





(C) 2013 Gabriel de Souza da Costa e Silva. This is an open access article distributed under the terms of the Creative Commons Attribution License 3.0 (CC-BY), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
For reference, use of the paginated PDF or printed version of this article is recommended.
Citation: Silva GSC, Roxo FF, Oliveira C (2013) Pareiorhina hyptiorhachis, a new catfish species from Rio Paraíba do Sul basin, southeastern Brazil (Siluriformes, Loricariidae). ZooKeys 315: 65–76. doi: 10.3897/zookeys.315.5307
Pareiorhina hyptiorhachis is described from Ribeirão Fernandes and Rio Pomba, Rio Paraíba do Sul basin, Brazil. The new species is distinguished from its congeners (Pareiorhina brachyrhyncha, Pareiorhina carrancas, Pareiorhina cepta, and Pareiorhina rudolphi) by the presence of a conspicuous ridge on the trunk posterior to the dorsal fin (postdorsal ridge), simple teeth, a completely naked abdomen, a round dorsal profile of the head, greater suborbital depth and greater head width. We discuss the distributional pattern of the new species and its congeners and hypothesize that headwater capture is responsible for the distribution of Pareiorhina species across different watersheds in southeastern of Brazil.
ResumoPareiorhina hyptiorhachis é descrita do ribeirão Fernandes e do rio Pomba, bacia do rio Paraíba do Sul, Brasil. A nova espécie é distinguida de suas congêneres (Pareiorhina brachyrhyncha, Pareiorhina carrancas, Pareiorhina cepta, e Pareiorhina rudolphi) por caracteres relacionados a presença de uma quilha conspícua na superficie pós-dorsal do tronco, dentes simples, abdomen completamente nú, perfil dorsal da cabeça arredondado, altura do sub-orbital e largura da cabeça. Nós discutido o padrão de distribuição da nova espécie e de seus congêneres e a hipótese de que a captura de cabeceira é responsável pela distribuição das espécies de Pareiorhina através das diferentes bacias hidrográficas do sudeste do Brasil.
Cascudinhos, Taxonomy, Freshwater, Neoplecostominae, Neotropical Region, Cascudinhos, Taxonomia, Água Doce, Neoplecostominae, Região Neotropical
The genus Pareiorhina was proposed by
An examination of the fish collections at the Laboratório de Biologia e Genética de Peixes de Botucatu (LBP) and Museu de Ciências e Tecnologia, Pontifícia Universidade Católica do Rio Grande do Sul (MCP) revealed the existence of an undescribed Pareiorhina species from the Rio Paraíba do Sul basin, Brazil. This new species is formally described herein.
All measurements were taken from point to point to the nearest 0.1 mm using digital calipers (except the postdorsal ridge depth, which was measured using a stereomicroscope and analyzed using the software Axio Vision Release 4.8.2). Counts were taken from the left side when possible. In the description, counts are followed by their frequencies in parentheses. The measurements followed
All examined material was housed at the following Brazilian institutions: LBP (Laboratório de Biologia e Genética de Peixes, Universidade Estadual Paulista Júlio de Mesquita Filho, Botucatu - SP); MCP (Museu de Ciências e Tecnologia, Pontifícia Universidade Católica do Rio Grande do Sul, Porto Alegre - RS); MZUSP (Museu de Zoologia da Universidade de São Paulo, São Paulo - SP); and NUP (Coleção Ictiológica do Núcleo de Pesquisas em Limnologia, Ictiologia e Aquicultura, Universidade Estadual de Maringá, Maringá - PR).
urn:lsid:zoobank.org:act:1D6D4D43-68CF-485B-9ABC-8FFB270E2460
http://species-id.net/wiki/Pareiorhina_hyptiorhachis
Figure 1; Table 1MZUSP 111956, female, 33.6 mm SL, Brazil, Minas Gerais State, municipality of Santa Bárbara do Tugúrio, Ribeirão Fernandes, a tributary of Rio Pomba, Rio Paraíba do Sul basin, 21°14'47"S, 43°34'07"W, 19 Jun 2011, Ferreira AT, Roxo FF, Silva GSC.
Brazil, Minas Gerais State, municipality of Santa Bárbara do Tugúrio, Rio Paraíba do Sul basin. LBP 12248, 2 males, 4 females, 1 c&s, 26.6–34.8 mm SL, collected with holotype. NUP 14331, 1 female, 29.6 mm SL, collected with holotype. LBP 1093, 1 male, 33.4 mm SL, Ribeirão Fernandes, 21°14'47"S, 43°34'07"W, 12 Oct 2001, Oliveira JC, Alves AL, Sato LR. LBP 8368, 5 females, 27.9–34.4 mm SL, Rio Pomba, 21°14'07"S, 43°30'50"W, 19 May 2009, Oliveira C, Silva GJC, Roxo FF, Pereira TNA. LBP 12257, 1 female, 27.2 mm SL, Rio Pomba, 21°14'07"S, 43°30'50"W, 19 Jun 2011, Ferreira AT, Roxo FF, Silva GSC. MCP 29432, 3 male, 1 female, 2 unsexed, (1 juvenile not measured) 23.8–39.0 mm SL, Ribeirão Fernandes, 21°14'47"S, 43°34'07"W, 12 Aug 2001, Oliveira JC, Alves AL, Sato LR.
Pareiorhina hyptiorhachis is distinguished from its congeners, except for Pareiorhina carrancas, by the presence of a postdorsal ridge (vs. the absence of a postdorsal ridge). The new species differs from Pareiorhina carrancas by having a more elevated postdorsal ridge, (Fig. 2; 16.7–26.8% of CP depth vs. 4.47–9.03%; table 1). Additionally, the new species can be distinguished from Pareiorhina cepta by having a naked abdomen (vs. having small plates covered with odontodes irregularly distributed on the abdomen); from Pareiorhina brachyrhyncha and Pareiorhina cepta by having unicuspid teeth (vs. teeth with a minute lateral cusp); from Pareiorhina carrancas and Pareiorhina rudolphi by having the anterior profile of the head rounded in dorsal view (vs. elliptical; Fig. 3) and by having a greater suborbital depth (35.0–40.5% of HL vs. 27.4–34.2% in Pareiorhina carrancas and 24.5–31.8% in Pareiorhina rudolphi). Moreover Pareiorhina hyptiorhachis is distinguished from its congeners by having a wider head (100.1–108.6% of HL vs. 91.7–98.1% in Pareiorhina brachyrhyncha, 82.9–96.2% in Pareiorhina carrancas, 83.4–90.5% in Pareiorhina cepta and 77.8–82.1% in Pareiorhina rudolphi).

Pareiorhina hyptiorhachis, sp. n., MZUSP 111956, 33.6 mm SL, holotype from Ribeirão Fernandes, Rio Paraíba do Sul basin, municipality of Santa Barbara do Tugúrio.

a Pareiorhina hyptiorhachis, sp. n., paratype, LBP 12248, 29.2 mm SL, showing the conspicuous postdorsal ridge b Pareiorhina carrancas, LBP 8380, 38.2 mm SL, showing the poorly-developed postdorsal ridge c Pareiorhina rudolphi, LBP 8044, 40.5 mm SL, showing the absence of a postdorsal ridge.

a Pareiorhina carrancas, LBP 8380, 36.5 mm SL, showing the elliptical anterior profile of the head elliptical in dorsal view b Pareiorhina rudolphi, LBP 8044, 42.0 mm SL, showing the elliptical anterior profile of the head in dorsal view c Pareiorhina hyptiorhachis, new species, holotype, MZUSP 111956, 33.6 mm SL, showing the rounded anterior profile of the head in dorsal view.
Morphometric and meristic data are given in Table 1. In lateral view, dorsal profile of body strongly convex from snout tip to distal margin of supraoccipital; straight from supraoccipital to dorsal-fin origin; slightly decreasing to end of caudal peduncle. Ventral surface of body, slightly concave at head portion, straight to convex from posterior end of head to pelvic-fin insertion, and straight but angled to posterior end of caudal peduncle. Snout tip rounded in dorsal view. Nostril small. Trunk and caudal peduncle rectangular in cross-section.
Morphometric data for Pareiorhina hyptiorhachis. SD = Standard Deviation, IO = Interorbital, OD = Orbital Diameter, CP = Caudal Peduncle.
| Pareiorhina hyptiorhachis n=21 | ||||
|---|---|---|---|---|
| Holotype | Range | Mean | SD | |
| Standard length (SL) | 33.6 | 26.6–38.8 | 31.0 | 3.0 |
| Percents of Standard length (SL) | ||||
| Predorsal length | 44.2 | 41.5–48.8 | 44.7 | 1.6 |
| Preanal length | 59.2 | 56.1–65.9 | 60.5 | 2.4 |
| Head length | 31.7 | 28.6–35.5 | 31.8 | 1.5 |
| Cleithral width | 32.8 | 30.4–36.9 | 33.5 | 1.8 |
| Dorsal-fin unbranched ray length | 21.2 | 20.3–24.1 | 22.2 | 1.1 |
| Base of dorsal fin length | 15.4 | 14.3–18.3 | 16.5 | 1.1 |
| Thorax length | 18.1 | 15.1–19.6 | 17.0 | 1.3 |
| Pectoral-fin unbranched ray length | 20.5 | 20.5–26.0 | 22.6 | 1.5 |
| Abdomen length | 27.0 | 22.6–30.1 | 26.2 | 1.6 |
| Pelvic-fin unbranched ray length | 22.5 | 17.7–26.6 | 22.9 | 2.1 |
| Anal-fin length | 15.3 | 13.7–17.8 | 15.6 | 0.9 |
| Ventral unbranched caudal ray | 24.9 | 20.3–30.5 | 25.2 | 2.8 |
| Caudal-peduncle depth | 9.0 | 8.3–11.0 | 9.39 | 0.7 |
| Postanal length | 34.9 | 31.6–38.1 | 33.9 | 1.4 |
| Anal width | 15.4 | 11.3–16.0 | 14.0 | 1.5 |
| Percents of Head Length (HL) | ||||
| Head width | 103.8 | 100.1–108.6 | 103.8 | 2.6 |
| Head depth | 61.7 | 53.5–62.8 | 56.9 | 2.3 |
| Snout length | 63.1 | 58.0–64.7 | 61.2 | 1.9 |
| Interorbital width | 37.7 | 34.8–40.7 | 38.0 | 1.4 |
| Orbital diameter | 11.4 | 11.1–15.5 | 12.7 | 1.6 |
| Suborbital depth | 39.2 | 35.0–40.5 | 37.3 | 1.6 |
| Mandibular ramus | 18.1 | 16.0–23.4 | 19.9 | 1.9 |
| Other measurements (expressed as percentages) | ||||
| Anal width/cleithral width | 47.1 | 32.0–49.7 | 42.0 | 5.1 |
| IO/OD | 29.5 | 21.6–42.1 | 33.5 | 4.5 |
| IO/Mandibulary ramus | 50.6 | 44.1–62.5 | 52.4 | 5.6 |
| Predorsal length/first ds ray length | 47.9 | 45.7–54.5 | 49.7 | 2.6 |
| Postanal length/CP depth | 25.8 | 24.8–31.1 | 27.6 | 1.9 |
| Pelvic-fin length/CP depth | 40.0 | 36.0–46.8 | 41.1 | 3.0 |
| Ventral unbranched caudal ray/CP depth | 36.2 | 32.1–46.0 | 37.5 | 4.3 |
| Postdorsal ridge depth/CP depth | 19.0 | 16.7–26.8 | 21.5 | 3.4 |
Greatest body depth at dorsal-fin origin. Body progressively narrowing posteriorly from cleithrum. Head flat to slightly convex between orbits; superior margin of orbits elevated. Eye small, orbital diameter 11.1–15.5% of HL, situated dorsolaterally just posterior of midpoint of head. Rostral margin of snout with minute, posteriorly-directed odontodes; numerous small odontodes on dorsal portion of head. Opening of swimbladder capsule small. Perforations of compound pterotic distributed on whole bone, greater and more concentrated on its ventral margin; its openings nearly rounded in median region, and irregular along inferior and posterior margins of bone. Lips large; oral disk rounded, papillose; premaxillary teeth 22 (1), 29 (1), 30 (1), 32 (1), 33 (1), 34 (2), 36 (1)*, 37 (2), 38 (1), 39 (2), 40 (2), 42 (2) or 44 (1). Dentary teeth 17 (1), 21 (1), 23 (1), 28 (1), 30 (2), 32 (2)*, 33 (2), 34 (2), 35 (1), 36 (1), 39 (1), or 45 (1). Teeth unicuspid. Maxillary barbel short and free distally.
Dorsal-fin rays ii, 7; dorsal-fin originating at vertical through posterior end of pelvic-fin base; distal margin slightly convex. Pectoral-fin rays i, 6; distal margin slightly convex; unbranched pectoral-fin ray reaching middle of unbranched pelvic-fin ray; unbranched pectoral-fin ray covered with large and pointed odontodes. Pelvic-fin rays i, 5; distal margin of fin slightly convex; tip of adpressed pelvic fin almost reaching anal-fin origin; unbranched pelvic-fin ray covered with conspicuously pointed and well-developed, and uniformly distributed odontodes which are larger at ventral portion. Anal-fin rays i, 5; distal margin slightly convex. Caudal fin rays i, 7-7, i. Adipose fin absent. Caudal fin truncated with ventral unbranched principal ray longer than dorsal ray.
Body entirely covered by bony plates, except for ventral surface of head, abdomen and region overlaying swimbladder capsule. Dorsal series of plates 24–26, mid-dorsal 17–21, median perforated plates 24–26, mid-ventral 17–22, and ventral 19–22. Trunk with conspicuous, elongated, postdorsal ridge formed by 13–15 raised, unpaired, median plates; ridge continuous posteriorly with procurrent caudal-fin rays. Head lacking crest. Head and body plates covered with minute, uniformly sized and distributed odontodes. Seven pairs of ribs associated with vertebrae 8–14. Ribs slender and poorly ossified. Total vertebrae 29.
Supraorbital sensory canal with four pores; pore s1 located on prenasal plate below nasal plate; pore s3 located on posterior portion of nasal; pore s6+s6 located between frontal plates, on horizontal line through anterior limits of eye; pore s8 located on division between frontals, sphenotic and supraoccipital plates, just above eye. Infraorbital sensory canals with six pores; pore io1 located on anterior portion of first infraorbital; pore io2 located in medial region between first and second infraorbitals; pore io3 located in medial region between second and third infraorbitals; pore io4 located in medial region between third and fourth infraorbitals; pore io5 located in medial region between fourth and fifth infraorbitals and pore io6 located between sixth infraorbital and sphenotic. Preopercular canal with three pores; pore pm2 located on ventral portion of cheek plate, pore pm3 located between cheek plate and preopercle; pore pm4 located between preopercle and compound pterotic. Two postotic pores; pore po2 located just above of branchial slit; pore po3 located in region of overlying opening of swim-bladder capsule.
Two body-coloration patterns observed. First pattern (Fig. 1): Ground color of dorsal surface of head and body yellowish brown. Ventral surface of body and head lighter than dorsal with dark spots of melanophores widely separated. Three dark saddles on dorsal surface of trunk (in some specimens not present), most anterior one inconspicuous. Lateral portion of body with inconspicuous dark stripe from head to caudal fin. Pectoral, pelvic and dorsal fins with three irregular, poorly defined bands. Caudal fin with variegated blotches. Second pattern (Fig. 4): Ground color of body uniformly dark except, ventral portion of body mostly clear; Fins with inconspicuous, irregularly defined bands: one in anal fin, two in pectoral and pelvic fins. Dorsal and caudal fins entirely dark.

Additional coloration pattern of coloration of Pareiorhina hyptiorhachis, sp. n., LBP 12257, female, 27.2 mm SL.
Males with a papilla at urogenital opening and fewer teeth in premaxillary 22–39 (vs. 32–44 females) and 17–32 dentary (vs. 30–45 females).
The specific name, hyptiorhachis is a combination of Greek, hyptios = supine, lying on the back, and rhachis = ridge, midrib, and is in reference to the conspicuous postdorsal ridge found in this species.
Pareiorhina hyptiorhachis is known from Rio Pomba and one of its tributaries, the Ribeirão Fernandes, in the municipality of Santa Barbara do Tugúrio, Minas Gerais State, Brazil (Fig. 5). This species inhabits moderate to fast-flowing streams, with a substrate of rocks and sand and margins covered by aquatic vegetation. Specimens were collected in association with loose stones, on the streambed. The new species is syntopic throughout its distribution with Astyanax sp., Characidium sp., Geophagus brasiliensis, Harttia cf. carvalhoi, Imparfinis sp., Neoplecostomus microps, Trichomycterus cf. alternatus, and Trichomycterus sp.
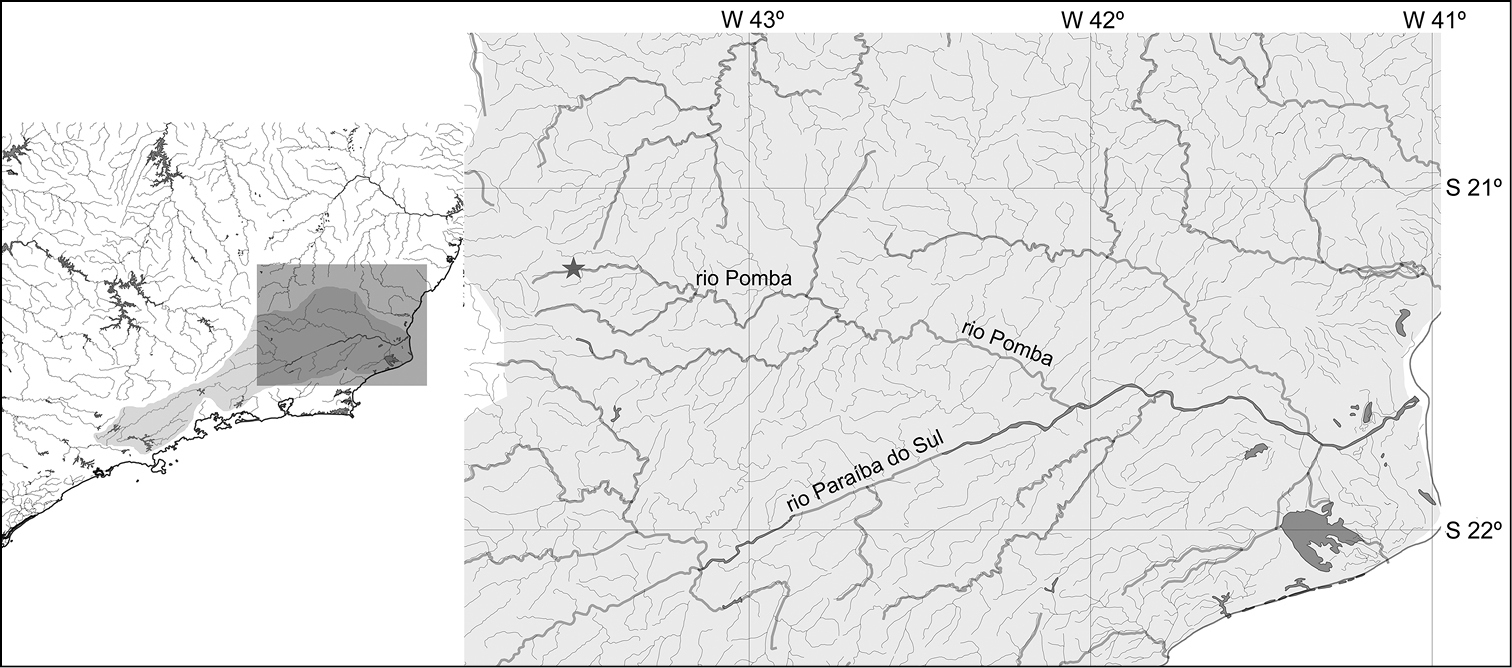
The Rio Paraíba do Sul basin indicating the type locality of Pareiorhina hyptiorhachis in Ribeirão Fernandes, a tributary of Rio Pomba, Rio Paraíba do Sul basin, 21°14'47"S, 43°34'07"W.
Pareiorhina hyptiorhachis is similar to Pareiorhina carrancas from the upper Rio Paraná basin. The two species share unicuspid teeth and the presence of a postdorsal ridge of unpaired plates, although the postdorsal ridge is better developed in Pareiorhina hyptiorhachis (all female and male samples) (Fig. 2). Moreover, the new species has more raised median unpaired plates in the postdorsal ridge (13–15 vs. 10–13 in Pareiorhina carrancas). The close relationship between Pareiorhina hyptiorhachis and Pareiorhina carrancas suggested by the molecular data of
Pareiorhina is distributed across three hydrographic basins, with Pareiorhina rudolphi, Pareiorhina brachyrhyncha and Pareiorhina hyptiorhachis from the Rio Paraíba do Sul basin; Pareiorhina carrancas from the upper Rio Paraná basin; and Pareiorhina cepta from the Rio São Francisco basin.
Isbrueckerichthys alipionis: LBP 7373, 17, 31.7–81.6 mm SL, municipality of Iporanga, SP, Rio Ribeira de Iguape basin; LBP 2660, 1, 55.1 mm SL, municipality of Iporanga, SP, Rio Ribeira de Iguape basin. Kronichthys subteres: LBP 515, 31, 28.4–61.9 mm SL, municipality of Iporanga, SP, Rio Ribeira de Iguape basin. Neoplecostomus microps: LBP 8036, 38, 41.3–65.0 mm SL, municipality of Piquete, SP, Rio Paraíba do Sul basin. Neoplecostomus franciscoensis: LBP 6489, 50, 42.8–55.9 mm SL, municipality of São Bartolomeu, MG, Rio São Francisco basin. Neoplecostomus paranensis: holotype, MZUSP 38572, 71.4 mm SL, municipality of Cajuru, MG, Rio Grande basin. Pareiorhaphis splendens: LBP 1117, 20, 32.0–100.0 mm SL, municipality of Morretes, PR, Coastal Drainage. Pareiorhaphis steindachneri: LBP 739, 6, 33.8–49.0 mm SL, municipality of Jaraguá do Sul, SC, Coastal Drainage. Pareiorhina brachyrhyncha: LBP 12240, 50, 26.4–36.9 mm SL, municipality of Pindamonhangaba, SP, Rio Paraíba do Sul basin. Pareiorhina carrancas: LBP 8380, 24, 21.3–38.2 mm SL, municipality of Carrancas, MG, Rio Grande basin. Pareiorhina cepta: holotype, MZUSP 111095, 41.5 mm SL, municipality of São Roque de Minas, MG, Rio São Francisco basin, paratypes, LBP 10261, 1, 30.2 mm SL, municipality of São Roque de Minas, MG, Rio Paraíba do Sul basin, LBP 10287, 13, 21.5–43.6 mm SL, municipality of São Roque de Minas, MG, Rio Paraíba do Sul basin, LBP 11835, 19, 25.1–44.0 mm SL, municipality of São Roque de Minas, MG, Rio Paraíba do Sul basin. Pareiorhina rudolphi: LBP 8044, 18, 31.7–48.9 mm SL, municipality of Piquete, SP, Rio Paraíba do Sul basin. Pseudotocinclus juquiae: LBP1081, 2, 29.0–31.9 mm SL, municipality of Juquitiba, SP, Coastal Drainage. Pseudotocinclus tietensis: LBP 2931, 3, 38.6–62.3 mm SL, municipality of Salesópolis, SP, Rio Tietê basin.
The authors thank Anderson L. Alves, Alex T. Ferreira, José C. Oliveira, Luciana R. Sato, Guilherme J. C. Silva, and Tiago N. A. Pereira for helping with the fish sampling of fishes; Mahmoud Mehanna for help with the etymology; and Edson H. L. Pereira for reading the manuscript and providing valuable suggestions. This research was supported by the Brazilian agencies FAPESP (Fundação de Amparo à Pesquisa do Estado de São Paulo; proc. 2012/01622-2 to GSCS, and proc. 2010/01610-9 to FFR) and MCT/CNPq (Conselho Nacional de Desenvolvimento Científico e Tecnológico).