





(C) 2013 Corey S. Whisson. This is an open access article distributed under the terms of the Creative Commons Attribution License 3.0 (CC-BY), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
For reference, use of the paginated PDF or printed version of this article is recommended.
Six species of Gastrocopta have been identified from the Pilbara region, Western Australia, by means of comparative analyses of shell and mtDNA variation. Three of these species, Gastrocopta hedleyi, Gastrocopta larapinta and Gastrocopta servilis, have been recorded in the Pilbara for the first time. Gastrocopta sp. CW1 is probably new to science and might be endemic to the region. By contrast, Gastrocopta hedleyi, Gastrocopta larapinta and Gastrocopta mussoni are shown to be widespread.
Australia, Pupilloidea, Pulmonata, 16S, COI
Gastrocopta Wollaston, 1878 is the most speciose pupillid genus in Australia with twelve recorded species (
Previous works have focussed mainly on the northern, eastern and southern parts of coastal Australia and to a lesser degree on the mid-west and central parts of Australia (
Since Pokryszko’s (
This paper (1) presents new data on Gastrocopta in the Pilbara, establishing new records and range extensions; (2) tests the taxonomic significance of morphological characters commonly used for the identification and delimitation of species by using a mitochondrial phylogeny; (3) provides comparative remarks on shell morphology of Gastrocopta species; (4) indicates systematic issues that require clarification by further studies. For detailed comparative analyses of shell characters we refer to
All Gastrocopta material from the Pilbara in the malacological collections of the Western Australian Museum was examined. Additional specimens from the private collection of Mr Vince Kessner and from the collection of the Field Museum of Natural History, Chicago were also included. In total 545 Gastrocopta lots were studied with distributional maps being plotted by use of the online vector map software available at www.planiglobe.com.
Species identifications were based on shell characters, with particular emphasise on the size, shape and quantity of apertural barriers. Specimens were photographed and measured using a Leica MZ16A microscope with Leica DFC500 camera. DNA was extracted from entire specimens taken from their shell by use of a QIAGEN DNA extraction kit for animal tissue following the standard procedure of the manual. Fragments of the mitochondrial 16S rRNA (16S) and of the COI genes were amplified by PCR using the primer pairs: 16Sar and 16Sbr (
Abbreviations used for depositories of material are: FMNH, Field Museum of Natural History, Chicago, United States; VK, Vince Kessner Private Collection, Adelaide River, Australia; WAM, Western Australian Museum, Perth, Australia. For shell aperture barrier terminology we followed
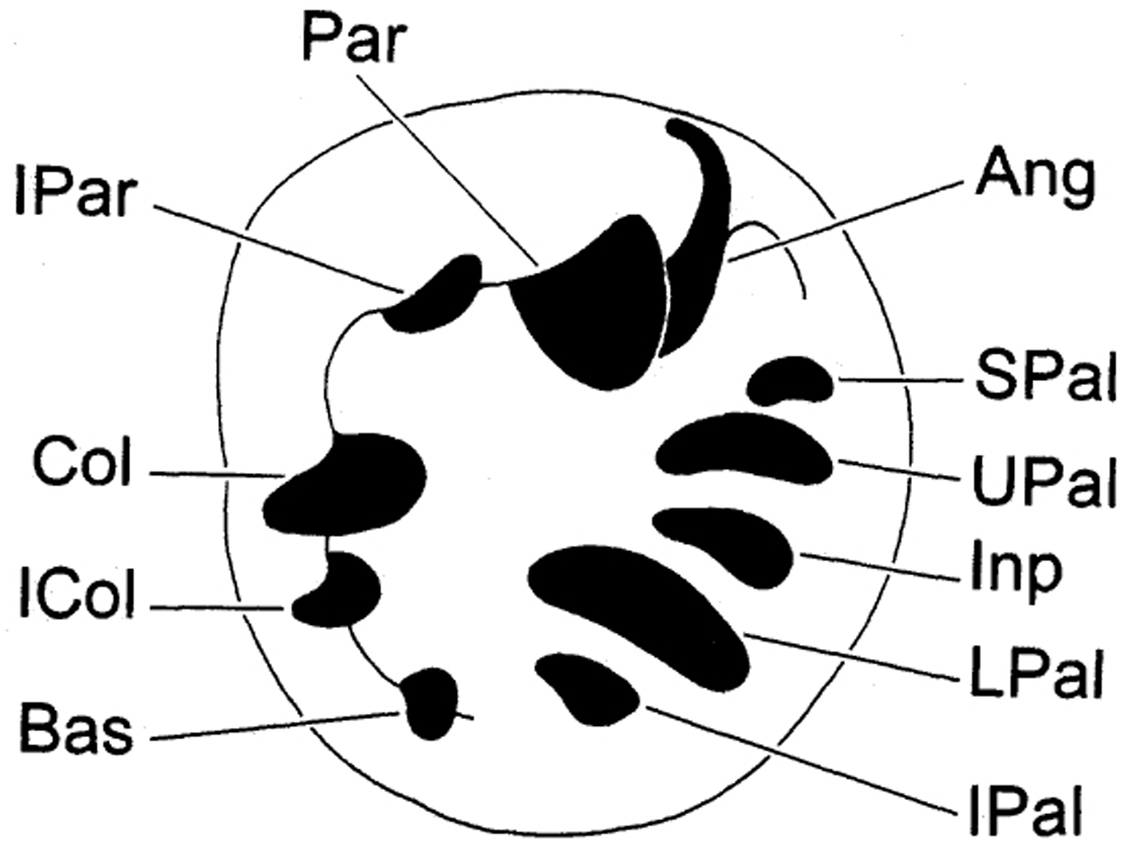
Apertural barriers of Gastrocopta (clockwise in aperture). IPAR Infraparietal Tooth PAR Parietoangular Tooth ANG Angular Tooth SPAL Suprapalatal Tooth UPAL Upper Palatal Tooth INP Interpalatal Tooth LPAL Lower Palatal Tooth IPAL Infrapalatal Tooth BAS Basal Tooth COL Columellar Tooth.
Six species of Gastrocopta were recorded from the Pilbara region (Table 1). Four species are endemic to Australia, one species is introduced and one species requires further investigation (Gastrocopta sp. CW1). Another species, Gastrocopta bannertonensis was only collected from the inner mid-west region of Western Australia and was not discussed in this paper.
Mean maximum shell height and width of Gastrocopta species from the Pilbara region.
| Species | N | Mean max. shell height (mm) | Mean max. shell width (mm) |
|---|---|---|---|
| Gastrocopta larapinta | 17 | 2.273 | 1.115 |
| Gastrocopta larapinta (Kalgan Pool) | 3 | 2.311 | 1.042 |
| Gastrocopta mussoni (ovate) | 7 | 2.278 | 1.251 |
| Gastrocopta mussoni (cylindrical) | 7 | 2.212 | 1.064 |
| Gastrocopta hedleyi | 14 | 2.017 | 1.014 |
| Gastrocopta margaretae (Pilbara) | 14 | 2.128 | 0.955 |
| Gastrocopta margaretae (SWA) | 13 | 2.466 | 1.128 |
| Gastrocopta sp. CW1 | 14 | 1.664 | 0.895 |
| Gastrocopta servilis | 14 | 2.255 | 1.070 |
http://species-id.net/wiki/Gastrocopta_hedleyi
Fig. 2BNarrabri, New South Wales.
Western Australia: Abydos (64km W of Marblebar): 21.1343°S, 119.1259°E (WAM S64439). Burrup Peninsula: 20.6080°S, 116.7670°E (WAM S60089); 20.6141°S, 116.7548°E (WAM S60226); 20.6066°S, 116.7681°E (WAM S60227); 20.5833°S, 116.8000°E (WAM S60228, WAM S60230, WAM S60349, WAM S61117-21); 20.6102°S, 116.7607°E (WAM S60353); 20.6232°S, 116.7784°E (WAM S60402); 20.6166°S, 116.7833°E (WAM S60475); 20.6119°S, 116.7587°E (WAM S60477); 20.6238°S, 116.7777°E (WAM S60480); 20.6300°S, 116.7800°E (WAM S65167); 20.5858°S, 116.8044°E (WAM S65168). Cloud Break area: 22.2997°S, 119.3737°E (WAM S60416). Hope Downs: 23.0865°S, 119.3184°E (WAM S42661); 23.0379°S, 119.2124°E (WAM S42663); 23.0952°S, 119.2022°E (WAM S59553); 23.1030°S, 119.2917°E (WAM S59555). Kalgan Pool area: 23.1872°S, 119.6958°E (WAM S58079); 23.1877°S, 119.6965°E (WAM S58091). Kangeenarina Gorge: 22.0588°S, 117.8549°E (WAM S60085). Karajini National Park: 22.4782°S, 118.5598°E (WAM S65307); 22.9797°S, 118.5891°E (WAM S65310); 22.8446°S, 118.5403°E (WAM S65314); 22.3714°S, 118.2989°E (WAM S65336). Marillana Station: 22.4285°S, 119.2043°E (WAM S81440). Mount Brockman area: 22.4815°S, 117.2384°E (WAM S83560). Mt Farquhar area: 22.4815°S, 116.8108°E (WAM S83564); 22.4932°S, 116.8679°E (WAM S83586). Nullagine area: 22.3848°S, 119.9696°E (WAM S58093); 22.3210°S, 119.4442°E (WAM S80958). Orebody 35°E(ca. 8km W of Newman): 23.4047°S, 119.6052°E (WAM S64713); 23.3943°S, 119.6316°E (WAM S64715-6, WAM S64718, WAM S64734, WAM S64753); 23.4108°S, 119.5715°E (WAM S64720); 23.3994°S, 119.5843°E (WAM S64722); 23.4045°S, 119.6211°E (WAM S64726); 23.4182°S, 119.5847°E (WAM S64730, WAM S64744); 23.4049°S, 119.6052°E (WAM S64732); 23.4045°S, 119.6247°E (WAM S64735); 23.4049°S, 119.6053°E (WAM S64737, WAM S64747); 23.4003°S, 119.6524°E (WAM S64740); 23.4137°S, 119.5826°E (WAM S64741); 23.4029°S, 119.6021°E (WAM S64742); 23.4003°S, 119.5721°E (WAM S64745); 23.3947°S, 119.5913°E (WAM S64750). ca. 35km E of Paraburdoo: 23.1300°S, 117.8984°E (WAM S41446); 23.1663°S, 117.9484°E (WAM S41447); 23.1670°S, 117.9597°E (WAM S41448). ca. 7.07.5km NW of Tom Price: 22.6500°S, 117.7185°E (WAM S42668); 22.6423°S, 117.7451°E (WAM S42672). Phil’s Creek: 22.7319°S, 119.1940°E (WAM S59376). Sulphur Springs: 21.1475°S, 119.2269°E (WAM S60229). Wonmunna: 23.1216°S, 119.0498°E (WAM S65971); 23.1428°S, 119.0099°E (WAM S65976); 23.1266°S, 119.0470°E (WAM S65992, WAM S81027, WAM S81085); 23.1255°S, 119.0797°E (WAM S80937); 23.1287°S, 119.0904°E (WAM S80938); 23.1393°S, 119.0182°E (WAM S80939); 23.1220°S, 119.0611°E (WAM S80941); 23.1436°S, 119.0064°E (WAM S81001, WAM S81073); 23.1185°S, 119.0649°E (WAM S81004); 23.1592°S, 118.9928°E (WAM S81025, WAM S81062); 23.1632°S, 118.9770°E (WAM S81033, WAM S81054, WAM S81168, WAM S81176); 23.1615°S, 119.0020°E (WAM S81048, WAM S811056); 23.1185°S, 119.0649°E (WAM S81091); 23.1596°S, 118.9703°E (WAM S81052); 23.1283°S, 119.0736°E (WAM S81059); 23.1331°S, 119.0154°E (WAM S81103); 23.1356°S, 119.0463°E (WAM S81120); 23.1546°S, 118.9932°E (WAM S81122); 23.1314°S, 119.0774°E (WAM S81174). ca. 6km W of Wodgina Mine: 21.2383°S, 118.6519°E (WAM S65895); 23.1348°S, 119.0338°E (WAM S81074).
This species has previously been recorded fromnorthern New South Wales and from scattered localities in northern Queensland (Cape York Peninsula), central Australia (Glen Helen area) and northern Western Australia (King Leopold Ranges) (
Gastrocopta hedleyi shellsare slightly smaller (shorter) than those of other Gastrocopta species (excluding Gastrocopta sp. CW1) recorded from the Pilbara. They typically have (1) a large, usually rounded (sometimes acute) columellar tooth that is drooping at the anterior end (2) a high, strongly convergent upper palatal tooth (3) a long, high, strongly twisted parietoangular tooth that usually comes in close proximity to the upper palatal tooth (4) a prominent infraparietal tooth that is sometimes prolonged as thin ridge on parietal wall (5) often a strong basal tooth (6) very occasionally with a weak interpalatal tooth.
Some Gastrocopta hedleyi shells (particularly more elongate specimens) can be difficult to separate from the ovate form of Gastrocopta mussoni but (1) are smaller (slender) when sympatric (2) have a less rounded body whorl (3) have a more strongly sigmoid lower palatal tooth (4) have a larger upper palatal tooth that is usually strongly convergent with the lower palatal (5) have a longer, more strongly twisted parietoangular tooth (6) have a larger, more rounded columellar tooth that is usually drooping at the anterior end .
The cylindrical form of Gastrocopta mussoni is also very similar to Gastrocopta hedleyi but (1) has a lower, shorter and less twisted parietoangular tooth (usually at 45o angle in apertural view) (2) has a shorter and less sigmoid (usually straight) lower palatal tooth (3) generally lacks an infraparietal tooth (4) has a smaller upper palatal tooth (occasionally slightly convergent with lower palatal) (5) has a more acutely angled, slanted columellar tooth, rarely drooping at the anterior end.
There is considerable variation in the shell size and barrier length of specimens identified as Gastrocopta hedleyi during this study. Many specimens grouped as Gastrocopta hedleyi from the eastern Hamersley Range (eg. Wonmunna, Kalgan Pool) have reduced barriers and often a lower parietoangular tooth (nearing 45o angle in apertural view) but a large series shows a progression to shells that typically possess a large, strongly convergent upper palatal tooth and a strongly twisted parietoangular tooth.
The abundance and seemingly allopatric nature of Gastrocopta hedleyi on the Burrup Peninsula is intriguing. The large numbers are presumably related to its’ habitat requirement of either Fig tree, Cypress or Brigalow Stands among rocky substrates, and its’ preference for high calcium soils (

Gastrocopta species: A Gastrocopta sp. CW1 (WAM S60408, Exmouth) B Gastrocopta hedleyi Pilsbry, 1917 (WAM S61117, Burrup Peninsula) C–E Gastrocopta larapinta (Tate, 1896) C WAM S58005, Kalgan Pool D–E WAM S60368, Roy Hill Station F Gastrocopta margaretae (Cox, 1868) (WAM S42834, Bateman Sanctuary); G Gastrocopta servilis (Gould, 1843) (WAM S60237, Karratha) H–J Gastrocopta mussoni Pilsbry, 1917 H Ovate Form (WAM S42865, Roy Hill Station) I Cylindrical Form (WAM S59375, Phils` Creek) J Cylindrical Form (WAM S61070), Roy Hill Station) . (Scale Bar = 1mm).
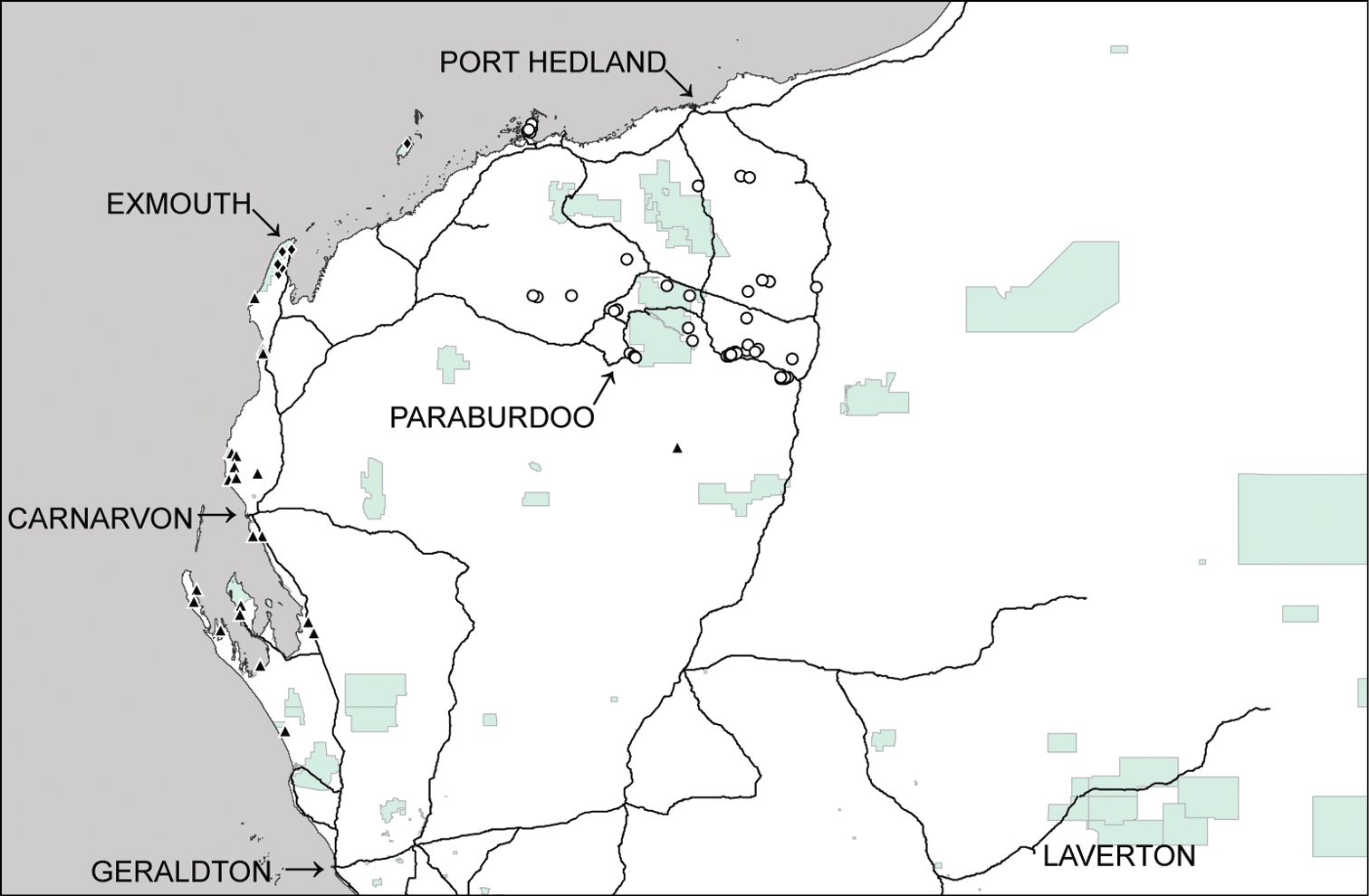
Distributional records of Gastrocopta hedleyi Pilsbry, 1917 (○), Gastrocopta margaretae (Cox, 1868) (▲) and Gastrocopta sp. CW1 (♦) in the Pilbara region. Shaded: Protected area.
http://species-id.net/wiki/Gastrocopta_larapinta
Fig. 2C–ECentral Australia.
Western Australia: Cane River: 22.0298°S, 115.4296°E (WAM S42993). Central Pilbara: 22.3855°S, 117.4667°E (WAM S58492, WAM S65790); 22.3200°S, 117.6194°E (WAM S65778); 22.2675°S, 117.7197°E (WAM S65780); 22.1350°S, 117.4728°E (WAM S58493); 21.0216°S, 117.0560°E (WAM S65817); 20.7506°S, 117.0096°E (WAM S65797). Christmas Creek: 22.4061°S, 119.7376°E (WAM S65612); 22.3985°S, 119.7930°E (WAM S65604). Cloud Break area: 22.3210°S, 119.4442°E (WAM S34453, WAM S80956); 22.2688°S, 119.3147°E (WAM S61128); 22.3652°S, 119.3409°E (WAM S65150); 22.3985°S, 119.4748°E (WAM S65156); 22.3949°S, 119.5019°E (WAM 65161); 22.3527°S, 119.4189°E (WAM S65135). Collier Rocks: 20.4071°S, 116.8514°E (VK 30297). Cy Creek: 22.8183°S, 114.0609°E (WAM S34381). Du Boulay Creek: 21.1833°S, 116.1833°E (WAM S34569). Fortescue Marsh area: 22.2930°S, 119.0606°E (WAM S61996-7); 22.4646°S, 119.7726°E (WAM S64651, WAM S64654); 22.4430°S, 119.7785°E (WAM S64649); 22.4252°S, 119.7235°E (WAM S64635); 22.3125°S, 119.2348°E (WAM S64634, WAM S64637); 22.2875°S, 119.1701°E (WAM S64650); 22.2822°S, 119.1276°E (WAM S42923, WAM S64644, WAM S64652, WAM S64694). Hope Downs: 23.1030°S, 119.5821°E (WAM S59298); 23.0919°S, 119.1875°E (WAM S59550). Jimblebar: 23.3693°S, 120.1958°E (WAM S41346). Kalgan Pool area: 23.1877°S, 119.6965°E (WAM S58005); 23.1874°S, 119.6957°E (WAM S80939). Koodaideri Corridor West (90.4km NW of Tom Price): 21.8833°S, 117.7000°E (WAM S83434). Marillana Station: 22.6260°S, 119.2834°E (WAM S34638); 22.5840°S, 119.3114°E (WAM S34632, WAM S34472); 22.5739°S, 119.2562°E (WAM S34474); 22.5666°S, 119.2327°E (WAM S34637); 22.5663°S, 119.2308°E (WAM S34633); 22.6013°S, 119.2913°E (WAM S80924); 22.5625°S, 119.2311°E (WAM S80908); 22.1269°S, 119.2123°E (WAM S80911); 22.4080°S, 119.0067°E (WAM S80914); 22.4285°S, 119.2043°E (WAM S8143940); 22.3476°S, 119.1518°E (WAM S81433); 22.4295°S, 119.1923°E (WAM S81446). Millstream National Park: 21.2000°S, 117.2667°E (WAM S60343); 21.6000°S, 117.1000°E (WAM S 61044); 21.5833°S, 117.1000°E (WAM S 61048); 21.5833°S, 117.0833°E (WAM S60944-5); 21.5833°S, 117.0667°E (WAM S 60947); 21.4255°S, 117.0535°E (WAM S81213); 21.2039°S, 117.0440°E (WAM S81267). ca. 30km NNE of Newman: 23.1164°S, 119.8865°E (WAM S64469). ca. 65km NW of Newman: 22.9169°S, 119.2128°E (WAM S80937). ca. 108118km N of Newman: 22.3132°S, 119.8599°E (WAM S65534, WAM S65673); 22.3134°S, 119.7886°E (WAM S65646); 22.2972°S, 119.8633°E (WAM S65530, WAM S65682); 22.2954°S, 119.8109°E (WAM S65652). North Star Mine: 21.2284°S, 119.0386°E (WAM S65720); 21.2104°S, 118.8769°E (WAM S65723). Phil’s Creek: 22.7412°S, 119.1959°E (WAM S59388, WAM S80934); 22.7320°S, 119.1836°E (WAM S59374). ca. 100km S of Port Hedland: 20.6066°S, 119.5016°E (WAM S80942). ca. 200km SSE of Port Hedland: 22.1554°S, 119.0216°E (WAM S83486). Robe River area: 21.8063°S, 116.0774°E (WAM S42832). 6km SW of Redmont Airport: 22.0195°S, 118.9816°E (WAM S83423). NNE of Rocklea Homestead: 22.7882°S, 117.4974°E (WAM S80977). Roy Hill Station: 22.4898°S, 119.8951°E (WAM S34703); 22.4547°S, 119.8709°E (WAM S42924, WAM S60428-9, WAM S65389); 22.4943°S, 119.9217°E (WAM S60359, WAM S60368, WAM S60373, WAM S60378); 22.5383°S, 119.9424°E (WAM S42831, WAM S42833, WAM S60232, WAM S60857, WAM S60861); 22.4793°S, 119.9420°E (WAM S42925, WAM S60363, WAM S60426, WAM S60853, WAM S60860); 22.6394°S, 119.9642°E (WAM S60398); 22.5769°S, 119.9952°E (WAM S603667, WAM S608545, WAM S60859); 22.5771°S, 120.0247°E (WAM S60422); 22.7058°S, 119.7082°E (WAM S60392); 22.6430°S, 119.9599°E (WAM S60396); 22.6076°S, 119.9826°E (WAM S60388); 22.6593°S, 119.9198°E (WAM S60397); 22.6642°S, 119.9458°E (WAM S61066, WAM S61069); 22.6431°S, 119.9642°E (WAM S61075, WAM S61077); 22.6225°S, 119.9634°E (WAM S610723); 22.5050°S, 119.9143°E (WAM S64455). Running Waters (east of Nullagine): 21.6819°S, 121.1254°E (WAM S58039, WAM S58050); 21.6806°S, 121.1261°E (WAM S58032). 15km W of Shaw River Airport: 21.6123°S, 119.2642°E (WAM S83412). Wonmunna: 23.1220°S, 119.0611°E (WAM S81087, WAM S81182); 23.1356°S, 119.0463°E (WAM S81107, WAM S81125, WAM S81164); 23.1216°S, 119.0498°E (WAM S81096); 23.1201°S, 119.0484°E (WAM S65993, WAM S80904). ca. 18-23km SE of Wodgina Mine: 21.2273°S, 118.8336°E (WAM S646089); 21.2871°S, 118.8671°E (WAM S64604, WAM S64616). ca. 20km NNE of Wodgina Mine: 21.0260°S, 118.7024°E (WAM S64610). Yule River area: 21.6961°S, 118.8604°E (WAM S83372).
This species has previously been recorded fromcentral Australia (southern part of Northern Territory) with fewer records in north-western Queensland (Gregory River Basin); eastern coast of Queensland and a single record from the Oscar Ranges, in the southern Kimberley region of Western Australia (
The shells of typical Gastrocopta larapinta specimens are distinguished from most other Gastrocopta species in the Pilbara by (1) their large size (2) usually the presence of three solid palatal teeth (the interpalatal tooth varying from a tiny callus to large tooth) (3) a short, solid parietoangular tooth that is usually deflected or curved moderately toward the columellar wall so that its anterior end is somewhat vertical in the apertural view (4) a long angular tooth that is generally fused (or connected via a translucent callus) to the parietoangular tooth, occasionally separate (particularly those that lack or have a small interpalatal tooth) (5) smaller, more rounded columellar tooth that curves or angles abruptly toward the columellar wall (6) usually the presence of an infraparietal tooth or basal tooth (or both).
Typical Gastrocopta larapinta shells with a small interpalatal tooth (or very occasionally no interpalatal tooth) appear considerably more variable in apertural barrier structure (particularly in high calcareous soils), making their separation from the cylindrical and elongate-ovate forms of Gastrocopta mussoni difficult. As such the following separation is tentative. Gastrocopta larapinta shells are typically (1) slightly to moderately larger (obese) (2) have a slightly smaller, more rounded columellar tooth (3) usually a much shorter parietoangular tooth that is positioned lower in apertural view (4) generally possesses an infraparietal tooth (5) often a slightly lower, less convergent upper palatal tooth (6) usually a less sigmoid lower palatal tooth.
The separation of Gastrocopta larapinta (small or no interpalatal tooth) with the cylindrical and elongate-ovate forms of Gastrocopta mussoni has proved extremely difficult and a more detailed molecular study is required to resolve this issue.
Some of those near west coast specimens (Cy Creek) included as cylindrical Gastrocopta mussoni contained mixed lots of Gastrocopta larapinta and Gastrocopta mussoni. Interestingly, Poykryszko had identified a few of these larger Cy Creek specimens as Gastrocopta larapinta, but included those records as Gastrocopta mussoni in her publication.
It is possible
As there is doubt surrounding the distinguishing morphological characters of cylindrical Gastrocopta mussoni and Gastrocopta larapinta shells with a small or no interpalatal tooth, the above separation is tentative and a more detailed genetic investigation is required.
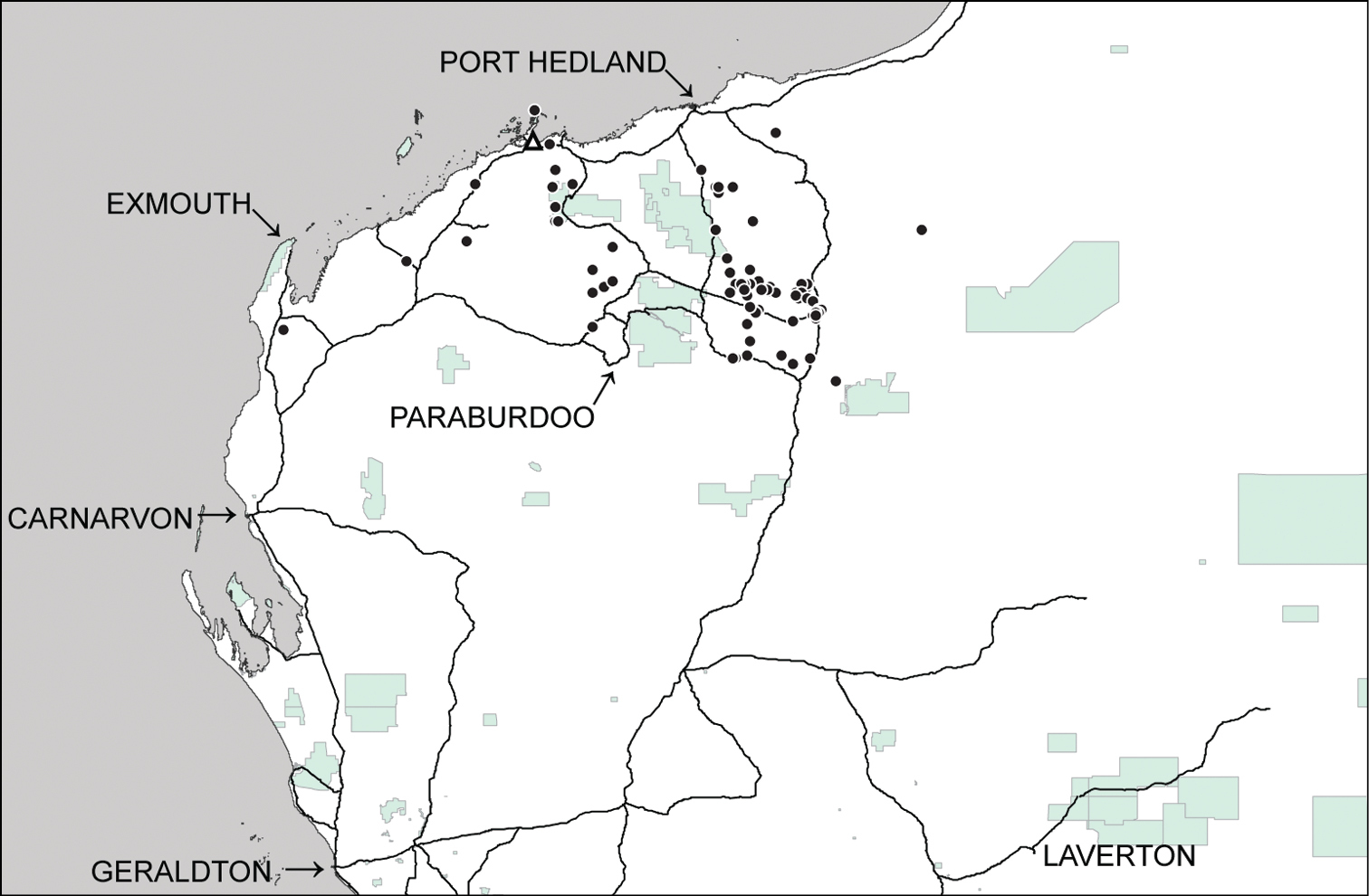
Distributional records of Gastrocopta larapinta (Tate, 1896) (●) and Gastrocopta servilis (Gould, 1843) (Δ) in the Pilbara region. Shaded: Protected area.
http://species-id.net/wiki/Gastrocopta_margaretae
Fig. 2FWallaroo, South Australia.
Western Australia: Bateman Sanctuary: 23.0552°S, 113.8234°E (WAM S42834). ~18km N of Boolathana Homestead: 24.4133°S, 113.7445°E (WAM S64708). Boolathana Station: 24.4127°S, 113.7631°E (WAM S64709). Bush Bay: 25.1175°S, 113.8063°E (WAM S42806); 25.1316°S, 113.7681°E (WAM S60355); 25.1136°S, 113.7311°E (WAM S64575); 25.1175°S, 113.8063°E (WAM S64577). Carrarang Station: 26.1666°S, 113.3500°E (WAM S34378). Cy Creek: 23.1000°S, 113.8000°E (WAM S34380). Dirk Hartog Island: 25.7166°S, 113.0667°E (WAM S14439); 25.8333°S, 113.0500°E (WAM S34398). Francois Peron National Park: 25.9760°S, 113.5707°E (WAM S60269); 25.9758°S, 113.5706°E (WAM S64706); 25.8752°S, 113.5497°E (WAM S61127). Lake Macleod area: 24.3449°S, 113.5194°E (WAM S65084); 24.3668°S, 113.5145°E (WAM S65093); 24.3544°S, 113.5098°E (WAM S65102); 24.4760°S, 113.5257°E (WAM S65108); 24.4598°S, 113.5013°E (WAM S65110); 24.3544°S, 113.5098°E (WAM S65121). 0.25 miles W of Nichol Springs: 24.1333°S, 118.4167°E (WAM S60270). ~25 miles N of turn off to Shark Bay on NW Coastal Highway: 26.0647°S, 114.3353°E (WAM S34459, WAM S60271, WAM S64585). 0.5 miles W of 512 mile peg on NW Coastal Highway: 26.1966°S, 114.3758°E (WAM S34379). Quobba Station: 24.4758°S, 113.4166°E (WAM S42829); 24.2448°S, 113.5353°E (WAM S61125); 24.1927°S, 113.4548°E (WAM S64576); 24.2233°S, 113.5036°E (WAM S64707). Salutation Island: 26.5333°S, 113.7667°E (WAM S34377). Winderabandi Point: 22.4929°S, 113.7258°E (WAM S60474). Zuytdorp: 27.2636°S, 114.0703°E (WAM S64710).
This species has previously been recorded fromthe western and southern coastal areas of Western Australia, the southern regions of South Australia and the area near Alice Springs in the Northern Territory. There is also an isolated record from the King Leopold Range in the north of Western Australia (
Shells of Gastrocopta margaretae are easily distinguished from other Gastrocopta species in the Pilbara by the presence of (1) a moderately to strongly folded columellar tooth (2) a generally large and transverse basal tooth (3) a high and long lower palatal tooth (4) an upper palatal tooth that is moderately to strongly convergent with the lower palatal (5) a weak to strong infraparietal tooth present.
The few specimens sequenced from the south coast of Western Australia (WAM S32048, WAM S32052) could represent genetic isolation by distance or perhaps a different species from those on the west coast (WAM S42834) but more molecular data are required. The southern specimens are (1) much larger with reduced apertural barriers (2) more strongly rounded whorls (conical) and (3) consistently lack an infraparietal tooth. Specimens resembling the smaller west coast form (ie. long apertural barriers and weak to strong infraparietal tooth) have also been recorded from the south west area of Western Australia (Whisson, pers. comm.) where it is often sympatric with Gastrocopta bannertonensis (Gabriel, 1930). It is not known whether there is a continuous distribution between the two areas. Until more detailed molecular work is undertaken we have maintained Pokryszko’s (
http://species-id.net/wiki/Gastrocopta_mussoni
Fig. 2H–JCalliungal (=Mt Morgan), Queensland.
Western Australia: Angelo River: 23.4331°S, 118.7329°E (WAM S65935). Anketell Point area: 20.6356°S, 117.0398°E (WAM S59990); 20.6719°S, 117.0965°E (WAM S599912); 20.7025°S, 117.0473°E (WAM S80936). Area C: 22.9104°S, 118.9664°E (WAM S60417). Barrow Island: 20.7833°S, 115.4000°E°E (WAM S34384); 20.8649°S, 115.4069°E°E (WAM S34455); 20.6666°S, 115.4667°E (WAM S42879, WAM S60847); 20.7921°S, 115.4573°E (WAM S59636); 20.7858°S, 115.4573°E (WAM S59637); 20.7997°S, 115.4403°E (WAM S59640); 20.7069°S, 115.4194°E (WAM S59642); 20.7866°S, 115.4547°E (WAM S59644, WAM S60413); 20.7938°S, 115.4575°E (WAM S59651); 20.8644°S, 115.3428°E (WAM S59652); 20.8101°S, 115.4270°E (WAM S60410); 20.6666°S, 115.4667°E (WAM S42879, WAM S60847); 20.7977°S, 115.4064°E (WAM S65123, WAM S65126, WAM S65127, WAM S65164); 20.7684°S, 115.4673°E (WAM S65174). Cane River Conservation Park: 22.1694°S, 115.5606°E (WAM S42961); 22.4321°S, 115.2895°E (WAM S42967); 22.2685°S, 115.6470°E (WAM S42974); 22.0975°S, 115.4942°E (WAM S42976); 22.2075°S, 115.5260°E (WAM S42981); 22.1451°S, 115.7249°E (WAM S42987); 22.0298°S, 115.4296°E (WAM S42992); 22.0131°S, 115.6325°E (WAM S42998). Cape Preston area: 20.8435°S, 116.2016°E (WAM S59141). Chichester Ranges: 22.0525°S, 118.9883°E (WAM S60407); 22.0516°S, 118.9884°E (WAM S42711, WAM S42713, WAM S42759); 22.1508°S, 119.0179°E (WAM S42709); 22.0503°S, 118.9934°E (WAM S42717); 23.1164°S, 119.8865°E (WAM S64460). Christmas Creek area: 22.4170°S, 119.8941°E (WAM S65603); 22.4078°S, 119.8767°E (WAM S65605); 22.4061°S, 119.7376°E (WAM S65611). Cloud Break area: 20.3216°S, 119.4418°E (WAM S34460); 22.3210°S, 119.4442°E (WAM S42876, WAM S60403, WAM S80957); 22.3181°S, 119.3788°E (WAM S60267); 22.2935°S, 119.3872°E (WAM S61122); 22.2997°S, 119.3737°E (WAM S61123); 22.2881°S, 119.2360°E (WAM S65137°S, WAM S65143°S, WAM S65153); 22.3251°S, 119.4458°E (WAM S65139); 22.3527°S, 119.4189°E (WAM S65144); 22.3652°S, 119.3409°E (WAM S65148); 22.3894°S, 119.4380°E (WAM S65155); 22.3985°S, 119.4748°E (WAM S65157); 22.3949°S, 119.5019°E (WAM S65213). Cy Creek: 22.8183°S, 114.0609°E (WAM S34382). Dolphin Island: 20.4833°S, 116.8500°E (WAM S34385). Du Boulay Creek: 21.1833°S, 116.1833°E (WAM S34568). Finucane Island: 20.2982°S, 118.5572°E (WAM S16108). Fortescue Marsh area: 22.2822°S, 119.1276°E (WAM S42926, WAM S64694, WAM S646523); 22.2938°S, 119.0732°E (WAM S61991); 22.5098°S, 119.1274°E (WAM S61993°S, (WAM S61995); 22.2872°S, 119.0301°E (WAM S646401, WAM S646556); 22.1322°S, 119.1983°E (WAM S64684); 22.2924°S, 119.0279°E (WAM S64687). East Hamersley Range: 22.8586°S, 119.6728°E (WAM S42921); 22.6335°S, 119.3289°E (WAM S64470). Hope Downs: 23.1474°S, 119.5191°E (WAM S42662); 23.1030°S, 119.2917°E (WAM S59548, WAM S59554); 23.0925°S, 119.2058°E (WAM S59551); 23.0878°S, 119.1609°E (WAM S59552). Jinayri area: 22.9219°S, 119.2036°E (WAM S42929); 23.0530°S, 119.2701°E (WAM S59239, WAM S592412, WAM S59244); 23.0129°S, 119.2371°E (WAM S59240, WAM S59243); 22.9058°S, 119.2000°E (WAM S59576, WAM S59596); 22.9275°S, 119.1275°E (WAM S59585); 22.9169°S, 119.2128°E (WAM S59587); 22.9219°S, 119.2036°E (WAM S59591); 23.0504°S, 119.2731°E (WAM S60406). Kalgan Pool area: 23.1877°S, 119.6965°E (WAM S439823). Kangeenarina Gorge area: 22.1186°S, 117.9427°E (WAM S61763); 21.8489°S, 117.3837°E (WAM S81210). Karratha area: 20.7166°S, 116.8500°E (WAM S60412); 21.4036°S, 116.9392°E (WAM S81217). SE of Karratha: 21.0698°S, 116.9702°E (WAM S81241, WAM S81246); 21.0300°S, 116.9997°E (WAM S81252, WAM S81271); 20.9853°S, 116.8811°E (WAM S81268). Lake Macleod: 24.3544°S, 113.5098°E (WAM S65120). Marillana Station: 22.5840°S, 119.3114°E (WAM S84075); 22.5663°S, 119.2308°E (WAM S34473); 22.5497°S, 119.2147°E (WAM S34476); 22.5851°S, 119.3142°E (WAM S34478). 22.5739°S, 119.2562°E (WAM S34634, WAM S80940); 22.5538°S, 119.2283°E (WAM S34635); 22.6260°S, 119.2834°E (WAM S34639); 22.5625°S, 119.2311°E (WAM S80907); 22.1269°S, 119.2123°E (WAM S80910); 22.5625°S, 119.2311°E (WAM S80913); 22.4080°S, 119.0067°E (WAM S80916); 22.6376°S, 119.3744°E (WAM S80917); 22.4285°S, 119.2043°E (WAM S81437); 22.3182°S, 119.1175°E (WAM S81442); 22.4295°S, 119.1923°E (WAM S81444). Meentheena Outcamp: 21.2671°S, 120.4570°E (WAM S58055); 21.2815°S, 120.4511°E (WAM S58099); 21.2816°S, 120.4508°E (WAM S58098). Millstream National Park: 21.6000°S, 117.1000°E (WAM S42838, WAM S42931, WAM S61043); 21.5833°S, 117.0833°E (WAM S42839, WAM S60943); 21.5833°S, 117.0667°E (WAM S60946); 21.4255°S, 117.0535°E (WAM S81212); 21.1781°S, 117.0461°E (WAM S81250); 21.2039°S, 117.0440°E (WAM S81301, WAM S81296). Mount Brockman area: 22.4815°S, 117.2384°E (WAM S83561). Mt Farquhar area: 22.4815°S, 116.8108°E (WAM S83563). Muiron Island: 21.6666°S, 114.3333°E (WAM S34383). Murray Hills: 22.1147°S, 118.5221°E (WAM S59996). ~60km NW of Newman: 23.0530°S, 119.2701°E (WAM S60235, WAM S60266, WAM S60405); 23.0878°S, 119.1609°E (WAM S59549); 23.0504°S, 119.2731°E (WAM S60404, WAM S60411); 22.9632°S, 119.2276°E (WAM S42928, WAM S60233). ~112km NNE of Newman: 22.3621°S, 119.9691°E (WAM S65650); 22.3132°S, 119.8599°E (WAM S 65672); 22.3871°S, 119.9664°E (WAM S 65694). ~110km N of Newman: 22.2954°S, 119.8109°E (WAM S65638); 22.3132°S, 119.8599°E (WAM S65672); 22.2972°S, 119.8633°E (WAM S65683). ~70km S of Newman: 23.7270°S, 119.7242°E (WAM S58073). North Star: 21.2523°S, 118.8334°E (WAM S65699); 21.1971°S, 118.8286°E (WAM S65706, WAM S65710); 21.2681°S, 118.9682°E (WAM S65713); 21.2104°S, 118.8769°E (WAM S65718); 21.2104°S, 118.8769°E (WAM S65719); 21.2319°S, 118.8263°E (WAM S65728). Nullagine area: 21.8221°S, 120.3409°E (WAM S61802). Orebody 35°E (ca. 8km W of Newman): 23.4108°S, 119.5715°E (WAM S64733); 23.3837°S, 119.6478°E (WAM S64748); 23.3712°S, 119.6127°E (WAM S64749); 23.3819°S, 119.6133°E (WAM S64751); 23.3970°S, 119.6138°E (WAM S64752). West of Pannawonica: 21.7000°S, 116.1667°E (WAM S42805); 21.8063°S, 116.0774°E (WAM S60268); 21.7202°S, 116.0705°E (WAM S60414, WAM S61034); 21.6298°S, 116.0206°E (WAM S60415). Point Quobba, near lighthouse: 24.4797°S, 113.4178°E (FMNH 201611); Phils` Creek area: 22.7320°S, 119.1836°E (WAM S 42878, WAM S 59372); 22.7316°S, 119.1931°E (WAM S 59371, WAM S59375, WAM S59383); 22.7351°S, 119.1856°E (WAM S59373); 22.7384°S, 119.1916°E (WAM S59377); 22.7412°S, 119.1959°E (WAM S59378); 22.7448°S, 119.1927°E (WAM S59379); 22.7412°S, 119.1959°E (WAM S59380); 22.7366°S, 119.1811°E (WAM S59381); 22.7316°S, 119.1798°E (WAM S59382). ~40km S of Port Hedland: 20.6095°S, 118.6661°E (WAM S81404). NNE of Rocklea Homestead: 22.8101°S, 117.4734°E (WAM S80990). Roy Hill Station: 22.6642°S, 119.9458°E (WAM S34588); 22.4943°S, 119.9217°E (WAM S42828, WAM SS60376); 22.4396°S, 119.9453°E (WAM S42837); 22.4898°S, 119.8951°E (WAM S42875, WAM S60234); 22.5383°S, 119.9424°E (WAM S42927); 22.7058°S, 119.7082°E (WAM S60393); 22.6347°S, 119.9698°E (WAM S60394); 22.8174°S, 119.9473°E (WAM S60395, WAM S60399); 22.5771°S, 120.0247°E (WAM S60421, WAM S60866); 22.4793°S, 119.9421°E (WAM S60427); 22.5039°S, 120.0210°E (WAM S60864); 22.5566°S, 119.9700°E (WAM S60865); 22.7489°S, 119.9221°E (WAM S61067); 22.7195°S, 119.9395°E (WAM S61068, WAM S61071); 22.6365°S, 119.9639°E (WAM S61070); 22.6431°S, 119.9642°E (WAM S61074, WAM S61076); 22.5050°S, 119.9042°E (WAM S64448); 22.5843°S, 120.0172°E (WAM S64453).Running Waters (east of Nullagine): 21.6819°S, 121.1254°E (WAM S58050); 21.6815°S, 121.1270°E (WAM S58059). W end of Telfer Road: 21.3290°S, 121.1390°E (WAM S58044). NW of Tom Price: 22.3734°S, 117.4631°E (WAM S34566); 22.1350°S, 117.4728°E (WAM S65775); 22.1519°S, 117.5428°E (WAM S65781); 22.2997°S, 117.6378°E (WAM S65787); 22.3855°S, 117.4667°E (WAM S65788). Weeli Wolli Creek: 22.6166°S, 119.4000°E (WAM S60231). Near Wodgina Mine: 21.1831°S, 118.6569°E (WAM S34567, WAM S65843, WAM S65869); 21.2871°S, 118.8671°E (WAM S64618); 21.1789°S, 118.6463°E (WAM S65841, WAM S65855); 21.1737°S, 118.6503°E (WAM S65844); 21.2383°S, 118.6519°E (WAM S65903). Wonmunna: 23.1436°S, 119.0064°E (WAM S65977, WAM S81002, WAM S81026); 23.1355°S, 119.0384°E (WAM S65983); 23.1596°S, 118.9703°E (WAM S65979); 23.1632°S, 118.9770°E (WAM S65985); 23.1615°S, 119.0020°E (WAM S 65990); 23.1201°S, 119.0484°E (WAM S65993); 23.1287°S, 119.0904°E (WAM S 65996); 23.1283°S, 119.0736°E (WAM S65998); 23.1268°S, 119.0673°E (WAM S81032); 23.1309°S, 119.0689°E (WAM S81036, WAM S81167); 23.1428°S, 119.0099°E (WAM S81053); 23.1355°S, 119.0384°E (WAM S81081); 23.1169°S, 119.0396°E (WAM S81099); 23.1255°S, 119.0797°E (WAM S81094); 23.1216°S, 119.0498°E (WAM S81096, WAM S81171); 23.1393°S, 119.0182°E (WAM S81104); 23.1210°S, 119.0632°E (WAM S81109); 23.1356°S, 119.0463°E (WAM S 81110, WAM S81127); 23.1185°S, 119.0649°E (WAM S81114); 23.1255°S, 119.0797°E (WAM S81116); 23.1201°S, 119.0484°E (WAM S81119); 23.1331°S, 119.0154°E (WAM S81170); 23.1216°S, 119.0498°E (WAM S81178); 23.1220°S, 119.0611°E (WAM S81180). Yandi Mine: 22.8200°S, 119.2500°E (WAM S61774).
This species has previously been recorded from central Australia (the southern part of Northern Territory), with a few records from the mid-west coast and northern Western Australia; northern Northern Territory; northern and north-eastern parts of Queensland and South Australia (
Cylindrical and elongate-ovate forms of Gastrocopta mussoni can be mistaken for Gastrocopta larapinta specimens (with a small or absent interpalatal tooth) but (1) are slightly to moderately slender (2) have a higher, usually longer parietoangular tooth (3) have a larger, more strongly slanted and acutely angled columellar tooth, with its posterior edge often forming a prominent wide ridge along the columellar wall (4) quite frequently have an upper palatal tooth that is slightly (occasionally moderately) convergent with the lower palatal (see also earlier section on Gastrocopta larapinta) . Gastrocopta mussoni very occasionally possesses a small interpalatal tooth, usually located close to the upper palatal.
The typical ovate form of Gastrocopta mussoni can be confused with Gastrocopta hedleyi (particularly those with reduced apertural barriers) but (1) are larger (obese) when sympatric (2) have a less sigmoid lower palatal tooth and (3) have a smaller, less convergent upper palatal tooth (see also earlier section on Gastrocopta hedleyi).
There appears to be two size forms in Gastrocopta mussoni, the larger ovate form and smaller, slender cylindrical form, and in agreement with
The ovate form of Gastrocopta mussoni appears to be most common in the Pilbara. Prior to Pokryszko’s
Some of those specimens tentatively identified as the elongate-ovate form of Gastrocoptamussoni during this study (Wonmunna; Cy Creek; Cloud Break, Barrow Island) are (1) much larger (obese) than the usual elongate-ovate form (2) have the parietoanangular tooth lower (45o) (3) usually have a supraparietal tooth and (4) quite frequently possess a small interpalatal tooth. These specimens may prove to be the somewhat variable Gastrocoptalarapinta with a small or no interpalatal tooth, but in the absence of a larger series of specimens and more detailed molecular data, we have left them as Gastrocopta mussoni.
The nature of many cylindrical Gastrocopta mussoni identified by Poykrosko (1996) and during this study requires more work. It is probable we have lumped the slender form of Gastrocopta larapinta from Kalgan Pool (no interpalatal tooth) withcylindrical Gastrocopta mussoni.
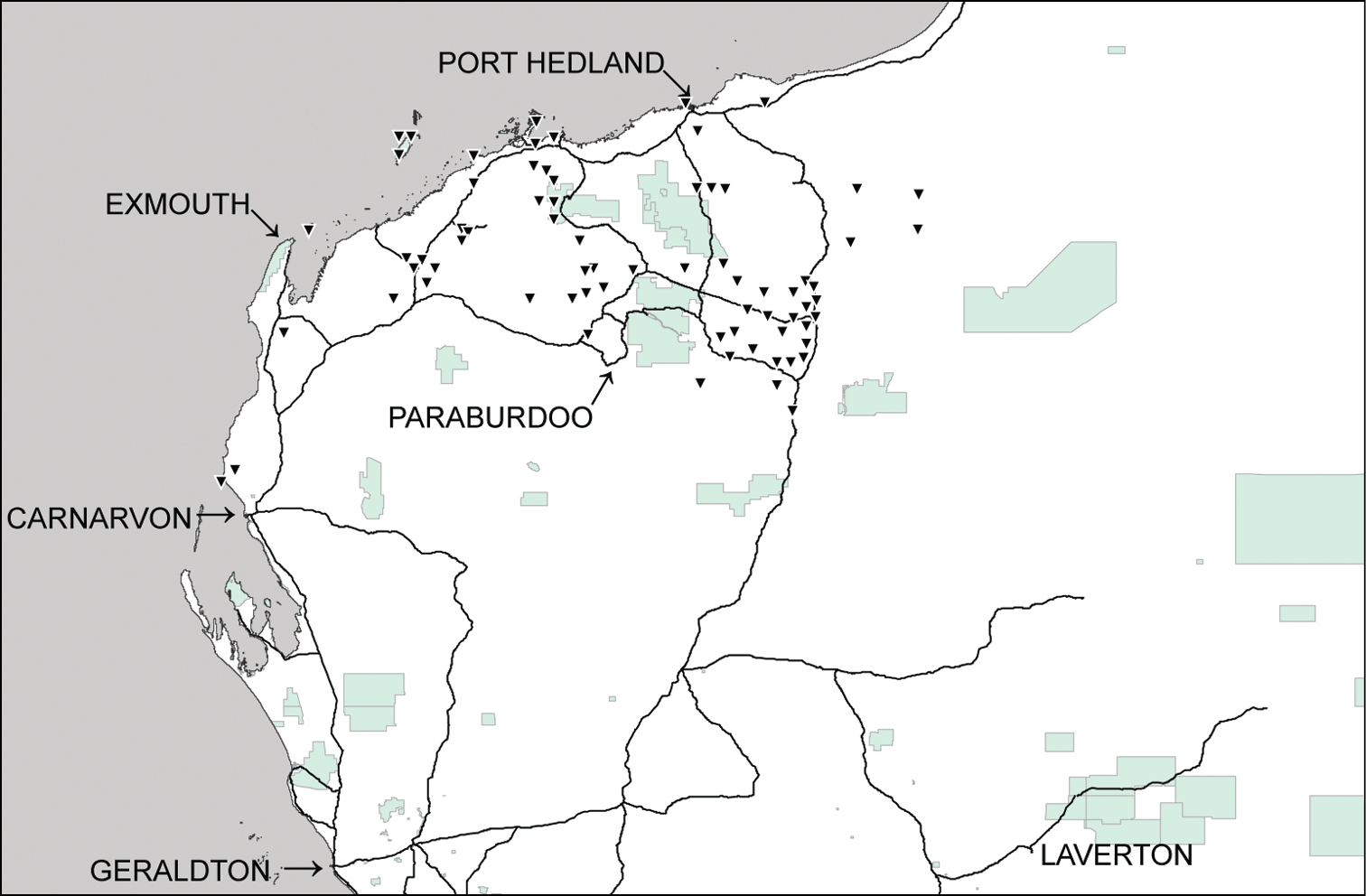
Distributional records of Gastrocopta mussoni Pilsbry, 1917 (▼) in the Pilbara region. Shaded: Protected area.
Western Australia: Barrow Island: 20.7069°S, 115.4194°E (WAM S59641). Cape Range No. 2 Deep Well: 21.9500°S, 114.0333°E (WAM S14132). Cape Range (cave): 22.1166°S, 113.9833°E (WAM S34394); 22.1500°S, 114.0000°E (WAM S34395, WAM S60409, WAM S60831); 22.0833°S, 113.9833°E (WAM S34396); 22.1833°S, 113.9833°E (WAM S80955). Exmouth rubbish tip: 21.9166°S, 114.1167°E (WAM S60408)
This species is recorded from the Cape Range and from an isolated site on Barrow Island (Figure 3).
Shells of Gastrocopta sp. CW1 are easily recognized by their (1) small size (2) very solid, non-lamellate columellar tooth that projects horizontally from the columellar wall (shelf-like), slightly drooping at anterior end (3) long sigmoid lower palatal tooth (4) large, transverse upper palatal tooth (5) presence of a suprapalatal tooth.
Gastrocopta sp. CW1. is very similar to Gastrocopta stupefaciens and Gastrocopta recondita but (1) is smaller (2) has longer apertural barriers and (3) has a thick, solid, non-lamellate columellar tooth and is here within regarded as a new species. Some of Solems` Gastrocopta recondita specimens from limestone outcrops near Katherine (station WA-685) and Lake Argyle (station WA-248) have a similar columellar tooth structure and their relationship to Gastrocopta sp. CW1 needs further work.
http://species-id.net/wiki/Gastrocopta_servilis
Fig. 2Gnear Matanzas, Cuba.
Karratha area: 20.7385°S, 116.8357°E (WAM S9932, WAM S60237).
This species has previously been recorded from just north of Broome (Quondong Point) across northern Australia to mid-eastern Queensland and offshore islands (
The shells of Gastrocopta servilis are easily distinguished from other Pilbara Gastrocopta by their (1) strongly rounded whorls (2) short, straight columellar tooth which is perpendicular to the mid-columellar wall (3) very long angular tooth which is fused with the parietoangular tooth (4) weak to absent basal tooth and (5) weak to absent upper palatal tooth.
Gastrocopta servilis has been a recent introduction to the residential gardens of Karratha.
Two mitochondrial gene fragments, COI and 16S, have been analysed. The data sets contained 27 sequences of Western Australian Gastrocopta (five each of Gastrocopta bannertonensis and Gastrocopta mussoni, 12 of Gastrocopta larapinta, two or three, respectively, of Gastrocopta margaretae, and two of Gastrocopta hedleyi) as well as 16 Genbank sequences of several American Gastrocopta species that stem from the study of
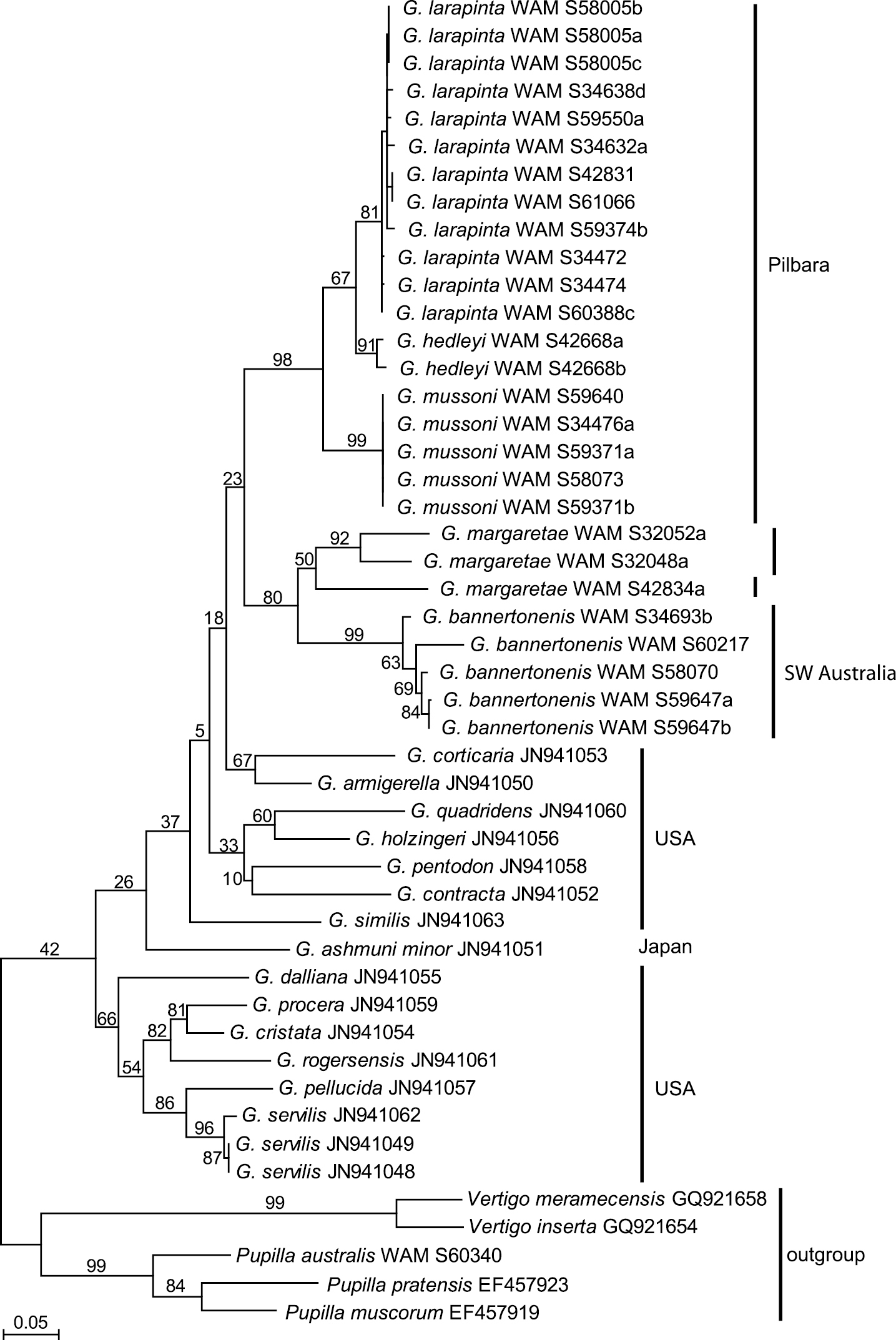
Maximum Likelihood phylogram for COI based on analysis of 27 new sequences of Gastrocopta from Western Australia and the 16 Genbank sequences made available by
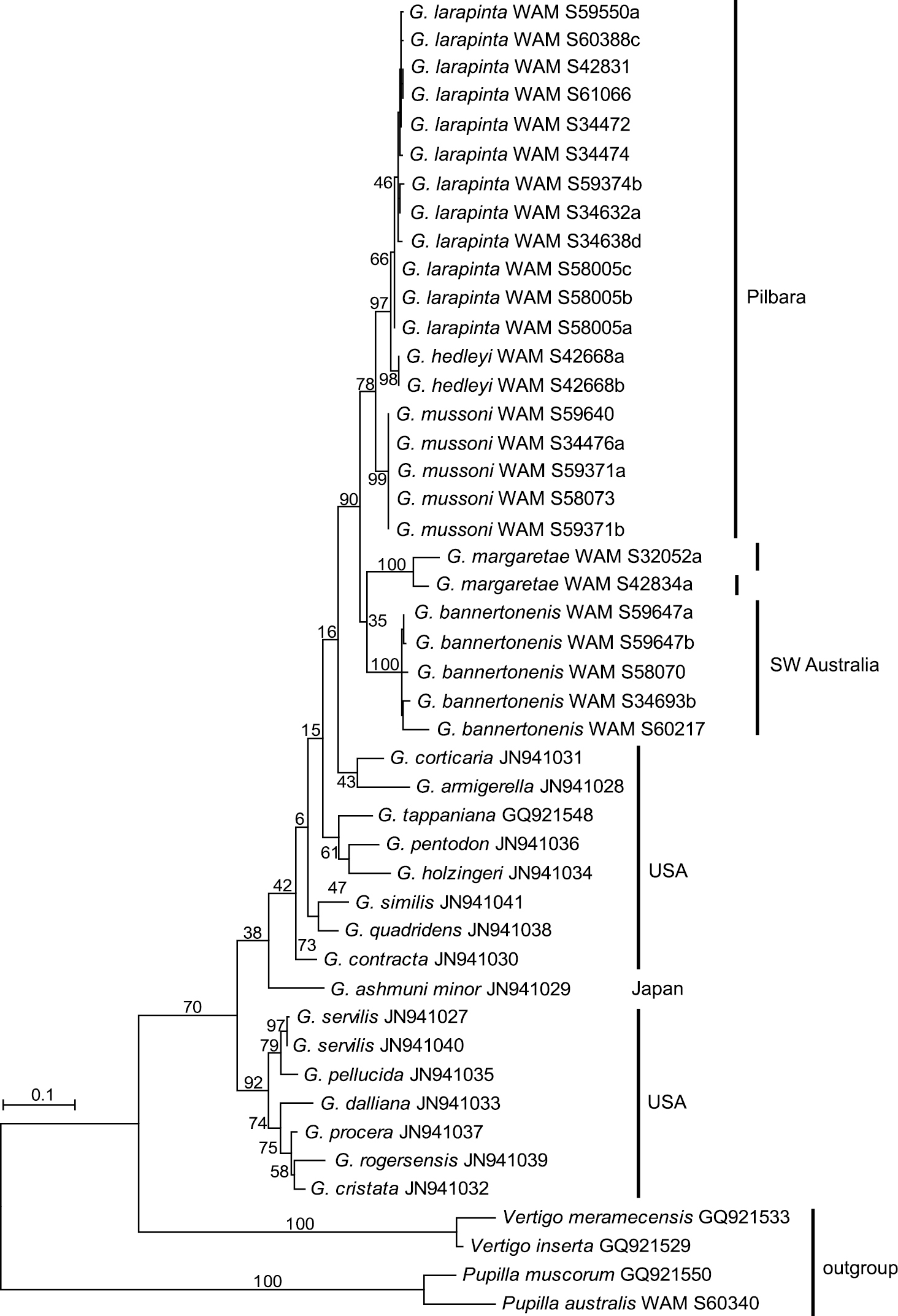
Maximum Likelihood phylogram for 16S based on analysis of 26 new sequences of Gastrocopta from Western Australia and the 16 Genbank sequences made available by
Average pair-wise genetic distances in COI. Shown are (p) uncorrected p-distances, and (TN93) corrected distances by using the model of Tamura-Nei (
| Distance | G. marg | G. bann | G. lara | G. muss | G. held | |
|---|---|---|---|---|---|---|
| G. marg | p | 0.103 | 0.121 | 0.133 | 0.128 | 0.129 |
| TN93 | 0.160 | 0.195 | 0.218 | 0.202 | 0.204 | |
| G. bann | p | 0.030 | 0.147 | 0.132 | 0.142 | |
| TN93 | 0.036 | 0.263 | 0.211 | 0.244 | ||
| G. lara | p | 0.008 | 0.080 | 0.044 | ||
| TN93 | 0.008 | 0.113 | 0.052 | |||
| G. muss | p | 0.000 | 0.081 | |||
| TN93 | 0.000 | 0.115 | ||||
| G. held | p | 0.012 | ||||
| TN93 | 0.013 |
Average pair-wise genetic distances in 16S. Shown are (p) uncorrected p-distances, and (TN93) corrected distances by using the model of Tamura-Nei (
| Distance | G. marg | G. bann | G. lara | G. muss | G. held | |
|---|---|---|---|---|---|---|
| G. marg | p | 0.045 | 0.102 | 0.094 | 0.082 | 0.095 |
| TN93 | 0.053 | 0.139 | 0.126 | 0.103 | 0.126 | |
| G. bann | p | 0.021 | 0.080 | 0.076 | 0.076 | |
| TN93 | 0.024 | 0.106 | 0.096 | 0.099 | ||
| G. lara | p | 0.008 | 0.040 | 0.021 | ||
| TN93 | 0.008 | 0.046 | 0.023 | |||
| G. muss | p | 0.000 | 0.041 | |||
| TN93 | 0.000 | 0.046 | ||||
| G. held | p | 0.000 | ||||
| TN93 | 0.000 |
Based on shell morphology, all Gastrocopta species recorded here (except Gastrocopta sp. CW1) have relatively large distributional ranges. Although limited, the molecular data supports the shell-based delineation of the six species recognized herein. The molecular data also confirms the large distributional ranges of Gastrocopta larapinta and Gastrocopta mussoni by including samples from areas that are about 100 and 550 kilometres apart, respectively. The apparently widespread distribution of Gastrocopta species is probably due to the common ability in which a single Gastrocopta adult can self-fertilize their eggs and establish a new population (
Some of the species Gastrocopta hedleyi and Gastrocopta sp. CW1 are at the southern limits of their range. It is probable that Gastrocopta hedleyi has arrived in the Pilbara as a result of dispersal potential whereas Gastrocopta sp. CW1, found only in the Cape Range in the Pilbara (and an isolated record from Barrow Island) might represent relictual populations from the Miocene. Both Cape Range and Barrow Island contain moist, well sheltered limestone gorges and caves.
Other recorded species represent a range extension from the red centre. These include Gastrocopta mussoni and Gastrocopta larapinta which are common in the Pilbara. This is not suprising given their affinity to arid or semi-arid environments, which persist throughout much of the Pilbara. Future collecting will no doubt show a mostly continuous distribution for these species between the Pilbara and the red centre.
The present CO1 and 16S molecular data set, although small (only 27 individuals sequenced) mostly supports the taxonomic revision of
The Australian species are less well differentiated by means of evolutionary divergence than the American Gastrocopta species, which are separated from each other by interspecific pair-wise
There appears to be tremendous variation in shell shape and size between and within populations of some Gastrocopta species, and often this is associated with variation in apertural barrier structure. This can make separation of species difficult, particularly Gastrocopta larapinta, Gastrocopta hedleyi and Gastrocopta mussoni which share similar apertural barrier structures. It is advisable to collect a large series of individuals so the wide variation in apertural barrier structures can be seen.
In summary, Gastrocopta hedleyi, Gastrocopta larapinta, Gastrocopta. sp. CW1 and Gastrocopta servilis are recorded from the Pilbara region for the first time. Gastrocopta servilis has been a recent introduction to the residential gardens of Karratha. Gastrocopta hedleyi, Gastrocopta larapinta and Gastrocopta mussoni were shown to be common across the Pilbara. Gastrocopta sp. CW1may represent an undescribed species.
We are grateful to the following colleagues, who generously supplied information, photographs or specimens: Jochen Gerber, Vince Kessner, Jeff Nekola, Shirley Slack-Smith, Amanda Lawless, Alison Miller, Mandy Reid, Chris Rowley, Michael Shea and Janet Waterhouse.