





(C) 2013 Robert Perger. This is an open access article distributed under the terms of the Creative Commons Attribution License 3.0 (CC-BY), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
For reference, use of the paginated PDF or printed version of this article is recommended.
Citation: Perger R, Grossi PC (2013) Revision of the rhinoceros beetle genus Oryctophileurus Kolbe with description of a new species, the male of O. varicosus Prell, and notes on biogeography (Scarabaeoidea, Dynastinae, Phileurini). ZooKeys 346: 1–16. doi: 10.3897/zookeys.346.6114
The genus Oryctophileurus is reviewed and its validity is supported by a combination of the following apomorphic characters: a single cephalic horn with lateral carina, pronotal cavity with ocellate punctures and two teeth or tubercles close behind the anterior pronotal margin. The male of Oryctophileurus varicosus Prell, 1934, is described for the first time. A new species, Oryctophileurus guerrai Perger & Grossi sp. n., from subhumid Tucuman-Bolivian forest in the Southern Bolivian Andes is described. The new species is distinguished from its closest relative, O. armicollis Prell, 1911, by a narrower distance between the inner teeth of the dorsal pronotal protuberances and a reduced area of weakly developed ocellate punctures above the posterolateral pronotal margin. The occurrence of Oryctophileurus species in areas of endemism along the eastern slope of the tropical Andes suggests that these populations represent biogeographic “relicts”, and the discovery of Oryctophileurus guerrai sp. n. in the southern Bolivian Andes suggests that this area is underrated with respect to insect diversity and endemism.
El género Oryctophileurus es revisado y su validez confirmada por la combinación de los siguientes caracteres apomórficos: un cuerno cefálico con carina lateral, cavidad pronotal con orificios ocelados y dos dientes o tubérculos detrás del margen anterior pronotal. El macho de Oryctophileurus varicosus Prell, 1934, es descrito por primera vez. Una nueva especie Oryctophileurus guerrai Perger & Grossi sp. n. es descrita del bosque subhúmedo Tucumano-Boliviano en el sureste de los Andes. La nueva especie se distingue por su parentesco más cercano a O. armicollis Prell, 1911, por la estrecha distancia entre los dientes interiores de las protuberancias en la región dorsal pronotal y una reducida área con escasos orificios ocelados debajo del margen posterolateral pronotal. Las especies de Oryctophileurus ocurren en áreas de endemismo a lo largo de la Vertiente este de los Andes tropicales y sugieren que estas poblaciones representan relictos biogeográficos, y el descubrimiento de Oryctophileurus guerrai Perger & Grossi sp. n. en el sudeste de los Andes Bolivianos indica que esta área está subestimada en cuanto a la diversidad y endemismo de insectos.
Andes, Melolonthidae, Neotropical, relictual species, South America, Tucuman-Bolivian forest
The evolution of exaggerated morphological traits such as cephalic and pronotal horns in male rhinoceros beetles (Dynastinae) has been of interest to biologists for centuries (
Oryctophileurus was established by
The purpose of this contribution is to stabilize the taxonomy of this group through the careful examination of all species originally described in Oryctophileurus and Amblyodus. A new species of Oryctophileurus from the southern Bolivian Andes and the male of Oryctophileurusvaricosus are described the first time, and their biogeography is reviewed and briefly discussed.
We examined 13 specimens deposited in the collection of the Museum für Naturkunde (Humboldt Universität), Berlin, Germany (ZMHB); the Museu de Zoologia, Universidade de São Paulo, São Paulo, Brazil (MZSP); and the Everardo and Paschoal Grossi Private Collection (Nova Friburgo, RJ, Brazil) (EPGC).
Additional specimens were collected during a biodiversity survey headed by the first author in the southern Bolivian Andes, in the northwestern buffer zone of the TariquíaFlora and Fauna National Reserve, department of Tarija, Bolivia. Several transects of about 6 km of subandine, subhumid, semi-deciduous Tucuman-Bolivian forest (
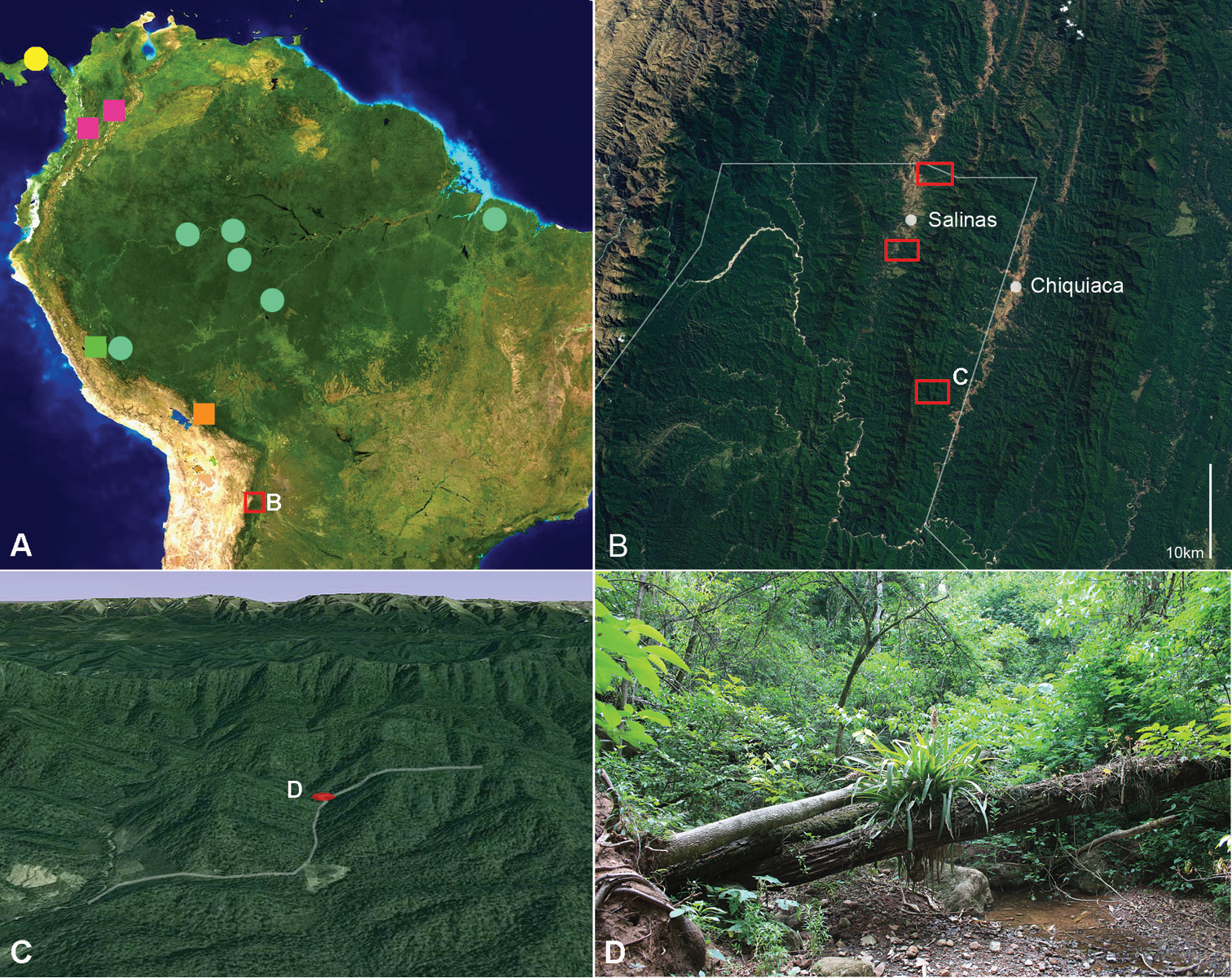
A map of distributional locations of South American Amblyodus and Oryctophileurus species (partly adapted from

Dorsal and lateralhabitus of males. A and B Amblyodus taurus Westwood, 1878 C and D Amblyodus castroi Grossi & Grossi, 2012, scale bar 5 mm.
Morphological characters were examined with a stereomicroscope, and specimens were sexed via genital dissection. Morphometric measurements were taken with a digital caliper. Body length was measured from the apex of the clypeus to the apex of the pygidium.
A map (Fig. 1A) with the known distribution of treated taxa is included to facilitate the interpretation of biogeographical relationships. Distributional data from the literature is only considered when based on properly identified specimens. Records of Oryctophileurus species from Central America (
The following abbreviations were used: department, dep.; province, prov.; municipality, muni.
Phileurus nasicornis Burmeister, 1847 (original combination) (Fig. 3).
Species of Oryctophileurus are distinguished from other Phileurini by a combination of the following apomorphic characters: a single cephalic horn with lateral carina, pronotal cavity with ocellate punctures and two teeth or tubercles close to the anterior pronotal margin, teeth vertically positioned at about the same level as the outer eye margins.
The following characters were cited by
Based on the combination of apomorphic characters of Oryctophileurus that lacks in Amblyodus we suggest retaining the genus Oryctophileurus.
| 1 | Horn on frons not projecting above anterior edges of pronotal protuberance (Figs 3B, D). Distance between inner teeth of dorsal pronotal protuberance (Figs 3A, C) narrower than distance between eyes. Development of horn and pronotal protuberance equal in both sexes. Color dark brown | Oryctophileurus nasicornis (Burmeister) |
| – | Horn on frons projecting above anterior edges of pronotal protuberance in both sexes. Horn longer in males than in females of the same size. Color black | 2 |
| 2 | Development of pronotal protuberance subequal in both sexes. Elytral striae and interstitial punctures arranged in irregular lines, punctures enlarged (Figs 4A, C) | Oryctophileurus varicosus Prell |
| – | Pronotal protuberance more strongly developed in males than females of the similar size. Elytral striae and interstitial punctures arranged in regular lines | 3 |
| 3 | Distance between inner teeth of dorsal pronotal protuberance in male wider than width between eyes (Fig. 7A); in females separated by a concavity with ocellate punctures (Fig. 6C). Pronotal concavity accounts for 40–50 % of the dorsal pronotal surface in males. Ocellate punctures above posterior-lateral pronotal margin continuous (Fig. 7C) | Oryctophileurus armicollis Prell |
| – | Distance between inner teeth of dorsal pronotal protuberance in male as wide as width between eyes (Fig. 7B), in females separated by a small fissure only (Fig. 8C). Anterior pronotal concavity accounts for about 30% of the dorsal pronotal surface in males. Area of ocellate punctures above posteriolateral pronotal margin reduced laterally (Fig. 7D), punctures small, shallow and sparse | Oryctophileurus guerrai Perger & Grossi, sp. n. |
http://species-id.net/wiki/Oryctophileurus_nasicornis
Fig. 3Colombia: Cauca dep., Cauca Valley: 1 male with body length 19 mm, det. Endrödi 1976 (ZMHB); Boyacá dep., Muzo muni.: 1 female with body length 20 mm, S. Apollin coll., det. Ohaus, revised by
Color dark brown, moderately shining. Horn on frons not projecting above anterior edges of pronotal protuberance, weakly recurved, in female with truncate apex; ocular canthus rounded in male, and subquadrate in female. Pronotum broadly rounded, more in female. Distance between inner teeth of dorsal pronotal protuberance narrower than distance between eyes. Pronotal protuberance on same level with elytra. Development of horn and pronotal protuberance equal in both sexes. Elytra striate, with five well defined discal striae; striae regular, at sides weakly defined; punctures regular, about the same size of those at disc. Meso and metatibiae with 4 to 5 distal teeth. Pygidium regularly convex in both sexes, more in female, and densely and finely wrinkled. Parameres with basal half broad, apex slender, straight.

Dorsal and lateral habitus of Oryctophileurus nasicornis (Burmeister, 1847) A and B male C and D female, scale bar 5 mm.
The species is known only from the Cauca Valley in Colombia, which is surrounded by the parallel, peninsula-like projections of the Andean Cordillera Occidental and Cordillera Central and the Muzo municipality which is situated on the eastern slope of the Cordillera Oriental (Fig. 1A). These areas include wet premontane forest and, to a lesser extent, upper montane forests and paramos (
http://species-id.net/wiki/Oryctophileurus_varicosus
Figs 4; 5C–DPeru: holotype, female, body length 20.4 mm, unknown locality (ZMHB).
Peru: Junin dep.: Satipo prov., Chanchamayo Valley: 1 male with body length 20 mm (EPGC); Satipo prov., Rio Tambo valley, Paraíso Tuncama, ~1300 m a.s.l.: 1 female with body length 19.4 mm, (EPGC); Rios Pichis & Perene, 600–900 m a.s.l., Soc. Geog. De Lima col., 1 male (MZSP).
Color black, moderately shining to shining. Enlarged elytral punctures. Punctures and elytral striae arranged in irregular lines (Figs 4A, C). Horn widely projected above body in male. Pronotal protuberance comparably flat and developed as in female.

Dorsal and lateral habitus of Oryctophileurus varicosus Prell, 1934, A and B male C and D female, scale bar 5 mm.
Male. (Figs 4A, B). Body elongate, cylindrical. Surface entirely black, glabrous, moderately shiny to shiny; dorsally almost totally punctate; punctures ocellate, moderate to very large. Legs, sterna, second abdominal ventrite, and basal margin of pygidium setose.
Head. Surface laterally wrinkled, in frontal view rugose. Clypeus subtriangular, apex rounded and emarginated, weakly reflexed; clypeal carina absent; sides concave. Canthus widely rounded and extending into middle of eye. Frons with long, recurved, cylindrical horn with narrow apex; each side of horn with elongated, weak carina, anterior surface with slightly concave furrow, furrow reaching apex. Mandible tridentate, teeth upturned. Mentum with longitudinal furrow wider anteriorly and posterior concavity narrow with subparallel sides.
Pronotum. Shape subquadrate, narrower than elytra together. Discal area covered by ocellate punctures combined with C-shaped, coalescent punctures; discal surface flat and declivous anteriorly (Fig. 4A, B); anterior and lateral margins complete with a marginal bead, concave at middle; posterior marginal bead absent. Pronotal disc carinate, carina convex, smooth; anterior carina more pronounced; near anterior border with a conspicuous tubercle present in each anterolateral corner; posterior carina joined posteriorly on pronotal margin, bisinuous. Anterior angle acute, posterior rounded. Middle apex laterally with smooth convex carina, intercalated by rugose area and coarse punctures. Prosternal process long, trapezoidal, concave at base and posteriorly produced; base with a spine like posterior process.
Elytra. Striae irregular, and not defined, even laterally; punctures ocellate, irregular, larger on discal area and becoming smaller laterally and posteriorly; elytral apices densely punctate, punctures small to moderate; apical umbone convex, smooth. Scutellum triangular, densely punctate; punctures ocellate, moderate in size. Pygidium. In lateral view widely convex; surface totally punctate; punctures smaller and denser near anterior margin and sides, sparser and larger to apex; apex with marginal bead.
Legs. Protibia with 4 external teeth; basal tooth smaller. Apex of mesotibia with 4 teeth. Apex of metatibia with 5 teeth.
Aedeagus. Shape symmetrical (Fig. 5D), narrowing abruptly at middle; apex inflated, rectangle shaped, truncate; sides subparallel. In lateral view surface concave and with, acute, small projection near lateral base (Fig. 5C).

Dorsal and lateral view of aedeagus. A and B Oryctophileurusarmicollis Prell, 1911 C and D Oryctophileurus varicosus Prell, 1934, scale bar 1 mm.
Oryctophileurus varicosus was described by
The records cited here are the only known specimens.
http://species-id.net/wiki/Oryctophileurus_armicollis
Figs 5A–B; 6; 7A, CPeru: holotype, male, body length 18.5 mm (ZMHB).
Bolivia: La Paz dep., Nor Yungas prov., Caranavi, I-2003, 2 males with body length 18 mm and 20 mm and 1 female with body length 18 mm (EPGC); Route Coroico-Caranavi, XII-2008, 1 male (EPGC); La Paz dep., Calisaia, V-1925, G.L. Harrington col., 1 male (MZSP); Beni dep., Cosincho, VIII-1925, G.L. Harrington col., 1 female (MZSP).
Color black, strongly shining. Head anteriorly flat, not concave; canthus subquadrate, in some specimens weakly projected forward; horn in males strongly recurved, on about the same level or slightly higher than posterior pronotal protuberance. Mentum with longitudinal furrow flat and wide, as well as posterior margin. Distance between inner teeth of dorsal pronotal protuberance in male wider that width between eyes (Fig. 6A); in females separated by a concavity with occelate punctures; pronotal concavity accounts for about 40–50 % of dorsal pronotal surface in males. Ocellate punctures above posterior-lateral pronotal margin continuous (Fig. 7C). Each elytron with 10 striae; punctures ocellate, sometimes coalescent and elongated, decreasing in size to sides; interstriae smooth, convex. Striae and interstitial punctures arranged in regular lines Mesotibia with 3 to 5 apical teeth; metatibia with 4 to 6 apical teeth. Pygidium in males distinctly more convex than in females, moderately punctate, denser at base and sides. Aedeagus symmetric (Fig. 5A); parameres with apex rounded to subtriangle shaped (Fig. 5B).

Dorsal and lateral habitus of Oryctophileurusarmicollis Prell, 1911, A and B holotype male, body length 18.5 mm C and D female, body length 18 mm, scale bar 5 mm.
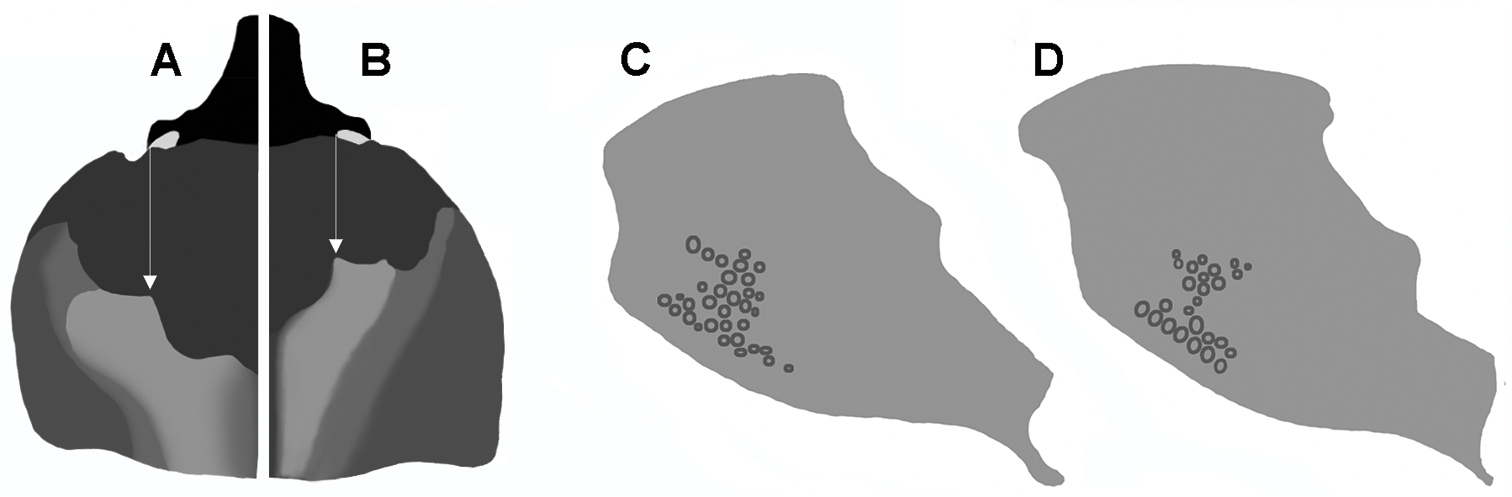
Pronotum of males with body length 20 mm, dorsal, higher areas light, inner teeth of transversal pronotal carina indicated by white arrow: A Oryctophileurus armicollis Prell B Oryctophileurus guerrai; ocellate punctures at posteriolateral pronotal surface, character state does not differ between sexes C Oryctophileurus armicollis D Oryctophileurus guerrai.
Oryctophileurus armicollis is known from Peru (
http://zoobank.org/71F1D594-7DC9-41E0-93DE-704DEDE75470
http://species-id.net/wiki/Oryctophileurus_guerrai
Figs 7B, D; 8Holotype: male, “Bolivia / Tarija / O’Connor province / Tariquía National Reserve / S21°59'01, W64°12'30 / 1008 m a.s.l. / Tucuman-Bolivian subhumid forest / gully close to small mountain river / 25-XI-2011 / R. Perger leg.” Allotype: female, same location data as the holotype, 20-XI-2011, F. Guerra leg.
Oryctophileurus guerrai sp. n. is distinguished from the morphologically similar Oryctophileurus armicollis by the distance between the inner teeth on the dorsal pronotal protuberance (in males as wide as width between eyes) (Figs 7B, 8A) and in females by the inner teeth separated by only a small fissure (Fig. 8C). In smaller males (body length 18.5 mm) of Oryctophileurus armicollis the distance between the inner teeth of the pronotal protuberance is wider than the width between the eyes and in larger males (body length 20 mm) as wide as the distance between the outer eye margins (Fig. 6A). In females of Oryctophileurus armicollis the inner teeth of the pronotal protuberance are separated by a comparably wide, continuous, parallel concavity containing ocellate punctures (Fig. 6C).

Dorsal and lateral habitus of Oryctophileurus guerrai sp. n., A and B holotype male, body length 20 mm C and D allotype female, body length 19 mm, scale bar 5 mm.
In both sexes of Oryctophileurus guerrai the ocellate punctures above the posteriolateral pronotal margin are smaller, shallower and sparser than in Oryctophileurus armicollis and the area of ocellate punctures above posteriolateral pronotal margin is reduced laterally (Figs 7D; 8B, D). In both sexes of other Oryctophileurus speciesthe pronotum posteriolaterally has a continuous area of distinctly developed, ocellate punctures.
In the male Oryctophileurus guerrai the pronotal protuberance is dorsally higher and the cephalic horn longer (Fig. 8B) than in similar-sized Oryctophileurus armicollis (Fig. 6B) and Oryctophileurus nasicornis (Fig. 3B). In Oryctophileurus nasicornis, both characters are less produced. In the male of Oryctophileurus varicosus (Fig. 4B), with its slightly larger body, the pronotum is much flatter than in the male of Oryctophileurus guerrai.
Holotype male (Figs 8A, B). Body length 20 mm, width 9 mm. Body elongate, cylindrical, head and pronotum brownish black dorsally, elytra black, body ventrally dark brown, dorsal surface glabrous. Legs, sterna, second abdominal ventrite and basal margin of pygidium setose.
Head. Surface smooth, finely punctate. Clypeus subtriangular, laterally emarginated, slightly upturned, and with acute apex. Cephalic horn recurving over pronotum, attenuate, apex narrowly rounded; surface at base coarsely punctate, with a lateral carina. Mandible tridentate with inner tooth more acute; teeth upturned. Antenna 10 segmented; club with antennomeres subequal in length.
Pronotum (Figs 7B, D; 8A, B). Shape subtrapezoidal, evenly rounded laterally, smooth, with 2 impressions posteriolaterally; posterior edges slightly obtuse. Basal half smooth, longitudinally ridged, dorsally bulging until transverse median carina, with 2 dorsolateral, coarsely punctate concavities on both sides; transverse median carina with 4 obtuse teeth, 2 inner teeth dorsally with small concavity, distance between inner teeth of dorsal pronotal protuberance as wide as width between eyes; anterior half of pronotum strongly concave, with ocellate punctures, 2 obtuse teeth close to anterior pronotal border, teeth at same level with lateral eye margins when seen from dorsal and lateral views. Prosternal process trapezoidal, concave at the base, posteriorly produced; base with spine-like process posteriorly.
Elytra. Surface smooth, with continuous, slightly convex carinae, weakly impressed interstriae, and with ocellate punctures. Pygidium. Shape convex in lateral view; surface densely punctate; punctures ocellate, moderately sized, elongate near basal margin.
Legs. Protibia with 4 teeth, basal tooth weakly developed. Meta- and mesotibia apically with 3 broaden, shovel-shaped teeth, each tooth additionally furnished with small, apical teeth. The specimen was found dead with damaged abdomen, soft parts and genitalia missing.
Female allotype (Figs 8C, D). Similar to male except by the following features: body length 19 mm, width 8.2 mm; head with cephalic horn less developed, reaching only dorsal pronotal protuberances when seen in lateral view; pronotum longer than high, dorsal longitudinal pronotal concavity about as narrow as width of cephalic horn, uppermost teeth of dorsal pronotal protuberances obtuse, separated by a small fissure.
The species is named after our friend and colleague, Fernando “Fideo” Guerra, for his lifetime commitment to the investigation of the Bolivian fauna. His participation in the actual survey in the southern Bolivian Andes has led to the discovery and description of several previously unknown taxa (e.g.,
Oryctophileurus guerrai is known only from the northwestern area of Tariquía National Reserve (Tarija department) in the southern Bolivian Andes (Fig. 1). The forest in the this area is considered subandine subhumid, semi-deciduous, Tucuman-Bolivian forest (TBF) (
The two individuals of the new species were collected in a narrow valley (elevation 1008 m a.s.l.) (Fig. 1C). The female was observed during the day on the floor of the densely vegetated, moist gully (Figs 1C, D). The male was found dead in a similar habitat. No individuals of the species were observed in subhumid forest along the slopes and during eight nights of sampling with a light trap close to the collection area. Like the other species of this genus, which are only known from few individuals, Oryctophileurus guerrai might be rare or has a cryptic way of life.
As in males of other dynastine taxa with exaggerated secondary sexual traits(e.g., Eberhard 1979;
While the two Amblyodus species occur in Amazon lowland forest (Amblyodus castroi Grossi & Grossi) and Central American mountain forests (Amblyodus taurus Westwood) (
As proposed for other Andean taxa (see
The discovery of Oryctophileurus guerrai extends the known distributional range of the genus more than 600 km southwards. In view of the mainly tropical Andes distribution of Oryctophileurus species, the presence of this genus in the southern Bolivian Andes, close to the Argentinean border, is surprising. Because of a change in orientation of the mountain ranges at the elbow of the Andes and local topographic features, the TBF is distinguished from Bolivian Yungas forest (BYF) by a more pronounced and prolonged dry season, occasionally accompanied by frost periods (
Nevertheless, the discovery of Oryctophileurus guerrai and other endemic TBF representatives of butterfly genera (
This hypothesis should be tested in further studies because human impact and low protection status of such ecoregion (see
We thank Fernando Guerra (Instituto de Ecología, Universidad Mayor de San Andrés, La Paz, Bolivia) for his support during the field trips. RP thanks Gonzalo Romero, Jairo Acosta, and Lorena Quintana (GeokineticsSrl Bolivia) for making available the logistic and financial support for the biodiversity study in Tariquía. The study was also made possible with the generous help of Saul Chavez Orosco (Director SERNAP), Julio Cesar Guerrero Ruíz (Director Tariquía), Fernando Villarte (SERNAP), Lic. Julieta Vargas (Colección Boliviana de Fauna, La Paz, Bolivia), and the park rangers of Tariquía.
PCG thanks Everardo J. Grossi for logistical support and Dr. Roberto A. Cambra T. (Museo de Invertebrados “G. B. Fairchild”, Universidad de Panamá) for the loan of the specimen of Amblyodus taurus and Dr. Nayr Aguiar (Universidade Federal do Amazonas) for the specimen of Amblyodus castroi illustrated here.
Dr. Olaf Jäger (Senckenberg Naturhistorische Sammlungen Dresden, Germany) is thanked for helping to locate the type material of Oryctophileurus armicollis and Dr. Joachim Willers and Dr. Johannes Frisch (Museum für Naturkunde, Berlin, Germany) for providing valuable images.
We are also grateful to Dr. J. Mark Rowland (University of New Mexico), Dr. Brett C. Ratcliffe (University of Nebraska State Museum), Dr. Andrey Frolov (Zoological Institute, St.Petersburg, Russia), Manuel Mena and the anonymous reviewer for commenting on an earlier draft of this article.
Open access to this paper was supported by the Encyclopedia of Life (EOL) Open Access Support Project (EOASP).