





(C) 2013 Xiao Qiao. This is an open access article distributed under the terms of the Creative Commons Attribution License 3.0 (CC-BY), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
For reference, use of the paginated PDF or printed version of this article is recommended.
Citation: Qiao X, Shih CK, Petrulevičius JF, Ren Dong R (2013) Fossils from the Middle Jurassic of China shed light on morphology of Choristopsychidae (Insecta, Mecoptera). ZooKeys 318: 91–111. doi: 10.3897/zookeys.318.5226
Choristopsychidae, established by Martynov in 1937 with a single isolated forewing, is a little known extinct family in Mecoptera. Since then, no new members of this enigmatic family have been described. Based on 23 well-preserved specimens with complete body and wings from the Middle Jurassic of northeastern China, we report one new genus and three new species of Choristopsychidae, two new species of the genus Choristopsyche Martynov, 1937: Choristopsyche perfecta sp. n. and Choristopsyche asticta sp. n.; one new species of Paristopsyche gen. n.: Paristopsyche angelineae sp. n.; and re-describe Choristopsyche tenuinervis Martynov, 1937. In addition, we emend the diagnoses of Choristopsychidae and Choristopsyche. Analyzing the forewing length/width ratios of representative species in Mecoptera, we confirm that choristopsychids have the lowest ratio of forewing length/width, meaning broadest forewings. These findings, the first fossil choristopsychids with well-preserved body structure and the first record of Choristopsychidae in China, shed light on the morphology of these taxa and broaden their distribution from Tajikistan to China, while increasing the diversity of Mesozoic Mecoptera in China.
Mecoptera, Choristopsychidae, new genus, new species, Middle Jurassic, China
Choristopsychidae is a rather obscure extinct family in the Order Mecoptera. The family, erected by
Recently, we collected 23 well-preserved fossils from the Daohugou Village, Ningcheng County, Inner Mongolia, China; Jiulongshan Formation, Middle Jurassic. Herein, based on their different morphological characters, we erect one new genus with one new species and two new species of Choristopsyche Martynov, 1937, and re-describe Choristopsyche tenuinervis Martynov, 1937, while emending diagnoses of Choristopsychidae Martynov, 1937 and Choristopsyche Martynov, 1937.
There are abundant well-preserved fossil insects from Daohugou, including 19 reported orders so far (
This study is based on 23 fossil specimens housed in the fossil insect collection of the Key Laboratory of Insect Evolution & Environmental Changes, College of Life Sciences, Capital Normal University, Beijing, China (CNUB; Dong Ren, Curator).
Photographs of whole specimens were taken with a Nikon D100 digital camera coupled to a Nikkor 105 mm macro lens. The specimens were examined using a Leica MZ12.5 dissecting microscope, and illustrated with the aid of a drawing tube attached to the microscope. Line drawings were made with CoreDRAW X4 graphic software.
The wing venation nomenclature used in this paper is based on the interpretations and system proposed by
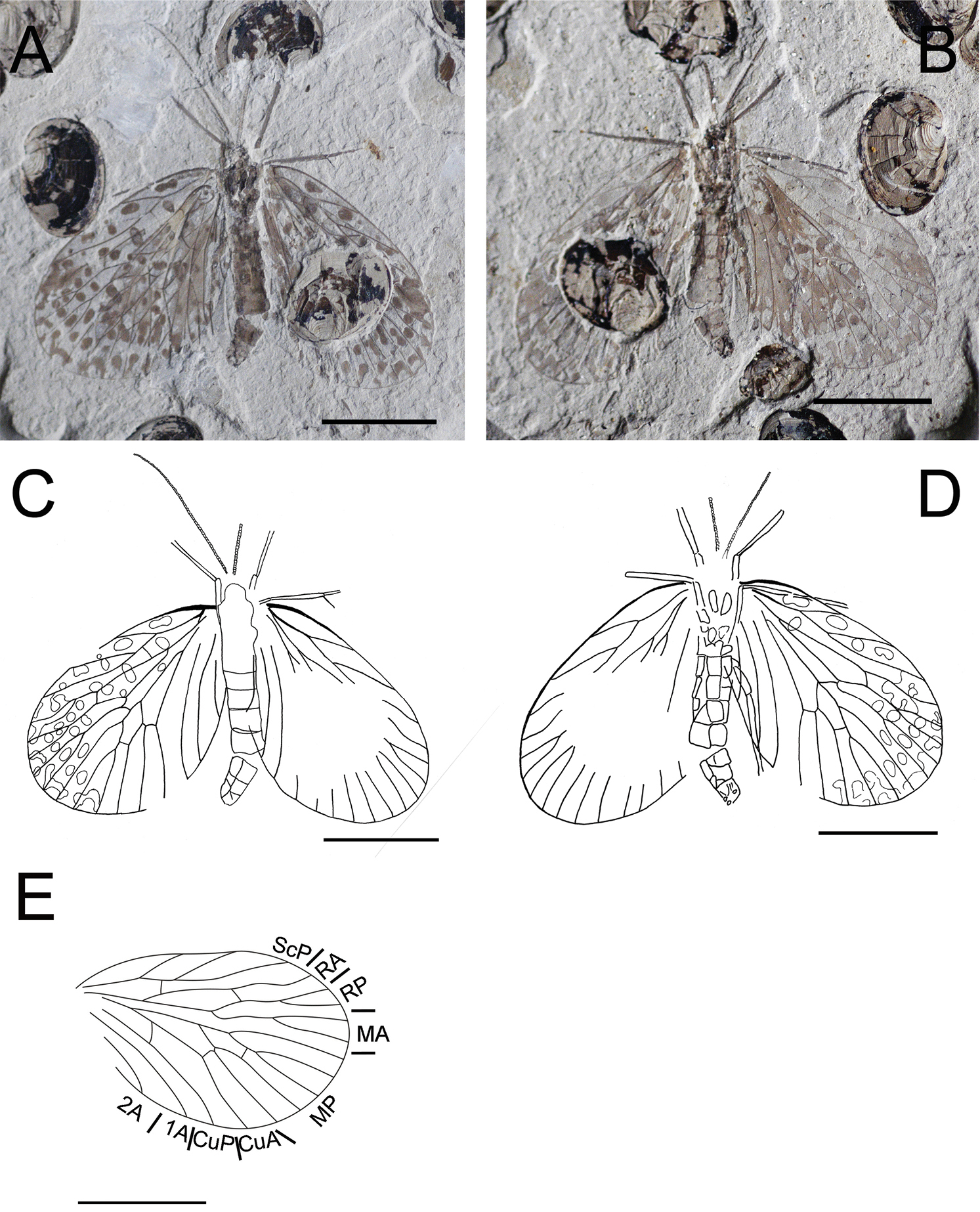
Photographs and line drawings of Choristopsyche tenuinervis Martynov, 1937 A Photograph of part, no. CNU-MEC-NN2011075p B Photograph of counterpart, no. CNU-MEC-NN2011075c C Line drawing of whole specimen of part, no. CNU-MEC-NN2011075p D Line drawing of whole specimen of counterpart, no. CNU-MEC-NN2011075c E Line drawing of forewing of part, no. CNU-MEC-NN2011075p. Scale bars represent 5 mm.
Forewing broad oval or subtriangular, field between C and ScP comparatively broad; ScP well developed and forked twice, forming three long branches; RA unforked, one crossvein between ScP and RA and between RA and RP; RP and MA both with two branches; MP with five branches, and the MP4+5 forking basal to the MP2+3 forking; MP and CuA merged at the base; CuA strongly bent at its mid point; an oblique crossvein between CuA and CuP; a curved crossvein between the midpoint of CuA and MP5; CuP, 1A and 2A almost parallel. Hind wing, similar in shape to the forewing but slightly smaller, ScP short, forked twice, the second bifurcation coalesces with RA for a short distance; RP and MA both with two branches; MP with five branches, the stem of MP4+5 forked earlier than that of forewing, and with a crossvein to CuA; CuA almost straight. Head, oviform with big and oval compound eyes; antennae long and filiform; small chewing mouthpart. Thorax: prothorax smaller than mesothorax and metathorax. Legs: long and slender, all legs nearly of the same shape, but hind legs longer than fore legs and mid legs, and femora wider than tibia, and tibia longer than femora. Abdomen slender, tapering apically, about eleven segments and the female terminal segment with cercus.
Choristopsyche tenuinervis Martynov, 1937 (Lower Jurassic of Tajikistan)
Forewing, the separation of RP+MA from RA distal to the separation of MP from CuA.
Type species (Choristopsyche tenuinervis Martynov, 1937), Choristopsyche perfecta sp. n. and Choristopsyche asticta sp. n.
http://species-id.net/wiki/Choristopsyche_tenuinervis
Figs 1, 2Forewing, RP+MA forking distal to MP forking.
CNU-MEC-NN2011075p/c (Fig. 1), a well preserved specimen with part and counterpart in dorsal view, with almost complete forewings, but partially preserved hind wings and body, and forewings overlapping hind wings. The terminus of abdomen is missing, sex unknown. Wings: Left forewing, length 11.0 mm, width 6.7 mm, broadly oval, RP forking distal to MA forking; MP2+3 forking basal to the forking of MA, and the stem of MP3 about twice as long as the stem of MP2+3; with one crossvein between RA and RP, and between MP2+3 and MP4; CuP, 1A, 2A single. Right forewing is similar to left forewing, but parts missing. Hind wings, smaller than forewings, overlapped by forewings, the venation visible but unclear. There are many spots on all four wings, symmetric between left and right wings.
In addition, there are nine new materials with analogous wing venation to that of specimen CNU-MEC-NN2011075p/c. They are listed as follows.
CNU-MEC-NN2011080 (Figs 2A, C), a well preserved specimen with clear wings, but parts of body, and the right forewing overlapping the right hind wing. Sex unknown. Wing: Right forewing, length 11.8 mm, width 6.7 mm, RP forking distal to the forking of MA; MP2+3 forking at about the same level as the forking of MA; the stem of MP3 about twice as long as the stem of MP2+3; with one crossvein between MP2+3 and MP4; CuP, 1A, 2A single. Left forewing is similar to right forewing, but the apex of the wing absent. Hind wings, length at about 10.1 mm, width 6.2 mm, similar to forewings but smaller. CNU-MEC-NN2009317 (Figs 2B, D), an almost complete specimen, female, with forewings overlapping hind wings, and nearly complete body, but legs absent in dorsal view. Wings: right forewing, length at about 9.5 mm, width 4.9 mm; RP forking distal to the forking of MA; MP2+3 forking at about the same level to the forking of MA; the stem of MP3 as long as the stem of MP2+3; CuP, 1A, 2A single. Left forewing is similar to right forewing. Hind wings, similar to forewings, but smaller; CuA almost straight. CNU-MEC-NN2009414 (Figs 2E, F), an almost complete preserved specimen in lateral view, female, with complete body and forewings, and right forewing overlapped with body and parts of left forewing, and right hind wing overlapped with left hind wing. The mouthparts are missing, the maxillary palpus with five segments visible. Abdomen: tapering apically, with eleven segments, and a pair of cercus can be visible, female. Wings: Left forewing, length at about 10.1 mm, width 6.4 mm, broadly oval, RP forking basal to the forking of MA; MP2+3 forking significantly basal to the forking of MA; the stem of MP3 about three times as long as the stem of MP2+3; with one crossvein between RA and RP, and between MA and MP1; CuP, 1A, 2A single. Right forewing is similar to left forewing, but there are crossveins between ScP and RA, RA and RP, MA and MP1. Hind wing: similar to forewing but smaller, RA straight, with one crossvein to RP. CNU-MEC-NN2009318 (Fig. 2G), a partially preserved specimen in dorsal view, with parts of forewings, hind wings and body, but the filiform antennae and venation visible, and the forewings overlapping the hind wings. CNU-MEC-NN2011070 (Fig. 2H), an almost completely preserved specimen in ventral view, with almost complete body and four wings, and hind wings overlapped with forewings. CNU-MEC-NN2011071 (Fig. 2I), a partially preserved specimen in dorsal view, with parts of body and forewings, but left hind wing is missing, and right hind wing is obscure. CNU-MEC-NN2009383 (Fig. 2J), a partially preserved specimen with four outspread wings and parts of body. CNU-MEC-NN2011083 (Fig. 2K), a specimen in dorsal view, female, with almost complete body but some legs not visible due to coverage by wings, and forewings overlapping hind wings. CNU-MEC-NN2011085 (Fig. 2L), a comparatively complete specimen in lateral view, male, and forewings overlapping hind wings; chewing mouthparts visible; Abdomen almost completely-preserved, the posterior six segments can be seen clearly, and abdomen bent at six and seven segment, but the posterior segments faint below the left forewing, the scorpion-like terminal visible.
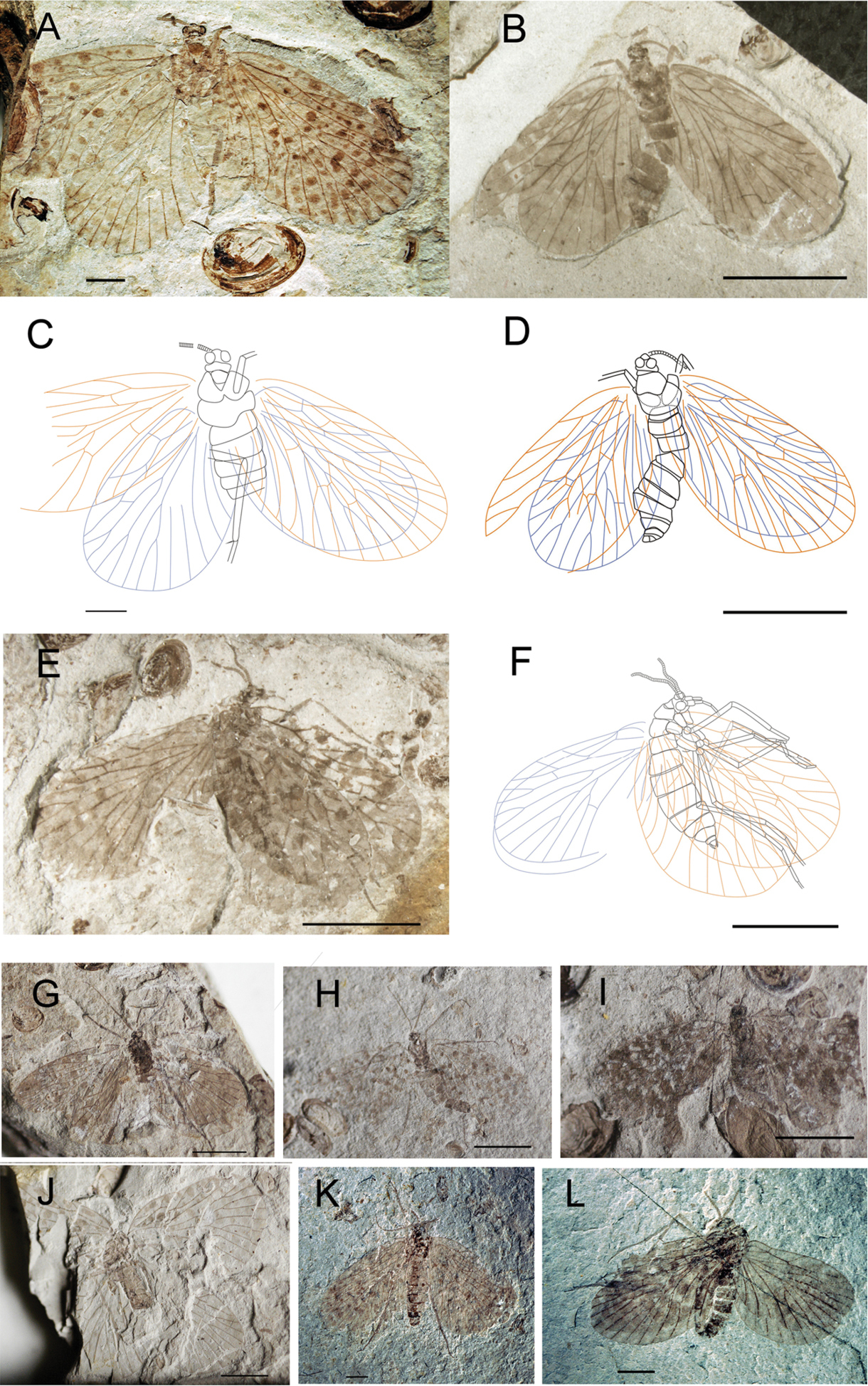
Photographs and line drawings of Choristopsyche tenuinervis Martynov, 1937 A Photograph of no. CNU-MEC-NN2011080 B Photograph of no. CNU-MEC-NN2009317 C Line drawing of no. CNU-MEC-NN2011080D Line drawing of no. CNU-MEC-NN2009317 E Photograph of no. CNU-MEC-NN2009414 F Line drawing of no. CNU-MEC-NN2009414 G Photograph of no. CNU-MEC-NN2009318 H Photograph of no. CNU-MEC-NN2011070 I Photograph of no. CNU-MEC-NN2011071 J Photograph of no. CNU-MEC-NN2009383 K Photograph of no. CNU-MEC-NN2011083 L Photograph of no. CNU-MEC-NN2011085. Scale bars ofA, C, K, L represent 2 mm. Scale bars of B, D, E, F, G, H, I, J represent 5 mm.
CNU-MEC-NN2011075p/c, CNU-MEC-NN2011080, CNU-MEC-NN2009317, CNU-MEC-NN2009414, CNU-MEC-NN2009318, CNU-MEC-NN2011070, CNU-MEC-NN2011071, CNU-MEC-NN2009383, CNU-MEC-NN2011083, CNU-MEC-NN2011085, deposited in CNUB.
Daohugou Village, Ningcheng County, Inner Mongolia, China, Jiulongshan Formation, Middle Jurassic (Bathonian-Callovian boundary interval, ca 164–165 Ma).
These ten specimens exhibit differences in the characters of “RP forking vs. MA forking”, “MP2+3 forking vs. MA forking” and “Length ratio of the stem of MP3 and the stem of MP2+3”, which are considered as intraspecific variations.
urn:lsid:zoobank.org:act:FA6A6E51-B3BD-459E-AED4-021539147BA3
http://species-id.net/wiki/Choristopsyche_perfecta
Fig. 3Forewing, RP+MA forking almost at the same level to MP forking.
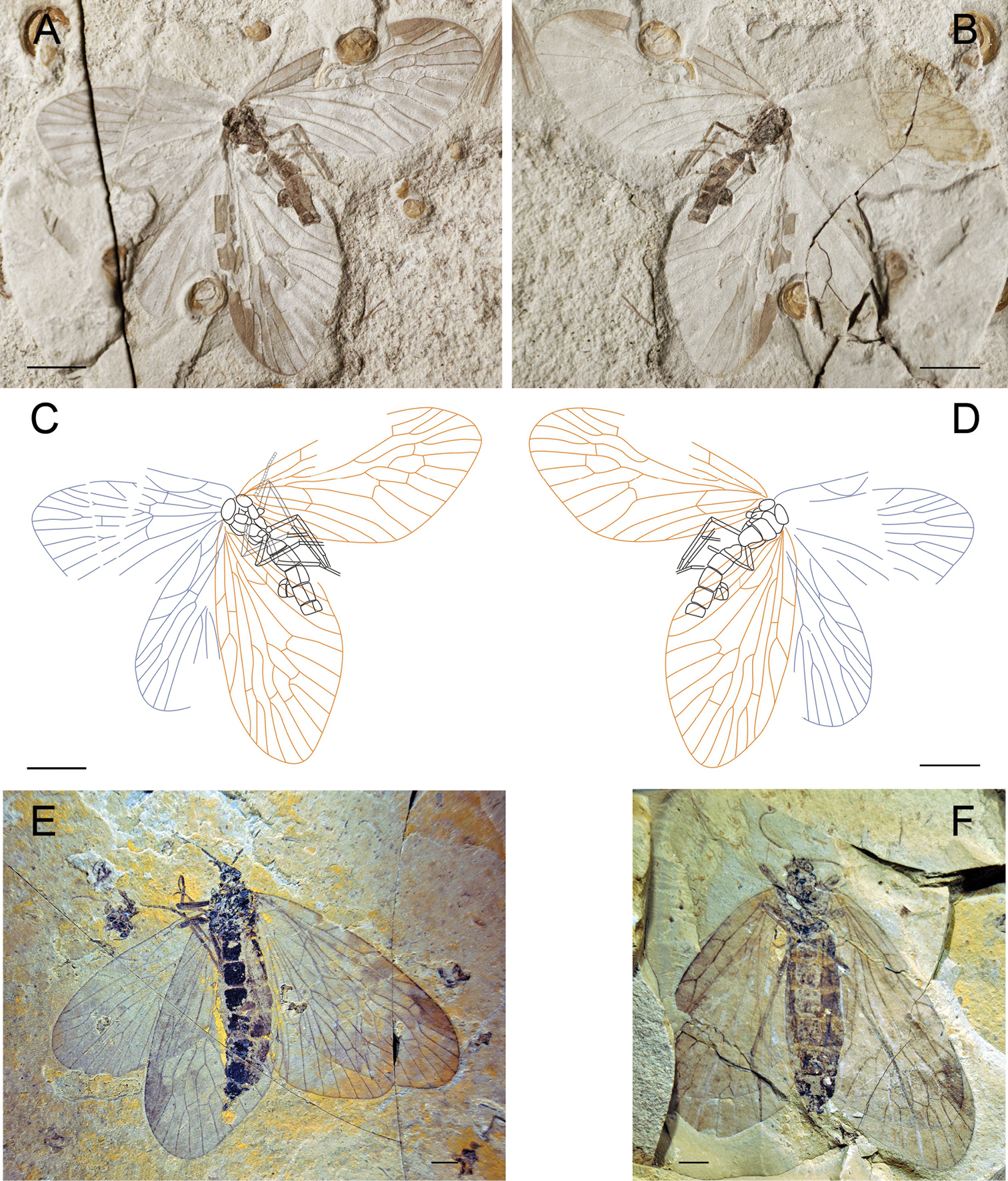
HolotypeCNU-MEC-NN2011082 (Figs 3A, B), an almost complete preserved specimen, maybe male, with four outspread wings, but with partially preserved body. Head is partially preserved, only with one compound eye visible. Thorax: prothorax smaller than mesothorax and metathorax in ventral view. Legs: some parts of fore legs and left hind leg visible. Abdomen: tapering apically, with six segments visible, but the terminal visible, maybe male. Wings: Right forewing, length 22.2 mm, width at about 11.4 mm, RP forking distal to the forking of MA; MP2+3 forking distal to the forking of MA; the stem of MP3 about twice as long as the stem of MP2+3; with one crossvein between CuP and 1A; CuP, 1A single. Left forewing is partially preserved, similar to right forewing. Hind wings, length 18.4 mm, width at about 10.3 mm, similar to forewings but smaller than forewings, and left hind wing partially preserved. There are many spots on all four wings, symmetric between left and right wings.

Photographs and line drawings of Choristopsyche perfecta sp. n. A Photograph of holotype, no. CNU-MEC-NN2011082 B Line drawing of holotype, no. CNU-MEC-NN2011082 C Photograph of paratype, no. CNU-MEC-NN2009352 D Photograph of paratype, no. CNU-MEC-NN2011079 E Line drawing of paratype, no. CNU-MEC-NN2009352 F Photograph of paratype, no. CNU-MEC-NN2011084. Scale bars of A, B, D, F represent 2 mm; scale bars of C, E represent 5 mm.
Paratypes: CNU-MEC-NN2009352 (Figs 3C, E), an almost complete preserved specimen, with clear wing venation and structure of body in side pressure, and forewings partially overlapped with hind wings, sex unknown. Wings: Left forewing, length 18.8 mm, width 10.0 mm, RP forking slightly distal to the forking of MA; MP2+3 forking at about the same level to the forking of MA; the stem of MP3 about twice as long as the stem of MP2+3; with one crossvein between MP1 and MA4, MP1 and MP2, MP2+3 and MP4; CuP, 1A, 2A, 3A single, and one crossvein between CuP and 1A. Right forewing is similar to left forewing. Hind wing, similar to forewing, but smaller. CNU-MEC-NN2011079 (Fig. 3D), female, a well-preserved specimen with complete body, and the terminal of abdomen visible, but the end of legs absent, and forewings overlapping hind wings, but parts of wings missing, RP forking distal to the forking of MA; Abdomen: tapering apically, with eleven visible segments, the tenth and eleventh segments smaller, and the eleventh segment with cerci visible. CNU-MEC-NN2011084 (Fig. 3F), a specimen with legs absent, and hind wings overlapped by forewings in dorsal view, sex unknown. Forewing, length 19.4 mm, width 10.0 mm, RP forking at the same level to the forking of MA; MP2+3 forking basal to the forking of MA; the stem of MP3 about twice as long as the stem of MP2+3.
Holotype CNU-MEC-NN2011082, Paratypes CNU-MEC-NN2009352, CNU-MEC-NN2011079, CNU-MEC-NN2011084, deposited in CNUB.
Daohugou Village, Ningcheng County, Inner Mongolia, China, Jiulongshan Formation, Middle Jurassic (Bathonian-Callovian boundary interval, ca 164–165 Ma).
The name is derived from the Latin word of perfectus, meaning “complete”.
These four specimens exhibit differences in the character of “RP forking vs. MA forking”, “MP2+3 forking vs. MA forking”, which is considered as intraspecific variations.
urn:lsid:zoobank.org:act:31DA887F-2987-4DAD-9520-5776FA0CD5CF
http://species-id.net/wiki/Choristopsyche_asticta
Fig. 4Forewing, RP+MA forking basal to MP forking.
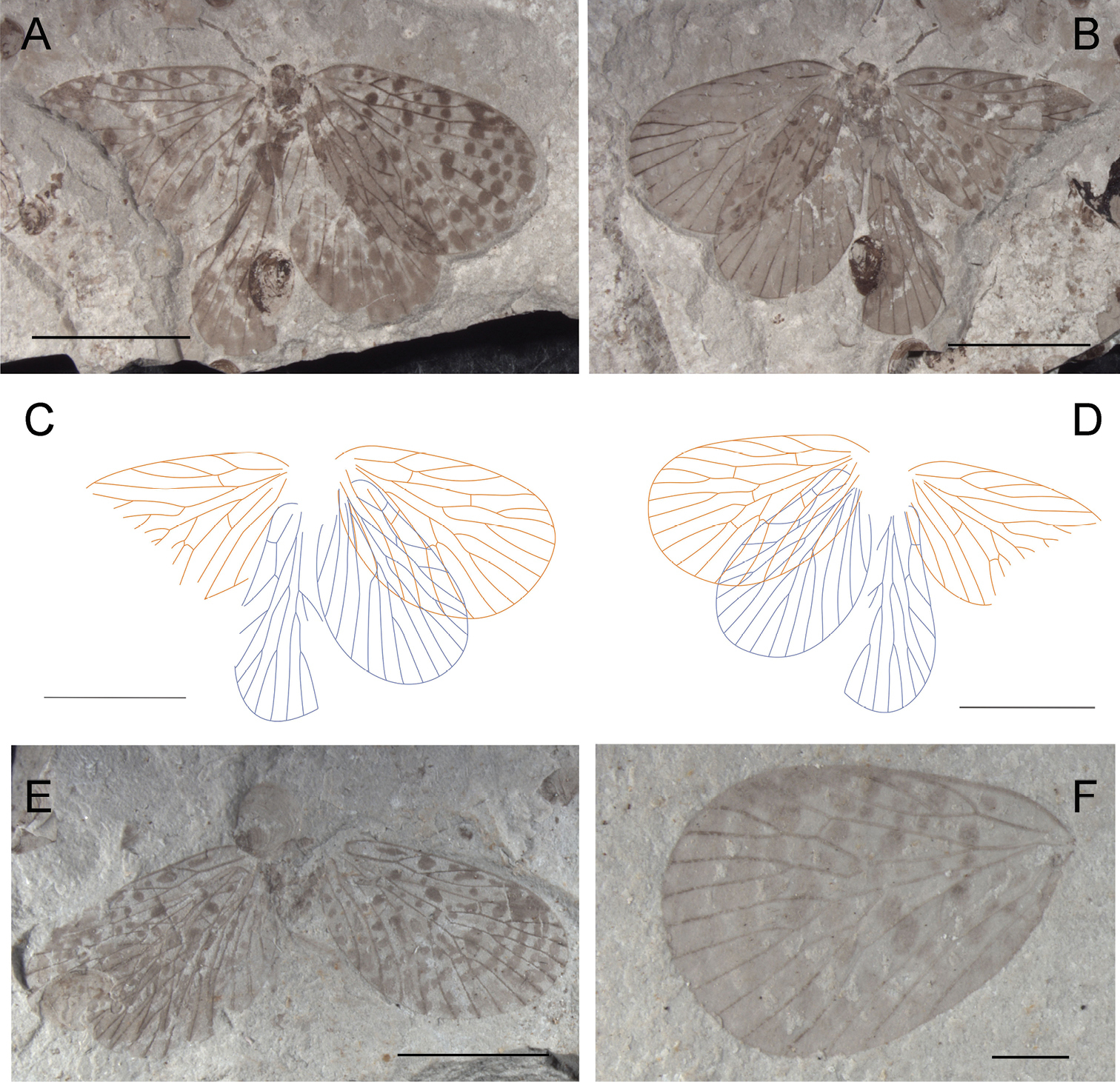
Holotype, CNU-MEC-NN2009394p/c (Figs 4A–D), an almost complete specimen, female, with well-preserved four outspread wings. Head: compound eyes are big and oval in ventral view, but mouthparts invisible and antenna partially preserved. Thorax: prothorax smaller than mesothorax and metathorax, visible in ventral view. Legs: all legs nearly the same shape and nearly completely preserved, long and slender in ventral view. Abdomen: slender and elongate, tapering apically, with eight visible segments. Wings: four wings are elongated and broad, with rounded apical margin. Forewings: Right forewing, length 20.7 mm, width 10.2 mm, almost triangular, dark color between C and RA; RP forking basal to the forking of MA; MP2+3 forking at about the same level to the forking of MA; the stem of MP3 about twice as long as the stem of MP2+3; the stem of MA strongly bent posteriorly; and the stem of MP2+3 strongly bent at its basal one third part; with one crossvein between MA and MP1, MP1 and MP2, and between MP2+3 and MP4; and one oblique crossvein between the base of CuA and CuP, and at the point one oblique crossvein between CuP and 1A, CuP, 1A, 2A single. Left forewing is similar to right forewing. Hind wings: similar to forewing, but slightly smaller, length at about 16.8 mm, width at about 10.0 mm; Right hind wing, with one crossvein between RA and RP+MA. Left hind wing is similar to right hind wing. No spots on entire wings.
Photographs and line drawings of Choristopsyche asticta sp. n. A Photograph of part of holotype, no. CNU-MEC-NN2009394p B Photograph of counterpart of holotype, no. CNU-MEC-NN2009394c C Line drawing of part of holotype, no. CNU-MEC-NN2009394p D Line drawing of counterpart of holotype, no. CNU-MEC-NN2009394c E Photograph of paratype, no. CNU-MEC-NN2011081 F Photograph of paratype, no. CNU-MEC-NN2011086. Scale bars of A–D represent 5 mm. Scale bars of E–F represent 2 mm.
Paratypes: CNU-MEC-NN2011081 (Fig. 4E), female, a well-preserved specimen, with complete body and wings; Head, oval with two big compound eyes, filiform antenna and chewing mouthparts; Abdomen, slender and elongate in lateral view, tapering apically and complete preserved, length at about 13.4 mm; Wings, forewings overlapping some parts of hind wings, and with clear venation; Forewing, length 19.8 mm, width 8.4 mm, RP forking basal to the forking of MA; MP2+3 forking at the same level to the forking of MA; the stem of MP3 about twice as long as the stem of MP2+3. Hind wing, length at about 16.6 mm, width 8.1 mm. CNU-MEC-NN2011086 (Fig. 4F), female, an fairly well-preserved specimen in dorsal view, forewings overlapping hind wings, with almost complete body; Forewing, length 20.8 mm, width at about 9.4 mm; RP forking basal to the forking of MA; MP2+3 forking at the same level to the forking of MA; the stem of MP3 about twice as long as the stem of MP2+3. Hind wing, length 17.3 mm, width at about 9.0 mm.
Holotype CNU-MEC-NN2009394p/c, Paratypes CNU-MEC-NN2011081, CNU-MEC-NN2011086, deposited in CNUB.
Daohugou Village, Ningcheng County, Inner Mongolia, China, Jiulongshan Formation, Middle Jurassic (Bathonian-Callovian boundary interval, ca 164–165 Ma).
The name is derived from the Latin word of astictus, meaning “no spots”.
urn:lsid:zoobank.org:act:B960BA0B-429E-4BCE-96D5-D59BDB7CC263
Paristopsyche angelineae sp. n..
Forewing, the separation of RP+MA from RA at about the same level as the separation of MP from CuA.
Type species: Paristopsyche angelineae sp. n..
The name is derived from the Greek word of paris-, meaning “equal”, and psyche, from the Greek, meaning “soul” or “mind”. The gender is feminine.
urn:lsid:zoobank.org:act:6C1AA33E-D1B1-4D52-9B7F-240CFA9FB1EB
http://species-id.net/wiki/Paristopsyche_angelineae
Figs 5, 6Forewing, RP+MA forking distal to MP forking.
Holotype, CNU-MEC-NN2011076p/c (Figs 5A–D), a well preserved specimen in dorsal view, with outspread clear wings, but parts of body visible. Some segments of head, thorax, legs visible, but faint. Wings: Right forewing, length 8.4 mm, width 5.5 mm, broadly oval with clear venation, RP forking distal to MA forking; MP2+3 forking at about the same level of the forking of MA; the stem of MP3 about three times as long as the stem of MP2+3; with one crossvein between MA and MP1+2+3, and between MP2+3 and MP4. Left forewing is similar to right forewing, but the apex of the wing absent. Right hind wing, length at about 7.5 mm, width 4.6 mm, similar to forewing, but smaller, and overlapping with forewing partially. Left hind wing is similar to left hind wing, but partly folded as preserved. There are many spots on all four wings, symmetric between left and right wings.
Photographs and line drawings of Paristopsyche angelineae gen. et sp. n. A Photograph of part of holotype, no. CNU-MEC-NN2011076p B Photograph of counterpart of holotype, no. CNU-MEC-NN2011076c C Line drawing of part of holotype, no. CNU-MEC-NN2011076p D Line drawing of counterpart of holotype, no. CNU-MEC-NN2011076c E Photograph of paratype, no. CNU-MEC-NN2009319 F Photograph of paratype, no. CNU-MEC-NN2011074. Scale bars of A–E represent 5 mm. Scale bar of F represents 1 mm.
Paratypes: CNU-MEC-NN2009319 (Fig. 5E), a partially preserved specimen with forewings and one hind wing, but body absent; forewing, length 9.0 mm, width 5.4 mm, RP forking distal to MA forking; MP2+3 forking at about the same level of the forking of MA; the stem of MP3 about three times as long as the stem of MP2+3. CNU-MEC-NN2011074 (Fig. 5F), a specimen with one complete and clear forewing, length 7.4 mm, width 5.2 mm, RP forking distal to MA forking; MP2+3 forking at about the same level of the forking of MA; the stem of MP3 about three times as long as the stem of MP2+3. CNU-MEC-NN2011069 (Figs 6A, B), a partially preserved specimen, with complete left wings and most of body except for the terminalia, but incomplete right wings in dorsal view. Wings: right forewing, length 11.2 mm, width 7.5 mm, broadly oval, field between C and RA wide; RP forking distal to MA forking; MP2+3 forking distal to the forking of MA; the stem of MP3 about twice as long as the stem of MP2+3; CuP, 1A, 2A, single. Right hind wing is similar to forewing, but slightly smaller. Left wings partially preserved. CNU-MEC-NN2011078 (Figs 6C, D), an almost preserved specimen in dorsal view, with outspread clear wings, but parts of body absent. Wings: Left forewing, length 10.7 mm, width 6.4 mm, RP forking at the same level of the forking of MA; MP2+3 forking basal to the forking of MA; the stem of MP3 about four times as long as the stem of MP2+3; with one crossvein between MP3 and MP4, and between 1A and 2A; CuP, 1A, 2A single. Right forewing, similar to left forewing, but individual asymmetry is shown by right wing having MP3 with two branches, not the typical one branch in the left wing. Hind wings, length 8.2 mm, width 6.2 mm, similar to forewings but smaller, and partially preserved. CNU-MEC-NN2011077 (Fig. 6E), a partially preserved specimen with complete hind wings but parts of forewings and body; Forewing, length at about 10.3 mm, RP forking at the same level of the forking of MA; MP2+3 forking basal to the forking of MA; the stem of MP3 about three times as long as the stem of MP2+3; Hind wing, length at about 7.9 mm, width 5.0 mm.
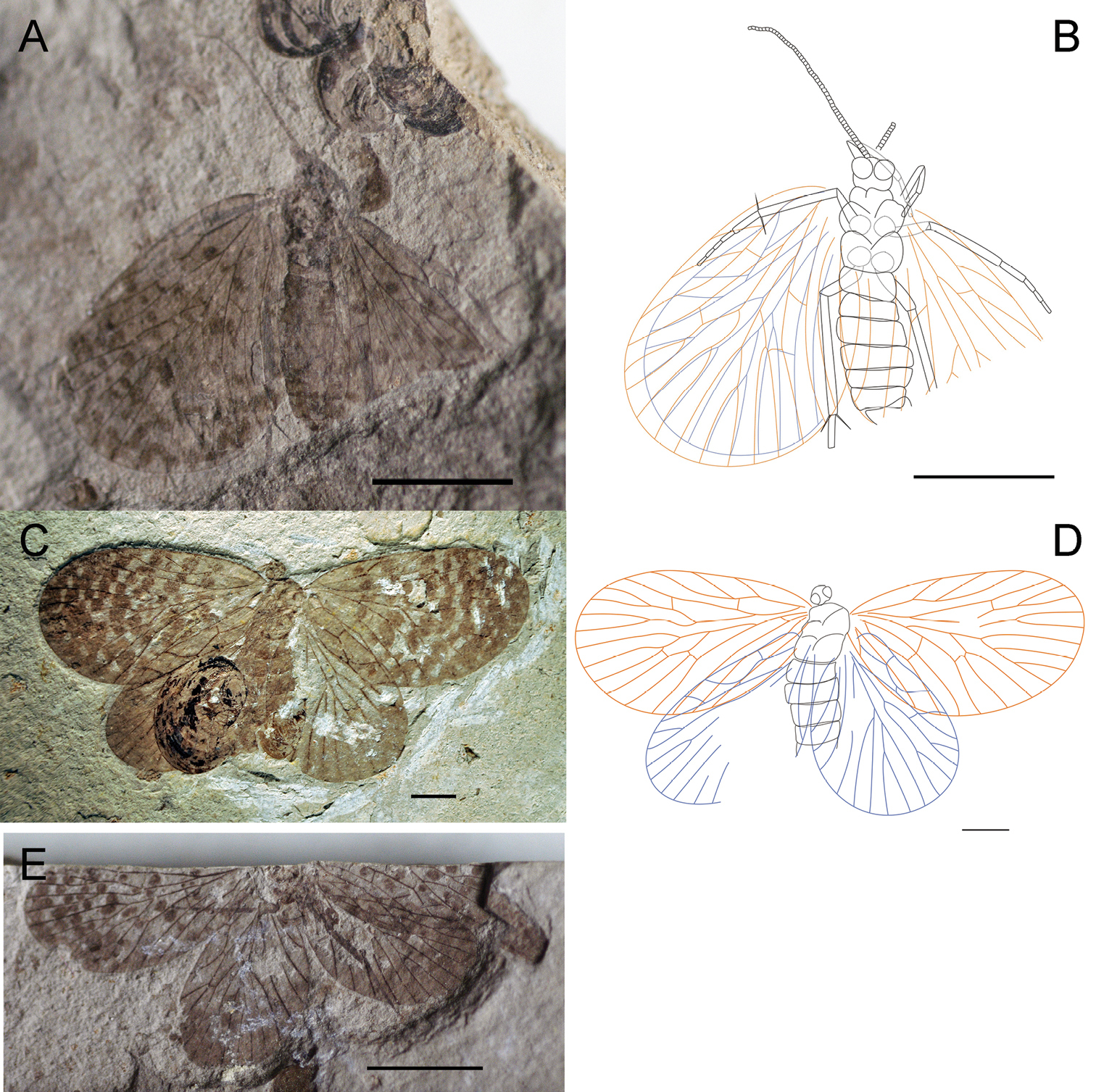
A–B Photographs and line drawings ofparatypesof Paristopsyche angelineae sp. n. A Photograph of paratype, no. CNU-MEC-NN2011069 B Line drawing of paratype, no. CNU-MEC-NN2011069 C Photograph of paratype, no. CNU-MEC-NN2011078 D Line drawing of paratype, no. CNU-MEC-NN2011078 E Photograph of paratype, no. CNU-MEC-NN2011077. Scale bars of A, B, E represent 5 mm. Scale bars of C-D represent 2 mm.
Holotype CNU-MEC-NN2011076p/c, Paratypes CNU-MEC-NN2011078, CNU-MEC-NN2011077, CNU-MEC-NN2011069, CNU-MEC-NN2009319, CNU-MEC-NN2011074, deposited in CNUB.
Daohugou Village, Ningcheng County, Inner Mongolia, China, Jiulongshan Formation, Middle Jurassic (Bathonian-Callovian boundary interval, ca 164–165 Ma).
The specific name is dedicated to Ms. Janet Angeline for her professionalism, dedication and accomplishments in her field and providing inspiration and support to CKS’s palaeontology studies.
These six specimens exhibit differences in the characters of “RP forking vs. MA forking”, “MP2+3 forking vs. MA forking” and “Length ratio of the stem of MP3 and the stem of MP2+3”, which are considered as intraspecific variations.
Based on studies of our specimens and the reported species of Choristopsychidae, we found the wing venation of this family is comparatively stable, that is, the numbers of branches of ScP, RP, MA, MP have almost no changes in these specimens. However we observed significant differences regarding the relative positions between bifurcation points of two different veins, especially the origination locations of the main veins. In the literature of Mecoptera taxonomy, these characters have been used as diagnostic characters for genera or species, such as in Orthophlebiidae, Aneuretopsychidae and so on (
Choristopsychids have relatively broad wings, either oval or subtriangular-shaped, with length/width ratio from 1.5 to 2.0, in contrast to long and narrow wings of most mecopterans. To compare relative forewing broadness for representative mecopterans, we summarize the data of forewing length, width and ratio of wing length/width in Table 1 and plot the ratio of wing length/width vs. wing length (in mm) in Fig. 7. The data and Fig. 7 indicate the family of Choristopsychidae has the lowest ratio, meaning broadest forewings among mecopterans. In addition, the data and Fig. 7 seem to show a general trend that for representative specimens of these families, the larger the body size, the narrower the forewings (comparatively higher ratio). The linear regression trend line is represented by Y1 = 0.036*X + 2.620. For example, the family of Cimbrophlebiidae have large body size, with forewing length from 25 mm to 30 mm, and high ratio, with L/W ratio from 3.5 to 4.4 (
Data of forewing length, width and length/width ratio of representatives of ten Families in Mecoptera.
| Family | Genus | Species | No. of fossil | Length of forewing (mm) | Width of forewing (mm) | Ratio of length/width |
|---|---|---|---|---|---|---|
| Aneuretopsychidae Rasnitsyn & Kozlov, 1990 | Jeholopsyche Ren, Shih & Labandeira, 2011 | Jeholopsyche liaoningensis Ren, Shih & Labandeira, 2011 | CNU-M-LB2005002 | 21.5 | 6 | 3.6 |
| Jeholopsyche completa Qiao, Shih & Ren, 2012 | CNU-MEC-LB2011062 | 16.5 | 5.2 | 3.2 | ||
| Jeholopsyche bella Qiao, Shih & Ren, 2012 | CNU-MEC-LB2011063 | 17 | 5.4 | 3.2 | ||
| Jeholopsyche maxima Qiao, Shih & Ren, 2012 | CNU-MEC-LB2011064 | 31.7 | 8.5 | 3.7 | ||
| Orthophlebiidae Handlirsch, 1906 | Orthophlebia Westwood, 1845 | Orthophlebia liaoningensis Ren, 1997 | LB95055 | 16 | 4.6 | 3.5 |
| Orthophlebia nervulosa Qiao, Shih & Ren, 2012 | CNU-MEC-NN2011060 | 27.5 | 6.0 | 4.6 | ||
| Eomeropidae Cockerell, 1909 | Tsuchingothauma Ren & Shih, 2005 | Tsuchingothauma shihi Ren & Shih, 2005 | M-NN200401 | 28 | 10.5 | 2.7 |
| Typhothauma Ren & Shih, 2005 | Typhothauma yixianensis Ren & Shih, 2005 | M-LB200401 | 18 | 8 | 2.3 | |
| Pseudopolycentropodidae Handlirsch, 1925 | Pseudopolycentropus Handlirsch, 1906 | Pseudopolycentropus janeannae Ren, Shih & Labandeira, 2010 | CNU-M-NN2005001 | 8 | 4 | 2 |
| Pseudopolycentropus novokshonovi Ren, Shih & Labandeira, 2010 | CNU-M-NN2005002 | 8 | 3.9 | 2.1 | ||
| Sinopolycentropus Shih, Yang & Labandeira, 2011 | Sinopolycentropus rasnitsyni Shih, Yang & Labandeira, 2011 | CNU-MEC-NN2010044 | 6.1 | 2.4 | 2.5 | |
| Cimbrophlebiidae Willmann, 1977 | Cimbrophlebia Willmann, 1977 | Cimbrophlebia flabelliformis Bruce, 2009 | UCCIPR L-18 F-763 | 28 | 7 | 4 |
| Cimbrophlebia leahyi Bruce, 2009 | TRUIPR L-018 F-1161 | 31 | 8 | 3.9 | ||
| Cimbrophlebia brooksi Bruce, 2009 | SR062005 | 31 | 7 | 4.4 | ||
| Cimbrophlebia westae Bruce, 2009 | SRUI099600 | 25 | 6 | 4.2 | ||
| Perfecticimbrophlebia Yang, Shih & Ren, 2012 | Perfecticimbrophlebia laetus Yang, Shih & Ren, 2012 | CNU-M-NN2010004 | 26.9 | 7.6 | 3.5 | |
| Nannochoristidae Tillyard, 1917 | Protochoristella Sun, Ren & Shih, 2007 | Protochoristella polyneura Sun, Ren & Shih, 2007 | CNU-M-NN2006049 | 7.5 | 2 | 3.8 |
| Protochoristella formosa Sun, Ren & Shih, 2007 | CNU-M-NN2006006 | 8 | 3 | 2.7 | ||
| Itaphlebia Sukatsheva, 1985 | Itaphlebia exquisita Liu, Zhao & Ren, 2010 | CNU-MEC-NN2009145 | 10.2 | 3.5 | 2.9 | |
| Itaphlebia laeta Liu, Zhao & Ren, 2010 | CNU-MEC-NN2009311 | 8.2 | 2.6 | 3.2 | ||
| Mesopsychidae Tillyard, 1917 | Lichnomesopsyche Ren, Labandeira & Shih, 2010 | Lichnomesopsyche gloriae Ren, Labandeira & Shih, 2010 | CNU-M-NN2005020 | 25 | 7 | 3.6 |
| Lichnomesopsyche daohugouensis Ren, Labandeira & Shih, 2010 | CNU-M-NN2005022 | 22 | 6.5 | 3.4 | ||
| Vitimopsyche Novokshonov & Sukatasheva, 2001 | Vitimopsyche kozlovi Ren, Labandeira & Shih, 2010 | CNU-M-HP2005001 | 24 | 8 | 3 | |
| Bittacidae Handlirsch, 1906 | Exilibittacus Yang, Ren & Shih, 2012 | Exilibittacus lii Yang, Ren & Shih, 2012 | CNU-M-NN2010001 | 7.5 | 2.2 | 3.4 |
| Preanabittacus Novokshonov, 1993 | Preanabittacus validus Yang, Ren & Shih, 2012 | CNU-MEC-NN2010005 | 18.7 | 5.6 | 3.3 | |
| Megabittacus Ren, 1997 | Megabittacus spatiosus Yang, Ren & Shih, 2012 | CNU-MEC-NN2010003 | 41.0 | 11.5 | 3.6 | |
| Formosibittacus Li, Ren & Shih, 2008 | Formosibittacus macularis Li, Ren & Shih, 2008 | CNU-M-NN2007001 | 23 | 5 | 4.6 | |
| Jurahylobittacus Li, Ren & Shih, 2008 | Jurahylobittacus astictus Li, Ren & Shih, 2008 | CNU-M-NN2007002 | 12.6 | 3.0 | 4.2 | |
| Panorpidae Latreille, 1805 | Panorpa Linnaeus, 1758 | Panorpa kunmingensis Fu & Hua, 2009 | 28-08-1985 | 10.0-10.3 | 2.1-2.5 | 4.1-4.8 |
| Panorpa kiautai Zhou, Hu & Wu, 1993 | 1982-03-25 | 14.0 | 3.0 | 4.7 | ||
| Panorpa choui Zhou, Hu & Wu, 1993 | 1986-07-20 | 14.0 | 3.5 | 4 | ||
| Neopanorpa Zhou, Hu & Wu, 1993 | Neopanorpa obstrusa Zhou, Hu & Wu, 1993 | 1987-06-20 | 11.0 | 2.2 | 5 | |
| Neopanorpa moganshanensis Zhou, Hu & Wu, 1993 | 1982-07-20 | 12.5 | 3 | 4.2 | ||
| Neopanorpa tengchongensis Zhou, Hu & Wu, 1993 | 1983-05-24 | 14 | 3 | 4.7 | ||
| Neopanorpa menghaiensis Zhou, Hu & Wu, 1993 | 1984-04-25 | 14.5 | 2.8 | 5.2 | ||
| Choristopsychidae Martynov, 1937 | Choristopsyche Martynov, 1937 | Choristopsyche tenuinervis Martynov, 1937 | 9.5 | 5 | 1.9 | |
| CNU-MEC-NN2011080 | 11.8 | 6.7 | 1.8 | |||
| CNU-MEC-NN2009317 | 9.5 | 4.9 | 1.9 | |||
| CNU-MEC-NN2009414 | 10.1 | 6.4 | 1.6 | |||
| Choristopsyche perfecta sp. n. | CNU-MEC-NN2009352 | 18.8 | 10.0 | 1.9 | ||
| CNU-MEC-NN2011082 | 22.2 | 11.4 | 1.9 | |||
| Choristopsyche asticta sp. n. | CNU-MEC-NN2009394 | 20.7 | 10.2 | 2.0 | ||
| Paristopsyche gen. n. | Paristopsyche angelineae sp. n. | CNU-MEC-NN2011069 | 11.2 | 7.5 | 1.5 | |
| CNU-MEC-NN2011078 | 10.7 | 6.4 | 1.7 | |||
| CNU-MEC-NN2011076 | 8.4 | 5.5 | 1.5 |
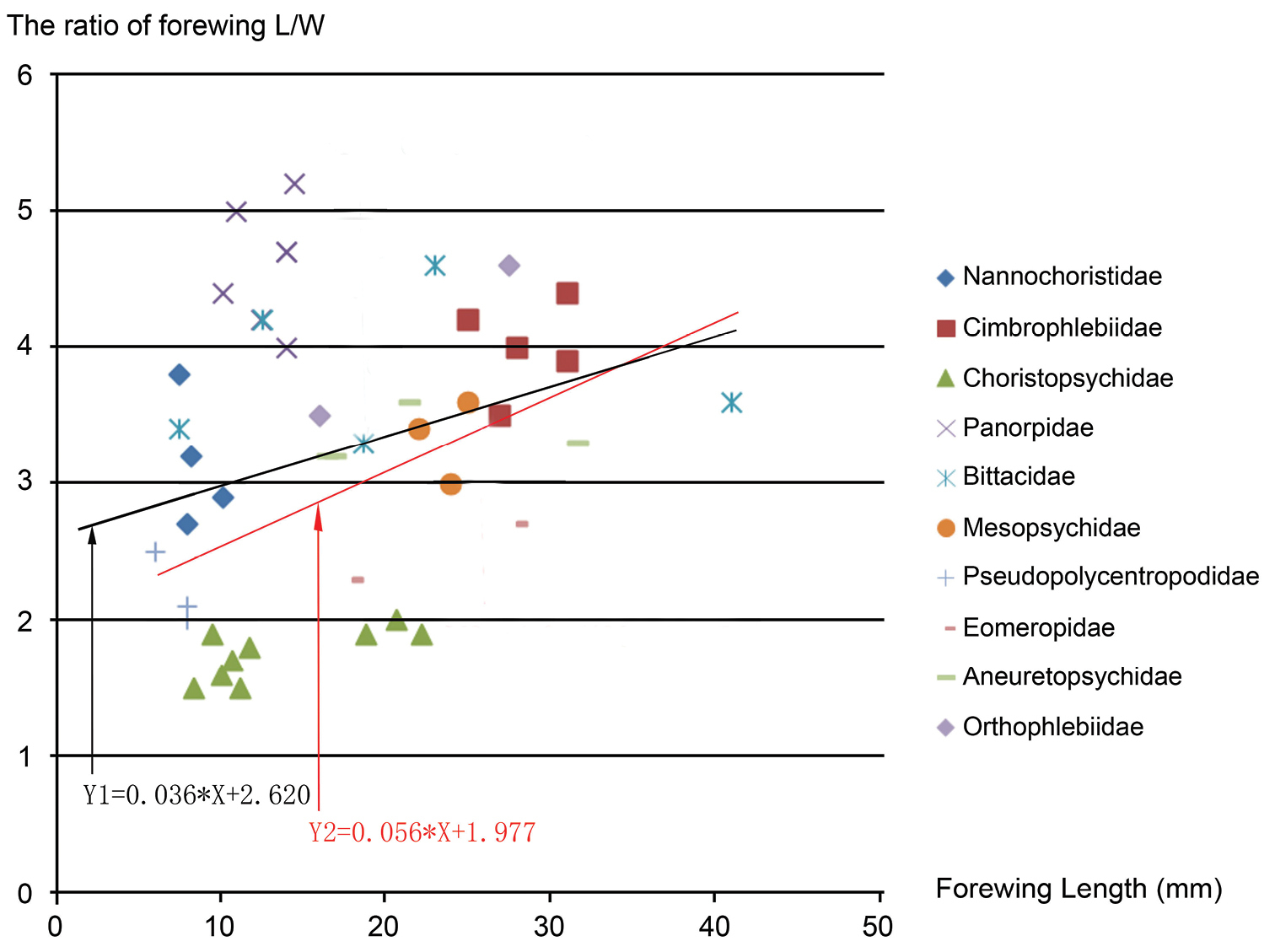
The ratio of forewing length (L)/width (W) vs. forewing length (in mm) of ten representative Families of Mecoptera. For all data points, the linear regression trend line is represented by Y1 = 0.036*X + 2.620. Excluding the data of Panorpidae, the linear regression trend line is Y2 = 0.056*X + 1.977.
The oval-shape forewings for choristopsychids are rather unique for mecopterans. Broad subtriangular (Choristopsyche asticta sp. n.) forewings can be found in Pseudopolycentopus (e.g. Pseudopolycentopus janeannae Ren, Shih & Labandeira, 2010, Pseudopolycentopus novokshonovi Ren, Shih & Labandeira, 2010, and Sinopolycentropus rasnitsyni Shih, Yang, Ren & Labandeira, 2011) with length/width ratio 2.1, 2.2 and 2.5 respectively (
Spots and bands of dark and light color are rather common for many mecopteran fossils from the Middle Jurassic of northeastern China. However, the patterns and many dark and light spots on all four wings, symmetric between left and right wings, are unique for choristopsychids (except for Choristopsyche asticta sp. n. without dots). It is likely that these spots and patterns on forewings might have served potential functions of mimicry, disruptive camouflage, or mate identification. The symmetric pattern between the left and right wings suggests that these dots may have been genetically controlled for an individual. We do not use the spots and bands as a diagnostic character.
The paratype of Paristopsyche angelineae sp. n. (CNU-MEC-NN2011078) exhibits individual asymmetry by having vein MP3 with two branches on the right wing, not the typical one branch on the left wing. Numerous cases of asymmetric variations within individual fossil insects have been reported from the Mesozoic of northeastern China. In the paratype of Synapocossus sciacchitanoae Wang, Shih & Ren, 2012 from Daohugou, China, the left and right forewings of CNU-HEM-NN2007008p/c show some individual variation, i.e., 1 mm of coalescence within the left wing and only a point contact on the right wing (
These new Chinese Choristopsychids, the first record in China, show many venational differences from the previously reported Choristopsyche tenuinervis Martynov, 1937. In addition, these new fossils with well preserved body structure and wings enhance our understanding of the morphological characters of this family, and provide a basis for future phylogenetic studies. Furthermore, these new species from China reveal that the early diversification of Choristopsychidae was well underway by the Middle Jurassic, while broadening the diversity of Mesozoic Mecoptera in China.
We appreciate Qiang Yang, Yongjie Wang, Weiting Zhang, Taiping Gao in the CNU Key Lab for their detailed comments and fruitful suggestion, and appreciate reviewers for their detailed comments. This research is supported by National Basic Research Program of China (973 Program) (2012CB821906), the National Natural Science Foundation of China (No. 31230065, 31272352, 41272006) and Project of Great Wall Scholar and KEY project of Beijing Municipal Commission of Education (grants KZ201310028033).