





(C) 2013 Bjarte H. Jordal. This is an open access article distributed under the terms of the Creative Commons Attribution License 3.0 (CC-BY), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
For reference, use of the paginated PDF or printed version of this article is recommended.
Citation: Bjarte H. Jordal (2013) Deep phylogenetic divergence between Scolytoplatypus and Remansus, a new genus of Scolytoplatypodini from Madagascar (Coleoptera, Curculionidae, Scolytinae). ZooKeys 352: 9–33. doi: 10.3897/zookeys.352.6212
Scolytoplatypodini Blandford is a monotypic tribe of ambrosia beetles found in Asia, Madagascar and Africa. Only three species are currently known from Madagascar and four additional species are here described as new to science. Phylogenetic analyses of morphological and molecular data revealed that four of the seven endemic species are deeply separated from all other species by genetic and distinct morphological characters and therefore placed in a new genus Remansus Jordal. The split between this ancient lineage and Scolytoplatypus Schaufuss was estimated to approximate Palaeocene age (63 Ma), extending the minimum age of ambrosia feeding for this tribe to the beginning of the Palaeocene‒Eocene thermal maximum (PETM). In addition to the ancient origin of Remansus in Madagascar during the Palaeocene, a second origin occurred in Scolytoplatypus no more than 13 Ma. A geographical origin of the latter in South-Eastern Africa was unequivocally inferred from the phylogenies.
Curculionidae, Scolytinae, Scolytoplatypodini, molecular phylogeny, biogeography, Madagascar, PETM
Madagascar has one of the highest diversity of plants and animals, a diversity reflected not only by the great number of plant and animal species, but also by a huge number of endemic lineages of higher taxonomic ranks. Forest insects are no exception to this pattern. Woodboring weevils in the subfamily Scolytinae and Platypodinae have dozens of genera and hundreds of species endemic to Madagascar, with several cosmopolitan genera forming particular voluminous species group radiations on this island (
Scolytoplatypus is currently the only recognised genus in the tribe Scolytoplatypodini, known from Asia, Madagascar and Africa (
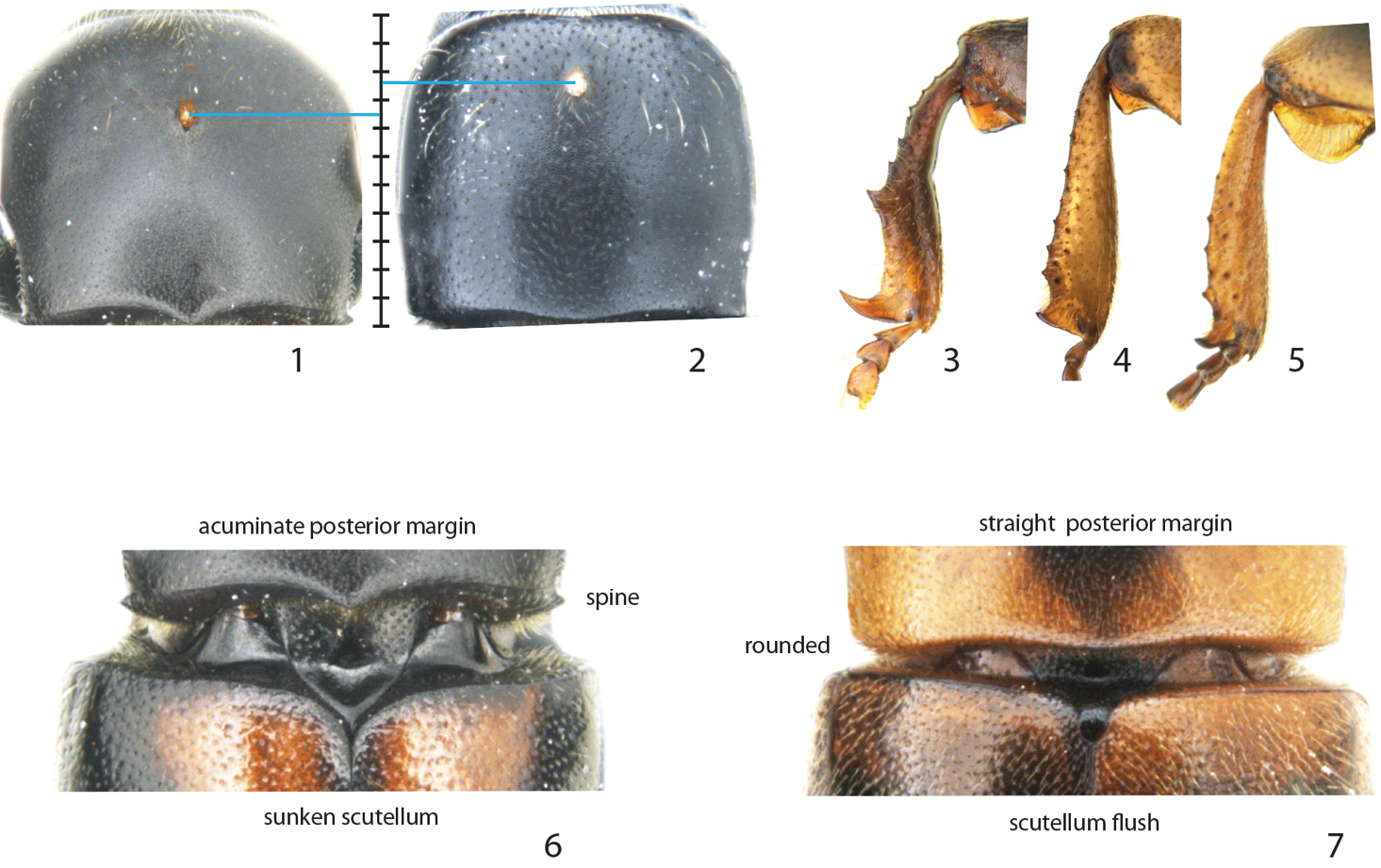
Diagnostic characters for Scolytoplatypus and Remansus. 1 Location of mycangia on the pronotum of Scolytoplatypus hova and 2 Remansus pygmaeus 3 male protibiae of Scolytoplatypus rugosus 4 Remansus serratus and 5 Remansus mutabilis 6 posterior part of pronotum and scutellum in Scolytoplatypus permirus and 7 Remansus mutabilis.
In Madagascar the genus has relatively low diversity with only three species reported, compared to a total of 12 species in Africa (
This paper presents a revision of the Malagasy species of Scolytoplatypodini and attempts to relate these species to the African and Asian members of the genus. A phylogenetic analysis of morphological and molecular data is presented to assess the number of independent clades that occur in Madagascar and to document the existence of a new genus Remansus for the most ancient clade of Malagasy species. Molecular data are used to place the Malagasy species in an evolutionary time frame, and to explore the geographical origin of the scolytoplatypodine fauna in Madagascar.
Material of Malagasy Scolytoplatypodiniwas available from California Academy of Science’s biodiversity inventory 2000–2002 and from the author’s field collecting in 2012. Additional material collected by the author was available from Africa and Asia (Table 1). The following acronyms are used for the material studied: CAS, California Academy of Science; ZMBN, University Museum in Bergen; NHMW, Naturhistorisches Museum Wien.
Material used for phylogenetic analyses, including their GenBank accession numbers.
| Taxon | Country | COI | EF1α | 28S | CAD |
|---|---|---|---|---|---|
| Polydrusus cervinus (Linnaeus) | Norway | HQ883729 | HQ883729 | HQ883568 | HQ883793 |
| Porthetes hispidus (Boheman) | South Africa | HQ883666 | HQ883737 | HQ883577 | HQ883805 |
| Scolytodes acuminatus Wood | Costa Rica | EU191844 | EU191876 | EU090351 | HQ883790 |
| Remansus mutabilis (Schedl) | Madagascar | KF758328 | KF758341 | KF758300 | KF758316 |
| Remansus pygmaeus Jordal, sp. n. | Madagascar | - | KF758338 | KF758294 | KF758310 |
| Remansus sahondrae Jordal, sp. n. | Madagascar | KF758331 | KF758347 | KF758303 | KF758319 |
| Remansus serratus Jordal, sp. n. | Madagascar | - | - | - | - |
| Scolytoplatypus africanus Eggers | Uganda | EU191866 | EU191898 | AF308391 | HQ883822 |
| Scolytoplatypus congonus Schedl | Cameroon | KF758321 | KF758334 | KF758290 | KF758306 |
| Tanzania | KF758322 | KF758335 | KF758291 | KF758307 | |
| Scolytoplatypus eutomoides Blandford | Papua N Guinea | HQ883679 | HQ883748 | EU090345 | HQ883823 |
| Scolytoplatypus fasciatus Hagedorn | South Africa | KF758324 | KF758337 | KF758293 | KF758309 |
| Scolytoplatypus hova Schaufuss | Madagascar Madagascar |
KF758326 KF758327 |
KF758340 KF758344 |
KF758298 KF758299 |
KF758314 KF758315 |
| Scolytoplatypus javanus Eggers | Sarawak | KF758333 | KF758349 | KF758305 | - |
| Scolytoplatypus neglectus Schedl | Cameroon | KF758332 | KF758348 | KF758304 | KF758320 |
| Scolytoplatypus permirus Schaufuss | Madagascar Madagascar Madagascar |
- - KF758325 |
KF758339 KF758342 KF758343 |
KF758295 KF758296 KF758297 |
KF758311 KF758312 KF758313 |
| Scolytoplatypus rugosus Jordal, sp. n. | Madagascar Madagascar |
KF758329 KF758330 |
KF758345 KF758346 |
KF758301 KF758302 |
KF758317 KF758318 |
| Scolytoplatypus truncatus Browne | Cameroon | KF758323 | KF758336 | KF758292 | KF758308 |
| Scolytoplatypus tycon Blandford | Japan | JX263861 | JX264142 | JX263764 | - |
Sequences were generated from the mitochondrial cytochrome oxidase 1 (COI) gene (690 bp) and the three nuclear genes CAD (490 bp), EF1α (857 bp) and 28S (865 aligned sites). DNA was extracted and amplified using the protocols and primers listed in
Morphological characters used in the phylogenetic analysis.
| 1 | Male frons convex (0); concave (1) |
| 2 | Male antennal club similar to female (0); prolonged (1) |
| 3 | Female pronotum with central mycangium absent (0); present (1) |
| 4 | Female mycangium placed about one quarter from anterior margin (0); one third or more from anterior margin (1); n/a (-) |
| 5 | Males with lateral fovea on pronotum absent (0); present (1) |
| 6 | Postero-lateral margin of pronotum straight in both sexes (0); constricted in females only (1); constricted in both male and female (2) |
| 7 | Posterior corner of pronotum rounded (0); with spine (1) |
| 8 | Posterior margin of pronotum straight (0); acuminate (1) |
| 9 | Scutellum is flush with elytra (0); or sunken and near invisible (1) |
| 10 | Striae on posterior half of the elytral disk of the male not impressed (0); strongly and broadly impressed (1) |
| 11 | Male declivity with interstriae 1, 3 and 5 on male declivity flat (0); convex (1); carinate (2) |
| 12 | Female declivity with all striae similarly impressed (0); not impressed or evident (1) |
| 13 | Base of declivity smooth (0); with a carinate ring or serration (1) |
| 14 | Female apex of the elytra rounded (0); extended into a flange (1) |
| 15 | Vestiture scattered and restricted to interstriae (0); uniformly pilose (1) |
| 16 | Females with a patch of longer setae close to the elytral apex absent (0); present (1) |
| 17 | Procoxae contiguous (0); separated (1) |
| 18 | Female protibiae: narrow, parallel-sided and smooth (0); broad, with coarse spines (1) |
| 19 | Male protibiae nearly parallel-sided (0); strongly curved and asymmetric (1) |
| 20 | Prosternum in males simple, smooth (0); or extended anteriorly into nodules or projections (1) |
| 21 | Dorsal side of profemur smooth (0); with spine (1) |
The timing of Malagasy origins was estimated in the software Beast (
Female. Frons flattened to convex; antennal club flat, pilose, without sutures; pronotum with a dorsal mycangium (except seven species), pronotum weakly to strongly constricted laterally in posterior third, hind corners rounded or acute, posterior margin straight to strongly bisinuate; scutellum visible, narrow and sunken or broader and flush with elytra; procoxae widely separated; protibiae broad, on its posterior side with coarse granules and blunt spines, some forming rugae.
Male. Similar to female except frons slightly to very strongly concave, pronotum without mycangium, some with anterior fovea on lateral sides (Asian species), laterally constricted or not in posterior third; protibiae generally smooth on posterior side, sides straight to strongly curved and asymmetrical, laterally with small or large lateral spines.
Scolytoplatypus Schaufuss and Remansus Jordal, gen. n.
Asia, Africa and Madagascar.
Scolytoplatypus permirus Schaufuss.
For a complete diagnosis of the genus and discussions on morphological features, see reviews by
The genus in Madagascar includes three possibly monophyletic species whose ancestor colonized Madagascar rather recently. They have all typical features of the African lineage, including a sharp lateral spine at posterior corners of the pronotum, the female mycangium about one-third or more from the anterior margin, a strongly bisinuate (acuminate) posterior margin of the pronotum, a narrow and sunken scutellum, and strongly asymmetrical male protibiae with a long and curved lateral distal spine. They lack a dorsal spine on the profemur and therefore key out in couplet 7 in
http://species-id.net/wiki/Scolytoplatypus_permirus
Figs 8–13Length 2.5–3.1 mm.
Male. Frons concave, marked at its upper margin by a small heart-shaped tubercle. Pronotum with a short spine in postero-lateral corners, posterior margin strongly bisinuate. Scutellum narrow, sunken between elytra. Elytral disk on posterior third and declivity with carinate interstriae and deeply excavated striae; interstriae 2, 4 and 6 not carinate on declivity; declivity in lateral profile gradually rounded.
Female. Similar to male except frons convex, pronotum with one large mycangial pore, posterior third of pronotum laterally constricted, posterior part of elytral disk smooth, declivity with shallowly impressed striae, interstriae 1, 3 and 5 weakly elevated, elytral apex broadly rounded, extended flange less transverse, with broad v-shaped emargination at suture; protibiae broad, posterior face tuberculate.
The amount of interstrial setae and granules on declivity varies considerably between individuals.
DNA barcodes in Table 1.
New records: Antsiranana Prov, Parc National Montagne d’Ambre, 12°30'52"S, 049°10'53"E, MA-01-01A-01, 21-26 Jan 2001, Malaise trap. Antsiranana Prov, Sakalava Beach [vegetated beach dunes], 12°15'46"S, 049°23'51"E, MA-01-04B-17, 13-20 Aug 2001, malaise trap. Fianarantsoa Prov, Forêt d’Atsirakambiaty, 7.6 km 285° WNW Itremo, 20°35'36"S, 046°33'48"E, BLF7155, 22-26 Jan 2003, EB09 sifted litter (leaf mold, rotten wood). Fianarantsoa Prov, Ranomafana National Park, Belle Vue trail, 21°15'59"S, 047°25'13"E, MA-02-09C-23, 31 Mar-7 Apr 2002; MA-02-09C-36, 24 Jul-4 Aug 2002, MA-02-09C-21, 19-26 Mar 2002; MA-02-09C-25, 14-23 Apr 2002, all in Malaise traps; JIRAMA water works, 21°14'55"S, 047°27'08"E, MA-02-09D-08, 21-24 Dec 2001, malaise trap. Toamasina Prov, Andasibe National Park, botanic garden near entrance, 18°55'35"S, 048°24'28"E, MA-01-08B-18, 01-10 Nov 2001, malaise trap [CAS]. Fianarantsoa prov, Ranomafana NP, Centre ValBio [-21.25, 47.42], alt. 950m, ex Dalbergia branch, 2012: 1x-7, B.Jordal leg; Vato trail [-21.29, 47.42], alt. 1100m, ex Xylopia branch, 2012: 4x-11, B.Jordal leg; Valo area [-21.31, 47.43], alt. 1100m, ex Albizia branch, 2012: 6x-9, B.Jordal leg. [ZMBN].
A fairly common species throughout the wet forests of Madagascar, where it frequently co-occurs with Scolytoplatypus hova Schaufuss. It breeds in small diameter branches, typically ranging between 2–5 cm. Previously known from Montagne d’Ambre and Joffreville in the north of the island, from Antananarivo, and in the east from Perinet in Andasibe national park (

Scolytoplatypus permirus Schaufuss. 8 male lateral view 9 male frons 10 male dorsal view 11 male declivity 12 female dorsal view 13 female declivity.
http://species-id.net/wiki/Scolytoplatypus_hova
Figs 14–19Length 3.5–4.5 mm.
Male. Frons concave, marked at its upper margin by a small protruding tubercle. Pronotum with a short spine in each postero-lateral corner, posterior margin strongly bisinuate. Scutellum narrow, sunken. Posterior half of elytral disk with carinate interstriae and deeply excavated striae; interstriae 2 closer to 3 than to 1 on disk, interstriae 2, 4 and 6 not carinate on declivity; transition from the smooth disk to posterior area with impressed striae angular, appearing hunchbacked, declivity in lateral profile angular.
Female. Similar to male except frons convex, pronotum with one large mycangial pore, posterior third of pronotum laterally constricted, striae not impressed, interstriae 1, 3 and 5 on declivity weakly elevated, declivity in lateral profile rounded, elytral apex transverse and extended between interstriae 1 and 3, with a narrow v-shaped emargination at suture.
DNA barcodes in Table 1.
New records: Antsiranana Prov, 7 km N Joffreville, 12°20'00"S, 049°15'00"E, MA-01-07-12, 13-16 May 2001, Malaise trap; Parc National Montagne d’Ambre [Petit Lac road], 12°31'13"S, 049°10'45"E, MA-01-01D-03, 29 Jan-11 Feb 2001, Malaise trap; Fianarantsoa Prov, Forêt d’Atsirakambiaty, 7.6 km 285° WNW Itremo, 20°35'36"S, 046°33'48"E, BLF7155, 22-26 Jan 2003, EB09 sifted litter (leaf mold, rotten wood); Fianarantsoa Prov, Ranomafana National Park, JIRAMA water works, 21°14'55"S, 047°27'08"E, MA-02-09D-08, 21-24 Dec 2001, malaise trap [CAS]. Fianarantsoa prov, Ranomafana NP, Vato trail [-21.29, 47.42], alt. 1100m, ex Xylopia branch, 2012: 4x-11, B.Jordal leg; Valo area [-21.31, 47.43], alt. 1100m, ex Albizia branch, 2012: 6x-9, B.Jordal leg. [ZMBN].
The biology and distribution of this species is similar to Scolytoplatypus permirus and these two species frequently occur on the same host plant. Previously reported from the Montagne d’Ambre area in the North, from Moramanga and Perinet (Andasibe in Toamasina) east of Antananarivo, from Infanadiana (Fianarantsoa) in the south-east, and Fort Dauphin in the south (

Scolytoplatypus hova Schaufuss. 14 male lateral view 15 male frons 16 male dorsal view 17 male declivity 18 female dorsal view 19 female declivity.
http://zoobank.org/823D10DE-FD29-44BD-BC99-F378CD4A6EE9
http://species-id.net/wiki/Scolytoplatypus_rugosus
Figs 20–25Holotype, male: “Madagascar: Fianarantsoa, Ranomafana, Centre ValBio [-21.25, 47.42], alt. 950m, ex Harungana madagascariensis, 2012: 29ix-13, B. Jordal” / “ZMBN/ENT-Scol-2” Allotype, female: same label as holotype. Paratypes: Madagascar: Fianarantsoa, Ranomafana, Vato trail [-21.29, 47.42] alt. 1100m ex Ocotea branch. 2012: 3x-2, B.Jordal leg, 2♀♀; same data but collecting code 5x-7, 1♂, 1♀. Madagascar: Morondava distr, Miandrivazo 246km W Antsirabe, D. Hauck lgt, 5.i.2002, 1♂. The holotype and five paratypes (“ZMBN/ENT-Scol-3 – ZMBN/ENT-Scol-7”) are deposited in the University Museum of Bergen (ZMBN), one paratype in Miloš Knížek collection, Prague.
Male. Protibiae strongly curved and asymmetrical, scutellum narrowly triangular and sunken between elytra, pronotum with a lateral spine in each posterior corner.
Female. With mycangial pore on pronotum; declivity identical to male; frons and protibiae typical dimorphic as for the genus.
This species is closely related to Scolytoplatypus fasciatus in south-eastern parts of Africa, but differs from that speciesby the lack of interstrial granules on declivity, by the more deeply impressed male frons, and by the uniform dark mature body colour. It differs further from all other African and Malagasy species by the transverse crest along the upper margin of the male frons. It is readily distinguished from males of the closely related Malagasy species Scolytoplatypus hova and Scolytoplatypus permirus by the much less impressed striae on posterior third of the elytral disk. From the African species Scolytoplatypus opacicollis Eggers, Scolytoplatypus obtectus Schedl and Scolytoplatypus fasciatus, it is distinguished by the extended flange of the female elytral apex, and the stouter body plan (1.7–1.8 versus 2.0–2.1 times longer than wide).
DNA barcodes in Table 1.
Male. Length 2.3-2.7 mm, 1.7–1.8× longer than wide; colour dark reddish brown to black.
Head. Eyes separated above by 4.2× their width. Frons concave from vertex to epistoma between inner eye margins, marked above by a distinct transverse crest at median third; impressed area weakly reticulated, with small shallow punctures separated by 1–2× their diameter, except smooth and shiny on a median triangular area on lower third. Vestiture consisting of short fine setae along the upper rim of concave area, and minute setae in punctures in concave area.
Pronotum 0.75× as long as wide, sides subparallel, surface finely reticulated with shallow punctures spaced by 1–2× their diameter; pronotal vestiture consisting of fine short setae arising from punctures, a few longer setae scattered close to anterior margin.
Elytra 1.0–1.1× longer than wide, 1.5–1.6× longer than pronotum; sides almost straight, broadly rounded behind, with an extended apical flange between suture and interstriae 5; striae impressed only on declivity, strial and interstrial punctures on disk confused, spaced by 1–1.5× their diameter; interstriae on declivity slightly raised, shrivelled, punctures asymmetric and confluent, surface strongly reticulate, mesh-like. Interstriae 10 weakly elevated to level of ventrite I. Vestiture consisting of minute setae in punctures only slightly longer than diameter of a puncture.
Legs. Procoxae separated by 0.6× the width of one coxa. Mesocoxae separated by 0.7× the width of one procoxae. Protibiae strongly asymmetrical, with one large laterally curved distal spine, one medium lateral spine and some 6–7 additional lateral small spiny granules.
Ventral vestiture. Metanepisternum with short, bifid setae.
Female. Similar to male in most respect, including the declivity, but differ by the convex frons and more widely separated eyes (4.5× their width), pronotum laterally constricted on posterior third, with mycangial pore on anterior third (0.35), by the broad protibiae with spines and granules on its posterior face, and by the broader prosternum being 0.8 as wide as one procoxa.
The Latin masculine adjective rugosus means ‘wrinkled’ or ‘shrivelled’, referring to the surface of declivity with wrinkled interstriae, confluent asymmetrical punctures and strongly reticulate cuticle.
Only known from the southern range of the Ranomafana National Park. The collections from Harungana (Hypericaceae) and Ocotea (Lauraceae) indicate a broad host plant range typical for the genus. Branches between 2–5 cm in diameter were colonized, where male and female joined in monogamous pairs, with the male guarding the entrance when the female excavated the egg tunnels.

Scolytoplatypus rugosus Jordal, sp. n. 20 male lateral view 21 male frons 22 male dorsal view 23 male declivity 24 female dorsal view 25 female declivity.
Scolytoplatypus mutabilis Schedl. Gender masculine.
Male. Frons concave, antennal funicle 6-segmented, club flattened and pilose without sutures; procoxae widely separated, protibiae narrow, nearly parallel-sided; pronotum not constricted laterally, posterior corners rounded, posterior margin nearly straight; scutellum large, flush with elytra.
Female. Similar to male except frons flat to slightly convex; pronotum with a median mycangium about one-quarter distance from the anterior margin, posterior lateral margins of pronotum weakly constricted; protibiae very stout, broad, with coarse granules and rugae on its posterior side.
Based on the Latin masculine participle meaning ‘left behind’ or ‘having endured’, referring to the first branch of Scolytoplatypodini that has remained in Madagascar since the origin of the tribe.
http://species-id.net/wiki/Remansus_mutabilis
Figs 26–31Holotype, male: Madagascar, Perinet, 16.XI.1952, K.E. Schedl, leg (NHMW).
Length 3.0–3.4 mm.
Male. Frons weakly concave, with a feeble longitudinal carina from epistoma to vertex. Elytra smooth, striae not impressed on disc and declivity, vestiture of dense, short, fine setae not in rows. Protibiae almost parallel-sided, with a moderately sized, curved lateral spine at distal end, and four additional smaller spines (granules) towards base. Procoxae rounded, separated by 0.7× the width of one coxa.
Female. Similar to male except frons weakly convex, eyes more widely separated; declivity near apex with two patches of longer and more densely placed setae; protibiae broad, with granules on posterior side. Procoxae very broad, separated by 0.8× the width of one coxa.
The female is here reported and diagnosed for the first time. It differs externally from the male only in those dimorphic features given in the diagnosis. This is the only species in Remansus with both male and female known.
DNA barcodes in Table 1.
New records: Fianarantsoa Prov, Forêt d’Atsirakambiaty, 7.6 km 285° WNW Itremo, 20°35'36"S, 046°33'48"E, BLF7155, 22–26 Jan 2003, EB09 sifted litter (leaf mold, rotten wood). Fianarantsoa Prov, Ranomafana National Park, JIRAMA water works, 21°14'55"S, 047°27'08"E, MA-02-09D-13 and MA-02-09D-08, 21-24 Dec 2001, malaise trap. Fianarantsoa Prov, Ranomafana National Park, radio tower, 21°15'03"S, 047°24'26"E, MA-02-09B-05, 28 Nov-6 Dec 2001, malaise trap, all material in CAS. Fianarantsoa prov, Ranomafana NP, Valo area [-21.31, 47.43], alt. 1100 m, ex Albizia branch, 6. Oct. 2012, B.Jordal leg. (ZMBN).
Previously reported from Anjanaharibe and Marojejy forest reserves in the Northern part of the island, and in Perinet east of Antananarivo (

Remansus mutabilis (Schedl). 26 male lateral view 27 male frons 28 male dorsal view 29 male declivity 30 female dorsal view 31 female declivity.
http://zoobank.org/BAA6B09A-B9A3-4664-8A5A-7980711414DA
http://species-id.net/wiki/Remansus_sahondrae
Figs 32–35Holotype, female: “Madagascar: Fianarantsoa, Ranomafana, Vato trail [-21.29, 47.42] alt. 1100m ex Eugenia branch. 2012: 2x-9, Sahondra Rahanitriniaina” / “ZMBN/ENT-Scol-8” Paratype, female: same data as holotype except museum label “ZMBN/ENT-Scol-9”. The holotype and paratype are deposited in the University Museum of Bergen (ZMBN).
Female. Pronotum with mycangium only one-quarter distance from anterior margin, posterior corners rounded, posterior margin straight; scutellum flat at the same level as elytra. It is closely related to Scolytoplatypus mutabilis, but distinguished from that species by the visible striae, larger elytral punctures, the coarse, short setae on the declivity, and by the much smaller body size.
DNA barcodes in Table 1.
Female. Length 2.2–2.3 mm, 1.9–2.0× longer than wide; colour black.
Head. Eyes separated above by 4.6× their width. Frons broadly flattened from vertex to epistoma, surface strongly reticulated, with large punctures spaced by their diameter, except smooth and impunctate on a triangular area on central lower third, a short median longitudinal keel dividing triangular area. Vestiture consisting of a few scattered fine setae.
Pronotum 0.8× as long as wide, sides almost straight, barely constricted on posterior third, surface reticulate with minute shallow punctures spaced on average by their diameter; center of median mycangial pore positioned from anterior margin about 0.24× the pronotal length; pronotal vestiture consisting of sparse fine short setae, with somewhat longer setae on anterior half.
Elytra 1.1× longer than wide, 1.6× longer than pronotum; striae not impressed, punctures in irregular rows, spaced within a row on disc by 2–3× their diameter, closer together towards declivital summit, almost subconfluent; interstrial punctures similar to those in striae, irregularly placed, confused. Interstriae 10 elevated to level of ventrite I. Declivity with small granules, strial punctures 2‒3× larger than on disk, oblong or asymmetric, subconfluent; vestiture on declivity consisting of stiff, short, curved setae, with a patch of longer setae on each side of suture close to apex; a subapical rim runs from the suture to about interstria 8.
Legs. Procoxae separated by 0.5× the width of one coxa. Mesocoxae separated by 0.6× the width of one procoxa. Protibiae broad, with one distal laterally curved spine, and many additional small spines or granules on its posterior side.
Ventral vestiture. Metanepisternum with long bifid setae.
Male: unknown.
Named after Sahondra Rahanitriniaina, our helpful Malagasy collegue who collected the type specimens and many other scolytine beetles during our joint field trip to Ranamafana National Park.
Only known from the type locality in the Ranomafana National Park. Two females were taken from a fallen branch of Eugenia (Myrtaceae).

Remansus sahondrae Jordal, sp. n., female. 32 lateral view 33 frons 34 dorsal view 35 declivity.
http://zoobank.org/5776128D-753D-4E2C-ABBB-C91FD86E7788
http://species-id.net/wiki/Remansus_pygmaeus
Figs 36–39Holotype, female: “Madagascar, Fianarantsoa, Ranomafana National Park, 5 km NE Centre ValBio [-21.24, 47.41]. Ex Weinmannia twig, 10. Oct. 2012, B. Jordal leg.” / “ZMBN/ENT-Scol-10”. The holotype is deposited in University Museum of Bergen (ZMBN).
Female. Posterior corners of pronotum rounded; scutellum flat at the same level as elytra; declivity characteristically truncated and marked by a circular blunt rim. One of the smallest known species in the tribe and the only species smaller than 2 mm in Africa and Madagascar. Closely related to Remansus serratus, but distinguished by the much smaller size (1.7 vs. 3.7 mm), the nearly glabrous elytral disk and posterior half of pronotum.
DNA barcodes in Table 1.
Female. Length 1.7 mm, 1.85× longer than wide; colour black.
Head. Eyes separated above by 4.0× their width. Frons broadly flattened from vertex to epistoma, surface strongly reticulated, with tiny shallow punctures spaced by 1.5–2× their diameter, except smooth and impunctate on a triangular area on central lower third. Epistoma elevated, shiny, with a short median carina extending from epistoma to impunctate area. Vestiture consisting of a few scattered fine setae.
Pronotum 0.8× as long as wide, sides almost straight, weakly constricted on posterior third, surface reticulate with minute shallow punctures spaced on average by 2× their diameter; median mycangial pore on anterior fifth round with a tuft of setae; pronotal vestiture consisting of sparse fine short setae, and about 20 much longer erect setae on anterior half.
Elytra 1.05× longer than wide, 1.5× longer than pronotum; sides almost straight, slightly constricted just before declivity, broadly triangular at apex; striae not impressed except weakly so at declivital margin; strial and interstrial punctures on disk entirely confused, shallow, with minute setae of variable length. Interstriae 10 elevated to level of ventrite I. Declivity dull, rugose, punctures variable but generally larger than on disk; vestiture on declivity of fine ground setae and fewer but coarser stiff and slightly curved short setae.
Legs. Procoxae separated by 0.5× the width of one coxa. Mesocoxae separated by 0.6× the width of one procoxae. Protibiae broad, with one large laterally curved distal spine, and five additional lateral small spines or granules, with small granules on the posterior face.
Ventral vestiture. Metanepisternum with bifid long setae.
Male: unknown.
The Latin masculine adjective pygmaeus pertaining to the mythical race of African dwarfs, referring to the relative small size for this species, being the smallest species of Scolytoplatypodini in Africa and Madagascar.
Only known from the type locality in the Ranomafana National Park. One female and one pupa were taken from a fallen branch of Weinmannia (Cunoniaceae), about 1.5 cm in diameter. The egg tunnel was transversely spiral shaped, with eight pupation chambers directed longitudinally, like bullet chambers in a revolver barrel. Based on empty pupal chambers the brood size in this type of host is 6–10 (n=3).

Remansus pygmaeus Jordal, sp. n. female. 36 lateral view 37 frons 38 dorsal view 39 declivity.
http://zoobank.org/94073590-56A8-45D0-9EF1-5D38FC16FD19
http://species-id.net/wiki/Remansus_serratus
Figs 40–43Holotype, male: “Madagascar, Antsiranana Prov, Parc National Montagne d’Ambre (Petit Lac Road) 12°31'13"S, 49°10'45"E, MA-01-01D-03, 29 Jan – 11 Feb 2001, Malaise trap”. The holotype is deposited in California Academy of Science (CAS).
Male. With a unique Amasa-like truncateddeclivity and rounded posterior angles of the pronotum. Characters suggest a close relationship to Remansus pygmaeus Jordal, but it is distinguished from that species by the much larger size, narrow body shape, the pilose body, and by the slightly elevated scutellum that is not entirely flushwith the elytra. It is further distinguished from males of Remansus mutabilis by the dentate upper declivital margin.
Male. Length 3.7 mm, 2.1× longer than wide; colour yellowish brown, darker brown on declivity and elytral lateral margins.
Head. Eyes separated above by 4.2× their width. Frons concave from vertex to epistoma via inner eye margins; upper half rugosely, densely punctured, lower half smooth and shiny. Vestiture consisting of fine setae increasing in length towards upper part of concave area, longest and most dense along upper margin from vertex to upper level of eyes.
Pronotum 0.9× as long as wide, sides subparrallel, brodest on anterior half, broadly, transversely rounded in front; surface finely reticulated with minute shallow punctures irregularly spaced by 1–4× their diameter; pronotal vestiture consisting of fine short setae, slightly longer close to anterior margin.
Elytra 1.2× longer than wide, 1.7× longer than pronotum; sides almost straight, broadly triangular at apex; decivital margin marked by a dentate rim, each incision marks the end of diskal stria; striae otherwise not impressed, interstriae on disk only reckognised by the lighter colour, punctures not clearly visible, confused and minute. Interstriae 10 elevated to level of ventrite I. Declivity rugosely granulated, largest granules on interstriae 1 (suture). Vestiture consisting of dense fine setae spaced on disk by less than one-third the length of each seta, on declivity about 2–3× longer than setae on disk.
Legs. Procoxae separated by 0.4× the width of one coxa. Mesocoxae separated by 0.5× the width of one procoxae. Protibiae narrow, with one larger distally lateral curved spine, and 6–7 additional lateral small spines, posterior face mainly smooth, with 4–5 tiny granules close to lateral edge.
Ventral vestiture. Metanepisternum mainly with long simple setae, a few shorter bifid setae anteriorly.
Female: unknown.
The Latin masculine adjective serratus means ‘serrated’, referring to the short pointed projections from interstriae at the declivital summit.
A single specimen was collected in a Malaise trap just south of Montagne d’Ambre.

Remansus serratus Jordal, sp. n., male. 40 lateral view 41 frons 42 dorsal view 43 declivity.
| 1 | Scutellum flat, broad, flush with the elytra and clearly visible; posterior edge of pronotum almost straight; female mycangium closer to anterior margin (0.22–0.28 of pronotum length); male protibiae almost parallel-sided; postero-lateral corners of pronotum without acute spine | Remansus Jordal, gen. n., 2 |
| – | Scutellum narrowly triangular, obliquely depressed between elytra; posterior edge of pronotum bisinuate; female mycangium closer to centre (0.33–0.36 of pronotum length); male protibiae asymmetric with large incision between lateral spines 1 and 2; postero-lateral corners of pronotum with an acute spine pointing laterad | Scolytoplatypus Schaufuss, 5 |
| 2 | Declivity abrupt, marked by a circum-declivital ring, surface rough, not shining, vestiture evenly distributed in both sexes | 3 |
| – | Declivity steep, gently rounded, not truncated, surface sub-shining, females with a patch of longer setae close to elytral apex | 4 |
| 3 | Larger, 3.7 mm long, elytra and pronotum densely covered by short fine setae | Remansus serratus Jordal, sp. n. |
| – | Smaller, 1.7 mm long, elytral disk and posterior half of pronotum almost glabrous | Remansus pygmaeus Jordal, sp. n. |
| 4 | Larger, 3.0–3.4 mm long, striae not impressed, all punctures small | Remansus mutabilis (Schedl) |
| – | Smaller, 2.2–2.3 mm, striae evident from base, slightly impressed on declivital summit, punctures on declivity much larger than on disk | Remansus sahondrae Jordal, sp. n. |
| 5 | Male and female declivity similar, without sharp interstrial carinae on posterior part of male elytral disk, interstriae on declivity only slightly raised and shrivelled, granules minute (2.4–2.7 mm) | Scolytoplatypus rugosus Jordal, sp. n. |
| – | Males with striae on posterior third of elytral disk deeply excavated between sharply elevated interstriae, male and female declivity with at least interstriae 1 and 3 distinctly raised towards apex, linear, with distinct granules (2.5–4.5 mm) | 6 |
| 6 | Larger, 3.5–4.5 mm; male interstriae 2 closer to 3 than to 1, disk and declivity profile subangular, hunchbacked | Scolytoplatypus hova Schaufuss |
| – | Smaller, 2.5–3.0 mm; male interstriae more regularly spaced, declivity profile rounded | Scolytoplatypus permirus Schaufuss |
The tree topology resulting from the Bayesian and parsimony analyses of the combined molecular data were near identical and all nodes except one received high support (Fig. 44). Scolytoplatypodini was monophyletic, with a clade consisting of the Malagasy species Remansus mutabilis, Remansus pygmaeus and Remansus sahondrae as the sister group to all species in Scolytoplatypus. The Asian species of Scolytoplatypus formed a sister clade to the Malagasy and African species of the genus. The three Malagasy species of Scolytoplatypus were nested in a derived monophyletic position in the Bayesian analysis, while Scolytoplatypus rugosus was sister to the South-East African Scolytoplatypus fasciatus in the parsimony analysis.
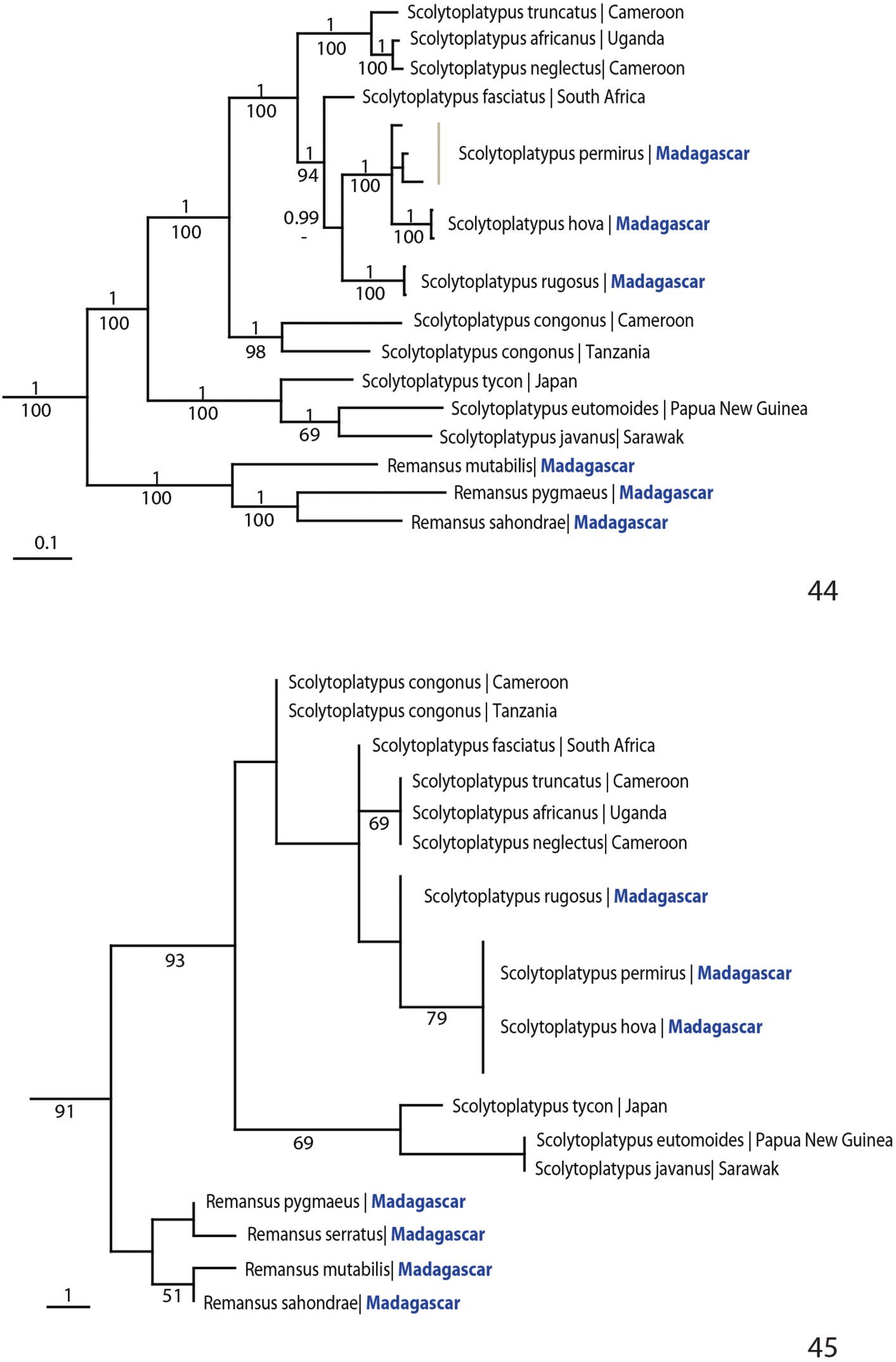
Phylogeny of Scolytoplatypodini. 44 Tree topology resulting from Bayesian analysis of 2902 nucleotide sites from one mitochondrial and three nuclear genes. Posterior probabilities are given above, parsimony bootstrap support below 45 Tree topology resulting from parsimony analysis of 21 morphological characters (Table 2, Appendix).
Parsimony analysis of the morphological data resulted in 30 most parsimonious trees. All trees revealed a strongly supported sister relationship between Remansus (including also Remansus serratus) and Scolytoplatypus. Some of the trees placed Scolytoplatypus congonus as the sister to the remaining African and Asian species of Scolytoplatypus, while in two of the trees the Asian species were sister to all African species (Fig. 45). The ambiguity was likely due to Scolytoplatypus congonus having two character states shared with Remansus, including a broad scutellum that is flush with the elytra and the nearly straight posterior margin of the pronotum (Fig. 7). However, Scolytoplatypus congonus has spiny hind corners of the pronotum, asymmetrical male protibiae and female mycangium behind anterior third as in other Scolytoplatypus.Molecular data unambiguously resolved this issue and strongly supported the Asian-African split in Scolytoplatypus, with Scolytoplatypus congonus as an early diverging lineage in Africa.
The strongly supported split between the two basal clades justify the designation of a new genus Remansus. Although largely defined by plesiomorphic characters such as the ‘normal’ male protibiae (Figs 4 and 5), rounded posterior corners of pronotum, and a visible large scutellum that is flush with the elytra (Fig. 7), the new genus is also supported by a uniquely derived feature in the female mycangium being displaced anteriorly (Fig. 2 vs. Fig. 1). Taken together with the large genetic divergence in mitochondrial and nuclear loci, the new genus appears to be well supported, and hence constitutes the second genus in the tribe Scolytoplatypodini.
The molecular data also distinguished the Asian species of Scolytoplatypus clearly from the African species of that genus. A distinction between these two clades is supported by several morphological characters in the phylogenetic analysis as was noted in two previous publications (
In addition to four species of Remansus, three additional species are found in Madagascar, in the genus Scolytoplatypus. The new species Scolytoplatypus rugosus was shown to be most closely related to the other two Malagasy species Scolytoplatypus hova and Scolytoplatypus permirus, although some analyses placed this species closer to Scolytoplatypus fasciatus (= Scolytoplatypus obtectus Schedl?), a species very similar to Scolytoplatypus opacicollis Eggers. The congruence between the Bayesian analysis of the molecular data and morphology suggests that the most likely scenario is a monophyletic group of Malagasy species, as sister clade to Scolytoplatypus fasciatus. The three Malagasy species of Scolytoplatypus share features on the female elytra by having a broad subtransverse apical flange, which, at least in comparison to the African species, seems unique. As typical Scolytoplatypus they share with all African species an acute spine at the postero-lateral corner of the pronotum, a bisinuate posterior margin of pronotum, a narrow scutellum that is sunken between the elytra, and similarly shaped antennal clubs in males and females. The three species therefore belong to the African group of Scolytoplatypus.
With the addition of several new taxa from Madagascar, the age estimated for the tribe Scolytoplatypodini becomes considerably older than previously reported (
Divergence time estimates for Scolytoplatypodini.
| Clade | Analysis A | Analysis B | ||
|---|---|---|---|---|
| median | 95 % CI | median | 95 % CI | |
| Scolytoplatypodini: stem age | 91.0 | 71.6‒106.4 | 59.1 | 45.0‒75.6 |
| Scolytoplatypodini: crown age (Remansus vs. Scolytoplatypus) | 63.1 | 45.6‒80.3 | 42.3 | 34.1‒53.0 |
| Scolytoplatypus: crown age (Asia split from Africa) | 51.6 | 36.7‒67.7 | 35.2 | 29.3‒40.7 |
| Remansus: crown age | 33.2 | 16.1‒50.8 | 22.0 | 9.9‒33.5 |
| Recent Madagascar clade: stem age | 13.1 | 6.9‒21.4 | 8.8 | 3.7‒13.9 |
| Recent Madagascar clade: crown age | 12.5 | 6.5‒18.6 | 7.3 | 3.2‒12.0 |
The ancient split between Remansus in Madagascar and the Asian/African clade of Scolytoplatypus is apparently younger than the latest Gondwanan vicariance event involving Madagascar. The separation of India from Madagascar occurred at least 80 Ma (
The author appreciates the help provided by MICET to organize our field work in Madagascar, and wish to thank the Madagascar National Park authorities for research and export permits. Also many thanks to the staff at Centre ValBio in Ranomafana NP for logistic and technical support. In addition a great thank you to the Department of Entomology at the University of Antananarivo, especially Sahondra Lalao Rahanitriniaina, for excellent field assistance. California Academy of Science provided loan of specimens, facilitated by Anthony Cognato at Michigan State University. The feedback provided by reviewers was of great help in improving the manuscript. I would also like to thank the editor for suggesting the genus name Remansus. This research was funded by grant 214232/F20 from the Norwegian Research Council.
Data matrix for morphological characters (Table 2) used in the phylogenetic analysis.
| Polydrusus | 0 | 0 | 0 | - | 0 | 0 | 0 | 0 | ? | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Porthetes | 0 | 0 | 0 | - | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Scolytodes | 0 | 0 | 0 | - | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 |
| Remansus mutabilis | 1 | 0 | 1 | 1 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 1 | 1 | 1 | 1 | 0 | 0 | 0 |
| Remansus pygmaeus | ? | ? | 1 | 1 | 0 | ? | 0 | 0 | 0 | ? | ? | 1 | 1 | 0 | 0 | 0 | 1 | 1 | ? | 0 | 0 |
| Remansus sahondrae | ? | ? | 1 | 1 | 0 | ? | 0 | 0 | 0 | ? | ? | 1 | 0 | 0 | 0 | 1 | 1 | 1 | ? | 0 | 0 |
| Remansus serratus | 1 | 0 | ? | ? | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 1 | 1 | ? | 1 | 0 | 1 | ? | 0 | 0 | 0 |
| Scolytoplatypus africanus | 1 | 0 | 1 | 0 | 0 | 1 | 1 | 1 | 1 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 1 | 0 | 1 |
| Scolytoplatypus congonus C | 1 | 0 | 1 | 0 | 0 | 1 | 1 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 1 | 0 | 0 |
| Scolytoplatypus congonus T | 1 | 0 | 1 | 0 | 0 | 1 | 1 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 1 | 0 | 0 |
| Scolytoplatypus eutomoides | 1 | 1 | 1 | 0 | 1 | 2 | 1 | 1 | 1 | 0 | 2 | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 1 | 1 | 0 |
| Scolytoplatypus fasciatus | 1 | 0 | 1 | 0 | 0 | 1 | 1 | 1 | 1 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 1 | 0 | 0 |
| Scolytoplatypus hova | 1 | 0 | 1 | 0 | 0 | 1 | 1 | 1 | 1 | 1 | 2 | 0 | 0 | 1 | 0 | 0 | 1 | 1 | 1 | 0 | 0 |
| Scolytoplatypus javanus | 1 | 1 | 1 | 0 | 1 | 2 | 1 | 1 | 1 | 0 | 2 | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 1 | 1 | 0 |
| Scolytoplatypus neglectus | 1 | 0 | 1 | 0 | 0 | 1 | 1 | 1 | 1 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 1 | 0 | 1 |
| Scolytoplatypus permirus | 1 | 0 | 1 | 0 | 0 | 1 | 1 | 1 | 1 | 1 | 2 | 0 | 0 | 1 | 0 | 0 | 1 | 1 | 1 | 0 | 0 |
| Scolytoplatypus rugosus | 1 | 0 | 1 | 0 | 0 | 1 | 1 | 1 | 1 | 0 | 1 | 0 | 0 | 1 | 0 | 0 | 1 | 1 | 1 | 0 | 0 |
| Scolytoplatypus truncatus | 1 | 0 | 1 | 0 | 0 | 1 | 1 | 1 | 1 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 1 | 0 | 1 |
| Scolytoplatypus tycon | 1 | 1 | 1 | 0 | 1 | 2 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 1 | 0 | 0 |