





(C) 2013 Fahrettin Küçük. This is an open access article distributed under the terms of the Creative Commons Attribution License 3.0 (CC-BY), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
For reference, use of the paginated PDF or printed version of this article is recommended.
Citation: Küçük F, Gülle İ, Güçlü SS, Çiftçi Y, Erdoğan Ö (2013) A new Pseudophoxinus (Teleostei, Cyprinidae) species from Southwestern Anatolia, with remarks on the distribution of the genus in western Anatolia. ZooKeys 320: 29–41. doi: 10.3897/zookeys.320.4447
Pseudophoxinus burduricus sp. n. is described from drainages of Salda and Burdur lakes, southwestern Turkey. It is distinguished from other Anatolian Pseudophoxinus by a combination of characters: lateral line incomplete, with 21–39 (commonly 26–37) perforated scales and 47–57+1-2 scales in lateral series; 10½–12½ scale rows between lateral line and dorsal fin origin, 3–4(5) scale rows between lateral line and the pelvic fin origin; dorsal fin commonly with 7½ branched rays; anal fin commonly with 6½ branched rays; 7–8(9) gill rakers on the first branchial arch; a faint and diffuse epidermal black stripe from eye to caudal fin base in alive and preserved individuals; mouth slightly subterminal, tip of mouth cleft on about level of lower margin of eye; snout rounded, its length greater than eye diameter. Comparison is given with all Pseudophoxinus species from western Anatolia.
Western Anatolia, Cyprinidae, Taxonomy, Pseudophoxinus, new species
According to
Morphological and phylogenetic distinctness of Pseudophoxinus egridiri (
As stated by
According to molecular data presented by
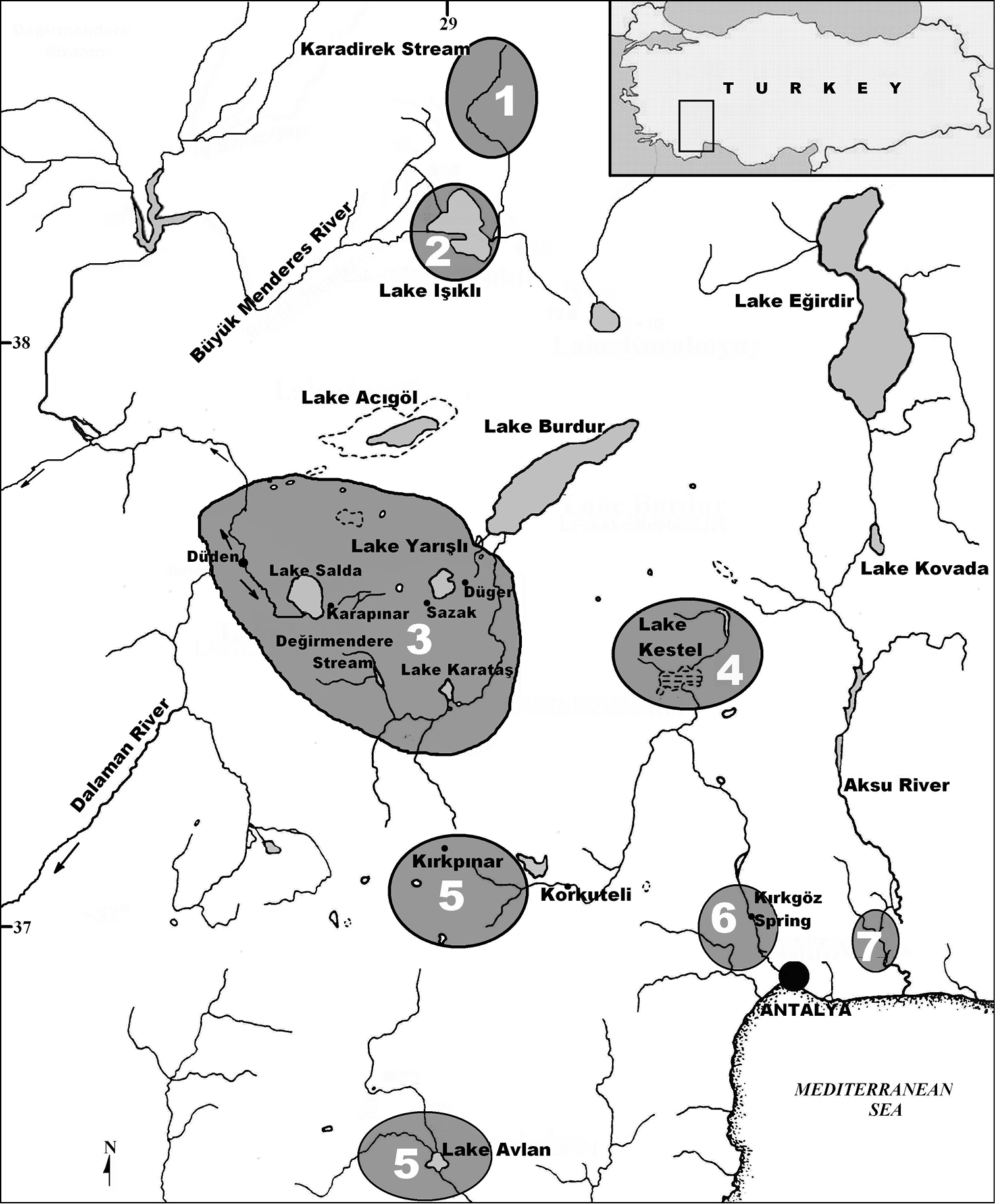
Map showing localities of Pseudophoxinus species in western Anatolia1 Pseudophoxinus maeandricus 2 Pseudophoxinus maeandri 3 Pseudophoxinus burduricus sp. n. 4 Pseudophoxinus ninae 5 Pseudophoxinus evliyae 6 Pseudophoxinus antalyae 7 Pseudophoxinus alii
Fish specimens were caught by pulsed DC electrofishing equipment and killed by over anaesthetization, preserved in 5% formalin. Material is deposited in: IFC-ESUF, Inland Fishes Collection, Eğirdir Fisheries Faculty of Süleyman Demirel University. Counts and measurements follow
The morphometric characters of the two species of Pseudophoxinus from Turkey were compared by Principal Component Analysis (PCA) using a covariance matrix on log–transformed measurements and counts with the software package PAST version 1.8 (
Abbreviations. CSO, supraorbital canal; CIO, infraorbital canal; CPM, preoperculo-mandibular canal; HL, lateral head length; SL, standard length. IFC-ESUF, Inland Fishes Collection, Eğirdir Fisheries Faculty of Süleyman Demirel University, Turkey. IUSHM, Istanbul University, Science Faculty, Hydrobiology Museum, Istanbul.
http://zoobank.org/5734C2B8-1D58-40E4-ABD6-43A9C610B1AB
http://species-id.net/wiki/Pseudophoxinus_burduricus
Figures 2, 3IFC-ESUF 0427, female, 62.80 mm SL; Turkey, Burdur Prov., Değirmendere Creek, Karamanlı, Lake Burdur drainage; 37°24'18"N, 29°49'06"E, 07 November 2009, coll. F. Küçük, İ. Gülle and Ö. Erdoğan.
IFC-ESUF 0428, 17 (11 males, 6 females), 39.90–86.69 mm SL; same as holotype.
IFC-ESUF 0236, 7, 45.31–54.40 mm SL; Burdur Prov., Salda Stream near Lake Salda; F. Küçük, M.A. Atalay, 13 June 1998. –IFC-ESUF 0289, 3, 31.98–71.56 mm SL, Burdur Prov., Düğer Spring; F. Küçük, A. Altun, M. Telli, 05 August 2006. –IFC-ESUF 0429, 5, 51.34–80.08 mm SL, Burdur Prov., Sazak Spring near Lake Yarışlı; F. Küçük, İ. Gülle, S.S. Güçlü, 13 July 2009. -IFC-ESUF 0430, 6, 41.91–55.95 mm SL, Burdur Prov., Salda Stream near Lake Salda; F. Küçük, İ. Gülle, S.S. Güçlü, 13 December 2009. –IFC-ESUF 0449, 8, 46.10-60.43 mm SL, Burdur Prov.; Salda Stream near Lake Salda; F. Küçük, İ. Gülle, 16 April 2010. –IFC-ESUF 0475, 7, 46.10–60.43 mm SL, Burdur Prov.; Dereköy Stream; F. Küçük, İ. Gülle, S.S. Güçlü. 18 May 2012.
Pseudophoxinus burduricus is distinguished from all other species of Anatolian Pseudophoxinus by the following unique combination of characters: head short, its length equal or slightly greater than body depth at dorsal fin origin; mouth slightly subterminal, the tip of the mouth cleft on approximately level of with lower margin of eye; snout rounded, its length greater than eye diameter; a faint and diffuse epidermal black stripe from eye to caudal fin base in alive and preserved individuals; pared fins and caudal peduncle distinctly sexual dimorphic (male with longer pelvic and pectoral fins and slenderer caudal peduncle); lateral line incomplete, with 21–37 (commonly 26–37) perforated scales and 47–57+1-2 scales in lateral series (commonly 50–55); 10½–12½ scale rows between lateral line and dorsal fin origin; 3–4 rarely 5 scale rows between lateral line and the pelvic fin origin; 7–8 (9) gill rakers on the first branchial arch; pharyngeal teeth 5–4 or 5–5, slightly serrated and hooked at tip; dorsal fin commonly with 7 (8)½ branched rays; anal fin with 6 (7)½ branched rays.
See Figs 2–3 for general appearance and Tables 1-2 for morphometric and meristic data.
A Moderately deep-bodied, elongate and wide headed species. Dorsal profile of body slightly convex in predorsal area, ventral profile more convex than dorsal profile. Predorsal distance 52–57% SL, mean 55.0 and preanal distance 69–74% SL, mean 72.0. Head short, its length 26–28% SL, mean 26.8, approximately 1.0–1.1 times body depth at dorsal-fin origin, and its dorsal profile slightly convex on snout. Head depth at interorbital region 2.1–2.7 times eye diameter and 1.3–1.6 times interorbital distance. Mouth slightly subterminal, posterior extremity of upper jaw slightly in front of anterior margin of eye. Snout rounded, its length 27–33% HL, mean 29.6, greater than eye diameter. Caudal peduncle slightly deep, its depth 1.4–1.9, mean 1.6 times in its length.

Pseudophoxinus burduricus sp. n. IFC-ESUF 0427, holotype, 62.80 mm SL, female; Turkey: Değirmendere Creek, Burdur.

Pseudophoxinus burduricus sp. n. IFC-ESUF 0428, paratype, 65.82 mm SL, female; Turkey: Değirmendere Creek, Burdur.
Morphometry of Pseudophoxinus burduricus sp. n. (holotype IFC-ESUF 427, paratypes IFC-ESUF 428, n=17) and Pseudophoxinus ninae (IFC-ESUF 263, n=15).
| Pseudophoxinus burduricus | Pseudophoxinus ninae | ||
|---|---|---|---|
| Holotype | Paratypes | ||
| In percent of standard length | |||
| Head length | 26.9 | 25.7–27.8 (26.8)±0.6 | 27.1–30.7 (28.6) ±1.0 |
| Body depth of dorsal fin origin | 26.6 | 24.1–27.1 (25.7) ±0.7 | 27.8–32.1 (29.3) ±1.2 |
| Predorsal distance | 56.6 | 51.6–57.4 (55.0) ±1.7 | 56.6–60.9 (58.0) ±1.1 |
| Prepelvic distance | 52.1 | 50.4–54.7 (52.6) ±1.5 | 53.9–56.7 (55.7) ±0.9 |
| Preanal distance | 72.4 | 68.7–74.0 (72.0) ±1.5 | 72.6–76.5 (74.5) ±1.1 |
| Distance between pectoral and anal-fin origins | 48.4 | 43.0–51.1 (48.1) ±2.5 | 47.3–51.3 (49.0) ±1.1 |
| Distance between pectoral and pelvic-fin origins | 26.7 | 24.6–31.2 (27.7) ±1.8 | 27.3–30.6 (29.0) ±0.8 |
| Distance between pelvic and anal-fin origins | 21.8 | 17.7–22.8 (20.0) ±1.6 | 18.3–22.2 (20.0) ±1.4 |
| Dorsal fin depth | 20.8 | 16.9–22.1 (20.1) ±1.7 | 16.9–21.7 (19.6) ±1.5 |
| Anal fin length | 18.3 | 15.2–20.6 (17.8) ±1.5 | 14.1–20.1 (17.0) ±1.7 |
| Pectoral fin length | 19.4 | 17.6–25.0 (21.3) ±2.1 | 17.7–22.1 (19.8) ±1.4 |
| Pelvic fin length | 15.6 | 13.7–19.1 (16.3) ±1.6 | 13.6–18.6 (15.7) ±1.4 |
| Caudal peduncle length | 20.5 | 17.5–23.0 (20.1) ±1.4 | 17.3–21.3 (19.0) ±1.3 |
| Caudal peduncle depth | 13.1 | 11.4–14.9 (12.6) ±0.9 | 12.7–14.9 (13.5) ±0.6 |
| In percent of head length | |||
| Snout length | 29.4 | 26.5–32.9 (29.6) ±1.9 | 22.0–28.9 (25.4) ±1.8 |
| Eye diameter | 22.6 | 22.0–25.9 (24.5) ±1.4 | 20.5–23.3 (21.9) ±0.9 |
| Interorbital distance | 38.3 | 34.6–40.5 (38.2) ±1.6 | 30.6–36.3 (33.6) ±1.9 |
| Head width at nape | 59.8 | 55.3–61.7 (58.7) ±1.6 | 53.8–63.2 (58.0) ±2.8 |
| Head depth at interorbital region | 60.6 | 50.2–62.2 (56.7) ±3.3 | 49.7–58.2 (53.7) ±2.6 |
| Head depth at nape | 79.4 | 76.5–85.3 (80.7) ±2.8 | 70.8–80.7 (74.9) ±1.5 |
| Operculum depth | 42.0 | 40.0–48.8 (43.4) ±2.3 | 36.5–46.6 (40.1) ±2.6 |
| Lower jaw length | 36.0 | 32.6–38.6 (35.5) ±1.2 | 31.3–35.6 (33.5) ±2.1 |
Meristic features of the western Anatolian Pseudophoxinus species.
| Species | Lateral series | Lateral line | Pharyngeal teeth | Total vertebrae | Abdominal vertebrae | Caudal vertebrae |
|---|---|---|---|---|---|---|
| Pseudophoxinus alii | 41–44 | 38–41 | 5–5 | 37–39 | 21–23 | 16–17 |
| Pseudophoxinus antalyae | 52–64 | 42–59 | 5–5 | 37–39 | 20–22 | 16–17 |
| Pseudophoxinus anatolicus |
93–109 | 78–93 | 5–5 | 41–42 | 23–24 | 17–18 |
| Pseudophoxinus battalgilae |
53–61 | 53–60 | 5–5 | 37–38(39) | 20–21 | 17–18 |
| Pseudophoxinus burduricus sp. n. | 47–57 | 21–39 | 5–4(5) | 36–39 | 21–22 | 15–17 |
| Pseudophoxinus crassus |
65–78 | 62–73 | 5–5 | 40 | 22 | 18 |
| Pseudophoxinus elizavetea |
60–68 | 33–60 | 5–5 | 36–37 (38) | 22 | 14–16 |
| Pseudophoxinus evliyae | 52–66 | 16–30 | 5–4 | 36–37 | 21–22 | 14–16 |
| Pseudophoxinus hittitorum | 83–96 | 83–94 | 5–5 | 39 | 21–22 | 17–18 |
| Pseudophoxinus maeandri | 41–45 | 19–27 | 5–4 | 35–36 | 19–20 | 15–16 |
| Pseudophoxinus maeandricus |
66–67 | 58–65 | 5–5 | 36–37 | 20–21 | 16 |
| Pseudophoxinus ninae | 46–53 | 10–32 | 5–4 | 36 | 21 | 15 |
(*) from
(**)
Lateral line incomplete, usually reaching above anal fin origin, 32 perforated scales in the holotype (26–37 in paratypes), 47–57+1-2 scales in lateral series. Dorsal fin with 3 simple and 7½ (n=18, in one specimen 8½) branched rays, outer margin slightly convex. Anal fin with 3 simple and 6 (15)½ or 7 (3)½ branched rays, outer margin slightly convex. Pectoral fin with 13–14 branched rays, outer margin straight or slightly convex. Pelvic-fin with 7 branched rays. Caudal fin forked, lobes rounded. There is no pelvic axillary lobe and keel between posterior pelvic fin base and anus. Pharyngeal teeth 5–4 or 5–5, slightly serrated, hooked at tip (developed on the left side). Gill rakers short and thick, with 7–8 (9) in outer side of first gill arch. Preoperculo-mandibular (CPM) and infraorbital (CIO) sensory canals disconnected, CSO with 10–12 pores, CIO with 14–19 pores, CPM with 14–18 pores, total vertebrae 36–39, 21–22 abdominal and 15–17 caudal vertebrae, vertebral formulae: 36–39:21–22+15–17.
Sexual dimorphism. In Değirmendere population, there is no tubercules on snout and head in males, which have longer pelvic and pectoral fins and slender caudal peduncles than females. In Lake Salda population, on the other hand, tubercules present in males on entire body (concentrated on operculum) and all fins except for the caudal fin.
Coloration. Body silvery, dorsal light brown or olive green and scales irregular with small epidermal spots in specimenslarger than50 mm SL. There is a faint epidermal black or violet (in Sazak population) stripe along lateral midline from eye to caudal fin base in alive. The dark stripe indistinct or slightly distinct in anterior part of body but distinct in posterior part of body in preserved specimens. Lateral line scales with small brown to black spots above and below pores in some individuals. Fin membranes whitish or light grey, rays with black-spotted. In individuals smaller than 50 mm SL: body silvery, dorsal dusty grey, ventral pearl grey.
Pseudophoxinus burduricus is known only from the lakes and their sources in Lake Burdur Endorheic Basin: Değirmendere Creek, Lake Karataş, Düğer and Sazak (or Kümbet) springs, Dereköy Stream, Lake Salda and Salda Stream (Fig. 1). Değirmendere Creek is a 5 km long creek flowing into the Karamanlı reservoir, which is connected with artificial reservoir Lake Karataş. Düğer spring is a source of Lake Burdur, while Sazak Spring is a source of Lake Yarışlı. Dereköy Stream is an approximately 5 km long rivulet formerly draining into Çorak (or Akgöl) Lake which, due to a small reservoir constructed in 1970, no more can reach the lake. The new species can be encountered in the shallow parts of the reservoir and the small stream flowing into it.
Other species present were: Chondrostoma fahirae (Ladiges, 1960), Oxynoemacheilus anatolicus Erk’akan, Özeren & Nalbant, 2008 and Oncorhynchus mykiss (Walbaum, 1792) (an escape from fish farms in Karamanlı reservoir)in Değirmendere, in Düğer Spring only Oxynoemacheilus anatolicus and in Salda Stream only Aphanius splendens (Kosswig & Sözer 1945).
The species is named after the Burdur Province where the type locality is located.
In this study, we also morphologically compared the new species with Pseudophoxinus ninae (the Onaç Stream and Kestel Swamp), Pseudophoxinus evliyae (dried Lake Söğüt [Kırkpınar Village, Korkuteli] and source of Lake Avlan [Elmalı]), Pseudophoxinus maeandri [Lake Işıklı], Pseudophoxinus maeandricus (Karadirek Stream [Upper Büyük Menderes basin]), Pseudophoxinus alii (Köprüçay and Ilıca Stream), Pseudophoxinus battalgilae (Manavgat River basin, Lake Akgöl [Ereğli], Lake Çavuşcu [Ilgın] and Lake Suğla [Seydişehir]), Pseudophoxinus fahrettini (Köprüçay River basin [Bağıllı Village and Değirmenözü Stream]), Pseudophoxinus antalyae (Kırkgöz Spring, Karamanlı Stream and tributaries of Düden canal in Antalya), and Pseudophoxinus elizavetae (Sultansazlığı [Kayseri]).
Pseudophoxinus burduricus is most similar to Pseudophoxinus ninae (Fig. 4). Itis distinguished from Pseudophoxinus ninae by having fewer branched pelvic fin rays (7, vs. 8 or 9), slightly fewer gill rakers in outer side of the first gill arch (7–8, rarely 9, vs. 8–9) and a longer caudal peduncle (caudal peduncle length 1.4–1.9 times caudal peduncle, vs. 1.3–1.4). It further differs from Pseudophoxinus ninae by having a shorter predorsal distance (52–57% SL, mean 55.0, vs. 57–61, mean 58.0), a shorter preanal distance (69–74% SL, mean 72.0, vs. 73–77, mean 74.5), a somewhat shorter head (head length 26–28 % SL, mean 26.8, vs. 27–31, mean 28.6), and narrower and relatively deeper hyomandibular bone (vs. wide and shallow) (see Figs 5a, b, c). Also Pseudophoxinus burduricus and Pseudophoxinus ninae were compared by Principal Component Analysis (PCA). The PCA was performed in using 18 morphometric characters of the two Pseudophoxinus species. The PCA clearly separated Pseudophoxinus burduricus from Pseudophoxinus ninae (Fig. 6). Variables loading on the first metric PC I–II are given in Table 3.

Pseudophoxinus ninae IFC-ESUF 0263, 66.37 mm SL; Turkey: Pınargözü Spring-Bucak.
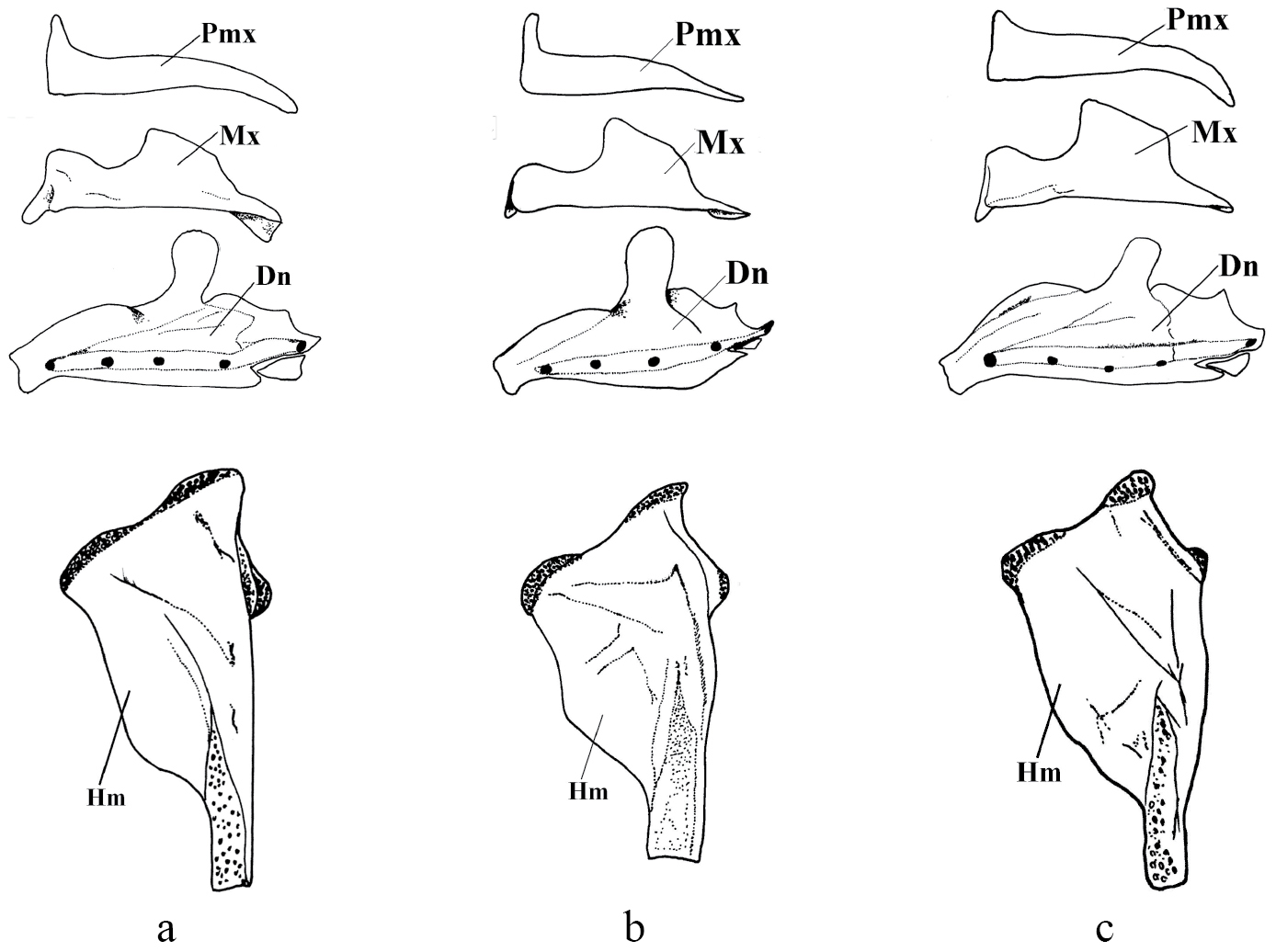
Left jaws and hyomandibular bones of Pseudophoxinus burduricus sp. n. (a) Pseudophoxinus ninae (b)and Pseudophoxinus maeandri (c) (Pmx: premaxilla, Mx: maxilla, Dn: Dentale, Hm: Hyomandibulare)
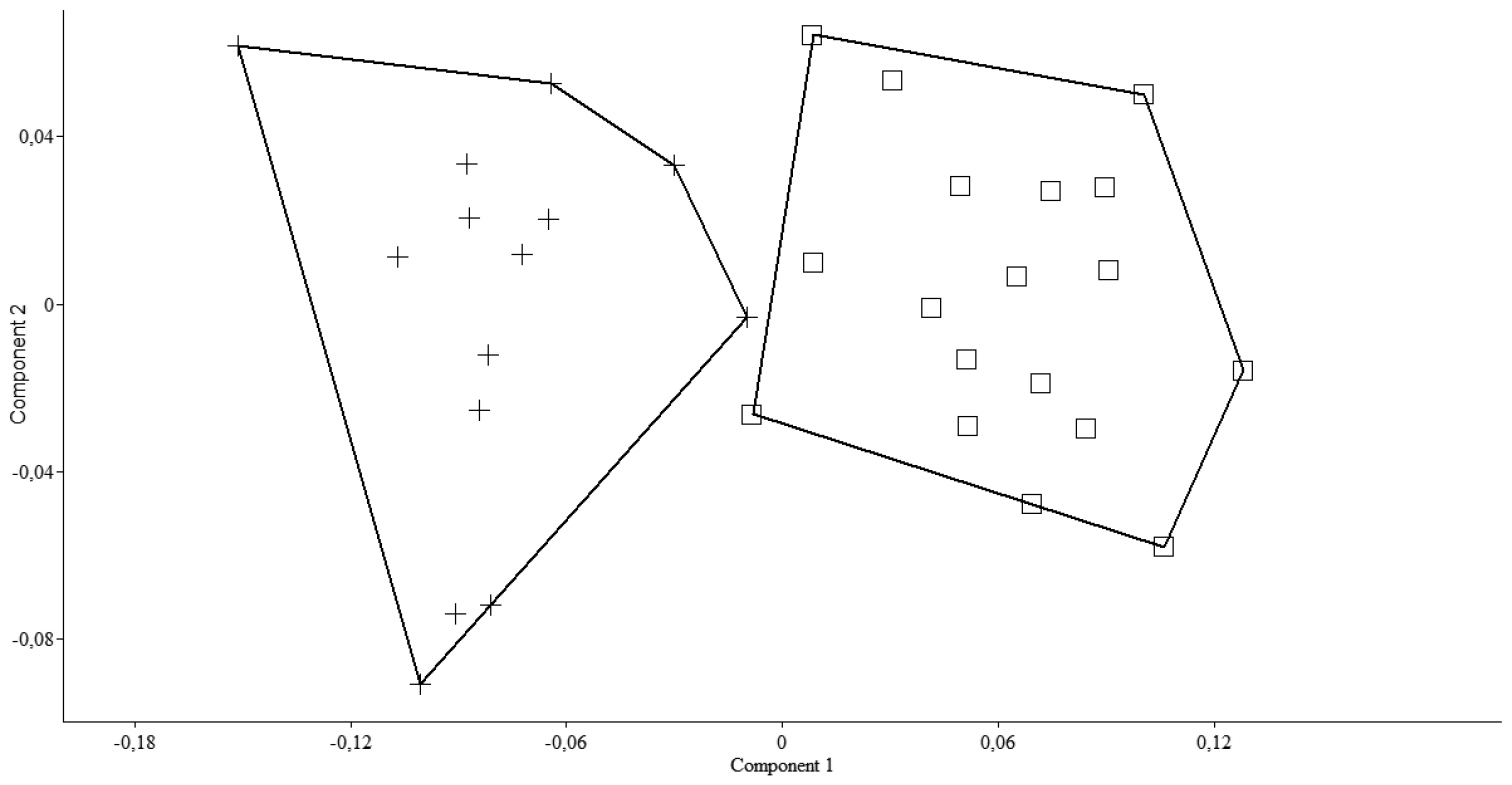
A scatter plot of the scores of the first two principal components (PC I, PC II) for 32 specimens of the two Pseudophoxinus species (Pseudophoxinus burduricus sp. n. (+) and Pseudophoxinus ninae (□), based on 18 morphometric characters.
Character loading on principal components I–II for 18 measurements taken on 32 specimen of two Pseudophoxinus species (Pseudophoxinus burduricus sp. n. and Pseudophoxinus ninae).
| Morphometric features | ||
|---|---|---|
| In percent of standard length | PC I | PC II |
| Head length | 0.196 | -0.103 |
| Body depth of dorsal-fin origin | 0.335 | 0.222 |
| Predorsal length | 0.140 | 0.119 |
| Prepelvic length | 0.147 | 0.145 |
| Preanal length | 0.083 | 0.156 |
| Dist. from pectoral-fin origin to anal fin | 0.060 | 0.331 |
| Dist. from pectoral-fin origin to pelvic fin | 0.140 | 0.396 |
| Dist. from pelvic-fin origin to anal fin | 0.012 | 0.368 |
| Length of caudal peduncle | -0.300 | -0.383 |
| Depth of caudal peduncle | 0.180 | 0.112 |
| In percent of head length | ||
| Snout length | -0.445 | 0.322 |
| Eye diameter | -0.285 | -0.118 |
| Interorbital distance | -0.420 | 0.166 |
| Head width (at operculum) | -0.054 | 0.300 |
| Head depth (at interorbital region) | -0.202 | 0.080 |
| Operculum depth | -0.290 | 0.155 |
| Head depth operculum | -0.208 | 0.164 |
| Length of lower jaw | -0.172 | 0.164 |
Pseudophoxinus burduricus is distinguished from Pseudophoxinus evliyae by having fewer scales in lateral series (47–57+1-2, vs. 54–64+1-2), fewer branched pelvic fin rays (7, vs. 8), fewer gill rakers in outer side of first gill arch (7-8, rarely 9, vs. 8-9 to 10 in some specimens), fewer scales between lateral line and dorsal fin origin (10½-12½, vs. 13½-15½), and a faint epidermal black or violet stripe along lateral midline from eye to caudal fin base (vs. black prominent stripe). Pseudophoxinus burduricus is distinguished from Pseudophoxinus maeandri by having more lateral line and lateral series scales (21–39, vs. 19–27 and 47–57+1-2, vs. 40–44+1-2 respectively), more total vertebrae, 36-39 (vs. 35–36) (see Table 2). Pseudophoxinus burduricus differs from Pseudophoxinus maeandricus by having an incomplete lateral line (vs. complete), a shorter pelvic-fin (reaching to anus, vs. not reaching), wider and deeper head (head width at nape 55-62 % HL, vs. 46–48; head depth at nape 77–85 % HL, vs. 67–73). Pseudophoxinus burduricus isdistinguished from Pseudophoxinus alii by having more scales in the lateral series (47–57+1-2, vs. 38-43+1-2), fewer perforated scales (21–39, vs. 38–41) and smaller eyes (eye diameter 22–26 % HL, vs. 26–32). Pseudophoxinus burduricus is distinguished Pseudophoxinus antalyae by presence of a faint black or violet lateral stripe (vs. plain golden or orange stripe when alive), rounded snout (vs. pointed) and shorter pharyngeal teeth. Pseudophoxinus burduricus is distinguished from Pseudophoxinus battalgilae by an incomplete lateral line (vs. complete), fewer gill rakers in outer side of first gill arch (7–8, rarely 9, vs. 13–16), fewer branched anal fin rays (6–7, vs. 8, respectively), and the absence of a keel between the pelvic fin base and the anus (vs. presence). Pseudophoxinus burduricus is distinguished Pseudophoxinus fahrettini by an incomplete lateral line (vs. complete), fewer perforated scales lateral line (21–39, vs. 73–88) and fewer gill rakers in outer side of first gill arch (7–8, rarely 9, vs. 11–13). Pseudophoxinus burduricus is distinguished Pseudophoxinus elizavetae by having fewer scales in the lateral series (47–57+1-2, vs. 56-62+2-3), fewer gill rakers in outer side of first gill arch (7–8, rarely 9, vs. 11–13).
Pseudophoxinus alii: IFC-ESUF 0169, 13 paratyps, 53.33–98.48 mm SL; Antalya Prov.: Ilıca Stream at Manavgat, F. Küçük, 05 May 1996.
Pseudophoxinus antalyae: IFC-ESUF 0159, 10, 64.07–97.10 mm SL; Antalya Prov.: Düden Canal, W.V.Neer, F. Küçük, R. Wildekamp, M. Ünlüsayın, 28 July 1996.
Pseudophoxinus battalgilae: IFC-ESUF 0161, 18, 46.51-109.68 mm SL; Antalya Prov.: Oymapınar Dam Lake at Manavgat, F. Küçük, 05 May 1996.
Pseudophoxinus elizavetae: IFC-ESUF 0174b, 10, 49.29-67.47 SL; Kayseri Prov.: Sultansazlığı, M.A. Atalay, 23 August 2004.
Pseudophoxinus evliyae: IFC-ESUF 0237, 26, 25.38–57.51 SL; Antalya Prov.: Kırkpınar-Korkuteli, F. Küçük, İ. Gülle, 10 May 1998. IFC-ESUF 0269, 10, 26.17–66.92 mm SL; Antalya Prov.: Kırkpınar-Korkuteli, F. Küçük, T. Şahan, 25 May 2007. -IFC-ESUF 0268, 1, 73.39 mm SL; Antalya Prov.: Kazanpınarı-Elmalı, F. Küçük, T. Şahan, 25 May 2007.
Pseudophoxinus maeandri: IFC-ESUF 0248, 8, 46.70–55.96 mm SL; Denizli Prov.: Lake Işıklı source, F. Küçük, M.A. Atalay, N. Bogutskaya & A.Naseka, 14 August 2006.
Pseudophoxinus maeandricus: 3, 60.71–78.70 mm SL; Afyon Prov.: Karadirek Stream-Sandıklı, V. Yeğen, 29 June 2006.
Pseudophoxinus ninae: IFC-ESUF 0263, 4, 48.9–67.5 mm SL; Burdur Prov.: Pınargözü-Bucak, F. Küçük, T. Şahan, 25 May 2007. –IUSHM 33900-928, 15, 46.4–68.4 mm SL; Burdur Prov.: Onaç Stream, M. Özuluğ, J. Freyhof, 12 June 2006.
This study was supported by the Research Fund of Süleyman Demirel University (grant SDÜ-08.M.1354). The authors thank to Dr. M. Özuluğ (İstanbul Üniversity) and V. Yeğen (Ministry of Food, Agriculture and Livestock) for donating some fish specimens, and Dr. Davut Turan (Recep Tayyip Erdoğan University) and three anonymous reviewers for commenting on earlier versions of the manuscript.