





(C) 2013 Olga L. Makarova. This is an open access article distributed under the terms of the Creative Commons Attribution License 3.0 (CC-BY), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
For reference, use of the paginated PDF or printed version of this article is recommended.
Citation: Makarova OL, Lindquist EE (2013) A new species of the gamasid mite genus Arctoseius Thor, 1930 (Parasitiformes, Mesostigmata, Ascidae) from Russia with a key to the multidentatus species-group. ZooKeys 313: 9–24. doi: 10.3897/zookeys.313.5317
A new gamasid mite species belonging to the genus Arctoseius Thor, 1930 is described from Russia. Arctoseius koltschaki sp. n. is distributed in the plain and mountain tundras from Khibiny Mountains to Chukotka on the north and to West Sayan Mountains on the south. A diagnosis and a key for identification of species comprising the multidentatus species-group (Arctoseius multidentatus Evans, 1955; Arctoseius wisniewskii Gwiazdowicz & Kamczyc, 2009; Arctoseius sexsetus Lindquist & Makarova, 2011; Arctoseius haarlovi Lindquist & Makarova, 2011; and Arctoseius koltschaki sp. n.) are given.
Gamasid mites, Arctic, Arctoseius koltschaki sp. n., multidentatus species-group, identification key
Arctic landscapes are populated by at least 26 mite species belonging to the genus Arctoseius Thor, 1930, which constitutes about half of the total diversity of this genus (
In total 59 females, 23 males and 19 nymphs were found in 25 localities. Setal notation for the idiosoma follows
lD/wD length-to-width ratio of the dorsal shield;
J(1–4)min/jmax ratio of the length of dorsocentral setae inserted on the opisthonotal and podonotal regions, notably the shortest among J1–4 and the longest in the series j;
(J4–J4)/(J3–J3) ratio of transverse distances between bases of opisthonotal setae J4 and J3 in pairs;
Z5/(Z5–Z5) ratio of setae Z5 length to the distance between them;
lSt/wSt, lAn/wAn, lVA/wVA length-to-width ratios of the sternal, anal, and ventrianal (in males) shields, respectively;
lCo/lD ratio of the length of the corniculus and of the dorsal shield, %;
lCh/lD ratio of the length of the chela and of the dorsal shield, %;
lCh/lCo ratio of the length of the chela and of the corniculus;
lExI/lD, lTIV/lD ratio of the length of leg I or tarsus IV and of the dorsal shield;
lTIII/wTIII length-to-width ratio of tarsus III, width taken at basistarsus.
In the male description, the features common with the female are omitted.
Holotype and most of paratypes are deposited in Zoological Institute, Russian Academy of Sciences, Saint Petersburg (ZIRAS). Part of paratypes, 5 females, 1 male, 2 N2, from Suntar-Khayata Range, 1500 m a.s.l. (above sea level), spotty tundra (on one slide) – in Canadian National Collection of Insects, Arachnids and Nematodes, Ottawa (CNC).
urn:lsid:zoobank.org:act:25C72458-8447-4610-BCA1-EC2EBC33D2A6
http://species-id.net/wiki/Arctoseius_koltschaki
Figs 1–25Holotype, female: EAST SIBERIA, YAKUTIYA, Khalerchinskaya Tundra, Chukochiya River, mosses in shrub tundra, IX 1975, leg. E.V. Gordeyeva (ZIRAS).
Paratypes: 5 females, 1 male, 2 N2, EAST SIBERIA, Suntar-Khayata Range, upper reaches of Kyubyume River, 63°13'N, 139°36'E, 1500 m a.s.l., spotty tundra, 29.VII 2002, leg. O.L. Makarova (CNC); 1 male, same district and collector, 1960 m a.s.l., snow bed near small stream, sedge turf, 1.VIII 2002; 1 female, delta of Indigirka River, Russko-Ustinskaya Protoka, hummocky Dryas-forb tundra, 14–16.VII 1994, leg. A.B. Babenko; NOVAYA ZEMLYA ARCHIPELAGO, 1 male, Southern Island, Pan’kova Zemlya, lower part of slope, sedge-willow-moss tundra, VIII 1995, leg. S.V. Goryachkin; 1 female, 1 male, Northern Island, Krestovaya Bay, tundra, VIII 1993, leg. V.I. Bulavintsev; 1 female, BOL’SHEZEMEL’SKAYA TUNDRA, Vorkuta Region, Sivaya Maska vicinity, tundra, 20.VII 1978, leg. N.A. Kuznetsova; 3 females, 1 male, KOLA PENINSULA, Khibiny Mountains, Eastern Petrelius Pass, Dryas-sedge-moss tundra, 28.VI 2006, leg. A.B. Babenko (ZIRAS).
Other material: NORTH-EAST ASIA, 5 females, 1 male, Chukotka, Daurkin Peninsula, lower reaches of Chegitun River, forb-sedge-moss tundra, 30.VIII 1996, leg. M.V. Berezin; 5 females, 3 males, Magadan Region, upper reaches of Kolyma River, Peak Aborigen vicinity, Pinus pumila litter, 24.VIII 2006, leg. A.V. Alfimov; 1 female, Olskoye Plateau, upper reaches of Ola River, 60°39'N, 151°16'E, 1220 m a.s.l., forb meadow, 11.VIII 2011, O.L. Makarova; EAST SIBERIA, delta of Kolyma River, Pokhodskaya Edoma, 1 female, litter in lower part of slope; 1 female, wet grassy meadow, 18.VII 1994, leg. A.B. Babenko; delta of Yana River, Shirokostan Peninsula, vicinity of Ledyanoye Lake, 72°25'N, 141°00'E, 1 female, south slope of valley, forb-grassy meadow; 1 male, high river terrace, Dryas-Carex tundra, 4–6.VIII 1994, leg. A.B. Babenko; 3 females, 1 N1, Yana Bay, Makar Island, 6.VIII 1985, leg. V.I. Bulavintsev; Suntar-Khayata Range, upper reaches of Kyubyume River, 3 males, 1 N2, 1300 m a.s.l., lichen Larix-forest, litter, 25.VII 2002, 2 females, 1 male, 1 N1, 1800 m a.s.l., snow bed, under Rhododendron aureum, 2 males, 5 N2, 2 N1, 1800 m a.s.l., hummocky Eriophorum-community, 28.VII 2002, 1 female, 1 N2, 1800 m a.s.l., lichen-willow tundra, 9.VII 2002, 1 female, 1 N2, 1 N1, 2000 m a.s.l., mossy tundra, 15.VII 2002, 4 females, 3 males, 2 N2, 1 N1, Dryas-Carex-Eriophorum bog, 19.VII 2002, leg. O.L. Makarova; 1 female, Cherskogo Range, Ust-Nera vicinity, 500 m a.s.l., grassy meadow near stream, 22.VII 1992, leg. N.A. Kuznetsova; TAYMYR PENINSULA, 1 male, N Taymyr, Khariton Laptev Coast, Opalovaya River, seaside sand-rubble terrace with Salix polaris, 15–17.VIII 1994, leg. A.B. Babenko; Taymyr Lake, Cape Blizhnyi, 2 females, Tetraplodon moss cushion in tundra, 28.VII 1994, 1 female, lemming hill within stony tundra on elevation, grassy turf, 10.VIII 1994, 2 females, lemming burrow, 17.VIII 1994, 5 females, 1 N1, lemming hill within spotty tundra, grassy turf, 17.VIII 1994, leg. O.L. Makarova; 1 female, NW Taymyr, upper reaches of Kolomejtseva River, lemming hill in spotty tundra, grassy turf, 21.VIII 1997, leg. O.L. Makarova; 2 females, 1 male, NW Taymyr, Ragozinka River, spotty tundra, 9.VII 1986, leg. A.B. Babenko; 1 female, NW Taymyr, mouth of the Tareya River, 73°15'N, 90°35'E, dry Dryas-community, 22.VII 2010, O.L. Makarova; 1 female, SW Taymyr, vicinity of Pyasino Lake, Nyapan’ Upland, zoogenic meadow on a hill, 16.VII 1999, O.L. Makarova; VAIGATCH ISLAND, 1 male, no other data, VII 1984, leg. V.I. Bulavintsev; NOVAYA ZEMLYA ARCHIPELAGO, 1 male, no other data, leg. V.I. Bulavintsev; BOL’SHEZEMEL’SKAYA TUNDRA, 3 females, Yugor Peninsula, Cape Belyi Nos, 25.VI 1983, leg. V.I. Bulavintsev; 1 female, Vorkuta city vicinity, IX 2009, leg. E.M. Perminova; 1 female, Pechora Bay, Kuznetskoye Lake, polar fox hill, litter under Cornus suecica, 25.VIII 1994, leg. A.B. Babenko; KOLGUEV ISLAND, 3 females, 1 male, 2 N2, 1 N1, tundra litter, VII 2011, leg. S.B. Rosenfeld; SOUTH SIBERIA, WEST SAYAN MOUNTAINS, 1 female, 50°23'N, 90°26'E, Tsagan-Shibetu Range, Mugur-Aksy vicinity, 2800 m a.s.l., tundra, litter, 22.VII 1993, leg. S.K. Stebaeva.
Adults of middle size, yellowish or brownish, with idiosoma rather narrow and appendages of normal proportions. Idiosomal shields moderately sclerotized, very finely punctate (punctation rarely visible), with clearly reticulate ornamentation only on sternal, genital or sternitigenital shields; dorsal shield smooth with distinct sigillae. Most body setae of moderate size, needle-shaped. Some setae of distal leg segments, especially of leg IV, elongated and finely tipped, subapical setae av-1 and pv-1 on tarsi II-IV strongly formed and blunt.
Female. Idiosomal dorsum. Dorsal shield 528-616 × 248-304, narrowing posteriad, lD/wD ca 1.96–2.24, its maximal width at level of setae s4 (Fig. 1); lateral incisions of moderate length (30–44). Podonotal region normally with 17 pairs of simple setae (z3 present); z1 and s1 seldom asymmetrically absent; s2 usually on soft cuticle, sometimes asymmetrically on shield margin, sometimes symmetrically absent; s6 rarely off shield. Opisthonotal region with 13–14 pairs of setae (S2 always absent, S3 or S5 rarely asymmetrically off shield margin). Among podonotal setae, z1 (13–22) and s1 (20–35) distinguished by clearly shorter lengths, seta j1 30–36, j2 32–42, length of other setae 30–50. On opisthonotal region, setae J1-4 (25–35) slightly shorter than others (30–44), except J5 clearly shortest (9–17) and Z5 clearly longest (48–63); J(1–4)min/jmax 0.65–0.80, Z5/(Z5-Z5) 0.76–1.26; (J4-J4)/(J3-J3) 1.31–2.00. Dorsal shield with 5 pairs of gland pores: gdj4, gdj6, gdz5, gdz6, all poorly visible and gdZ3 distinct. All marginal setae on soft cuticle (their length 28–44, only r3 39–48); 4 (rarely 3) setae in series r, 5 setae in series R; marginal poroid Rp in usual position between setae R3 and R4.
Idiosomal venter. Base of tritosternum narrow (20–35 × 16–22); laciniae with sparse large barbs, free for ca 0.8–0.9 of lengths, their fused basal area fimbriated anteriorly (Fig. 20); rarely barbs are hardly separated (Fig. 21); length of laciniae free part 60–80. Presternal platelets large, consolidated with each other and sternal shield, lineate and clearly punctate (Fig. 6). Sternal shield commonly longer (146–164) than wide (124–160), lSt/wSt 1.00–1.23, merged with endopodal platelets between coxae II-III. Endopodal projections between coxae I-II small, nearly bacilliform, commonly separated, their lateral part encompassing opening of gland gvb. Sternal shield entirely reticulated; anterior margin straight or slightly concave, posterior margin straight. Sternal shield with typical setae st1-3 (32–44), lyrifissures iv1-3, and with vestiges of gland gv1 on posterior margin. Setae st4 (24–34) on soft cuticle. Endopodal strips between coxae III and IV free, rather narrow, partly hidden under epigynal flap. Epigynal shield (140–164 × 72–82) distinctly reticulated (Fig. 9), broadly axe- or flask-shaped, with evenly convex hyaline flap well distant from sternal shield, and posterior margin broadly convex; lateral margins widening behind level of setae st5, but st5 (26–36) and paragenital poroids iv5 remain on soft cuticle. Opening of gland gv2 on soft cuticle close beside end of peritrematal-exopodal strip behind coxa IV. Two pairs of postgenital platelets in fold of soft cuticle adjacent to posterior margin of epigynal shield, another pair between setae JV1 and ZV1. Anal shield rather small, ovate, weakly reticulated (Fig. 10), longer (94–112) than wide (76–93), lAn/wAn 1.08–1.30; paranal setae (36–46) variably inserted, usually between mid-level of anus and its posterior margin, and often nearly as long as postanal seta (42–64); opening of gland gv3 inconspicuous; cribrum ordinary. Opisthogastral setae (8 pairs, JV1–5, ZV1, 2, 4) of moderate length (most setae 30–40), JV5 longest (35–52), ZV1 shortest (20–34). Anterior metapodal platelets small, of variable form; posterior metapodal platelets elongate, often bacilliform (24–39 × 4–7). Exopodal platelets between coxae II and III, as well as between coxae III and IV, small, triangular, rarely visible in ventral aspect. Peritrematal shield reduced, of uneven width, with angular extension at level of coxa I; its anterior end abutting dorsal shield; its posterior edge connecting with exopodal strip enveloping coxa IV posteriorly; lyrifissures ip1–3 and glands gp1, 2 present. Peritreme shortened (128–156 × 8–11), extending to mid-level of coxa II anteriorly. Spermathecal apparatus seldom visible, with very narrow tubuli, thin-walled sperm receptacle and sperm duct strongly expanded medially (Fig. 8).
Gnathosoma. Gnathotectum (28–39 × 40–56) triramous, with projections of almost equal length, sometimes divided apically (Figs 23, 24); middle process sometimes narrower basally than lateral ones. Subcapitulum always slightly longer (116–128) than wide (104–120) (Fig. 7). Deutosternum with 7 (rarely 8) rows of denticles (15–22 denticles in each row); groove delineated laterally, its width 15–19 (Fig. 7). Hypostomatic pair hp3 (46–58) longer than other subcapitular setae (hp1 31–40, sc26–42, hp2 18–26); all setae simple, attenuate. Corniculi somewhat elongated, 46–53 × 15–19; lCo/lD 7.66–9.56. Internal malae slightly shorter than corniculi, gradually tapering to tip, with lateral margins roughly fimbriated basally. Chelicera medium-sized, length of middle article 132–156; movable digit moderate sized (50–56, lCh/lD 8.65–9.71), slightly longer than corniculus (lCh/lCo 1.03–1.17). Fixed digit of chela ending in apical trident, masticatory surface with a row of 5–6 teeth in paraxial position and pilus dentilis in antaxial position (Figs 2–4). Movable digit bidentate. Palp length 180–204; internal seta of trochanter (39–43) slightly longer than external seta (34–38); palpi with typically specialized setae on femur (al) and genu (al1, al2) large, thick, with oblique tip.
Legs. Legs of moderate length (I 512–624, II 432–480, III 352–424, IV 512–592); leg I similar in length to dorsal shield, lExI/lD 0.91–1.09. Length of tarsi I 152–180, II 108–134, III 96–132, IV 160–192; tarsi of normal proportions: lTIV/lD 0.28–0.34, lTIII/wTIII 3.25–4.10. Leg II significantly thicker than others (width of genu II 56–68, genu I 36–40, genua III, IV 44–48). Leg chaetome in general as described for genus (
Male. Idiosomal dorsum. Dorsal shield 446–552 × 228–260, more narrowed posteriad than in female (lD/wD ca 2.05–2.17); lateral incisions on average shorter than in female (20-40); chaetome in general as in female, but setae commonly slightly shorter (by 5–15 %); 9 pairs of marginal setae (r2-r5, R1-R5) on soft cuticle; s2 usually on soft cuticle, rarely on shield margin or absent.
Idiosomal venter. Tritosternum base (16–20 × 12–16) slightly smaller than in female (Fig. 17). Presternal platelets connected with sternitigenital shield. Sternitigenital shield united with endopodal platelets developed between coxae II-III, partly separated from ones situated between coxae III-IV and only abut ones developed between coxae I-II; its posterior margin slightly convex or truncate; length of shield 212–248, width 124–140, its narrowest part (44–48) between coxae IV. Sternitigenital shield without reticulation in the central and posterior areas, with setae st1–3 (30–38) longer than setae st4, 5 (20–28) and distinct lyrifissures iv1–3; vestiges of glands gv1 not discernible. Ventrianal shield well separated from sternitigenital and peritrematal shields, subtriangular, often with small anterior ledge, fully reticulated, its length 188–224, width 184–240, lVa/wVa 0.83–1.09, normally consolidated with metapodal platelet sigillae laterally; its anterolateral margins evenly convex, cribrum formed as in female. Ventrianal shield with 6 pairs of opisthogastral setae (JV1–4, ZV1, 2), 2–3 pairs of poroids and gland opening gv3, sometimes ZV1 asymmetrically absent, JV4 rarely asymmetrically off shield margin; ZV1, 2 (20–28) somewhat shorter than others (26–35), paranals 32–42, postanal seta 40–52; seta JV5 (34–40) on soft cuticle.
Gnathosoma. Gnathotectum as in female. Corniculi slightly thinner and more parallel than in female (Fig. 5). Internal malae unlike female finely and densely fimbriate. Cheliceral digits 46–52 (lCh/lD 8.58–10.94), longer than corniculus (42–47 × 12–15, lCh/lCo 1.02–1.30, lCo/lD 7.52–9.40). Fixed digit with dentition similar to female, but apical trident not developed and one denticle present subapically in antaxial position, apart from paraxial row of 5–6 denticles. Movable digit with one large denticle and tightly (at a sharp angle) sinuate spermatodactyl (length of free part 26–29), half protruding beyond tip of digit (Fig. 18).
Legs. Length of legs I to IV 492–596 (lExI/lD 0.96–1.13), 364–432, 336–416, and 472–568 respectively; length of tarsi I to IV 140–168, 96–124, 104–122, and 140–170 respectively. Leg II stouter than other legs, with dimorphically modified setae. Three setae on leg II, namely v2 on femur, av2 and mv on telotarsus, remarkably stout basally and attenuate apically, on elevated bases; opposable ventral setae v-1 on femur II and av on genu II and tibia II also usually slightly spinelike (Fig. 22). Other leg structure and setation generally as in female.
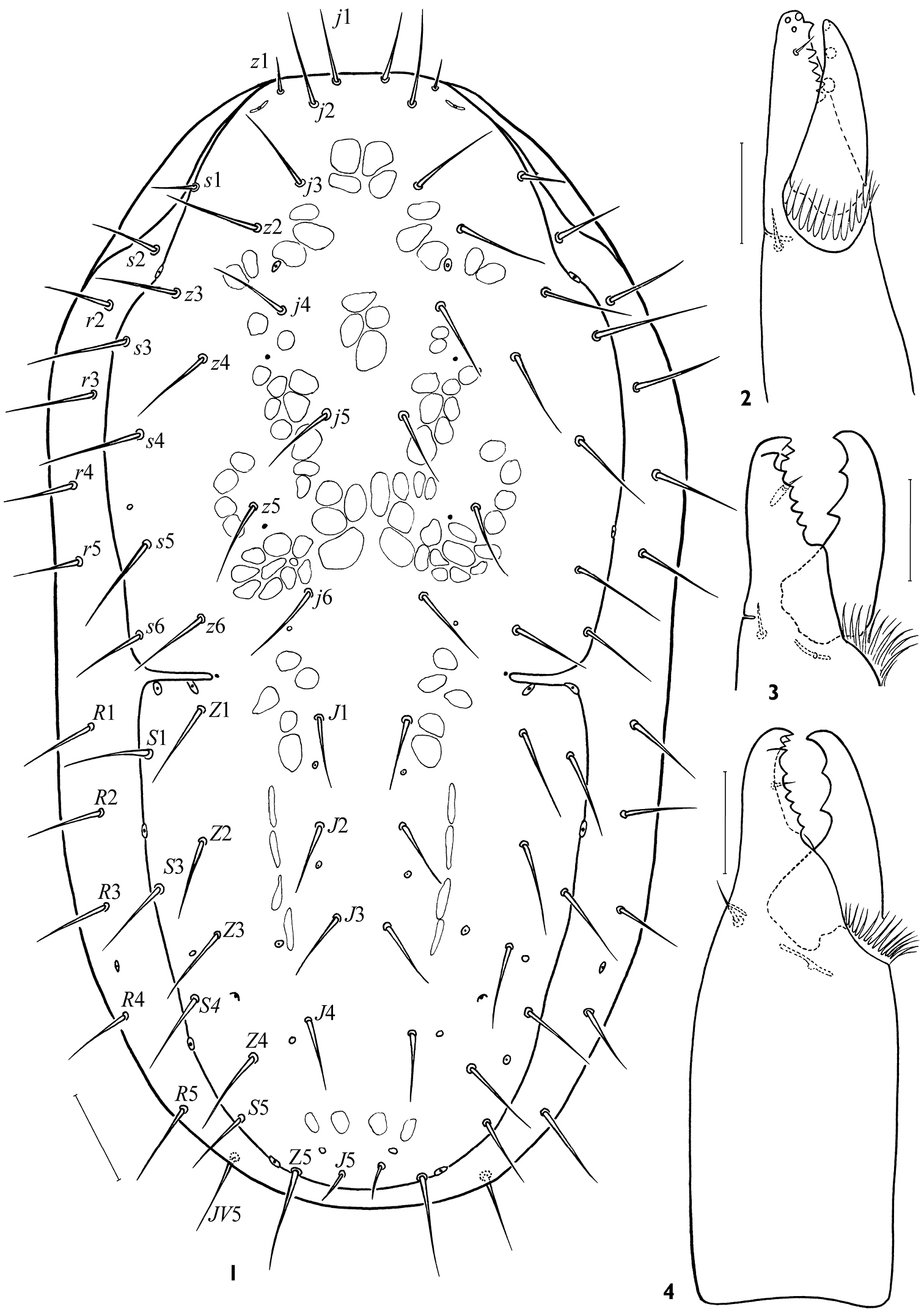
Arctoseius koltschaki sp. n., female 1 idiosomal dorsum 2–4 cheliceral digits. Scales: 1 – 50 μm, 2–4 – 25 μm.
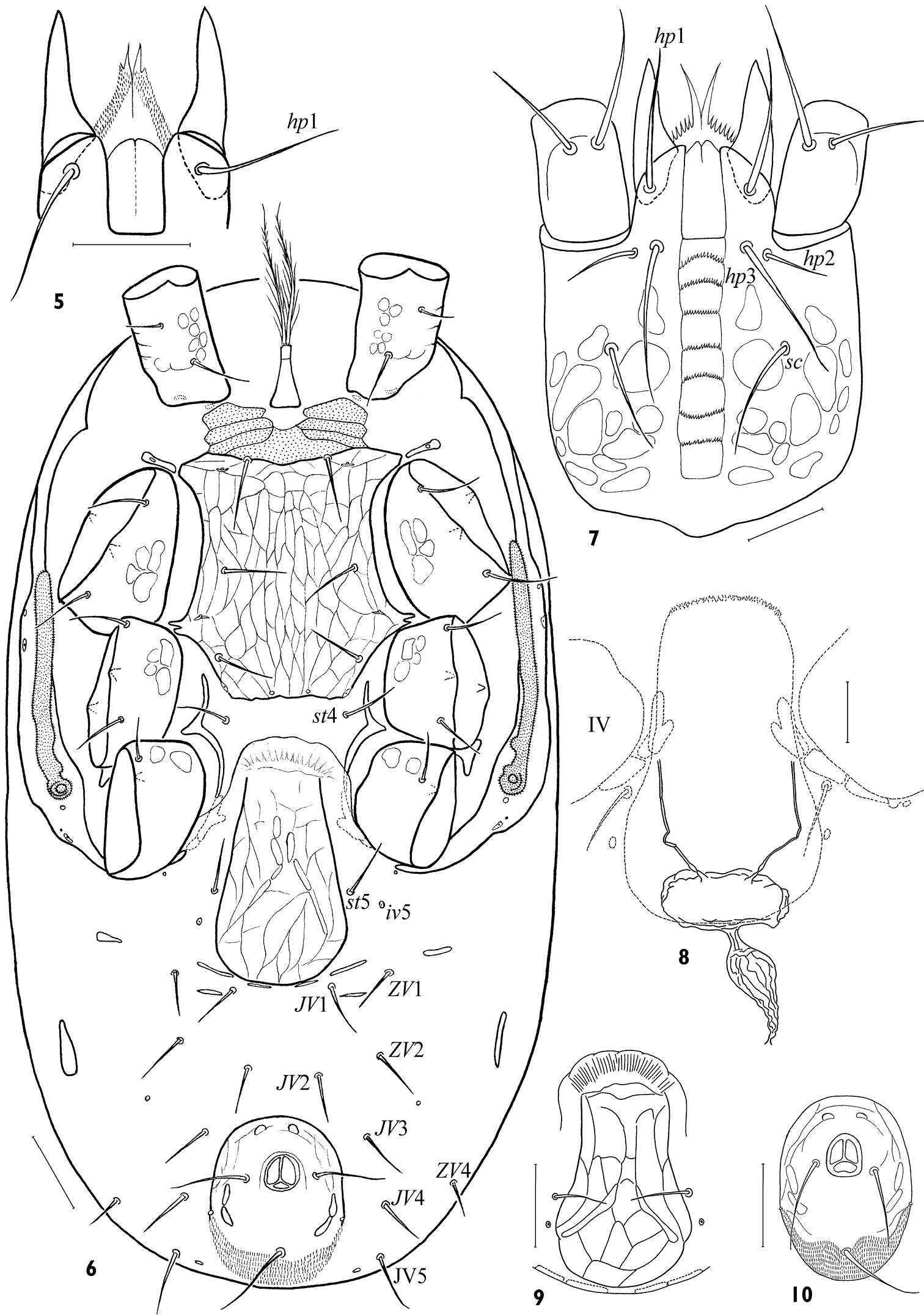
Arctoseius koltschaki sp. n., male (5) and female (6–10) 5 hypostome 6 idiosomal venter 7 subcapitulum 8 inner part of spermathecal apparatus 9 variant of genital shield form 10 variant of anal shield form. Scales: 5, 7, 8 – 25 μm, 6, 9, 10 – 50 μm.
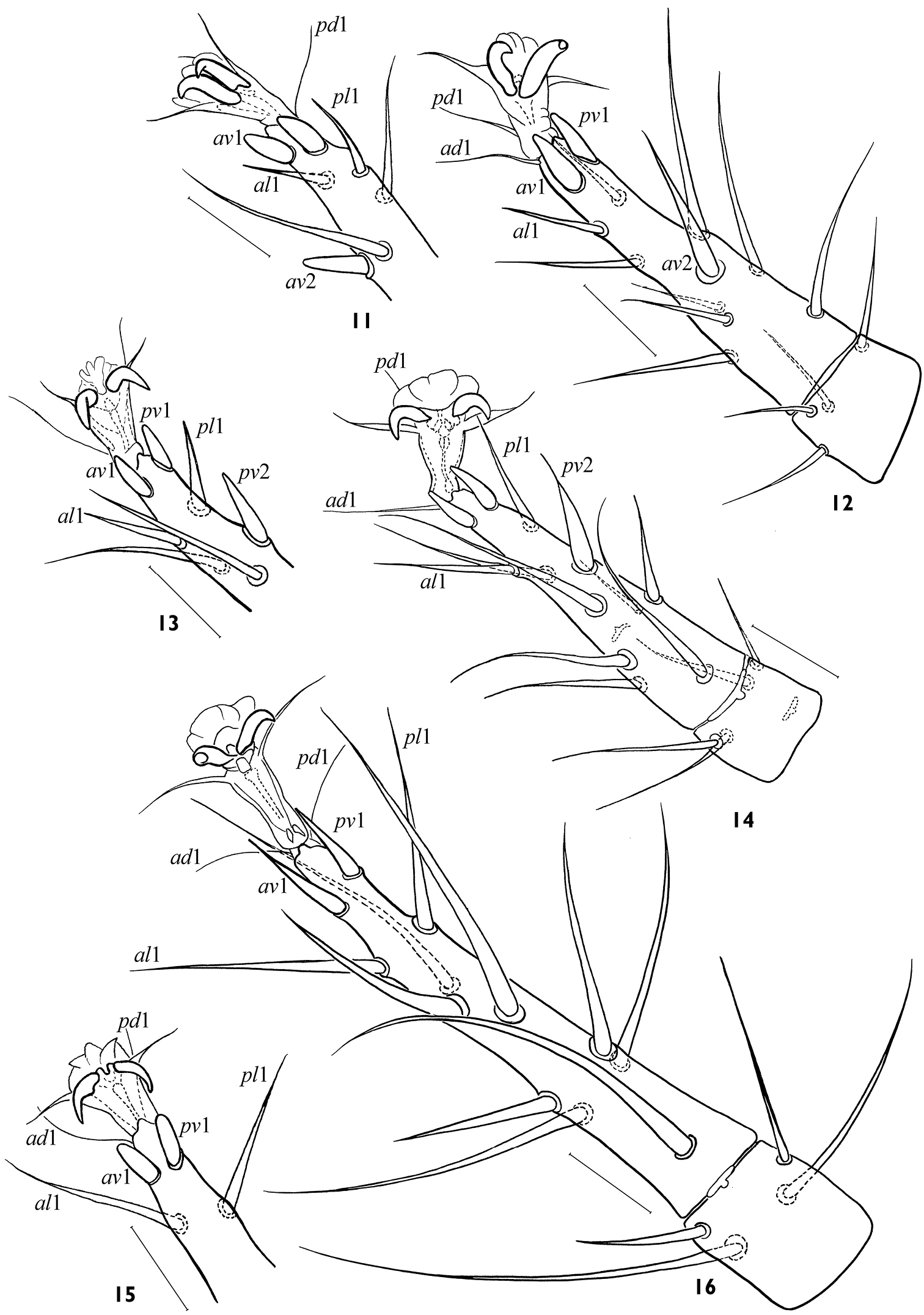
Arctoseius koltschaki sp. n., female 11, 12 variants of subapical setation of tarsus II 13, 14 variants of subapical setation of tarsus III 15, 16 variants of subapical setation of tarsus IV. Scales 25 μm.
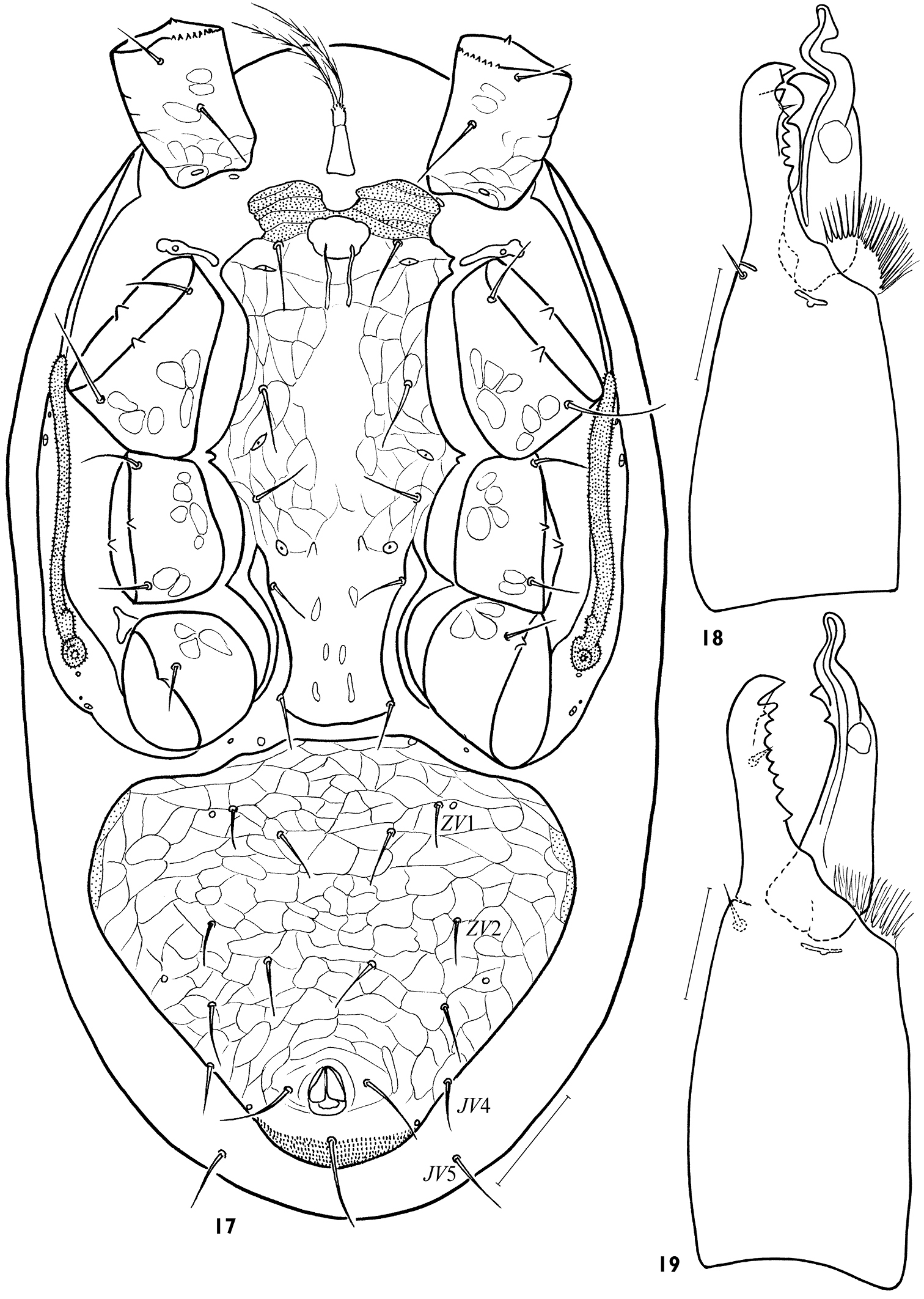
Males, Arctoseius koltschaki sp. n. (17, 18) and Arctoseius multidentatus Evans, 1955 (19) 17 idiosomal venter 18, 19 chelicera. Scales: 17 – 50 μm, 18, 19 – 25 μm.
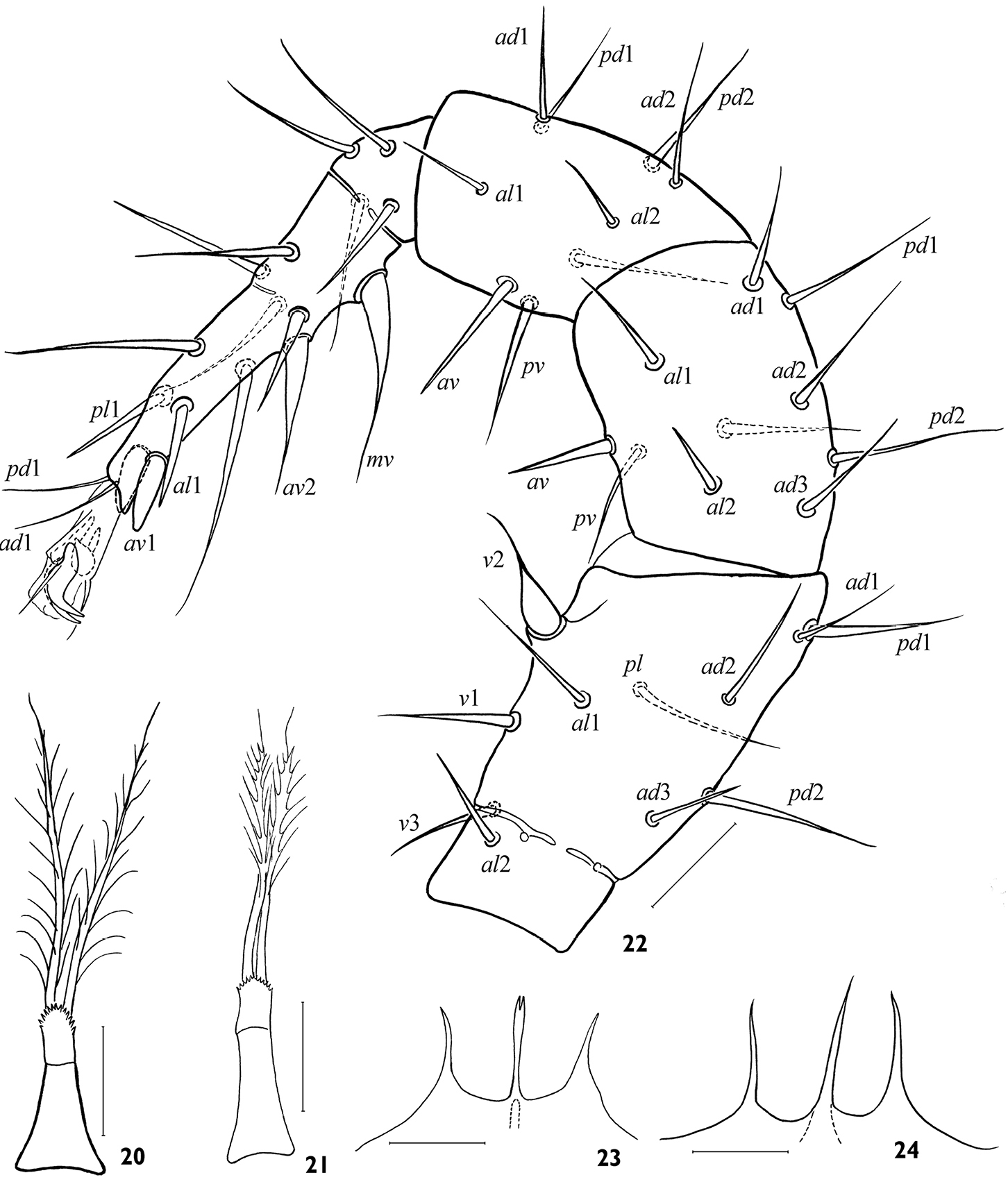
Arctoseius koltschaki sp. n., female (20, 21, 23, 24) and male (22) 20, 21 tritosternum 22 leg II 23, 24 gnathotectum. Scales 25 μm.
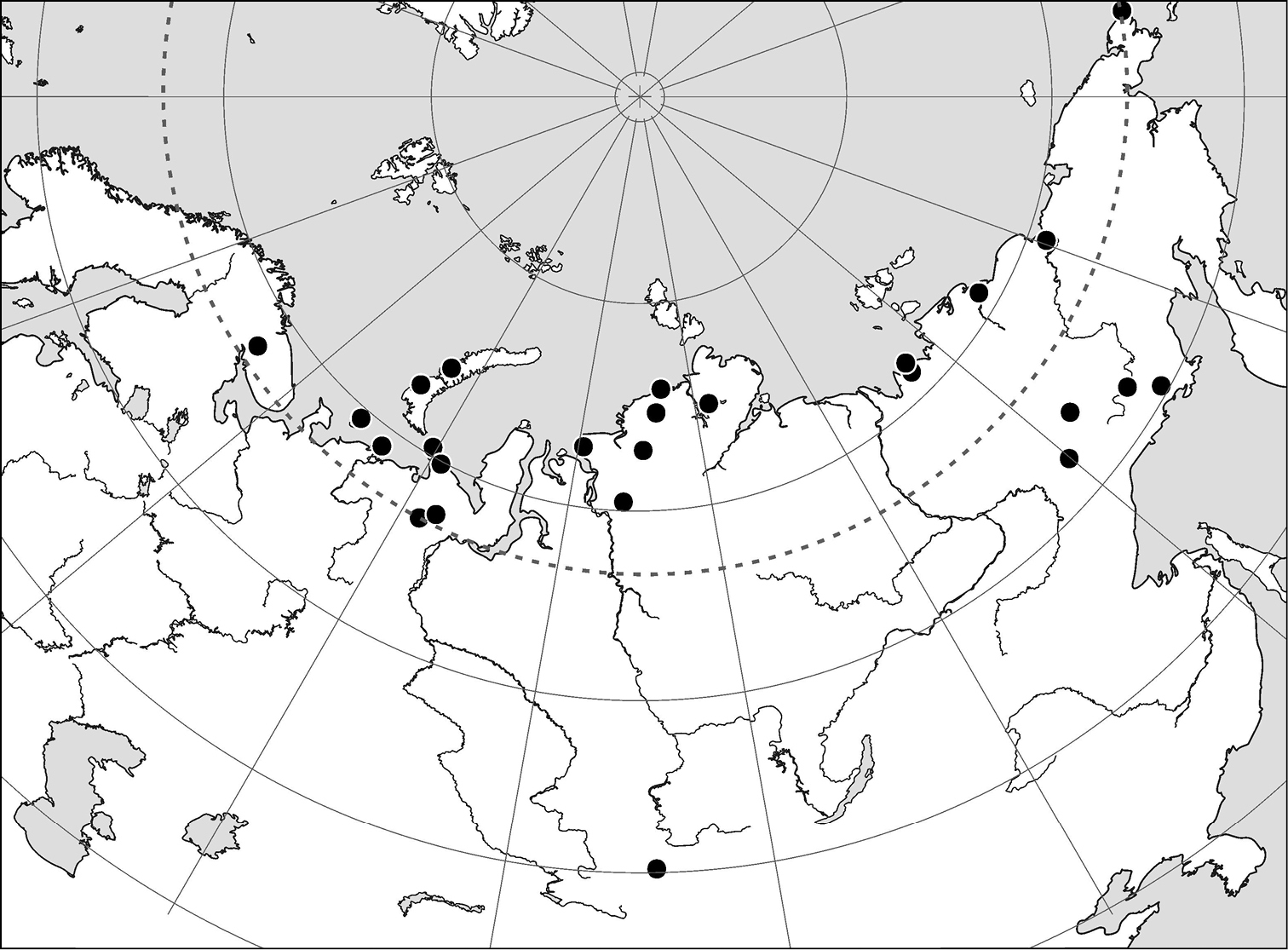
Records of Arctoseius koltschaki sp. n.
Adults of Arctoseius koltschaki sp. n. are similar to those of Arctoseius multidentatus Evans, 1955, but the dorsal shield in Arctoseius koltschaki sp. n. is often narrower (lD/wD in female 1.96–2.24, in male 2.05–2.17 instead of 1.91–2.05 and 1.85–2.03 respectively), and tarsi II-IV have subapical setae av1 and pv1 more stout and blunt (Figs 11–15). In male of Arctoseius koltschaki sp. n., the ventrianal shield is relatively wide (Fig. 17), subtriangular (narrowly oval in Arctoseius multidentatus), incorporating the metapodal platelets (free in Arctoseius multidentatus); the spermatodactyl is more tightly or sharply sinuate, and more angled apically (not as gently sinuate, and digitiform apically as in Arctoseius multidentatus) (cf. Figs 18 and 19). In female of Arctoseius koltschaki sp. n., the sternal shield is uniformly reticulated (whereas in Arctoseius multidentatus, the reticulation is developed laterally, and separately within an elongate posteromedial “window”). Female of Arctoseius koltschaki sp. n. is often smaller (dorsal shield length 528–616 instead of 566–688 in Arctoseius multidentatus).
The species named in honor of Admiral Alexander Koltschak, the prominent Arctic hydrologist and outstanding Citizen of Russia.
At present known from the Russian Arctic and Siberian mountain regions (Fig. 25).
Recorded from dry and humid tundra sites, various meadows including zoogenic ones (e.g. polar fox and lemming hills), snow-beds, bogs, larch forests and shrub (Salix spp., Rhododendron aureum, Pinus pumila) thickets.
The species is rather stable morphologically. The form of female epigynal and anal shields varies a little (Figs 6, 9, 10). The variability concerns mainly the form of tectum (Figs 23, 24), the position of setae s2, s6, S3, S5 (on or off shield margin), and the form and size of leg subapical setae (Figs 11–16). The robust subapical setae av1 and pv1 sometimes may be acuminate, more often on leg IV (Fig. 16). The deutosternum rarely has 8 (instead normal 7) rows of denticles. The lacinia of trito-sternum seldom has adnate barbs (Fig. 21). One male (from 23 ones) originating from Chukotka has free baculiform metapodal platelets (26 × 4).
Previously and provisionally, there were some indications about the presence of new species close to Arctoseius multidentatus Evans, 1955, namely Arctoseius sp. aff. multidentatus [Arctoseius koltschaki sp. n.], «Arctoseius haarlovi Lindquist, 1963 ms.» (
Idiosoma of moderate size (388–688 in females, 348–552 in males); dorsal shield rather narrow (lD/wD ca 1.9–2.6), not covering entire idiosoma, smooth, with distinct sigillae. Presternal platelets weakly sclerotized, lineate and punctate, fused with sternal shield. Sternal/sternitigenital shield often free from endopodal strips between coxae I–II (excluding Arctoseius sexsetus, Arctoseius wisniewskii), but united with them between coxae II–III. In females, anal shields small, usually not shorter than wide (lAn/wAn 0.85–1.40). In males, ventrianal shields with convex anterior margin, oval or roundish, leaving the metapodal platelets free on soft cuticle (exception – Arctoseius koltschaki sp. n. with wider, subtriangular ventrianal shield incorporating metapodal platelets). In both sexes, circumanal setae of similar lengths, with paranal setae relatively long, at least 0.7 as long as postanal seta. In both sexes, peritrematal shield reduced, of uneven width, with angulate extension between coxae I-II; peritreme shortened, extending anteriorly at most to mid-level of coxa II. Gnathotectum with three projections, these sometimes bifurcate apically. Corniculi rather long (lCo/lD ca 7–13), shorter or longer than cheliceral digits (lCh/lCo 0.74–1.44). In male, spermatodactyl straight or sinuate; leg II sometimes with dimorphically modified setae. Legs moderately long, leg I similar in length with dorsal shield.
The relatively long paranal setae are considered to be apomorphic for this species-group within the genus Arctoseius. The reduced peritremes and peritrematal shields, somewhat elongated corniculi, and complete lack of dorsal shield ornamentation (retention of deutonymphal condition) are also apomorphic, but are derived independently among some other Arctoseius species.
| 1 | Podonotal seta z3 and opisthonotal seta S1 absent; corniculi large (lCo/lD 11.52–13.41), longer than cheliceral digits (lCh/lCo 0.74–0.93); fixed cheliceral digit with at most 4 denticles besides terminal hook; tibia IV with 6 setae, pl absent; in male, dimorphically modified setae on leg II absent, spermatodactyl straight | 2 |
| – | Podonotal seta z3 and opisthonotal seta S1 present (Fig. 1); corniculi smaller (lCo/lD 6.90–10.98), shorter than cheliceral digits (lCh/lCo 1.02–1.44); fixed cheliceral digit polydont, with 6–14 denticles besides terminal hook (Figs 4, 18); tibia IV with 7 setae, pl present; in male, dimorphically modified setae on leg II well developed (Fig. 22), spermatodactyl clearly sinuate (Figs 18, 19) | 3 |
| 2 | Fixed cheliceral digit terminating with trident, behind which one denticle present; in male, single denticle of movable digit situated in subapical position. Female 444–552 μm, male 404–464 μm. Alaska, Eurasian Arctic, Siberian mountains | Arctoseius sexsetus Lindquist & Makarova, 2011 |
| – | Tip of fixed cheliceral digit of usual form, with three denticles forming a row behind apical hook; in male, single denticle of movable digit situated at a greater distance from apex, almost opposite pilus dentilis of fixed digit. Female 550 μm, male 500 μm, Stolovye Mountains, Poland | Arctoseius wisniewskii Gwiazdowicz & Kamczyc, 2009 |
| 3 | Smaller mites, dorsal shield length at most in female 465 μm, in male 420 μm; opisthonotal region with 13 pairs of setae (only one pair of setae present in area usually occupied by J4 and Z4); peritreme much reduced, usually not extending beyond mid-level of coxa III anteriorly. Female 388–465 μm, male 348–420 μm. Circumpolar | Arctoseius haarlovi Lindquist & Makarova, 2011 |
| – | Larger mites, dorsal shield length at least in female 528 μm, in male 446 μm; opisthonotal region with 14 pairs of setae (J1–5, Z1–5, S1, 3–5); peritreme terminating about mid-level of coxa II anteriorly (Figs 6, 17) | 4 |
| 4 | Tarsi II-IV with subapical setae av-1 and pv-1 stout and blunt (Figs 11–15); in male, ventrianal shield relatively wide, subtriangular, incorporating metapodal platelets (Fig. 17); spermatodactyl tightly sinuated, and angled apically (Fig. 18). Female 528–616 μm, male 446–552 μm. Eurasian Arctic and Metaarctic, South Siberia (West Sayan Mountains) | Arctoseius koltschaki sp. n. |
| – | Tarsi II-IV with subapical setae av-1 and pv-1 needle-shaped, but thicker than pl-1, al-1; in male, ventrianal shield usually narrow, often oval, leaving metapodal platelets free on soft cuticle; spermatodactyl more gently sinuated, and either simple or bluntly angled apically (Fig. 19). Female 566–688 μm, male 488–550 μm. Circumpolar | Arctoseius multidentatus Evans, 1955 |
Many friends and colleagues supplied material from which additional specimens of this new species were found – A.V. Alfimov, A.B. Babenko, M.V. Berezin, D.I. Berman, V.I. Bulavintsev, E.V. Gordeyeva, S.V. Goryachkin, N.A. Kuznetsova, E.M. Perminova, M.B. Potapov, S.B. Rosenfeld, S.K. Stebaeva. The Programs “Problems of Life Origin and Biosphere Formation” and “The Wildlife” of the Russian Academy of Sciences as well as the Russian Fond of Basic Researches supported the first author during certain aspects of this investigation, including field work. Arrangement of figures and the map were prepared for publication with thankful participation of K.V. Makarov. We are very grateful to all.