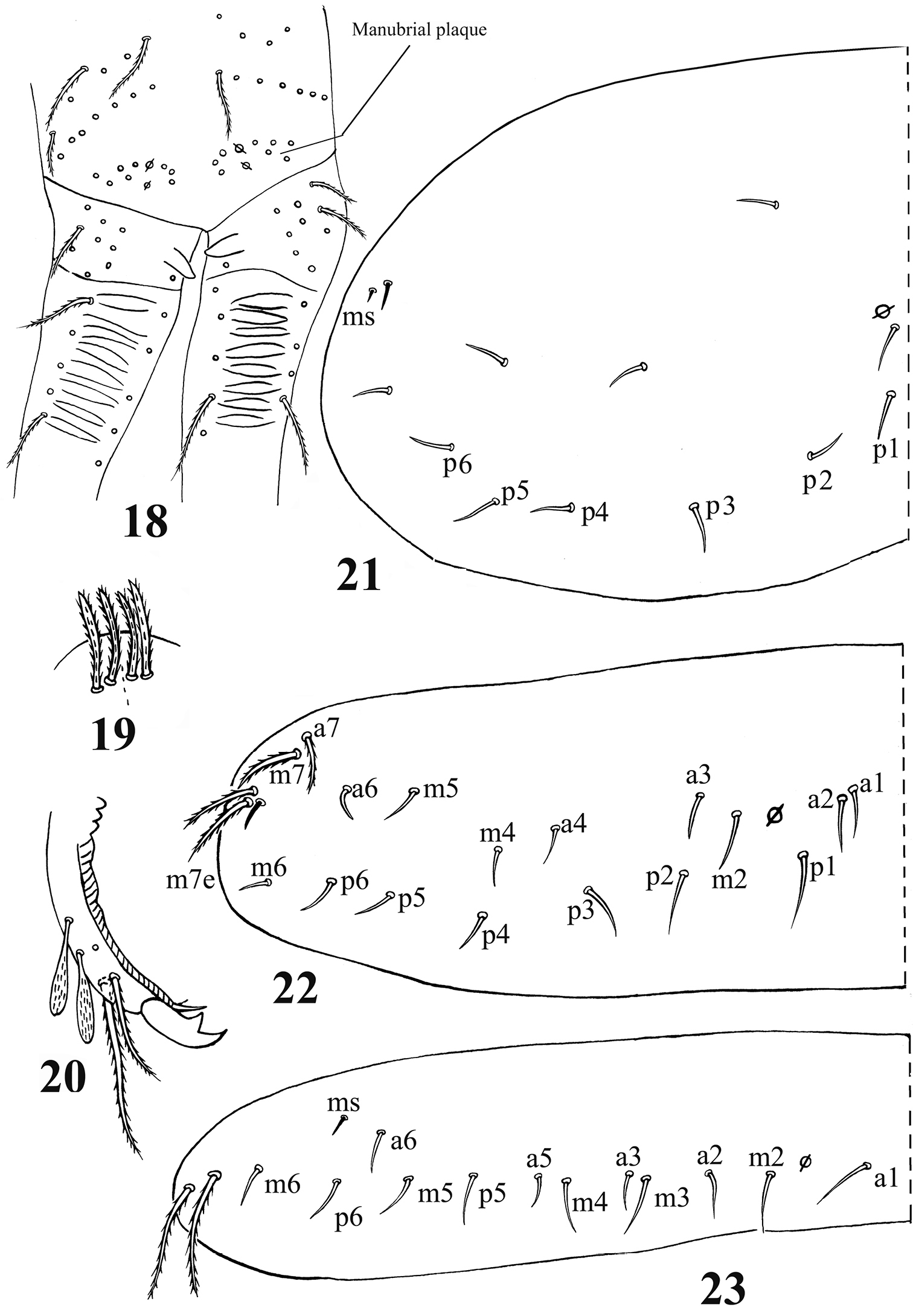
Acrocrytus zhujiensissp. n. 18 distal manubrium and basal dens 19 distal part of ventral manubrium 20 mucro 21–23 dorsal chaetotaxy 21 Th. II 22 Th. III 23. Abd. I.
Chaetotaxy. Dorsal cephalic mac as R0R1R2STP0; R1s absent. Body mac as 00/0100+3, S-chaetae as 21/11253, ms as 10/10100. Th. II slightly protruded over head, with 1–2 rows of ciliate “collar” mac, 2 antero-lateral S-chaetae (ms external to another S-chaeta), 6 (p1–6) smooth mic and 5 anterior unclear homology smooth mic (Fig. 21). Th. III with 1 S-chaeta external to m7; 15 (a1–4, a6, m2, m4–6 and p1–6) smooth mic, 3 (a7, m7 and m7e) ciliate mac and a lateral unclear homology ciliate chaeta (Fig. 22). Abd. I with 1 ms, 12 (a1–3, a5–6, m2–6 and p5–6) smooth mic and 2 lateral unclear homology ciliate chaetae (Fig. 23). Abd. II with 1 central S-chaeta (as), 1 (a2) ciliate mic, 13 (a3, a6–7, m3e, m4, m6–7, p4–7, p5p and el) smooth mic, 1 ciliate and slightly modified mic (mi), 2 (Lm and Li) ciliate, modified and fan-shaped mic, 2 (m3 and m5) ciliate mac; chaetae a2p and ml absent (Fig. 24). Abd. III with 1 central S-chaeta (as) and 1 lateral ms, 5 (mi, ml, a2, im, em and am6; ml sometimes present on one side) ciliate and slightly modified mic, 4 (Li, Lm, Ll and a6) strongly modified and fan-shaped mic, 8 (a3, a7, m3, m7, p3–5 and p7) smooth and subequal mic, 3 (pm6, m7a and p6) ciliate mac (Fig. 25). Abd. IV with 1 anterior (as) and 1 posterior (ps) short S-chaetae and 3 median elongate S-chaetae, 22 (A2–6, B2–3, Be2, C1–4, T1, T3, T5–6, Te3, D2–3, D1p, E1 and Fe1) smooth mic, 2 (C1p and T7) ciliate mic, 6 (m, a, s, D1, pi and pe) ciliate, strongly modified and fan-shaped mic, 13 (B4–6, De1, De3, D3p, E2–4, F2–3, F3p2, Fe4 and Fe6) ciliate mac, 4(E4p, E4p2, F2p and F3p2) mic (Fig. 26). Abd. V with 3 S-chaetae (Fig. 27).
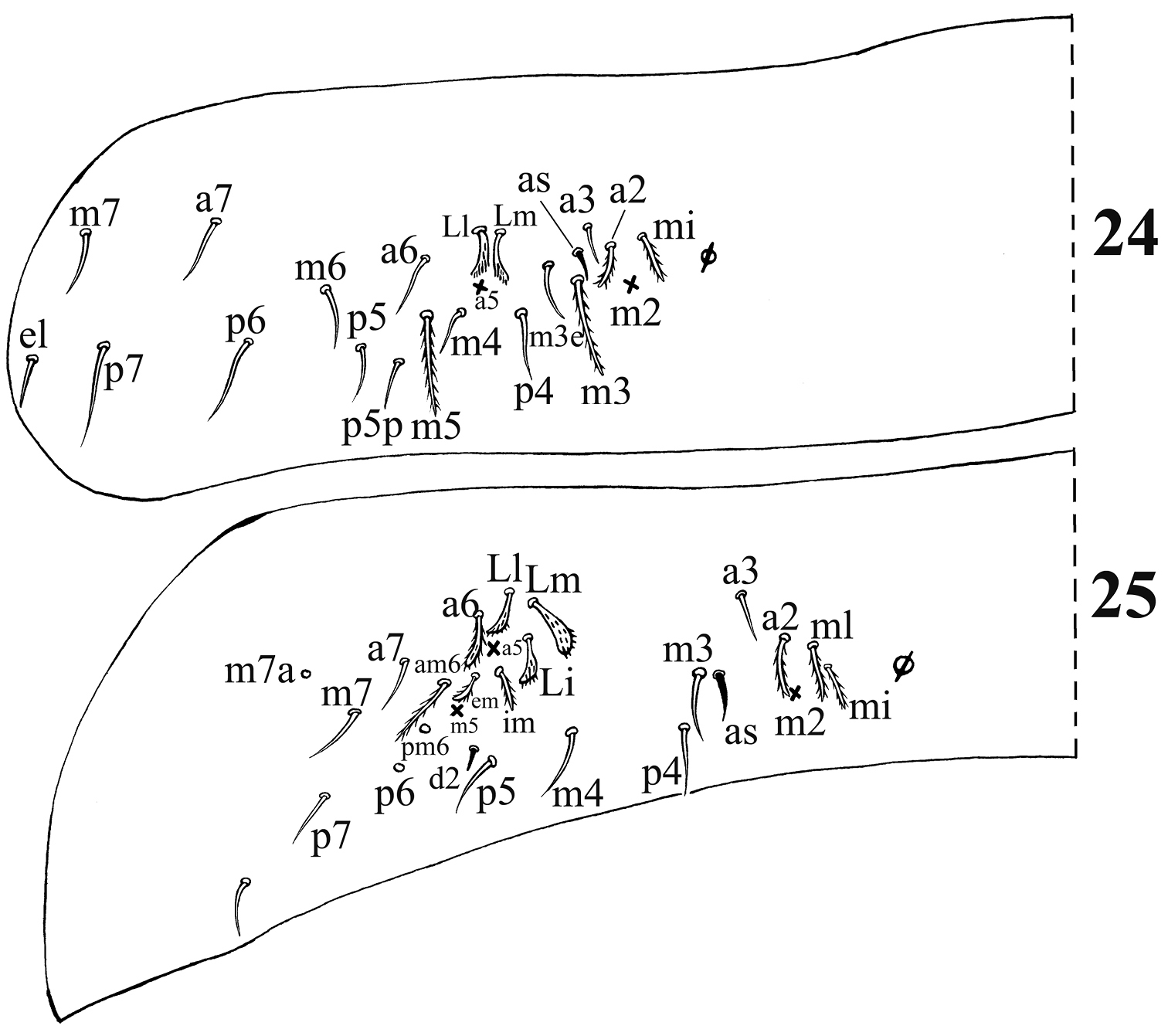
Dorsal chaetotaxy of Acrocrytus zhujiensis sp. n. 24 Abd. II 25 Abd. III (as: antero-lateral S-chaeta).
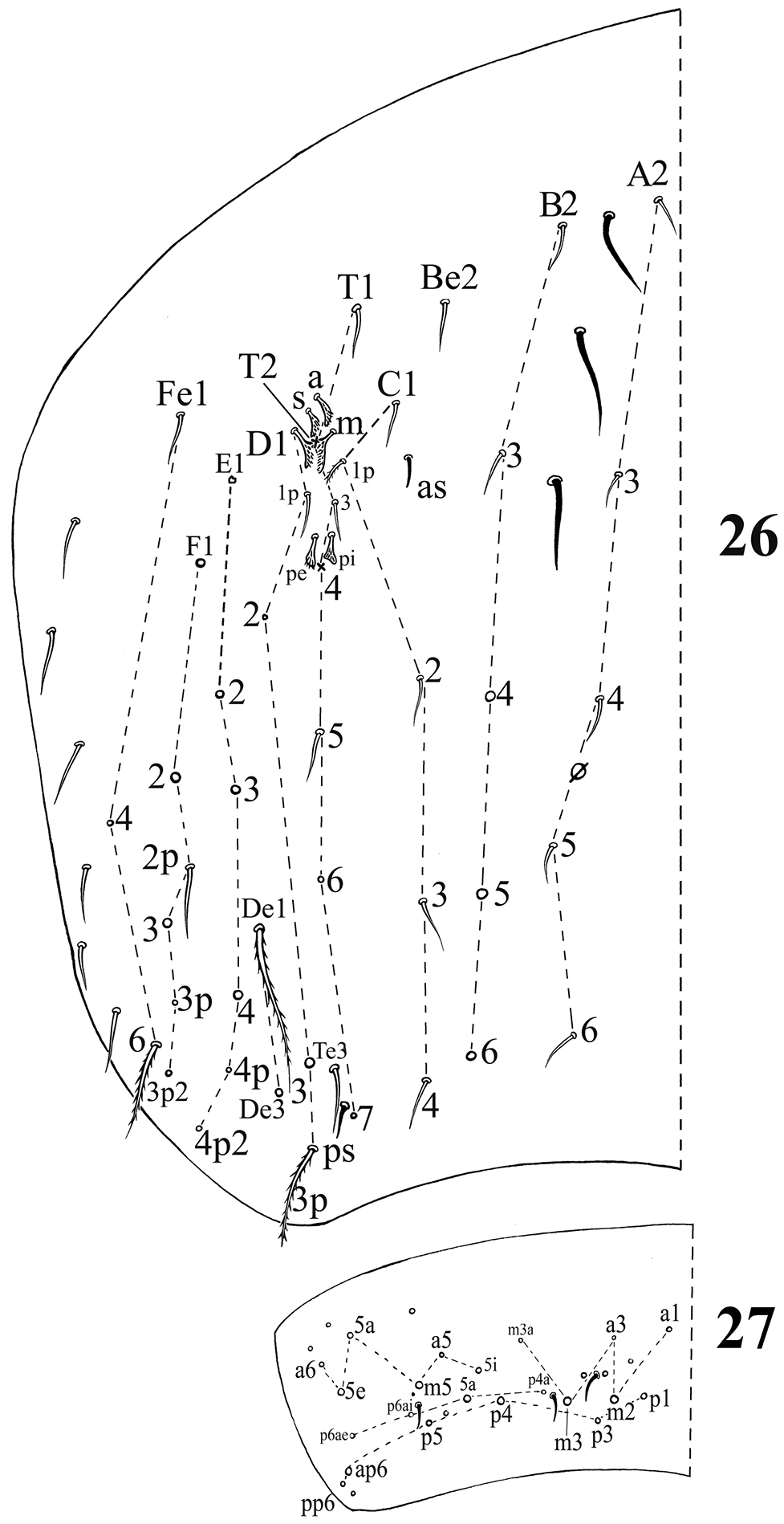
Dorsal chaetotaxy ofAcrocrytus zhujiensis sp. n. 26 Abd. IV 27 Abd. V (ps: postero-sublateral S-chaeta).
In the leaf litter of Cunninghamia lanceolata, Cinamomum camphora along a lake.
This new species is characterized by colour pattern, clavate tenent hair, unscaled appendages (antennae, legs and ventral tube), 4 conical labral papillae, ventral tube with ciliate chaetae present on lateral flap and without smooth chaeta on posterior side, and ciliate mic a2 on Abd. II.
It is similar to Vietnamese Acrocyrtus baii Nguyen, 2005 in clavate tenent hair, claw, unscaled appendages (antennae and ventral tube). However, it can be easily distinguished from it by pigment absent on Th. II–III and Abd. II (versus present), ciliate labial chaetae EL1L2 (versus smooth), M1 subequal to M2 on labial base (versus M1 smaller than M2), absence of smooth chaetae on posterior face of ventral tube (versus 1+1 smooth chaetae) and unscaled legs (versus scaled).
urn:lsid:zoobank.org:act:0E689089-71E5-4FD4-AD50-3A2B23CE5595
http://species-id.net/wiki/Acrocyrtus_finis
Figs 28–37, Table 11 ♀ on slide, Taizhou City, Dalei Mountain, Zhejiang Province, CHINA, 29°02.25'N, 120°53.03'E, 25.X.2009, collection number S4023, collected by Zhi-Xiang Pan, deposited in Taizhou University.
11 ♀ on slide and 15 in alcohol, same data as holotype. 4 paratypes (2 ♀ on slide and 2 in alcohol) deposited in School of Life Sciences, Nanjing University and others in School of Life Sciences, Taizhou University, China.
Named after the type locality, which is the border (latin word “finis”) of the three adjacent cities.
Body length up to 1.2 mm.
Colour pattern. Ground colour from yellow to slightly brown, a pair of dark lateral patches of Abd. III and a pair of dark postero-lateral patches of Abd. IV, slightly violet pigment distributed on antennae and gradually darker towards tip, eye patches dark (Fig. 28). Scales hyaline, oval to circular (Fig. 29), present on head, body tergites and ventral side of furcula, and absent on antennae, legs and ventral tube.
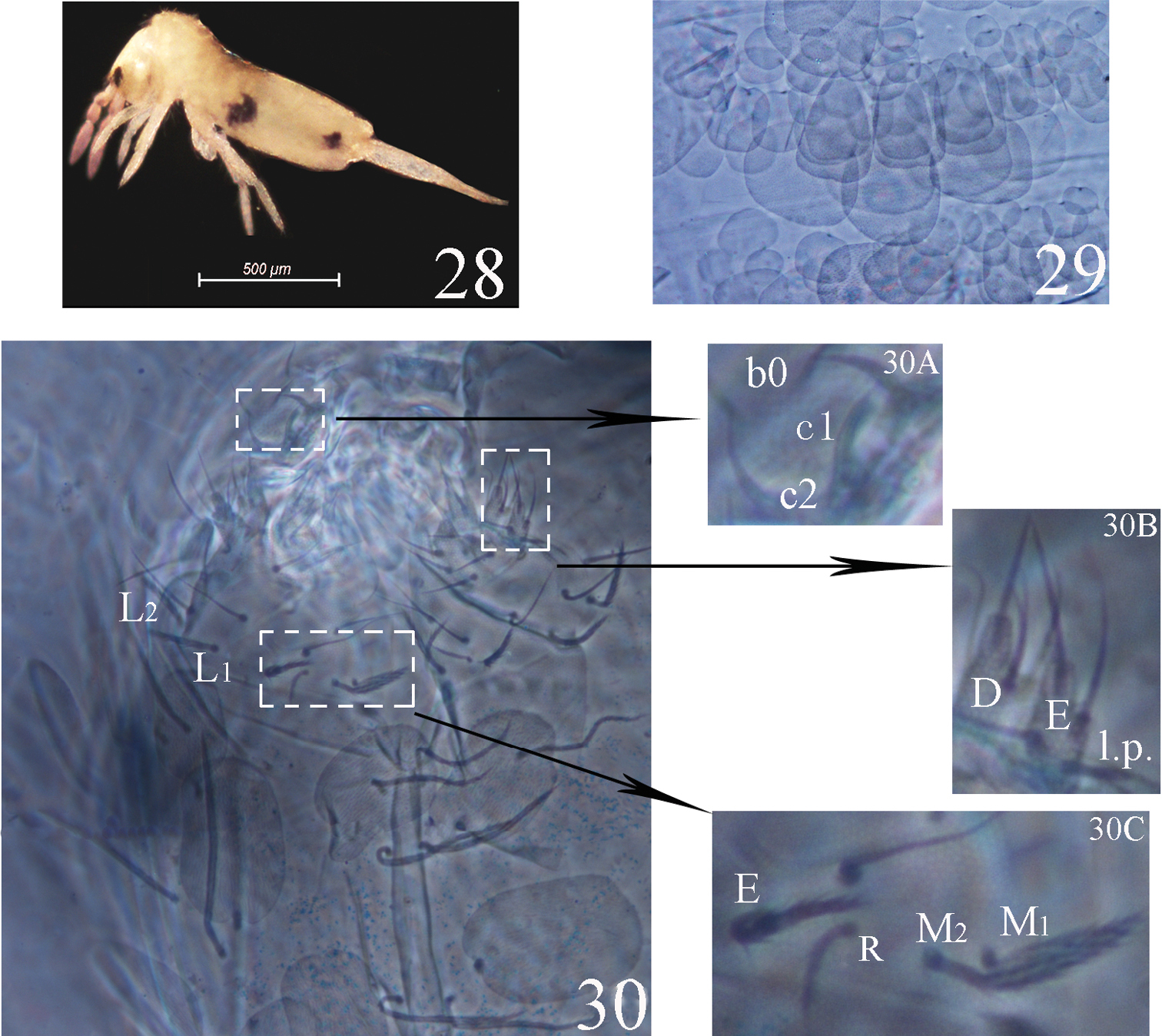
Acrocrytus finissp. n. 28 colour pattern, lateral view 29 body scale 30 labium and labrum (30A labral intrusion 30B labial papillae D, E and l.p. 30C labial base).
Head. Ommatidia 8+8, G and H smaller than others, interocular chaetae as p, r, t, q, s, v; chaeta s smooth, chaetae p, t, q ciliate, chaetae r and v transformed to scales. Antennae 1.4–2.0 times as long as cephalic diagonal. Antennal segmental ratio as I:II:III:IV = 1:1.3–1.9:1.5–2.5:2.5–5.0. Ant. I with 3 dorsal and 3 ventral basal spiny chaetae. Ant. II with 4 basal tiny spines, 11–14 short and 1 distal rod-like S-chaetae. Ant. III organ with 2 rod-like S-chaetae. Ant. IV without apical bulb. Anterior part of head with many ciliate and long, but not claviform chaetae. Cervical with 16 spiny chaetae, all subequal in length. Prelabral and labral chaetae as 4/5, 5, 4, prelabrals ciliate and others smooth, chaetae of c-row thicker than other row chaetae; labral intrusion V-shape; labral margin with 4 papillae. Clypeus without scales. Subapical chaeta of the maxillary outer lobe subequal to apical chaeta, 3 smooth sublobal hairs on sublobular plate. Labial palp with five papillae as A–E, respectively with 0, 5, 0, 4, 4 guard chaetae; lateral process of labial palp straight, thick with tip not reaching apex of papilla E. Chaetotaxy of labial base as M1M2REL1L2, all ciliate, chaeta r shorter than others. Chaetal row along labial groove with 3 ciliate chaetae, and other postlabial chaetae ciliate. Mandible with 4+5 (left+right) teeth (Fig. 30).
Leg. Coxae: I, with 5–7 ciliate mac and 2 pseudopores; II, with 6–7 ciliate mac in the anterior row, 7–9 ciliate mac in the posterior row and 3 pseudopores; III, with 6–7+3 ciliate mac and 2 pseudopores. Trochanteral organ with 10–14 smooth spiny chaetae. Unguis with 1 outer (at 1/5 distance from base), 2 lateral (at 1/4 distance from base) and 4 inner teeth (paired ones at 1/3, middle one at 2/3 and apical one at 3/4 distance from base to apical inner unguis), all tiny. Unguiculus slender and truncate with outer edge slightly serrate. Tenent hair clavate, subequal to inner margin of unguis, and slightly longer than unguiculus. Supraempodial chaeta subequal to unguiculus.
Ventral tube. Anterior face with 9–15 larger ciliate chaetae; posterior face without smooth chaetae; lateral flap with 6–7 smooth and 2–3 ciliate chaetae.
Furcula. Manubrial plaque with 2–3 inner, 4–6 outer ciliate chaetae and 2 pseudopores, ventral manubrium with 2+2 ciliate terminal chaetae. Dental tubercles conically pointed. Distal smooth part of dentes 1.5–2.0 times as long as mucro. Mucro bidentate, mucronal basal spine reaching subapical tooth with two accessory spinelets.
Chaetotaxy. Dorsal cephalic mac as R0R1R2TS, P0 sometime absent. Body mac as 00/0100+3, S-chaetae as 21/11253, ms as 10/10100. Th. II slightly protruded over head, with 2 antero-lateral S-chaetae (ms postero-external to another one), 6 (p1–6) smooth and subequal mic and 5 anterior smooth mic of unclear homology (Fig. 31). Th. III with 1 S-chaetae external to m7, 15 (a1–4, a6, m2, m4–6 and p1–6) smooth mic, 3 (a7, m7 and m7e) mac and one other mac of unclear homology (Fig. 32). Abd. I with 1 ms external to a6, 12 (a1–3, a5–6, m2–6 and p5–6) smooth mic and 2 lateral ciliate mac of unclear homology (Fig. 33). Abd. II with 1 central S-chaetae (as), 1 (mi) ciliate and blunt mic, 14 (a2–3, a6–7, m3e, m4, m6–7, p4–5, p5p, p6–7 and el) smooth and subequal mic, 2 (Lm and Li) ciliate and slightly expanded mic, 2 (m3 and m5) ciliate mac with tip expanded, chaetae a2p and ml absent (Fig. 34). Abd. III with 1 central S-chaeta (as) and 1 lateral ms, 5 (a2, mi, ml, em and am6) ciliate mic with tip expanded, 4 (Li, Lm, Ll and a6) ciliate modified and fan-shaped mic, 9 (im, a3, a7, m3, m7, p3–5 and p7) smooth mic, 3 (pm6, m7a and p6) ciliate mac (Fig. 35). Abd. IV with 1 anterior (as) and 1 posterior (ps) short S-chaetae and 3 elongate median S-chaetae, 22 (A2–6, B2, B3, Be2, C1–4, C1p, T1, T3, T5–6, D1p, D2–3 and Fe1) smooth mic, 15 (B4–6, T7, D3p, De1, De3, E1–4, F1–3, Fe4 and Fe6) ciliate and mac, 5 (Te3, E4p, E4p2, F3p and F3p2) as mic, 4 (m, a, s and D1) ciliate and strongly fan-shaped mic, 2 (pi and pe) ciliate mic with tip expanded (Fig. 36). Abd. V with 3 S-chaetae (Fig. 37). Abd. IV:Abd. III = 2.6–4.1:1.
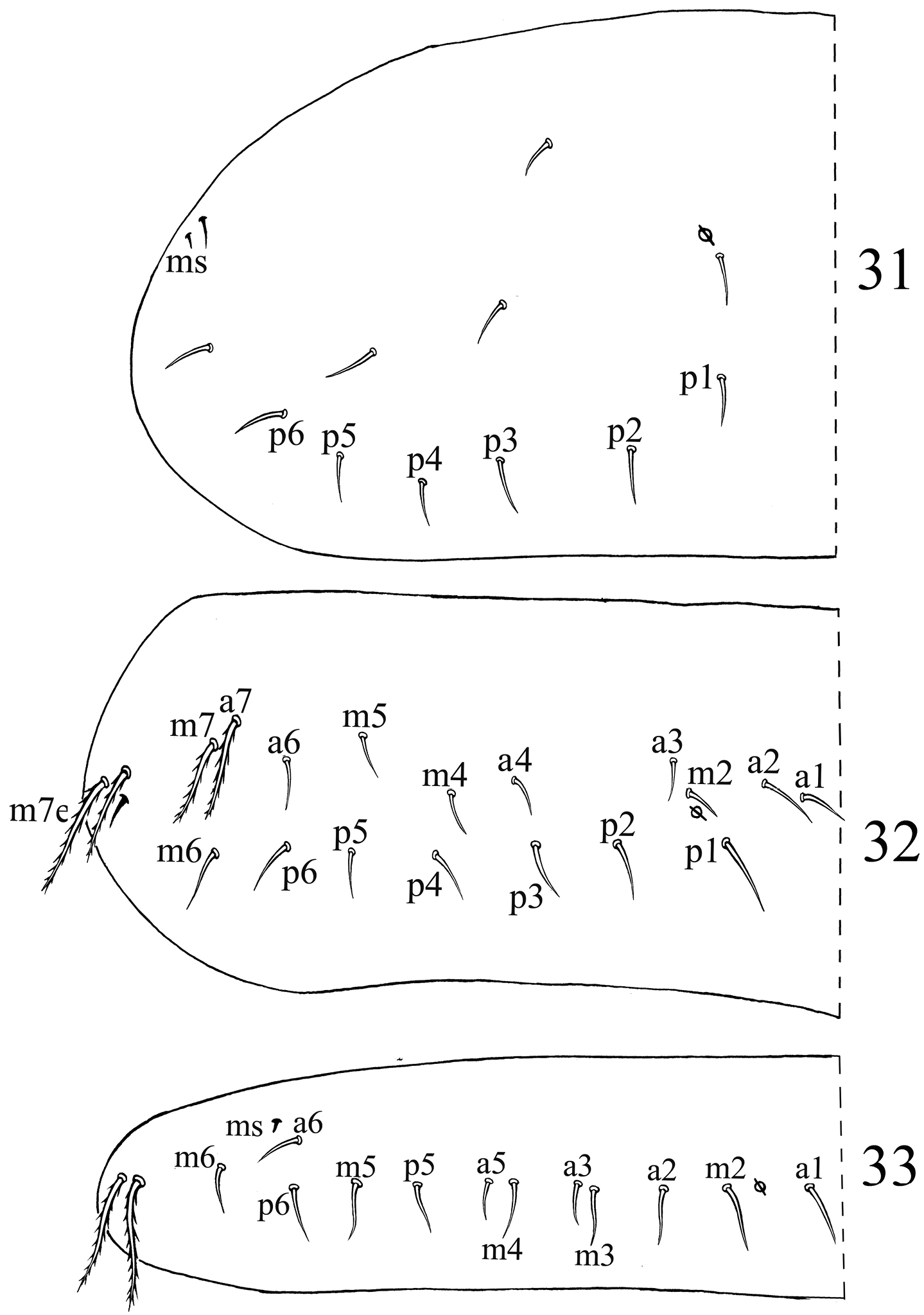
Dorsal chaetotaxyof Acrocrytus finis sp. n. 31 Th. II 32 Th. III 33 Abd. I.
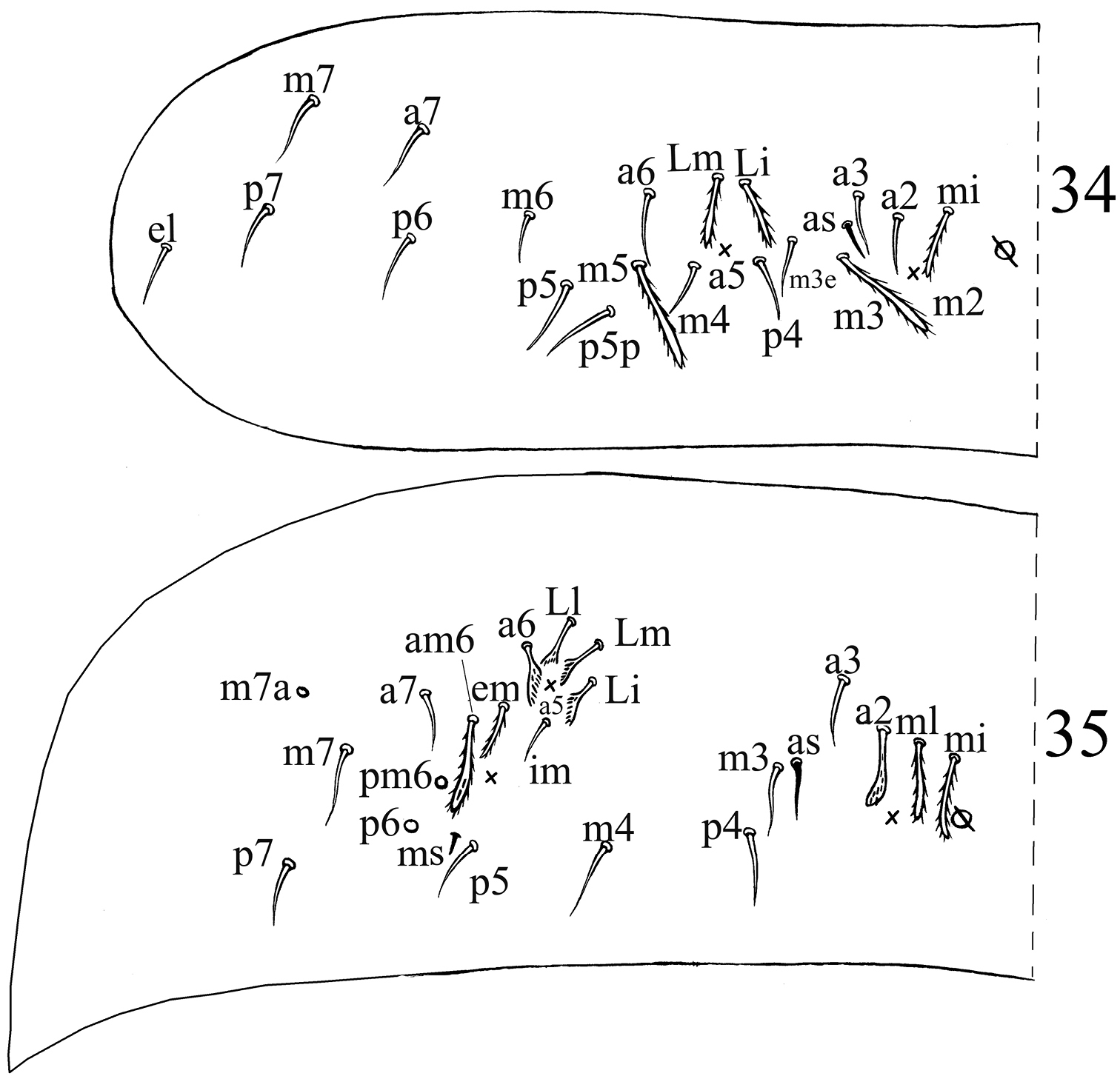
Dorsal chaetotaxyof Acrocrytus finis sp. n. 34 Abd. II 35 Abd. III.
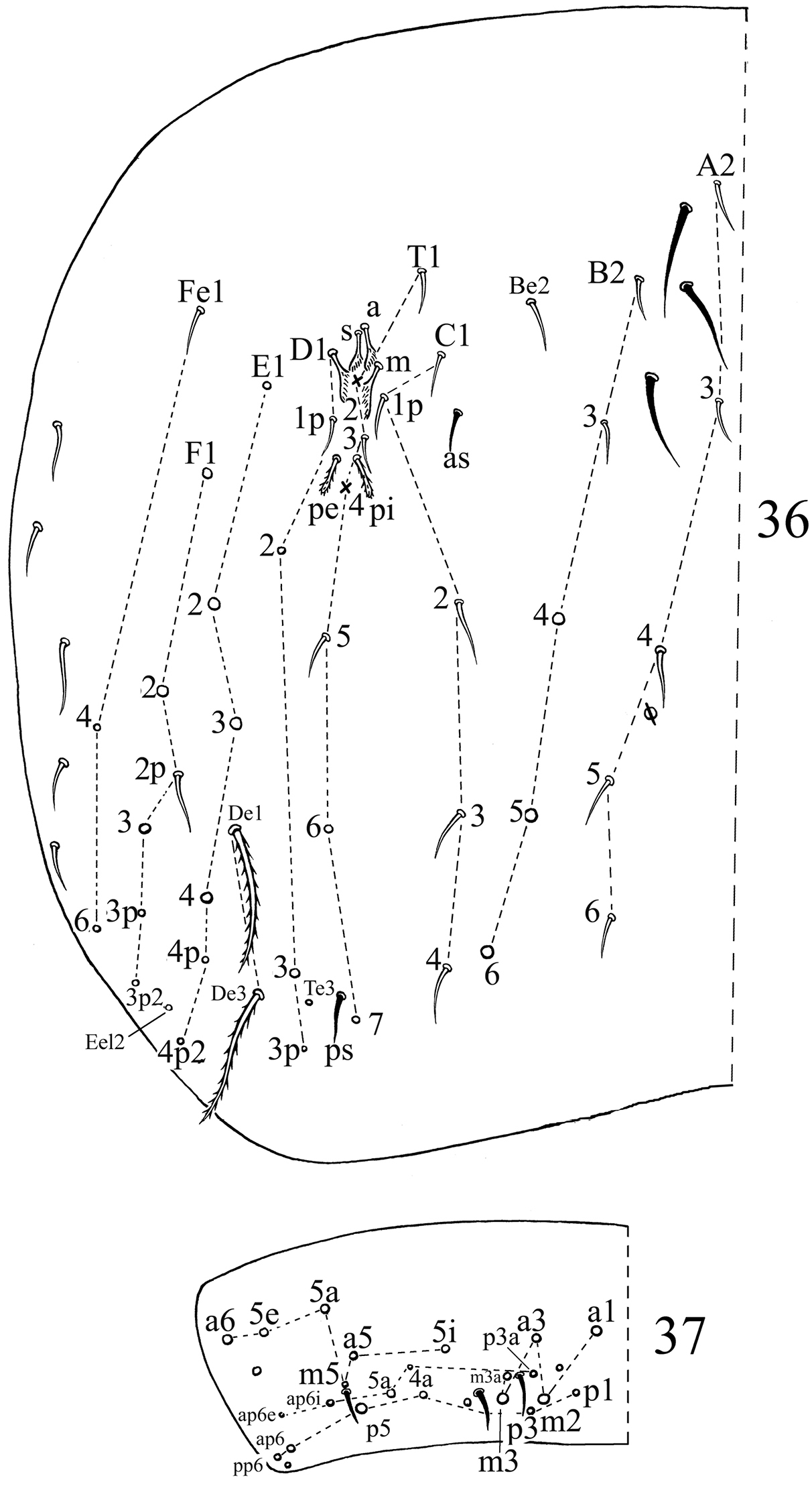
Dorsal chaetotaxyof Acrocrytus finis sp. n. 36 Abd. IV 37 Abd. V.
In leaf litter of Pinus massoniana, Schima superba and Cinamomum camphora, in bryophyta and on farmland.
The new species is easily distinguished from other Acrocyrtus by 4 abdominal lateral patches, morphology of interocular chaetae v, r and s, cephalic mac as R0R1R2ST, 4 papillae and thicken c-row chaetae on labrum, smooth mic a2, im and C1p on Abd. II, Abd. III and Abd. IV, respectively, unscaled appendages (antennae, ventral tube and legs).
The species is most similar to Acrocyrtus zhujiensis sp. n. in cephalic chaetotaxy, labral papillae, claw, furcula, macrochaetotaxy and S-chaetotaxy. However, the two species are different in colour pattern, morphology of chaetae a2, m3 and m5 on Abd. II, a2 and im on Abd. III, C1p on Abd. IV. Main differences between two new species are listed in Table 1.
Main differences between three similar species of Acrocyrtus.
| Acrocrytus zhujiensis sp. n. | Acrocrytus finis sp. n. | Acrocrytus baii | |
|---|---|---|---|
| Dark patches laterally on Abd. II | absent | absent | present |
| Dark patches postero-laterally on Abd. IV | absent | present | present |
| Apical bulb of Ant. IV | absent | absent | present |
| Number of labral papillae | 4 | 4 | 0 |
| Chaeta M2 on labial base | ciliate | ciliate | smooth |
| Chaeta R on labial base | slightly ciliate | slightly ciliate | reduced |
| Chaetae EL1L2 on labial base | ciliate | ciliate | smooth |
| Inner teeth on unguis | 4 | 4 | 3 |
| Smooth chaetae on posterior ventral tube | 0+0 | 0+0 | 1+1 |
| Chaeta a2 on Abd. II | ciliate | smooth | ? |
| Chaetae m3 and m5 on Abd. II | not expanded | expanded | ? |
| Chaeta a2 on Abd. III | not expanded | expanded | ? |
| Chaeta im on Abd. III | ciliate | smooth | ? |
| Chaeta C1p on Abd. IV | ciliate | smooth | ? |
| Distribution | China | China | Vietnam |
?: character not provided in original description
Dental tubercles are pointed in Acrocyrtus and rounded in Ascocyrtus. However, it is sometimes uneasy to recognize the shape of dental tubercles. Their shape maybe wrongly observed in different visual angles, pointed in lateral view but “rounded” in facial view. They need to be observed under various angles. Another interesting point is that some species of
The present study was supported by the National Natural Sciences Foundation of China (31101622) and the Knowledge Innovation Program of the Chinese Academy of Sciences (No. KSCX2-EW-J-28). Thanks should be given to Louis Deharveng (Museum National d’Histoire Naturelle, Paris, France) and two anonymous referees for their useful suggestions.