





(C) 2012 Rolando Bastida-Zavala. This is an open access article distributed under the terms of the Creative Commons Attribution License 3.0 (CC-BY), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
For reference, use of the paginated PDF or printed version of this article is recommended.
The exotic Indo-West-Pacific species, Ficopomatus uschakovi (Polychaeta, Serpulidae) is recorded for the first time in the Tropical Eastern Pacific from two sites in La Encrucijada Biosphere Reserve, Chiapas, a coastal lagoon in the Pacific south of Mexico. The means of dispersal of this serpulid species still remains unclear, as the nearest port (Puerto Chiapas) is 70 km to the south, and there are no port installations or shrimp cultures in the lagoon. The record of this serpulid species, apparently widely distributed in this coastal lagoon, has implications regarding possible effects on the brackish-water ecosystem, since the invasion event very well may have occurred several years ago. It is recommended that an exhaustive study be carried out in the coastal lagoons of Chiapas to evaluate the real distribution and the effects of this invasive species on the ecosystem. A complete description, including photographs and drawings, is provided.
ResumenSe registra por primera vez la especie exótica del Indo-Pacífico occidental, Ficopomatus uschakovi (Polychaeta, Serpulidae), en el Pacífico oriental tropical, en dos sitios de la reserva de la biosfera La Encrucijada, Chiapas, una laguna costera en el Pacífico sur de México. El medio de dispersión de este serpúlido no es claro, debido a que el puerto más cercano (Puerto Chiapas) está 70 km al sur de la laguna, tampoco hay instalaciones portuarias ni estanques de cultivo de camarón. El registro de esta especie de serpúlido, aparentemente bien distribuida en esta laguna costera, tiene implicaciones con respecto a posibles efectos sobre el ecosistema salobre, sobretodo considerando que el evento de invasión pudo suceder hace ya varios años. Se recomienda realizar un estudio exhaustivo en las lagunas costeras de Chiapas para evaluar la distribución real y los efectos de esta especie invasora en el ecosistema. Se incluye una descripción completa, con fotografías e ilustraciones de la especie.
Annelida, brackish-water, Chiapas, fouling, ecologic impacts, La Encrucijada, México, polychaete taxo-nomy, systematics
The genus Ficopomatus Southern, 1921 includes five species of brackish-water serpulids (
On the Pacific Coast of America, no indigenous species of Ficopomatus has been described (
The case of Ficopomatus enigmaticus, a serpulid widely dispersed around the world and with a long historical record of invasions, is a well-known example, because it can have dramatic impacts in invaded habitats building large, reef-like colonies in some coastal lagoons that cause major changes in benthic communities, especially in the Lake of Tunis (
The effects of Ficopomatus miamiensis in Urías lagoon were considered to be positive in the shrimp-ponds because the population of these serpulids (densities higher than 230, 000 ind/m2) helps to clean the water and control the suspended particulates. However, the impact of the colonies attached to the mangrove roots is negative, because the serpulid competes with other fouling and filter-feeding invertebrates such as native barnacles, mussels and oysters (
Ficopomatus uschakovi has been recorded from the Gulf of Guinea, West Africa since the 1950’s (
From 19–24 September, 2011, we visited several sites to collect marine invertebrates in Chiapas, including Puerto Chiapas (or Puerto Madero, as its old name), La Encrucijada Biosphere Reserve, Chocohuital, Boca del Cielo Lagoon, Paredón (Mar Muerto Lagoon) and ending in Oaxaca, in San Dionisio del Mar (Superior Lagoon). Serpulid samples were taken from hard substrates such as rocks, mangrove roots and artificial structures (e.g. piers, submerged buildings, etc.). The preliminary examination of the specimens included only the samples from La Encrucijada Biosphere Reserve because Ficopomatus tubes were immediately identified in the field. In the other sites in Chiapas and Oaxaca the typical tubes of Ficopomatus were not found. Most specimen lots were deposited in the Colección de Invertebrados Marinos, Universidad del Mar, Puerto Ángel, Oaxaca (UMAR-Poly), other samples were deposited in the collections of El Colegio de la Frontera Sur, Chetumal, Quintana Roo (ECOSUR), and in the Universidad Autónoma de Nuevo León, Monterrey, México (UANL). Topotypical specimens examined came from the Los Angeles County Museum of Natural History, Allan Hancock Foundation, Los Angeles, California, USA (LACM-AHF).
The specimens were fixed with 10% formalin and preserved in 70% alcohol. The line drawings were made using camera lucida, the photographs were taken with a digital camera (Nikon Coolpix). Opercula were generally found to be covered in silt and algae, and were, therefore, cleaned with a fine brush.
Standard measurements and counts were total length, measured from the most distal part of the operculum to the pygidium; thoracic length, from the apron to the base of the collar; thoracic width, measured across the collar region; the number of thoracic chaetigers, the number of radioles in each lobe of the branchial crown; opercular length, from the base of peduncle to the end plate; and opercular diameter, measured across the dorso-ventral axis of the end plate.
The abbreviations used in the text were as follows: OL (opercular length), OD (opercular diameter), THL (thoracic length), THW (thoracic width), TL (total length of the body), n (sample size), r (range of data), µ (mean), and ± (standard deviation).
Systematics Class Polychaeta Grube, 1850 Family Serpulidae Rafinesque, 1815 Genus Ficopomatus Southern, 1921Type species. Ficopomatus macrodon Southern, 1921 by monotypy.
http://species-id.net/wiki/Ficopomatus_uschakovi
Figs 1A–E, 2A–IPanadura River estuary, Madu Ganga estuary at Balapitiya and Ratgama Lake at Dodanduwa, Sri Lanka.
Chiapas, South Pacific of Mexico. Five specimens (ECOSUR), three specimens (UANL), more than 100 specimens (UMAR-Poly 112), La Encrucijada Biosphere Reserve, Barra San Juan, 15°09'58"N, 92°51'12"W, 0.5–1 m, submerged mangrove root (Rhizophora mangle), September 21, 2011, Rolando Bastida-Zavala et al. leg. Five specimens (ECOSUR), 20 specimens (UMAR-Poly 113), La Encrucijada Biosphere Reserve, Zacapulco, 15°11'37"N, 92°53'22"W, on rotting mangrove root and gastropod shell, 0–0.5 m, September 21, 2011, Rolando Bastida-Zavala et al. leg.
Two specimens (LACM-AHF N10947), Panadura River estuary, Sri Lanka, brackish water (donated by T.G. Pillai), October 9, 1961.
Mass occurrence is present (Fig. 2A, D); however, some specimens were solitary (Fig. 2E). The tube colour varies from pink to red in live material, changing to white, brown or orange in preserved material (Fig. 2D–E). They possess several prominent to shallow peristomes (Fig. 2D) or only low growth rings (annulations, Fig. 2E), but lack longitudinal ridges and alveoli.
The body is yellow; with 6–7 dark bands on the radioles (preserved material, Figs 1A, 2F). The thorax has five dark bands on the lateral side, between the notopodial bundles, although sometimes these bands are blurred. TL=6.7 mm (n=10, r:4–6.7, µ=5.2 ±1); THL=2 mm (n=10, r:1.4–2, µ=1.7 ±0.2); THW=0.7 mm (n=10, r:0.5–0.7, µ=0.6 ±0.1). The branchial crown has nine radioles (n=10, r:6–9, µ=7.4 ±0.8) on the left, and eight on the right (n=10, r:7–9, µ=8 ±0.7). Interradiolar membrane absent.
The peduncle is smooth, inserted in the left branchial lobe; lacks a constriction (Fig. 1D–E); OL=1.8 mm (n=9, r:1–1.9, µ=1.6 ±0.4). The operculum is spherical to oval in shape, with a slightly convex, flat or slightly concave horny plate (Fig. 1B–E). OD=0.6 mm (n=9, r:0.5–0.8, µ=0.6 ±0.1). The end plate bears 1–5 concentric rows of spines (Fig. 2H); the rows are sometimes incomplete or converge with the other rows. The spines are transparent (Fig. 2I).
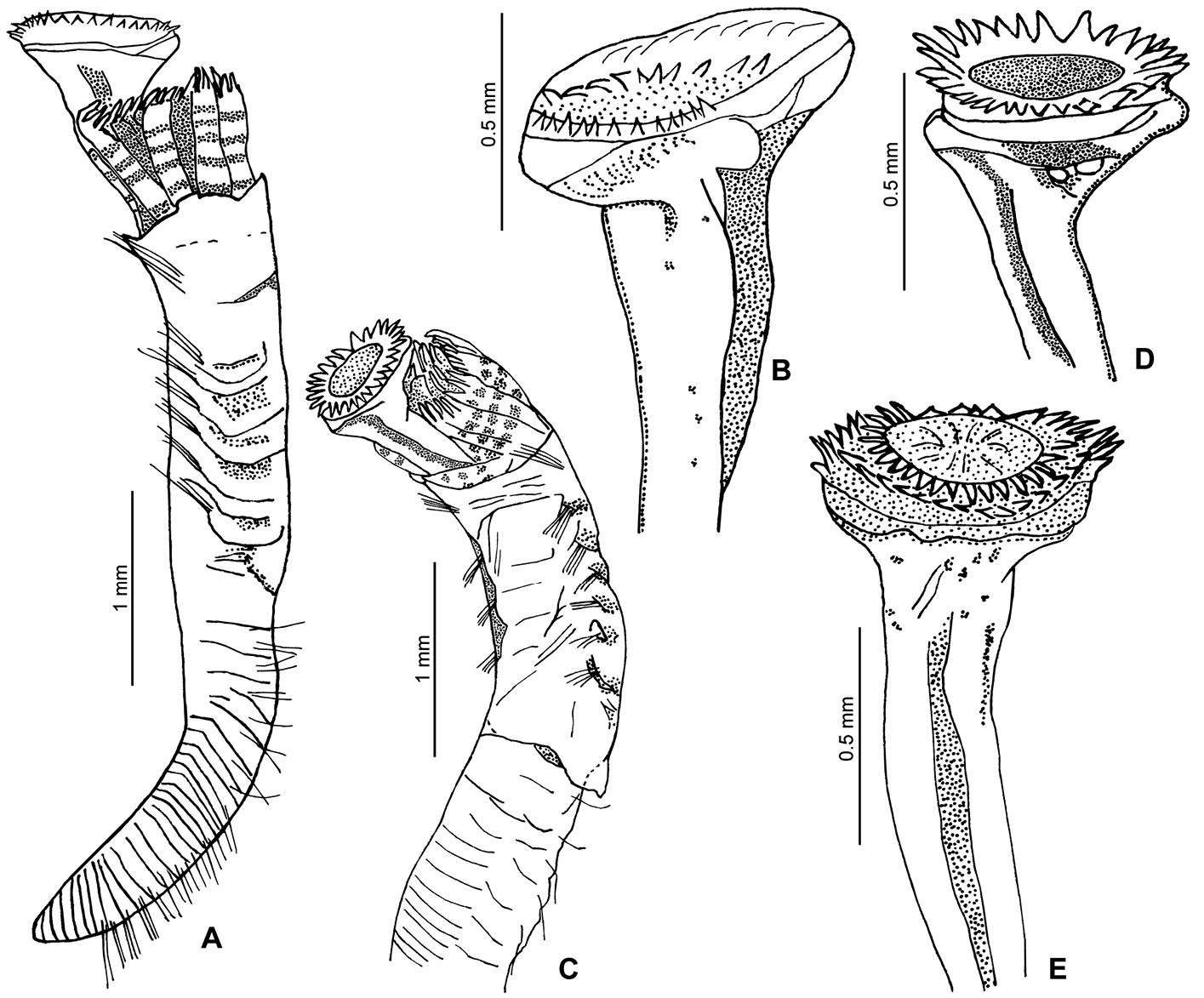
A–E Ficopomatus uschakovi, from La Encrucijada Biosphere Reserve, UMAR-Poly 112–113. A, C complete body in lateral and dorsal views; B, D–E opercula, lateral views.

A–I Ficopomatus uschakovi, from La Encrucijada Biosphere Reserve, UMAR-Poly 112–113. A tubes on mangrove roots B tubes on the shell of the gastropod Thaisella kiosquiformis C mangroves in the collecting site D tubes forming small aggregations E large, single tube F complete specimen in dorsal view G complete specimen with mass of sperm attached to the abdomen H operculum in dorsal view I operculum in aboral view.
The collar is entire, with well-developed lobes. The collar chaetae include coarsely serrated chaetae and hooded (capillary) chaetae. The thoracic membranes are fused dorsally, ventrally forming a small apron. The thorax has six chaetigers with hooded (limbate) chaetae, and saw-shaped uncini. A short achaetous region is present between the thorax and abdomen. Most of the abdominal segments possess geniculate chaetae and rasp-shaped uncini.
Originally limited to Indo-West Pacific and Gulf of Guinea (
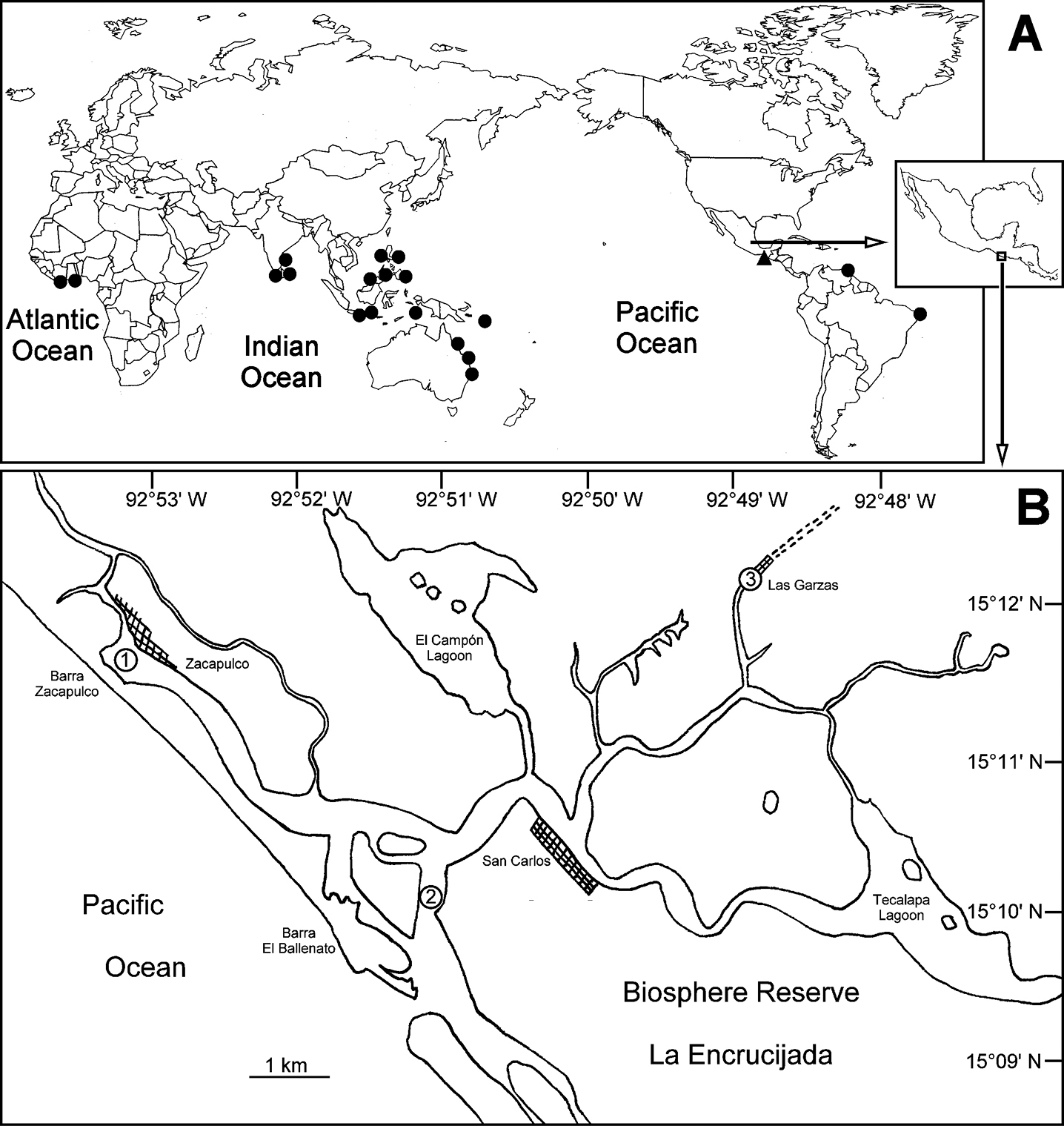
A World-wide distribution of Ficopomatus uschakovi. Triangles denote examined material, circles literature records (data from
Depth: Intertidal to 1 m. On mangrove roots (Fig. 2A, C) and on the shell of the gastropod Thaisella kiosquiformis (Duclos, 1832) (Fig. 2B); coastal lagoon with salinity ranges approximately 20–35 o/oo. On shells, stones and petiole bases of coconut leaves in the topotypical area (
Two specimens (TL=4 mm and 4.2 mm) have masses of sperm glued to the abdomen (Fig. 2G); however, eggs have not been found in the specimens studied in detail (n=10).
The specimens of Ficopomatus uschakovi from Chiapas are slightly bigger than the topotypical specimens (TL=4–4.2 mm, THL=1.1–1.4 mm, THW=0.7–0.8 mm, OL=1.1–1.2, OD=0.3–0.5 mm); however, the rest of the morphological characteristics are very similar.
Apart from the spines of the operculum, the main character that separates Ficopomatus uschakovi from Ficopomatus enigmaticus and Ficopomatus miamiensis is the dorsally fused thoracic membranes. This autapomorphy of Ficopomatus uschakovi is regarded to be of generic level by
In the local community the serpulid tubes on the mangroves are called ‘broca’ (more or less ‘drill’), just like all other sessile invertebrates with calcareous covers, such as barnacles and oysters.
The presence of a species, resembling Ficopomatus uschakovi, from La Encrucijada Biosphere Reserve was initially brought to the attention of the authors two years ago (S.I. Salazar-Vallejo et al. pers. comm. 2009). Previously, no other record of the species existed throughout the Tropical Eastern Pacific. However, the observations in the field show that Ficopomatus uschakovi is widely distributed in La Encrucijada, suggesting that the invasion event of this species may have occurred several years ago.
Although ballast water, sediment transport and fouling are the main means of dispersal of aquatic exotic species (
Mass occurrence of Ficopomatus uschakovi was observed (Fig. 2A), however, not to the extent of the reef-like structures, formed by Ficopomatus enigmaticus in Argentina (
We would like to thank Sergio Salazar-Vallejo, Luis Carrera-Parra (ECOSUR, Chetumal) and Ángel de León-González (UANL, Monterrey) for alerting us to the existence of this species in La Encrucijada. We thank Edmundo Aguilar López, Teófilo González and Candelario Girón (CONANP, Chiapas), for their support during the visit to La Encrucijada Biosphere Reserve. The help in the field of Jani Jarquín-González (INECOL, Xalapa), Carmen Méndez (CICIMAR, La Paz) and Violeta Jiménez (UNSIJ, Ixtlán) was invaluable; Esmeralda Morales (UMAR, Puerto Angel), is thanked for the fast processing of the samples. The gastropod shell was identified by Norma Barrientos (UMAR). The specimens from Sri Lanka were examined in a visit to Los Angeles in 2004, hosted kindly by Leslie Harris (LACM-AHF). We appreciated the corrections and comments of Harry ten Hove (NCB Naturalis, Leiden), one anonymous referee and Chris Glasby (as editor) to the latest version. This work is part of the study of marine exotic species made by the Mexican network of exotic species (UMAR-UANL-UABC), supported by the project Programa de Mejoramiento del Profesorado 103–5/09/1353.