





(C) 2013 Chongda Wang. This is an open access article distributed under the terms of the Creative Commons Attribution License 3.0 (CC-BY), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
For reference, use of the paginated PDF or printed version of this article is recommended.
Citation: Wang C, Ren D (2013) Nuurcala obesa sp. n. (Blattida, Caloblattinidae) from the Lower Cretaceous Yixian Formation in Liaoning Province, China. ZooKeys 318: 35–46. doi: 10.3897/zookeys.318.5514
Nuurcala obesa sp. n., in the cockroach family Caloblattinidae, is described from the Lower Cretaceous Yixian Formation (Liaoning Province, China) based on a combination of differential characters of fore- and hind wing venation, colouration and body structures. Systematically, Nuurcala (Vršanský, 2003), known from the Cretaceous sediments of Bon Tsagaan and Khurilt, Mongolia, is closely related to other genera of Caloblattinidae known from the Jurassic and Cretaceous localities in other parts of Asia.
Fossil cockroach, new species, Huangbanjigou, wing venation, colouration
Caloblattinidae, a large extinct family of cockroaches, was erected by Vršanský and Ansorge, it consists of over 50 genera comprising nearly 200 described species distributed in Argentina, Australia, Brazil, Burma, China, England, France, Germany, Greenland, Japan, Kazakhstan, Kirgizstan, Mongolia, Russia, Spain, Switzerland, and South Africa during Middle Triassic to Late Cretaceous (
Caloblattinidae was the dominant family from Upper Jurassic in Karatau, South Kazakhstan (
The strata of the Yixian Formation represent mainly lacustrine sediments intercalated with volcaniclastics, which contains a large number of Jehol Biota fossils, such as well-preserved dinosaurs, primitive birds, early mammals, fishes, ostracods, plants and abundant insects (
The four specimens were collected from the Yixian Formation, Huangbanjigou, Chaomidian Village, Beipiao City, Liaoning Province, China. All type specimens are deposited in the fossil insect collection of the Key Laboratory of Insect Evolution & Environmental Changes, Capital Normal University, Beijing, China. They were examined with a Leica MZ 12.5 dissecting microscope and illustrated with the aid of a drawing tube attached to the microscope. Line drawings were made with Photoshop CS 3.0 graphic software. Photographs of fossils were taken by a MZ12.5 dissecting microscope (Leica, Wetzlar, Germany), either dry or with alcohol.
The venation nomenclature used in this paper is based on the interpretation of
Nuurcala popovi Vršanský, 2003
Nuurcala popovi Vršanský, 2003. Bon Tsagaan Nuur, Bed 87/8, Mongolia; Barremian or Aptian, Early Cretaceous.
Nuurcala srneci Vršanský, 2008. Khurilt, Bed 210/24, Mongolia, Barremian or Aptian, Early Cretaceous.
Nuurcala sp. (collected by expedition of PIN led by M.B. Mostovski and P. Vršanský) Baissa, Transbaikalian Russia, ?Valanginian, Early Cretaceous.
Nuurcala sp. (collected by D. Davaadorj, deposited in the PCMAS, undescribed) Erdenyi Ula, Mongolia, Early Cretaceous.
?uppermost Jurassic; Lower - Upper Cretaceous; Asia.
urn:lsid:zoobank.org:act:1009795B-D0EA-41D4-A356-B3FEA7515871
http://species-id.net/wiki/Nuurcala_obesa
Figs 1–4Differs from Nuurcala popovi and Nuurcala srneci in having relatively small head, and antennal sockets conspicuous at sides, antennae long and thick; forewing with dark colouration except for R area, a dark maculae present at the edge of Sc area; hindwing with dark colouration; Legs with dark colouration, femora thick, tarsus with five segments and a claw (Fig. 1).
Holotype (Fig. 1): body medium to large size, length about 23.8 mm as preserved, total estimated length is 25.2 mm, and width 9.9 mm; Abdomen with 6–7 visible segments. head small, length 1.8 mm, and width 2.5 mm, antennal sockets conspicuous at sides. Pronotum, shield-like, vaulted, transversal, simple symmetrical zonal colouration at the margin, length 6.2 mm, width 6.9 mm. Forewings: length 22 mm, width 6 mm; one dark maculae present at the edge of Sc area, dark colouration with pale area in R; 55 veins at margin; intercalaries thick, all over wing surface; Sc 3 branches, shorter than clavus; R sigmoidal 15 branches, with undifferentiated Rs, reaching the anterior wing margin; M slightly curved with 9 branches, most posterior branches of M reaching wing apex; CuA slightly curved to posterior wing margin and 10 branches; CuP curved and simple; clavus long, more than a third of the wing’s length; A with tertiary branches. Hind wing (17 mm long as preserved vs. 22 mm long for forewing) with branched Sc; both R1 and RS about 9 veins; M 4 branches; Cu (±9) with additional blind branches that may reticulate, fan-like pleating present visible on forewing. Legs with dark colouration, femora thick, about 2 times as long as tibia, tarsus with five segments and a claw, spines obscure.
Paratypes (Figs 2–4): only forewing preserved, length range about 11.5–19 mm, width range about 5.2–5.7 mm; 42–48 veins at margin; Sc 3-5 branches, R sigmoidal 12–15 branches, M slightly curved and 4–9 branches, CuA 7–10 branches.
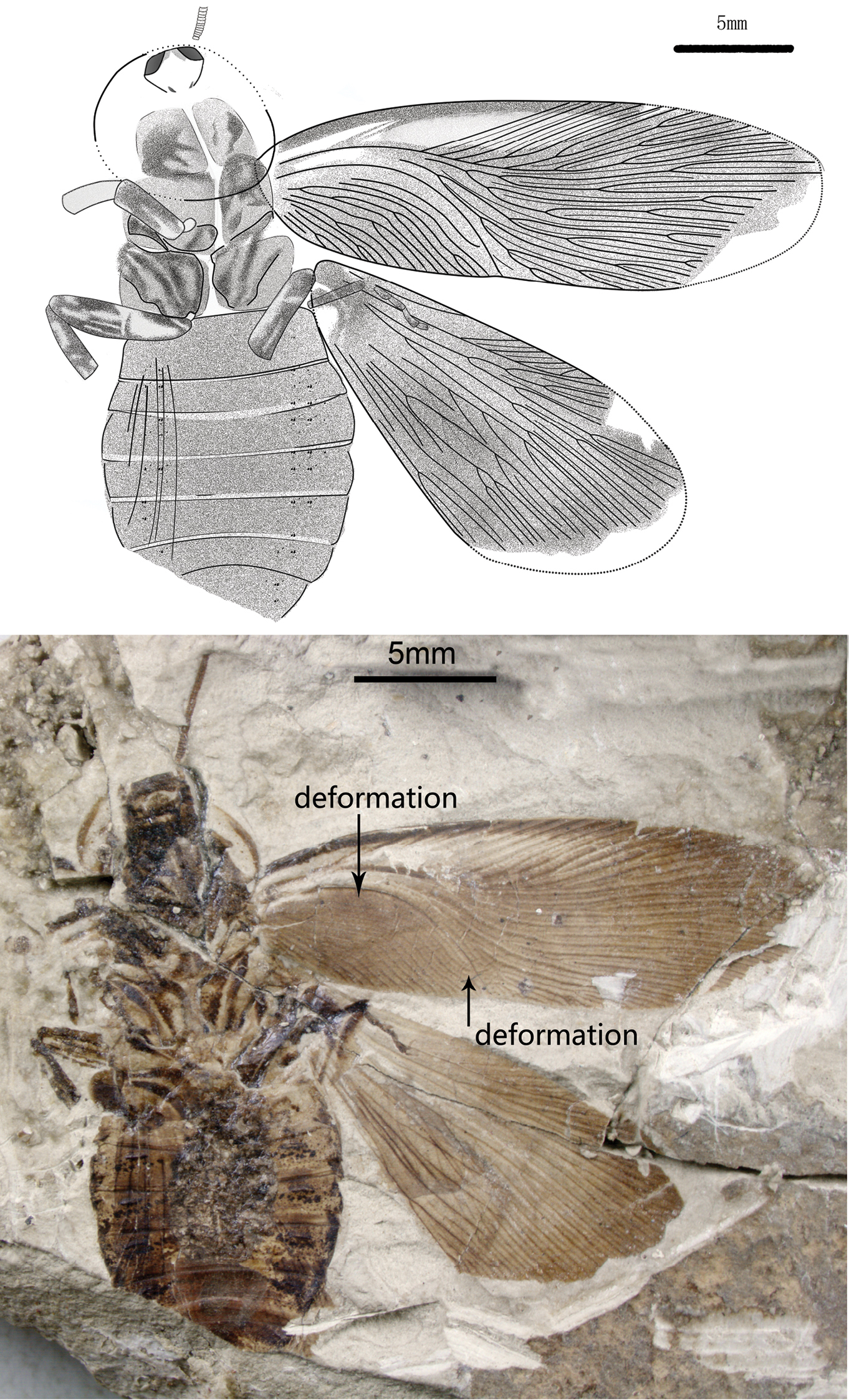
Nuurcala obesa, sp. n. Holotype, CNU-BLA-NN-2012055 A Line drawing B photograph. Scale bars = 5 mm.

Nuurcala obesa, sp. n. Paratype, CNU-BLA-NN-2012056 A Line drawing B photograph. Scale bars = 2 mm.

Nuurcala obesa, sp. n. Paratype, CNU-BLA-NN-2012057 A Line drawing B photograph. Scale bars = 2 mm.

Nuurcala obesa, sp. n. Paratype, CNU-BLA-NN-2012058; photograph. Scale bar = 2 mm.
Holotype, partially preserved specimen with both fore- and hind wings and a body, CNU-BLA-NN-2012055 (Fig. 1). Paratypes (three isolated forewings): CNU-BLA-NN-2012056 (Fig. 2), CNU-BLA-NN-2011057 (Fig. 3), CNU-BLA-NN-2011058 (Fig. 4).
Yixian Formation; Early Cretaceous; Huangbanjigou, Chaomidian Village, near Beipiao City, Liaoning Province, China.
The specific name is derived from the Latin word “obesus”, (meaning “fat”), refer to the relatively large abdomen.
We consider the present taxon belonging to the genus Nuurcala based on the following features: Body medium to large size, both wings have dark markings, and forewing with characteristic colouration pattern. The four specimens vary in size significantly (wing length from 11.5 to 22 mm), and the size range of this species supports that Caloblattinidae are highly variable in size (
Nuurcala obesa sp. n. is similar to Nuurcala popovi Vršanský, 2003 in the following aspects: head hypognathous, pronotum transverse ovoid, and forewing with distinct cubital space, rather wide, but differs from Nuurcala popovi Vršanský, 2003 by forewings with subparallel margins and characteristic markings (dark markings with pale area in R), Sc branched, R rich, M branched, Cu veins ending prior to the apex of the wing, A branched, CuA almost straight, and then curved to posterior wing margin, anal area wide.
Nuurcala obesa sp. n. differs from Nuurcala srneci Vršanský, 2008 by the latter having much bigger head and the veins less numerous than Nuurcala obesa. The new speciesdiffers from Nuurcala sp. (PCMAS) from Erdenyi Ula, Mongolia in having a bigger pronotum (in contrast to 5.8/6.2 mm; forewing length 21 mm).
The number of forewing veins in Nuurcala obesa sp. n. is differs from those of previously reported Nuurcala popovi and Nuurcala srneci. For comparison, the data are listed in Table 1. The total number of forewing veins of Nuurcala obesa (42–55 veins) is higher than that of Nuurcala srneci (about 44 veins), but much less than that of Nuurcala popovi (54–83 veins). The number of forewing M veins of Nuurcala obesa (4–9 veins) is lower than those in Nuurcala popovi (8–17 veins) and Nuurcala srneci (6–11). The differences of venation further justify the erection of Nuurcala obesa sp. n.
Variability of number of veins in forewings for three species of Nuurcala.
| Species | Number of veins in forewings | |||||
| Sc | R | M | Cu | A | Total | |
| Nuurcala obesa | 1–3 | 12–15 | 4–9 | 8–10 | 14–19 | 42–55 |
| Nuurcala popovi | 3–5 | 16–25 | 8–17 | 12–17 | 15–21 | 54–83 |
| Nuurcala srneci | ±3 | 13–16 | 6–11 | 8–14 | 5–7 | ±44 |
There are some taxa placed in other genera which show affinities to the Nuurcala. Nuurcala obesa sp. n. differs from Rhipidoblattina hebeiensis Hong, 1980 from the Middle Jurassic Jiulongshan Formation (
Yixian Formation is characterized by a high proportion of coloured species (
The family Caloblattinidae, starting with its first occurrence in the Middle Triassic, was important in almost all known ecosystems and dominated from the earliest Jurassic up to the latest Early Cretaceous (
We sincerely thank Dr. Peter Vršanský (GlU SAV, Bratislava) and Dr. ChungKun Shih for revision of the manuscript and are grateful to Dr. Yunzhi Yao, Dandan Wei, Qiang Yang, Taiping Gao, Qiuping Dong, Haijing Hu and other students of our laboratory for their kind help. This research is supported by the National Basic Research Program of China (973 Program) (grant 2012CB821906), the National Natural Science Foundation of China (No. 31230065, 31272352, 41272006), Project of Great Wall Scholar and KEY project of Beijing Municipal Commission of Education (grants KZ201310028033).