





(C) 2014 Juan M. Fuentes-Reinés. This is an open access article distributed under the terms of the Creative Commons Attribution License (CC BY 4.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
For reference, use of the paginated PDF or printed version of this article is recommended.
Citation: Fuentes-Reinés JM, Suárez-Morales E (2014) A new subspecies of Nitokra affinis Gurney, 1927 (Copepoda, Harpacticoida) from the Caribbean coast of Colombia. ZooKeys 378: 1–15. doi: 10.3897/zookeys.378.6695
Plankton samples from Laguna Navio Quebrado, La Guajira, northern Colombia, yielded male and female specimens of an harpacticoid copepod that was first identified as the widely distributed species Nitokra affinis Gurney, 1927 for which at least four subspecies have been described from different geographic areas. A more detailed examination of the Colombian specimens revealed that it differs from the other morphs so far considered as subspecies. The Colombian specimens differ from the other four known subspecies in the following aspects: (1) rostrum with long projection, (2) relatively long exopod of P1, almost as long as first endopodal segment, (3) endopodal and exopodal rami of P2 equally long, (4) a reduced number of endopodal setal elements of the male P5. It also differs from N. a. californica Lang in details of the ornamentation of the urosomites. Descriptions and illustrations of this new subspecies, the first one described from the Neotropical region, are presented together with a key to the five known subspecies of Nitokra affinis. As in many other cases of presumedly widespread species of harpacticoids, it is possible that N. affinis represents a species complex with more restricted distributional patterns, a notion that certainly deserves further study.
Harpacticoids, taxonomy, meiofauna, marine crustaceans, lagoon systems biota
The family Ameiridae is one of the most diverse among the harpacticoids; the genus Nitokra Boeck, 1865 is contained in this group. Species of this genus occur in fresh, brackish and marine water habitats (
In Colombia only three species and subspecies of Nitokra: the first record, involving the description of the subspecies Nitokra lacustris pacifica Reid, 1987 was published by
From a biological survey of a coastal lagoon system in the Caribbean coast of Colombia, specimens of a species tentatively identified as Nitokra affinis Gurney, 1927 were obtained; a further analysis of these specimens revealed that they show consistent differences with respect to the other four subspecific forms currently known. In this contribution we describe and illustrate this taxon and provide a key to the identification of the five subspecies of Nitokra affinis.
Samples of near-shore and open water plankton were taken from the Laguna Navío Quebrado, Colombia (11°25'N, 73°5'W) from April to December 2012. Samples were mainly in the littoral areas with vegetation (macrophytes and mangrove) but also from limnetic areas close to oyster banks. Water salinity ranged from 0 to 28‰. Water samples were collected using a bucket of 25 L at both littoral vegetation areas and open water. Samples were filtered with a standard zooplankton net with a 45 μm mesh and fixed and preserved in 70% ethanol. Dissected specimens and appendages were mounted in glycerine and sealed with Canada balsam. Drawings of the mounted appendages were prepared with a camera lucida and also photographed using a Kodak Easy Share C140 digital camera adapted to a compound microscope. The specimens were measured in lateral position, from the tip of rostrum to the posterior margin of the caudal rami. Morphological nomenclature follows the terminology proposed by
One female holotype (UARC133M) and one male allotype (UARC134M), ethanol-preserved. Paratypes: one female (UARC142M-147M) and one male (UARC136M-141M), plus two females and two males (UARC135M). Additional material: Six adult females, four adult males in authors’ (JF-R) personal collection. Two adult females, two adult males from same locality and date (ECO-CHZ-09088).
Laguna Navío Quebrado, La Guajira, Colombia (11°25'N, 73°5'W).
Habitus in lateral view as in Figure 1A. Body subcylindrical, tapering posteriorly, total body length measured from tip of rostrum to posterior margin of caudal rami ranging from 588 to 714 μm (average 660 μm, n = 6; holotype: 700 μm). Rostrum subtriangular with 2 sensilla and apical rostral projection which is about half the length of rostrum (Fig. 3H). Genital double-somite distinct dorsally and laterally, with partial ventral suture (Fig. 2A, B). Anterior ventral surface of genital somite with incomplete rows of spinules on medial outer surface, distal row of spinules and pair of sensilla. Succeeding urosomite with dorsal incomplete row of spinules on medial surface and distal row of spinules covering lateral margin and only part of ventral margin (Fig. 2A, B); same somite with ventral curved row of minute spinules on central surface, incomplete row on medial outer margin and 2 sensilla on posterior margin. Preanal somite with similar ornamentation pattern except for spinules encircling posterior margin of somite, absence of curved row on ventral surface and additional row of minute spinules. Anal somite with ventral and dorsal rows of spinules along posterior margin bordering insertion of caudal rami; anal operculum semicircular, ornamented with 14–20 large spinules, flanked by 2 sensilla and rows of minute spinules (Figs 1C, 2B). Caudal ramus short, subquadrate, with rounded margins, armed with 6 setae, seta I small, seta II as long as seta I, seta III on distal outer position, about 1.5 times as long as setae I and II. Setae IV and V thick, long, the former being longest; seta VI slightly longer than seta III. Dorsal seta VII uniarticulate at base. Ramus ornamented with spinules at insertion of setae (Figs 1F, 2A, B).
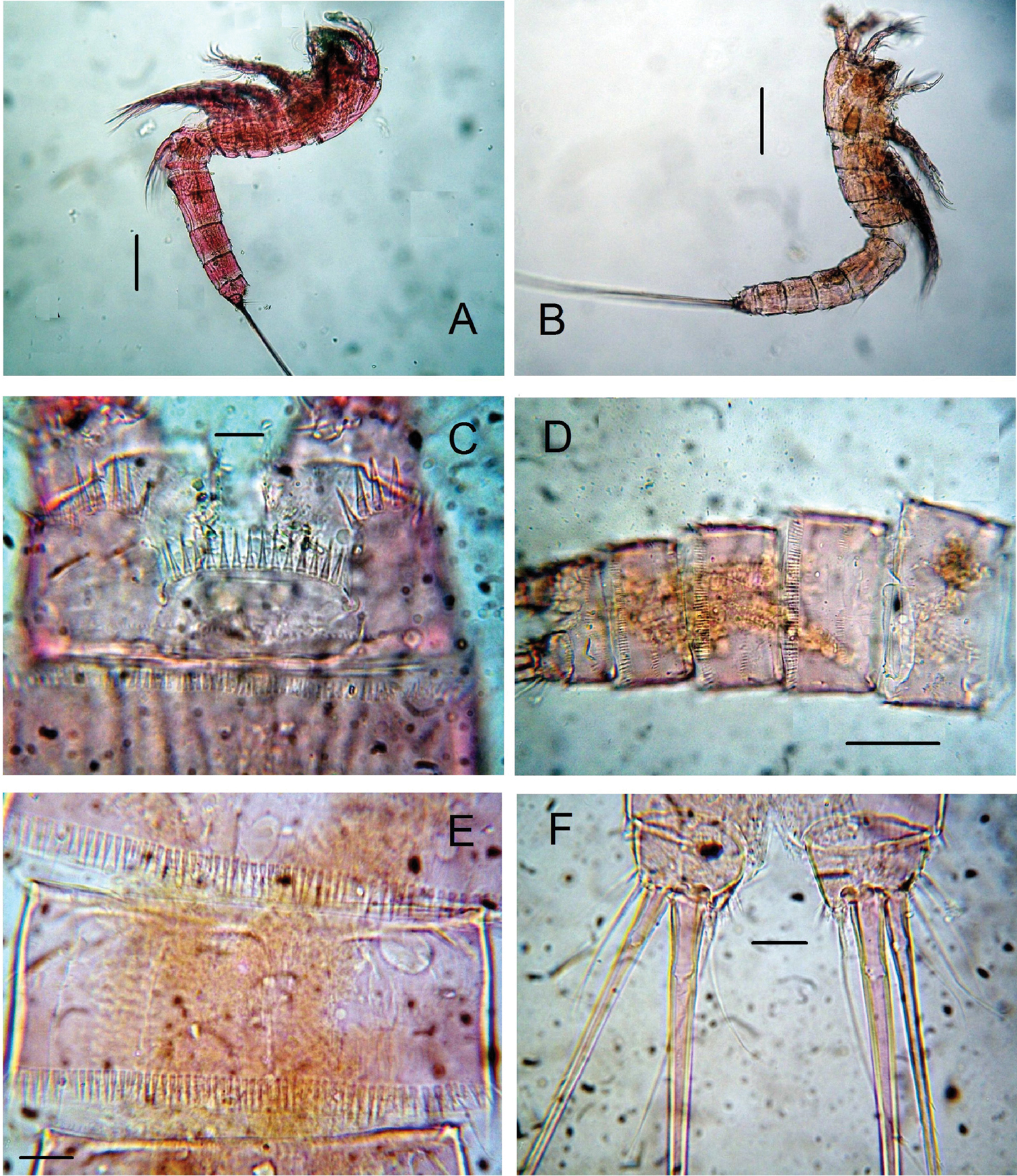
Nitokra affinis colombiensis ssp. n., from northern Colombia. A adult female, habitus, lateral view B adult male, habitus, lateral view C female, anal somite showing ornamentation of anal operculum D male, urosome, ventral view E male, third urosomite, ventral view F male, caudal rami, ventral view. Scale bars: A, B = 100 μm, C, E, F = 10 μm, D = 50 μm.
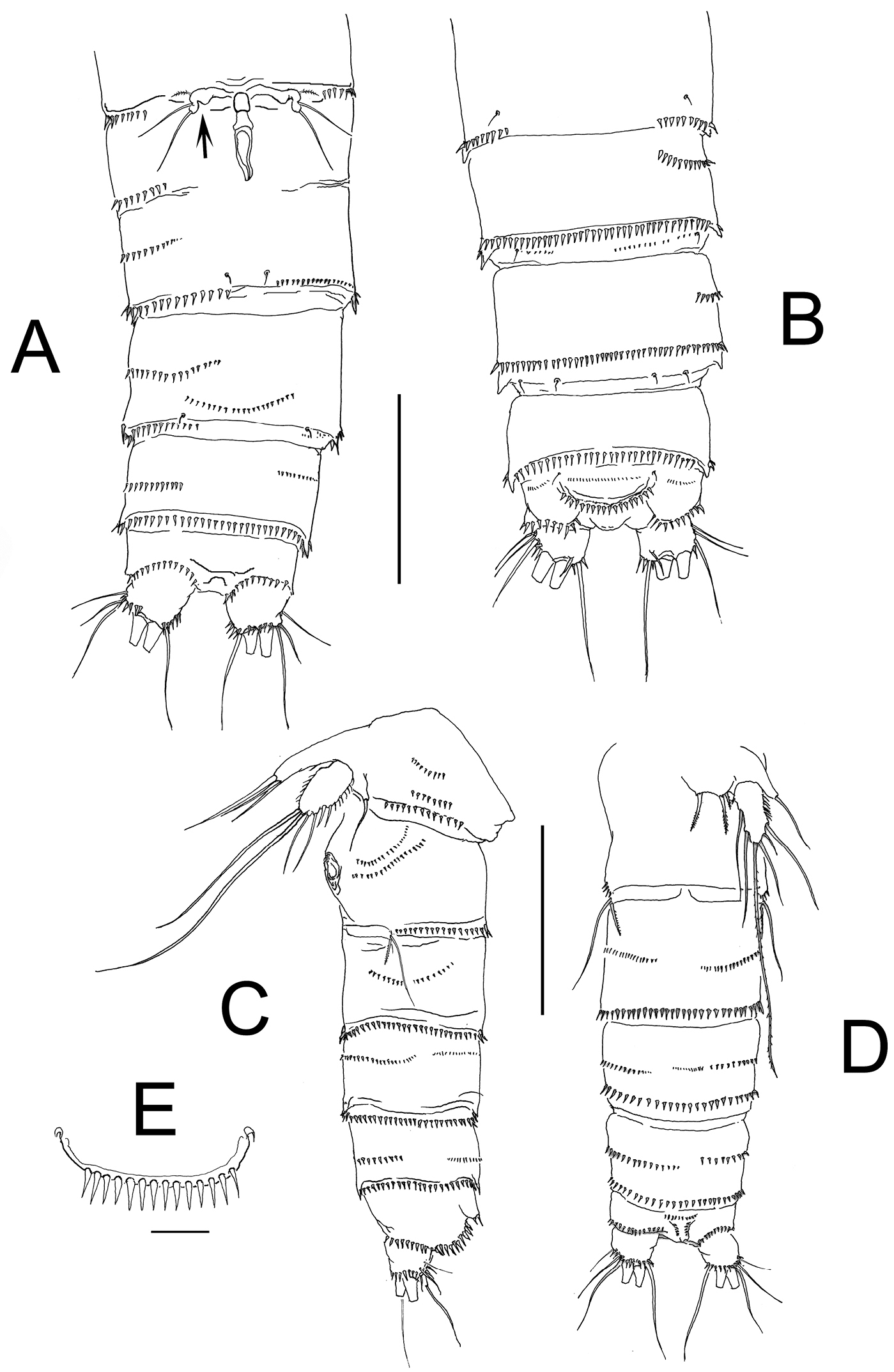
Nitokra affinis colombiensis ssp. n., from northern Colombia. A female, urosome, ventral view showing genital field and P6 B same, dorsal view, showing genital field and sixth leg plate, arrowed C male, urosome, lateral view showing P5 and P6 plate D same, ventral view E male, detail of ornamentation of anal operculum. Scale bars: A–D = 100 μm, E = 10 μm.
Antennule. 8-segmented, second segment about 1.5 longer than first and third segments, aesthetasc on fourth segment reaching well beyond distal end of terminal segment (Fig. 3A). Second segment longest, about 1.5 times as long as third segment and 2.1 times longer than preceding first segment. Fourth segment about as long as third segment. Armature as follows: 1(1), 2(7), 3(7), 4(3+aes), 5(2), 6(3), 7(3), 8(7).
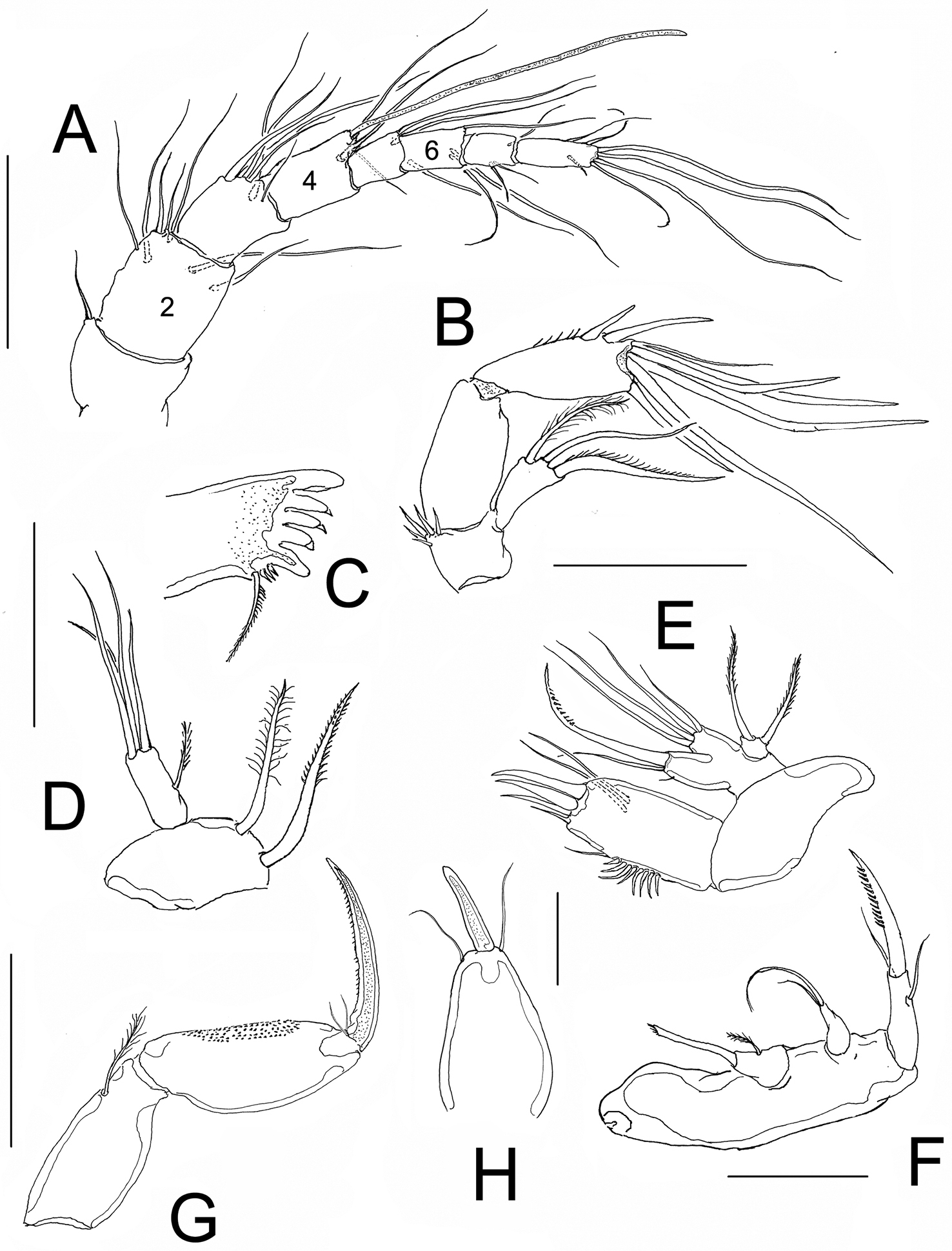
Nitokra affinis colombiensis ssp. n., adult female from northern Colombia. A antennule B antenna C mandible blade D mandibular palp E maxillule F maxilla G maxilliped H rostrum with rostral process. Scale bars: A–G = 50 μm, H = 10 μm.
Antenna. Basis with group of 5 unequal spiniform setae, first endopodal segment subrectangular, smooth, second endopodal segment with subdistal row of spinules on inner margin, with 2 lateral inner spines and 6 distal elements, outermost two of them basally fused at insertion. Exopod one-segmented with 3 setae, 2 pinnate and 1 smooth seta (Fig. 3B).
Mandible. Gnathobase with 5 large teeth, and long dorsal seta ornamented with short spinules (Fig. 3C). Mandibular palp 2-segmented, first segment (basis) with 2 setae. Endopodal segment with 1 short lateral and 4 long apical setae (Fig. 3D).
Maxillule. Arthrite of praecoxa ornamented with group of subequal spinules, arthrite armed with 2 subdistal setae and 4 distal elements. Coxal endite with 2 setae. Basis with 4 setae; exopod 1-segmented, with 2 setae, one slender, the other thicker, distally serrate (Fig. 3E).
Maxilla. Syncoxa naked, with 2 endites, proximalmost with single, slender modified element with distal tuft of setules and with short proximal seta; second endite with 2 unequal setae. Allobasis forming strong serrate claw with 2 accessory setae on proximal position (Fig. 3F).
Maxilliped. Subchelate. Syncoxa with single seta on inner distal corner, basis unarmed, with longitudinal patch of spinules. Endopod drawn into long and slender lightly serrate claw with 2 short accessory setae (Fig. 3G).
P1. Coxa with outer row of slender spinules. Basis with spinules bordering insertion of exopodal and endopodal rami, inner basipodal spine short, stout, reaching 1/3 of length of first endopodal segment. Outer basipodal spine short, stout, spinulated. EXP and ENP 3-segmented. Exopodal ramus shorter than first endopodal segment. Third exopodal segment with 2 apical geniculate setae. First endopodal segment about 2.9 longer than its width; third endopodal segment with terminal claw, the latter about 1.5 times as long as segment (Fig. 4A).
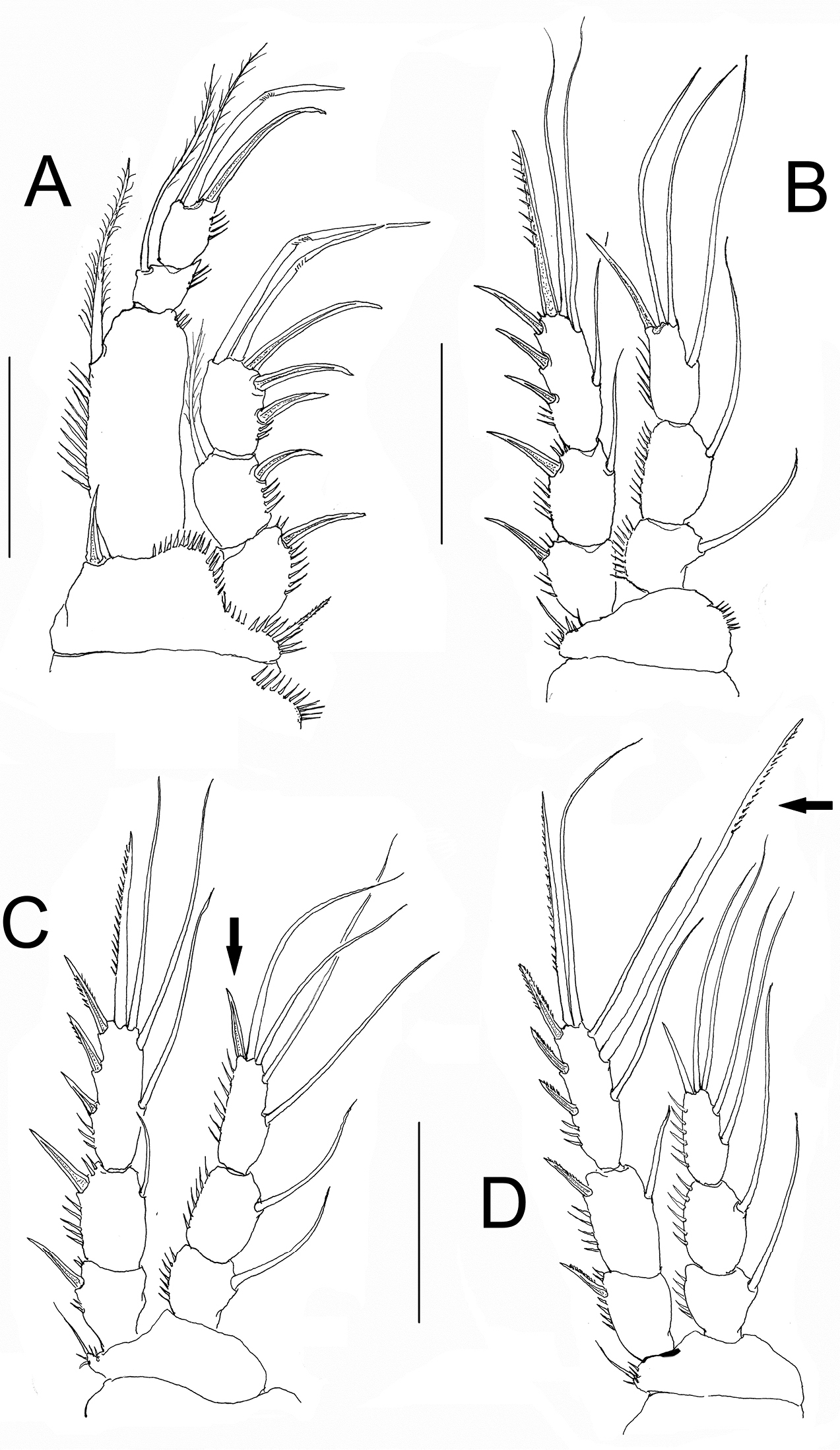
Nitokra affinis colombiensis ssp. n., adult female from northern Colombia. A first swimming leg (P1) B second swimming leg (P2) C third swimming leg (P3) D fourth swimming leg (P4) showing modified pectinate element (arrowed). Scale bars: A–D= 50 μm.
P2. Coxa with outer row of slender spinules. Basipod with 2 groups of spinules, as figured. Exopod and endopod 3-segmented. Exopod endopod equally long. First exopodal segment without inner seta, second and third exopodal segments with inner seta. Exopod without modified setae, outer margin of exopodal segments spinulated. Endopod 3-segmented, outer margin of segments ornamented with spinules (Fig. 4B).
P3. Coxa with outer row of slender spinules. Basipod with spinules only at insertion of outer basipodal seta. Exopodal and endopodal rami as in P2 except for shorter apical spiniform element (arrowed in Fig. 4C) and endopod slightly shorter than exopod (Fig. 4C).
P4. Coxa with outer row of slender spinules. Basipod as in P3. EXP longer than ENP (Fig. 4D). Middle inner seta of EXP3 thicker and longer than adjacent setae (arrowed in Fig. 4D).
P5. EXP subrectangular, about 1.66 longer than it is width, with 6 setae. Relative length of exopodal setae from inner to outer element as follows: 0.81, 1.00, 0.18, 0.38, 0.18, 0.56. Endopodal lobe quadrate, reaching almost halflength of EXP, with 5 spinulose setae, outermost being longest; relative length of setae from inner to outer elements as follows: 0.32, 0.37; 0.43, 1, 0.46 (Fig. 6A).
Armature formula of female P1-P5 as follows:
| P1 | P2 | P3 | P4 | P5 | |
|---|---|---|---|---|---|
| EXP | I-0;I-1;III, 2, 0 | I-0;I-1;III, I, 1, 2 | I-0;I-1;III, I, 1, 2 | I-0;I-1;III, I, 1, 3 | 6 |
| ENP | 0-1; 0-1;I, 2, 0 | 0-1;0-1;I, 2, 1 | 0-1;0-1;I, 2, 2 | 0-1;0-1;I, 2, 2 | 5 |
P6. Represented by narrow plate with subdistal lobe-like process marked by a notch (arrowed in Fig. 2A); plate bearing 3 elements, 2 equal slender setae and outer small spinulated seta (Fig. 6B).
Smaller than female, habitus in lateral view as in figure 1B. Total body length ranging from 518 to 574 μm (mean, 546 μm; n = 4; allotype: 518 μm). Ornamentation of urosomites resembling that of female except for position of rows of minute spinules on ventral surface of genital and preanal somites (Fig. 2C, D). Anal somite with row of small spinules on posterior margin at insertion of caudal rami. (Figs 1D, E, 2C, D). Rostrum, antennae and mouthparts as in female.
P1. As in female except for an additional row of spinules on the coxa, presence of small slender seta and 1 geniculate apical seta (arrowed in Fig. 5A), instead of 2 on ENP3, slenderer ENP1, and dimorphic modified inner basipodal spine (Fig. 5A, B).
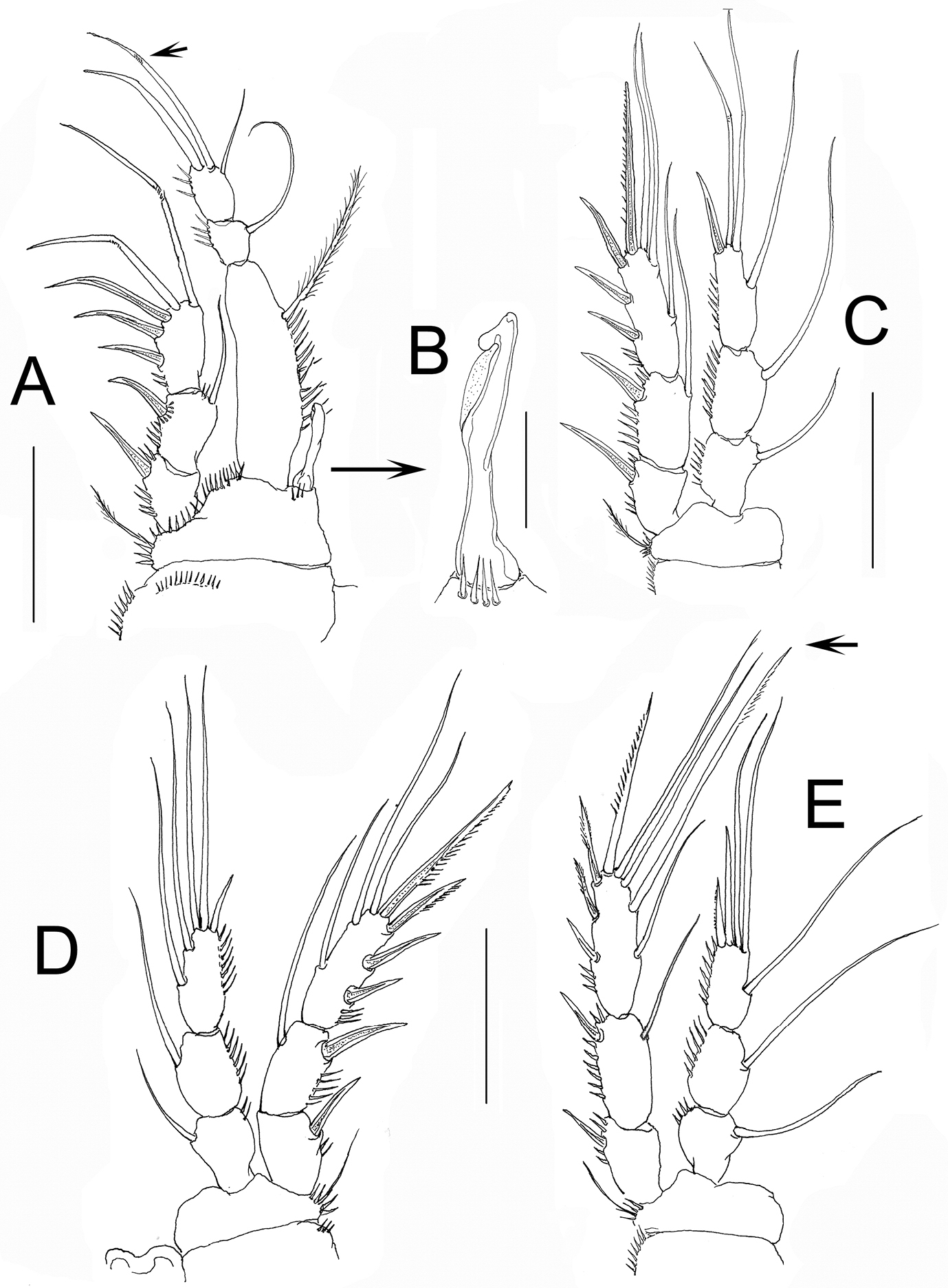
Nitokra affinis colombiensis ssp. n., adult male from northern Colombia. A first swimming leg (P1) B modified inner basipodal spine of P1 C second swimming leg (P2) D third swimming leg (P3) E fourth swimming leg (P4). Scale bars: A, C–E = 50 μm, B = 10 μm.
P2-P4. As in female (Fig. 5C-E), including thickened middle inner seta of EXP3 (arrowed in Fig. 5E) which is relatively shorter than in female.
P5. EXP subquadrate, armed with 6 setae, distal innermost being longest, reaching midlength of fourth urosomite (Fig. 2C, D). Baseoendopod with 3 unequally long setae, middle one longest, about twice as long as the other two (Fig. 6C).
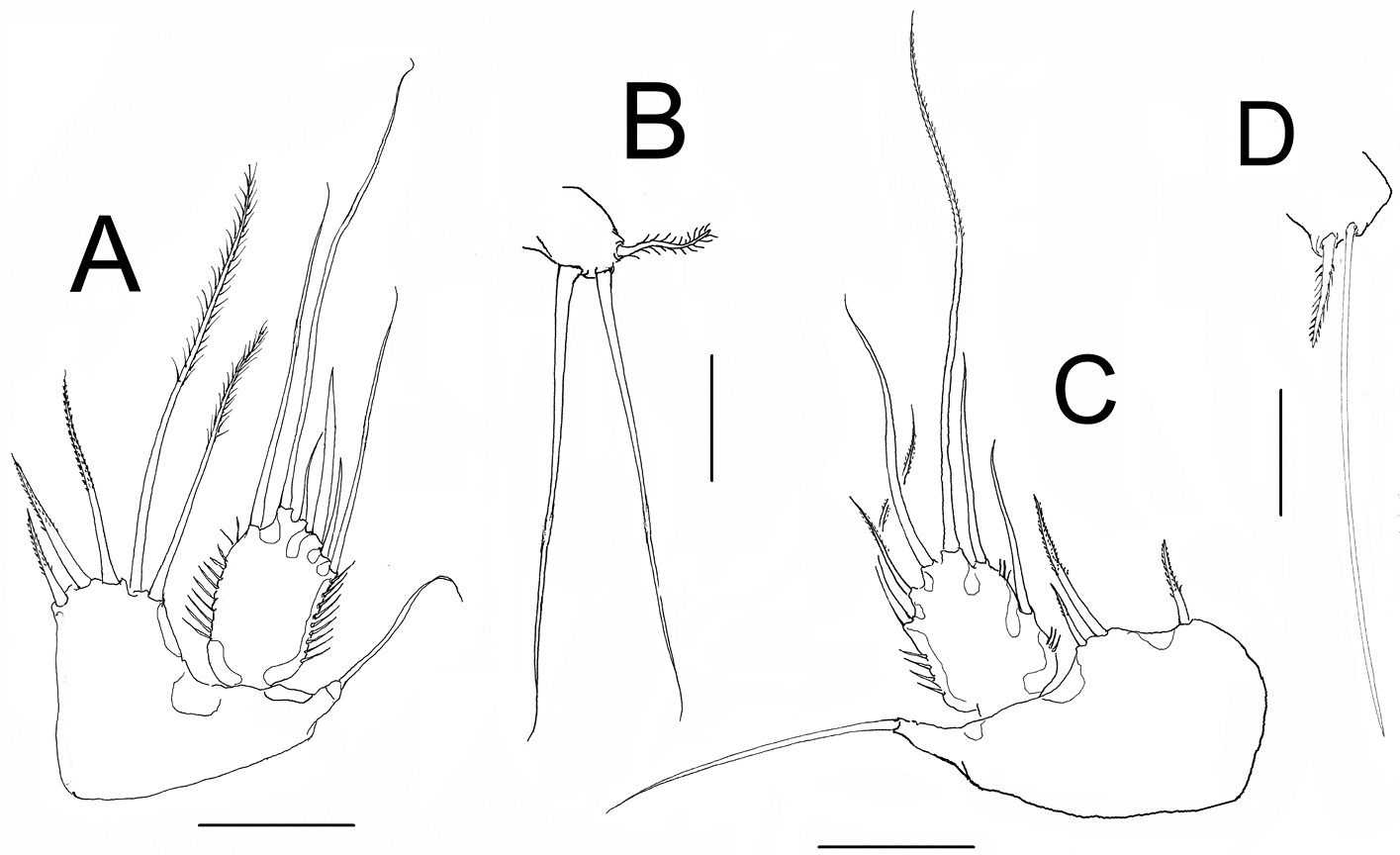
Nitokra affinis colombiensis ssp. n. from northern Colombia. Adult female: A fifth leg B sixth leg. Male: C fifth leg D sixth leg. Scale bars: A, C = 50 μm; B, D = 10 μm.
P6. With 2 unequal seta, inner one three times longer than outer seta (Figure 6D). Caudal rami as in female (Fig. 2C, D).
Variability. One male with 2 elements (instead of3) on ENP3 of P1.
The new subspecies is named after Colombia, the country from which it was first discovered.
The estuary Laguna Navío Quebrado has a surface area of 10.7 km2; it is characterized by the presence of an oyster bank in the limnetic area and vegetation (mangrove and beds of macrophytes) in the littoral zone. Water temperature ranged between 28 and 31 °C, salinity between 0-28 psu, and pH values were 7.8–8.3.
Based on the combination of the armature formula of the P1EXP2-3, three species groups can be recognized within the genus Nitokra (Gómez et al., 2012). The first group exhibits one inner seta and four elements on P1EXP2 and EXP3, respectively, the second group lacks an inner seta on P1EXP2 but bears five setae on P1EXP3. The third group exhibits one inner seta, and five setal elements on P1EXP2 and EXP3, respectively. Nitokra affinis Gurney is part of this third group. This is a very widespread species, recorded from different geographic regions, including the Atlantic and Pacific Oceans, the Mediterranean, the Black Sea, the Red Sea, the Caribbean, and Brazil (
Despite the fact that
The four known subspecific forms of Nitokra affinis are known from different geographic areas: Nitokra affinis affinis from the Suez Canal, European and Mediterranean waters, and Bermuda (
The Colombian specimen shares most characters with Nitokra affinis Gurney, and its subspecific forms, including the armature formula of P1-P4, the morphology of the mouthparts, the size proportions and armature of the caudal rami, and the number of setae on the female and male P5EXP. The new subspecies, Nitokra affinis colombiensis ssp. n. differs from its congeners in the following aspects: (1) in the Colombian specimens the rostrum has a long rostral projection. This structure has not been hitherto described or depicted in any other subspecies of Nitokra affinis; (2) the length of the EXP with respect to the enlarged P1ENP1 differs among these subspecies; in Nitokra affinis affinis and Nitokra affinis rijekana the exopod reaches about the point of insertion of the inner seta of the first endopodal segment (
Overall, the new subspecies most closely resembles Nitokra affinis californica, but some additional characters can be useful to separate these two species; the number of spines on the posterior margin of the female anal operculum is only 14-20 in the new subspecies (Fig. 2H) vs. +25 in Nitokra affinis californica (Lang, 1965, fig. 196b). The second antennular segment of Nitokra affinis californica is relatively longer (1.7 times as long as third segment) than in Nitokra affinis colombiensis (1.3). Also, the fourth segment is elongate in Nitokra affinis californica (3.3 times as long as wide) and clearly shorter (1.4) in the new subspecies. The ornamentation of the maxillipedal basis is represented by row of short hair-like elements in Nitokra affinis californica (Lang, 1965, fig. 197c) whereas this segment has a patch of spinules in the Colombian specimens. The shape and armature of the female sixth leg plate differs between these taxa, the two inner setae are unequally long in Nitokra affinis californica but these elements have the same length in the Colombian specimens (Figs 2A, 6B). Also, in the new subspecies the distal section of the plate has a subterminal notch (arrowed in Fig. 2A) which is absent in Nitokra affinis californica (Lang, 1965, fig. 196d). The shape of the male fifth leg exopod is clearly subrectangular in the new subspecies vs. subtriangular in Nitokra affinis californica (Lang, 1965, fig. 197 h). In addition, the middle apical seta of the male fifth leg exopod is distinctively long in the Colombian specimens, it reaches midlength of the fourth urosomite (Fig. 2C, D) whereas this seta is clearly shorter in the Californian subspecies, barely reaching beyond the second urosomite (
As in many other cases of presumedly widespread species of harpacticoids, it is possible that Nitokra affinis represents a species complex with more restricted distributional patterns, a notion already advanced by
| 1A | Female P1ENP1 less than 3.8 times as long as wide; EXP of P1 reaching the point insertion of inner seta of ENP1 (Figs 3A, 4A); male P5ENP with 3 setae; rostrum with rostral projection | Nitokra affinis colombiensis ssp. n. |
| 1B | Female P1ENP1 more than 3.8 times as long as wide; EXP of P1 with a different length; male P5ENP with 4 or 5 setae; rostrum without rostral projection | 2 |
| 2A | Middle inner seta of P4EXP3 (arrow in Fig. 3D) not longer and stronger than distal inner seta; male P5ENP with 5 setae | Nitokra affinis rijekana Petkovski, 1954 |
| 2B | Middle inner seta of P4EXP3 longer and stronger than the distal inner seta | 3 |
| 3A | Posterior edge of antepenultimate somite with incomplete spinules ring, dorsal to ventro-lateral only, ventral margin smooth | Nitokra affinis affinis Gurney, 1927 |
| 3B | Posterior margin of antepenultimate somite spinulose as a continuous ring | 4 |
| 4A | P1EXP short, not reaching insertion of inner seta of P1 ENP1; female P5EXP not elongated at distal half, about 1.3 as long as wide, innermost distal seta about as long as adjacent distal seta | Nitokra affinis stygia Por, 1962 |
| 4B | P1EXP long, reaching well beyond insertion of inner seta of P1ENP1, both rami equal in length; P5EXP elongated at distal half, about 1.5–1.8 as long as wide; innermost distal seta about 1.5 times as long as adjacent distal seta | Nitokra affinis californica Lang, 1965 |