





(C) Maurizio Mei. This is an open access article distributed under the terms of the Creative Commons Attribution License 3.0 (CC-BY), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
For reference, use of the paginated PDF or printed version of this article is recommended.
The European bone-skippers (Diptera: Piophilidae: Thyreophorina), long considered extinct, have recently been the object of much interest by dipterists after their unexpected rediscovery. Considerable faunistic work has been done on these flies in recent years. However, some nomenclatural and taxonomic issues still require attention. A neotype is designated for Thyreophora anthropophaga Robineau-Desvoidy, 1830 (now in the genus Centrophlebomyia Hendel, 1903) to fix the identity of this nominal species. Centrophlebomyia anthropophaga is recognized as a valid species. It is described and illustrated in detail, and information on its preimaginal instars is provided for the first time. Four Palaearctic species of Centrophlebomyia are recognized and reviewed and a key is provided for their identification. Centrophlebomyia orientalis Hendel, 1907 from northern India, is removed from synonymy with Centrophlebomyia anthropophaga and recognized as a valid species of Centrophlebomyia, stat. r. The nominal genus Protothyreophora Ozerov, 1984 is considered a junior synonym of Centrophlebomyia, syn. n.
Neotype designation, new synonymy, Protothyreophora, preimaginal instars, life history, systematics
It is undeniable that some organisms are able to provoke great curiosity, which may last through generations of students. There are several, often interrelated reasons for this: rarity for example, true or apparent, but also a certain aesthetic appeal or unusual biology. All of these factors have contributed to the long-term popularity of thyreophorine Piophilidae, particularly that of the European species. These unusual-looking flies, commonly known as bone-skippers, appear to be associated as necrophages with large vertebrate carrion, including human corpses, and have always been considered rare, almost legendary (cf.
The present work was triggered by the finding of several specimens of a species of Centrophlebomyia Hendel, 1903 in central Italy (Monte Velino, Central Apennines), during a study on the necrophilous insect fauna associated with carrion (
Male terminalia, pinned specimens and larvae were examined, uncoated, with a Hitachi TM1000 environmental scanning electron microscope (ESEM). Male terminalia were also slide mounted. Line drawings were made using a drawing tube. Figure 38 was prepared from composites of images captured using a DS-L1 digital camera (Nikon, Tokyo) mounted on a MZ 12.5 stereoscopic microscope (Leica, Wetzlar, Germany) and processed with Helicon Focus Pro software (Kharkov, Ukraine).
Male terminalia were dissected following the method described by
The material examined is deposited in the following collections (acronyms as used in the text):
MZUR Museum of Zoology, Sapienza Università di Roma, Italy;
NHMW Naturhistorische Museum, Wien, Austria;
TAU Department of Zoology, Tel Aviv University, Tel Aviv, Israel;
ZMUC Natural History Museum of Denmark, Zoological Museum, University of Copenhagen, Denmark.
Label data of type specimens are given verbatim using the following symbols:
/ end of a line and beginning of the next;
// end of a label and beginning of the next (from top to bottom on the same pin).
http://species-id.net/wiki/Centrophlebomyia
Figs 1−38Brownish, scathophagid-like flies, body length 4−8 mm. Body densely to moderately microtomentose and covered with long, fine setulae, especially in males. Frons with one or two upper reclinate orbital setae. Ocellar seta, medial and lateral vertical setae, and postvertical seta long and robust. Two to ten frontal setae usually arranged more or less regularly around lunula. Face with a strong, flattened median carina, antennal grooves deep. Parafacial with a patch of microtomentum at mid length (Fig. 15). Compound eye almost round in lateral view. Two pairs of strong vibrissae present, subequal in length and strength. Thoracic chaetotaxy as follows: 0−2 postpronotal setae (postpronotal setae usually absent in male of Centrophlebomyia furcata); 1 + 1 intra-alar, 1−2 + 3 strong dorsocentral setae, 1 postalar, 2 notopleural setae, 0−1 prescutellar acrostichal setae. Scutellum long, dorsally flattened, much more developed in male than in female, with two pairs of setae, apical pair very long and strong, almost spiniform in larger males. Dorsal surface of scutellum bare. Development of scutellum in male related to body size. Propleural seta strong. Katepisternum densely setulose, with one strong katepisternal seta at upper posterior margin. Anepisternum with a row of setae along posterior margin, one of them strong. Thorax (except scutellum) very finely setulose throughout, besides the strong setae.
Wing membrane hyaline. Costa more or less spinose (i.e., with a regular row of stronger setulae interspersed with the general costal setulae), spine-like setae stronger in male than in female. Anal vein fading out well before wing margin. Legs thickly setulose in both sexes (almost woolly in male). Fore femur with 5–6 weak posteroventral setae near apex, scarcely differentiated in male. Hind femur with 2–3 anteroventral setae near apex. Mid tibia with five apical setae on ventral side: middle and lateral ones strongest. Hind tibia with one short, curved apical seta on posteroventral surface. Tarsi unmodified.
Described by
Europe, North Africa, Middle East, Russian Far East, northern India (Kashmir and Darjeeling).
The generic diagnosis incorporates the characters given by
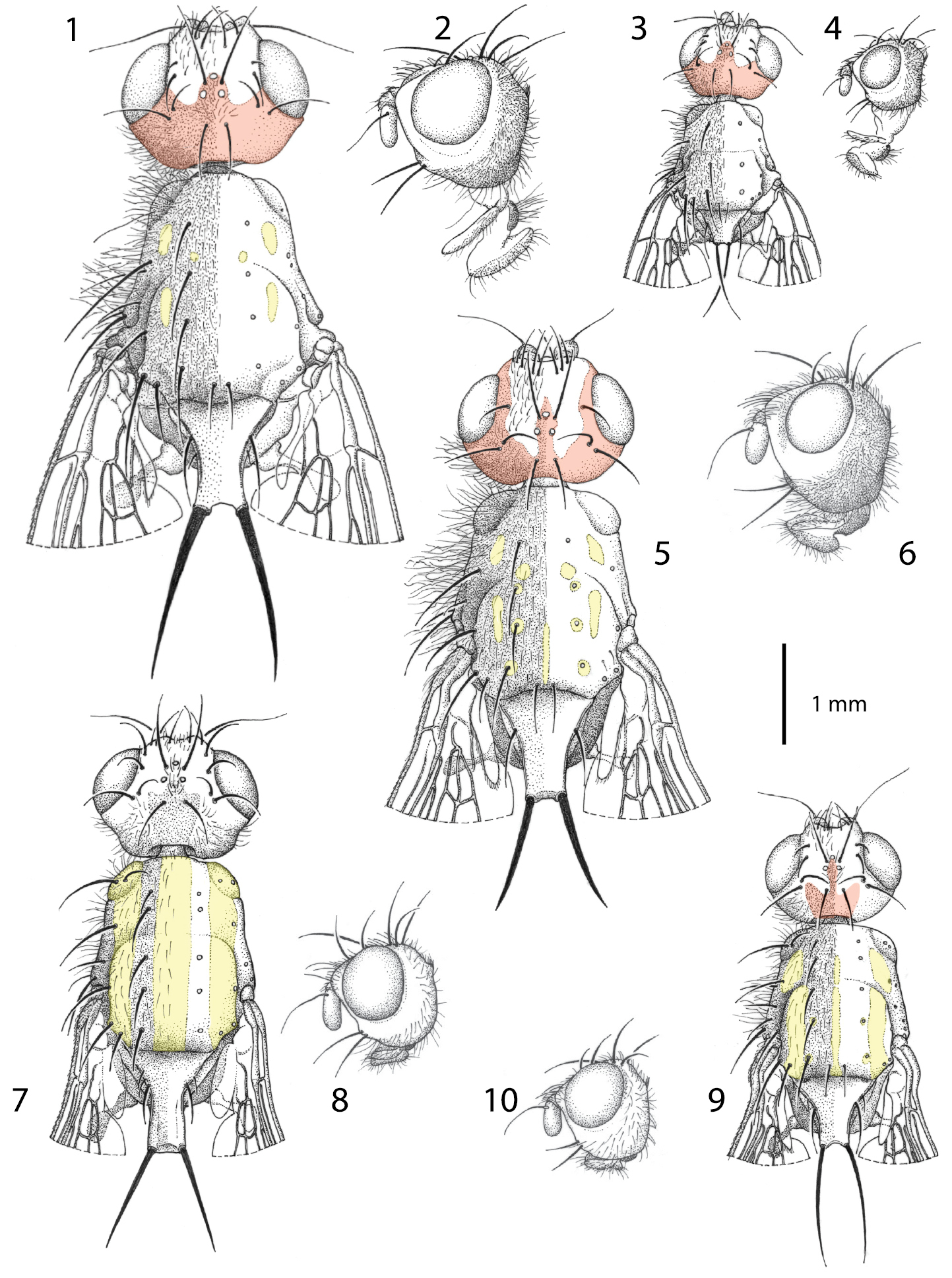
Males of Centrophlebomyia spp. 1, 3, 5, 7, 9 head and thorax in dorsal view 2, 4, 6, 8, 10 head in lateral view 1−4 Centrophlebomyia anthropophaga (Italy) 5−6 Centrophlebomyia furcata (Italy) 7−8 Centrophlebomyia grunini (Russian Far East) 9−10 Centrophlebomyia orientalis (India). In red the microtomentum pattern of head; in yellow the shiny, non microtomentose, pattern of thorax.
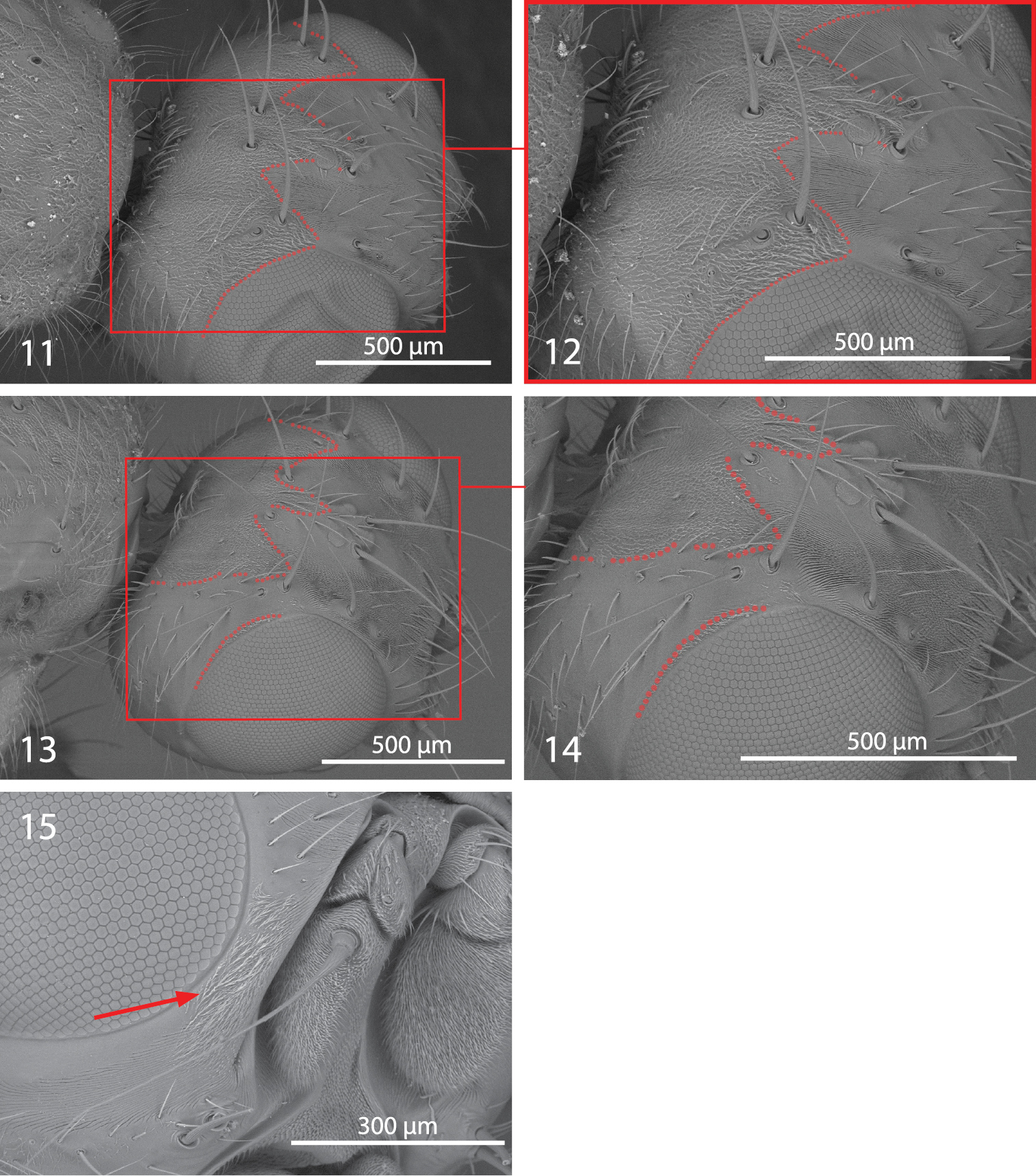
Centrophlebomyia spp. 11−14 head in dorsolateral view [dotted line indicates the border of microtomentum] 11−12 Centrophlebomyia anthropophaga (Italy) 13−14 Centrophlebomyia orientalis (India) 15 Centrophlebomyia anthropophaga (Italy), detail of head in anterodorsal view [arrow indicates microtomentum on parafacial].

Centrophlebomyia spp. 16−17 Centrophlebomyia anthropophaga (Italy), anterior part of thorax in dorsolateral view [red circles indicate postpronotal setae] 16 male 17 female 18−21 Wing 18 Centrophlebomyia anthropophaga (Italy), detail of third costal sector (Cs3) [in red the costal spine-like setae] 19−20 Centrophlebomyia grunini (Russian Far East) 21 Centrophlebomyia orientalis (India), detail of third costal sector (Cs3).
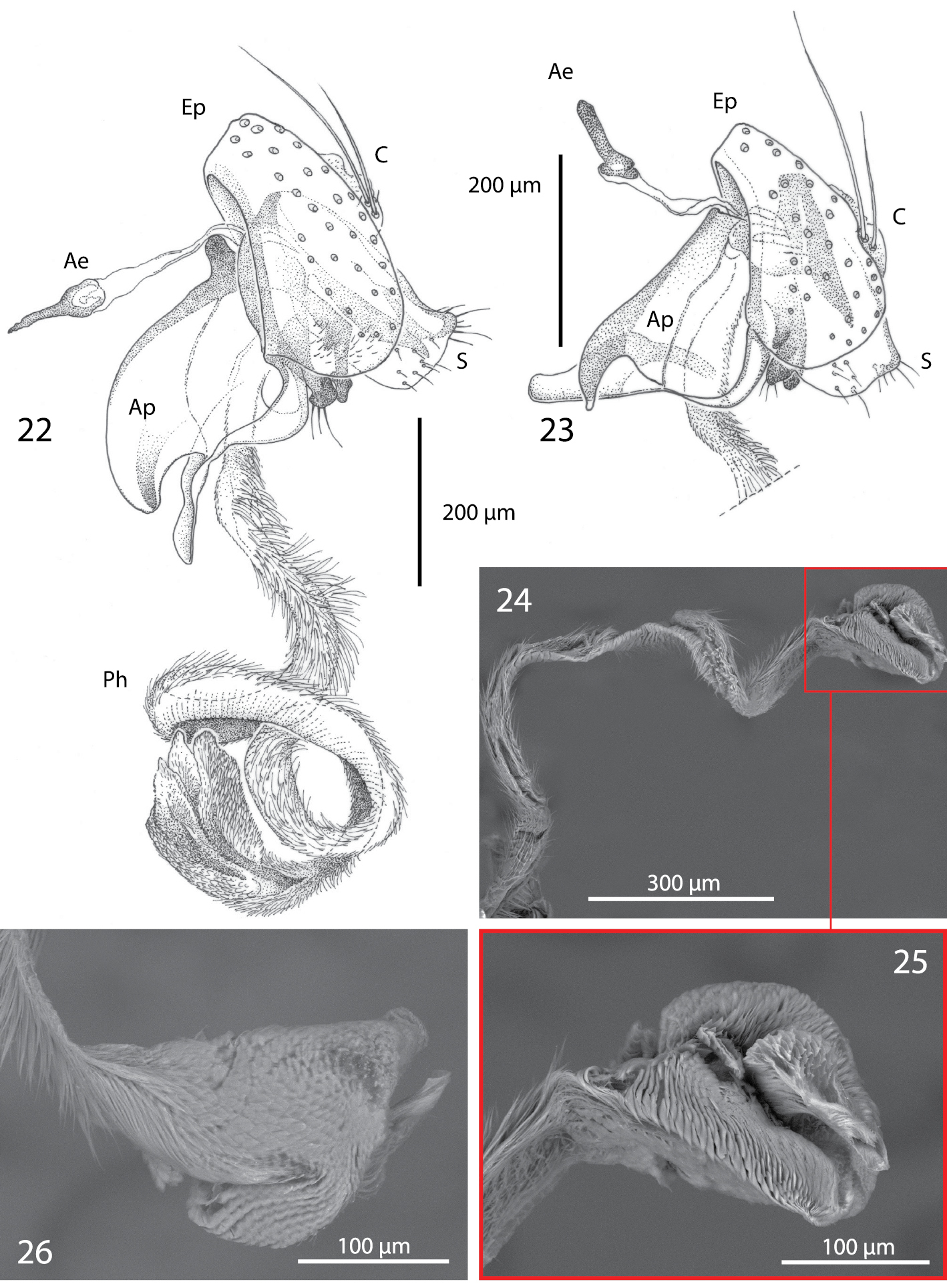
Male terminalia of Centrophlebomyia spp. [Ae = ejaculatory apodeme; Ap = phallapodeme; C = cerci; Ep = epandrium; Ph = phallus; S = surstylus] 22 Centrophlebomyia anthropophaga (Italy) 23 Centrophlebomyia orientalis (India) 24−26 Centrophlebomyia anthropophaga (Italy), phallus.
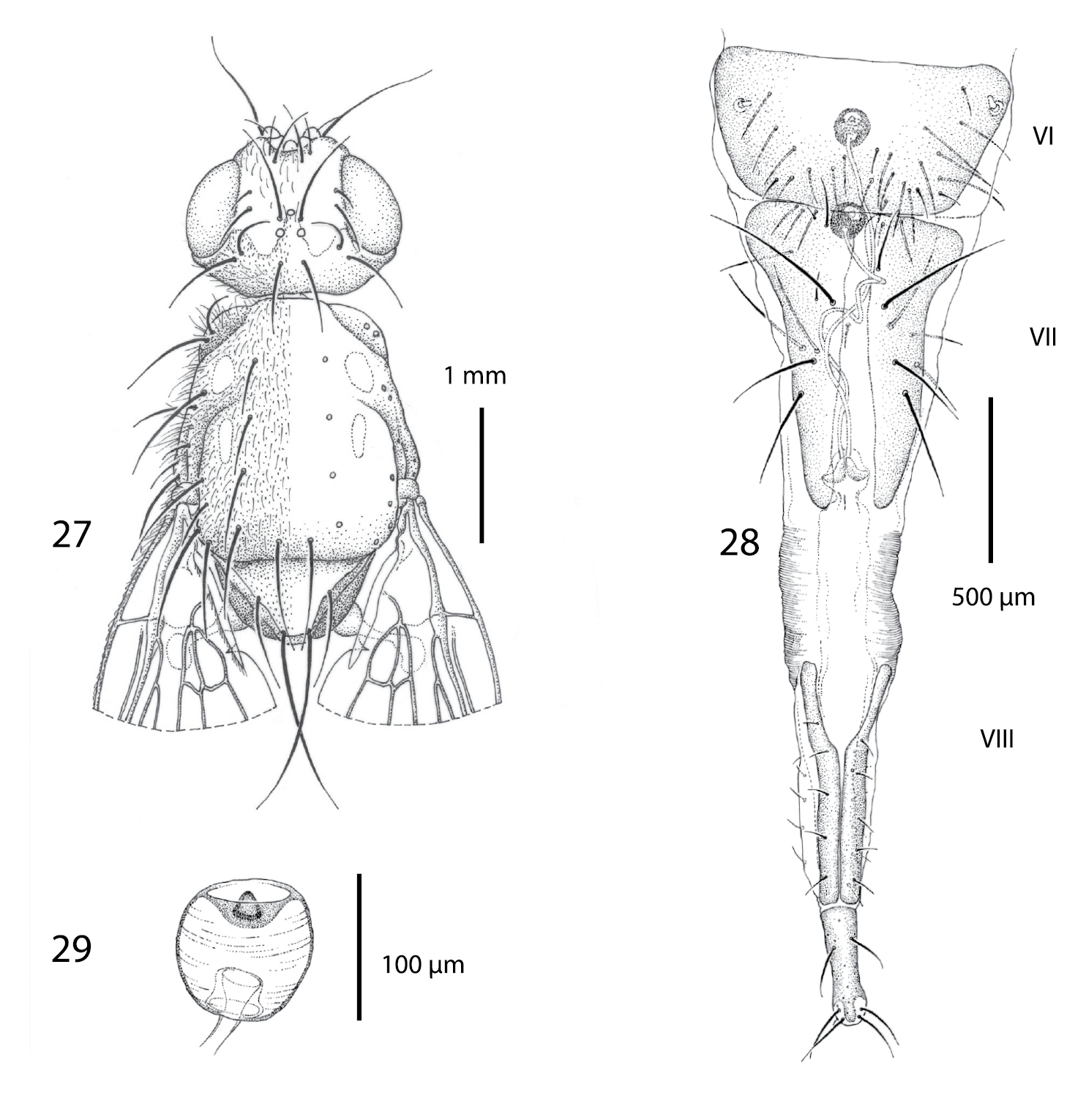
Female of Centrophlebomyia anthropophaga (Italy) 27 head and thorax in dorsal view 28 ovipositor in dorsal view 29 spermatheca.
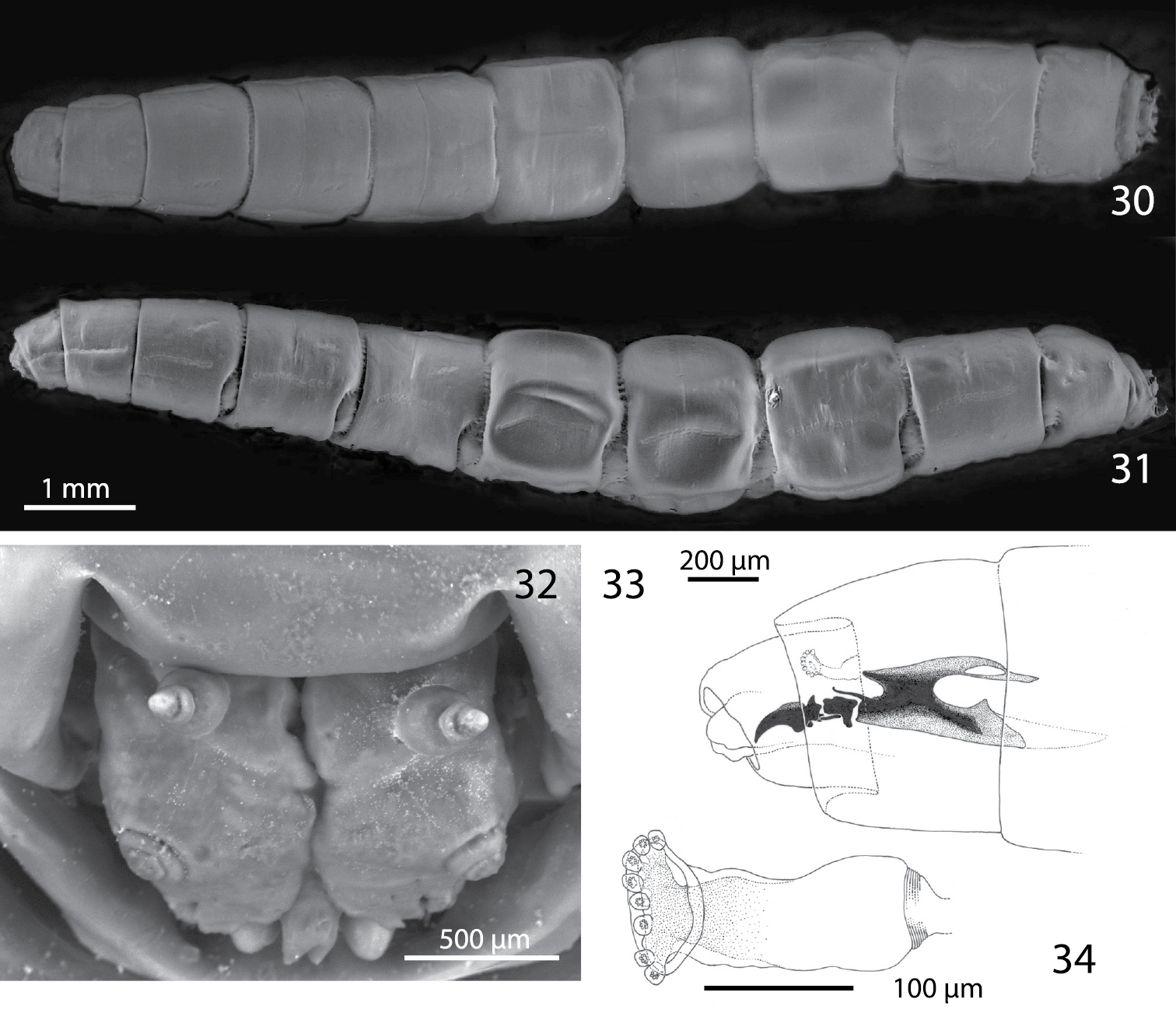
Third instar of Centrophlebomyia anthropophaga (Italy) 30 habitus in dorsal view 31 habitus in lateral view 32 head in frontal view 33 cephalopharyngeal skeleton in lateral view 34 aQnterior spiracle in lateral view.

Third instar and puparium of Centrophlebomyia anthropophaga (Italy) 35 last segment bearing posterior spiracle in posterior view 36 last segment bearing posterior spiracle in lateral view 37 posterior spiracles in posterior view 38 puparium in dorsal view.
http://species-id.net/wiki/Centrophlebomyia_anthropophaga
Figs 1−4, 11, 12, 15−18, 22, 24−38Neotype (designated below). Male (ZMUC), here designated, from Sardinia, Italy and bearing the following labels: SARDEGNA / Belvì (NU) / 10.X.’84 [39°57.889'N, 9°11.111'E] // Neotype ♂ / Thyreophora / anthropophaga / Robineau-Desvoidy, 1830 / M. Mei & P. Cerretti des. 2013 // Centrophlebomyia / anthropophaga / (Robineau-Desvoidy, 1830) / M. Mei & P. Cerretti det. 2013).
Other material examined. 5 ♂♂, 5 ♀♀, same data as neotype. 35 ♂♂, 3 ♀♀, Italy, Abruzzo, L’Aquila province, Riserva Naturale Orientata Monte Velino, Mandridi, 42°7.696'N, 13°22.247'E, 1270 m, 11.XI.2005, G. Lo Giudice, M. Mini, A. Vigna Taglianti legit. 20 ♂♂, 13 ♀♀, same data but 9.XII.2005; 13 ♂♂, 4 ♀♀, same data but 25.I.2006; 7 ♂♂, 1 ♀♀, same data but 25.X.2006; 2 ♂♂, same data but 9.XI.2006; 3 ♂♂, 5 ♀♀, same data but 16.XI.2006; 6 ♂♂, 4 ♀♀, same data but 30.XI.2006; 2♂♂, same data but XI.2006 reared from larvae, (see below); 13 ♂♂, 2 ♀♀, same data but 14.X.2007; 2 ♂♂, same data but 18.XI.2008.
Specimens from Sardinia were collected from a bag of dead, decaying snails (
?France (Paris), Italy (Sardinia, Central Apennines).
Colouration. Head, including antenna and palpus, usually reddish yellow; sometimes dorsal half of occiput black. Occiput, ocellar triangle, genal dilation, and parafacial covered with microtomentum. When seen in dorsal view, the microtomentum that covers the occiput anteriorly outlines a three-pointed crown on the frons between the medial posterior margin of eyes (Figs 1, 3, 11, 12); middle tip of crown corresponds to anterior ocellus; lateral tips, laterally confined by eyes, end about level with posterior ocelli or slightly anteriorly. Prementum black. Postpronotum at least partly reddish laterally (reddish colour usually not visible in dorsal view). Scutum black in ground colour, covered with thin microtomentum except around base of dorsocentral setae and two lateral, longitudinal shiny vittae, widely interrupted at level of transverse suture (Figs 1, 27) (suture well developed laterally up to level of dorsocentral row). Scutellum at least apically yellow. Legs usually entirely yellow, rarely tarsi darkened. Abdomen usually entirely yellow or light brown, but can vary from dark brown to shiny black dorsally in some females. Setae of whole body black. Wing hyaline.
Head (Figs 1−4, 11−12, 15, 27). Head about as wide as thorax. Eye almost round. Frons 2.0−2.5 times as wide as an eye in dorsal view. Parafacial 1/2–2/3 as wide as first flagellomere, both measured at mid length. Gena, in profile, 0.33–0.65 times as high as eye. Medial vertical seta well developed, reclinate. Lateral vertical seta well developed, about 4/5 of medial vertical seta, lateroclinate. One or (usually) two upper reclinate orbital setae; when two are present, then anterior one at most 1/3 as long as posterior seta, and distinctly thinner. Postocellar seta strong and reclinate, subequal in size to ocellar and medial vertical setae. Ocellar seta proclinate. Anterior margin of frons with 2–3 pairs of pro- and medioclinate, strong frontal setae. Fronto-orbital plate with scattered, short, proclinate or medioclinate setae, between posteriormost upper reclinate orbital seta and distal margin of pedicel. Vibrissa double, very strong. Antenna shorter than height of facial ridge; first flagellomere 1.3–2.0 times as long as pedicel. Occiput and genal dilation covered with scattered black setae. Palpus well developed, apically clavate, covered with fine black setae.
Thorax (Figs 1, 3, 16, 17, 27). Thorax covered with fine black setulae, those on scutum distinctly shorter than those on pleurae. Postpronotum with or without 1–2 very fine setae in male (Fig. 16), usually with 2, relatively strong setae in female (Fig. 17). One strong presutural and 2 postsutural supra-alar setae; posterior postsutural supra-alar seta short and thin. One presutural and 3 postsutural dorsocentral setae (Figs 1, 27) (2 postsutural dorsocentral setae may occur in smaller sized male specimens (Fig. 3)). One, short and weak, prescutellar acrostichal seta. Scutellum dorsally flat to slightly concave (ground plan trait of the Thyreophorina, McAlpine, 1977), more or less elongated posteriorly, with one lateral seta and one apical seta (Figs 1, 3); lateral seta usually much smaller than apical seta. Shape and size of scutellum strongly variable (Figs 1, 3, 27) between sexes and between males of different sizes (Figs 1, 3). Two notopleural setae. One anepisternal seta. One katepisternal seta. Legs robust, covered with long and fine setulae. Mid tibia with 3–5 robust ventral preapical setae. Claws well developed in both sexes, about as long as fifth tarsal segment in male, varying in length between 0.5 and 0.7 times as long as fifth tarsal segment in female. Ventral row of costal setae (specifically CS3) characterized by the presence of some longer and stouter setae placed at more or less regular intervals (Fig. 18).
Abdomen. Male: more or less elongated; tergite 1 laterodorsally covered with short erect hair-like setae, medially bare; tergites 2 and 3 laterodorsally and ventrally covered with long, hair-like setae that become shorter toward the midline of tergites. Tergites 4 and 5 evenly covered with long, erect hair-like setae. Female: abdominal setae distinctly shorter.
Male terminalia (Figs 22, 24−26). Epandrium short and convex. Surstyli massive, almost touching each other posteromedially; distal margin of surstylus slightly bent posteriorly. Cerci very small, bearing long setae. Phallapodeme, in lateral view, very large with an evenly convex dorsal margin (Fig. 22). Pregonite well sclerotized, relatively narrow and slightly bent posteriorly; basally fused to hypandrium; pregonite with 1−2 fine setae distally. Postgonite very long, well sclerotized and evenly bent anteriorly. Pregonite and postgonite pincer-like in relative position, almost touching each other distally. Epiphallus attached basally and well developed. Basiphallus very long, tubular, covered with fine pubescence and membranous. Distiphallus massive, slightly sclerotized, covered with fine pubescence as in basiphallus; distiphallus with two large laterodistal lobes.
Female terminalia (Figs 28−29). Ovipositor long and telescopically retracted within fifth segment. Tergites 6 and 7 relatively wide and more or less flattened. Tergite 8 longitudinally divided into two halves. Cerci not differentiated. Two rounded and well sclerotized spermathecae.
Both the larva and puparium of Centrophlebomyia anthropophaga (Figs 30−38) correspond well to features given by
On April 5th (n=15) and May 3rd (n=7), 2006, several mature larvae were collected from the soil a few centimetres below the sheep quarters used as bait for the pitfall traps set in the “vulture restaurant” (see above under “Remarks”). The larvae were then transferred into two petri dishes (12 cm diameter): one filled with potting soil, the other with natural soil collected with the larvae from under the carcass. Moisture was provided each week until midsummer. All larvae remained active, though only slightly so, during this time. By June 1st, five out of 22 larvae had died. The loss of larvae continued steadily and by the beginning of September only six larvae were left, three in the potting soil and three in the natural soil. In early October 2006, two puparia were found in each dish and all the remaining larvae were dead. The four puparia and small amounts of soil were isolated in smaller dishes. An adult male emerged in November from one of the puparia in the natural soil, and another adult (possibly a male) was found dead in its puparium in the potting soil. The remaining two puparia failed to produce adults.
Our observations are consistent with those of
http://species-id.net/wiki/Centrophlebomyia_furcata
Figs 5−61 ♀, 1 ♂, Israel, Tel Aviv, 17.XII.1977, A. Freiberg legit (MZUR); several males and females, same data (TAU). 16 ♂♂, 9 ♀♀, Italy, Latium, Monti della Tolfa, Mount S. Ansino, 332 m, 42°03'51.85"N, 11°59'47.05"E, 28.XII.2011, M. Mei legit, on dead sheep (MZUR).
Europe: Austria, Cyprus, France, Germany, Greece, Italy, Spain, United Kingdom; North Africa: Algeria; Middle East: Turkey, Israel.
http://species-id.net/wiki/Centrophlebomyia_grunini
Figs 7−8, 19−201 ♂, 1 ♀, each bearing the following labels: [Russia] Aмурская обл[аст] / Г. ЗеЯ [= Amur region, Zeya.] 1.VIII.1981 / A. Ozerov / PARATYPE (ZMUC).
Russian Far East.
Holotype male, bearing the following labels: [INDIA] Darjeeling / Juni / Frusthofer leg. // Centrophlebomyia / orientalis / Hendel [handwritten] // Type [red label] // Holotypus [red label] // Pseudotyreo - / phora orientalis / Hendel / A. Ozerov det., 1984 (NHMW).
INDIA (Kashmir) Gulmarg, 2600–3000 m, 17.VIII 5.IX.1978: 2 ♂♂, 1 ♀ (ZMUC).
Northern India (Darjeeling, Kashmir).
| 1 | Scutum mostly shiny black with two large, longitudinal, silver stripes of microtomentum aligned with, and along the entire length of, dorsocentral setae (Fig. 7 [where the three shiny black vittae are yellow highlighted]). Head almost entirely smooth, katepisternum without microtomentum. Costal vein without spine-like setae (Figs 19−20). [Gena 0.44–0.47 times as high as eye in lateral view (Fig. 8)] | Centrophlebomyia grunini (Ozerov) comb. n. |
| – | Scutum usually more extensively covered with microtomentum, and not showing the pattern described above (Figs 1, 3, 5, 9, 27). Head microtomentose at least on medial dorsal portion of postcranium, katepisternum at least partly microtomentose (e.g., dorsal and posterior to base of katepisternal seta). Ventral row of costal setae characterized by some longer and stouter (spine-like) setae placed at more or less regular intervals, especially on Cs3 (Figs 18, 21) | 2 |
| 2 | Gena 0.68–0.95 times as high as eye in lateral view (Fig. 6). One upper reclinate orbital seta (Figs 5, 6). Genal dilation and occiput entirely covered with microtometum. Occipital microtomentum extends anteriorly on fronto-orbital plate as shown in Fig. 5 [in red]. One presutural dorsocentral seta (Fig. 5). Katepisternum entirely covered with microtomentum. Male: postpronotal setae not differentiated (Fig. 5) | Centrophlebomyia furcata (Fabricius) |
| – | Gena 0.33–0.65 times as high as eye in lateral view (Figs 2, 4, 10). Two upper reclinate orbital setae (exceptionally only one in Centrophlebomyia anthropophaga) (Figs 2, 4, 10). Occipital microtomentum never extending anteriorly on fronto-orbital plate (Figs 1, 3, 9 [in red]). Occipital microtomentum extends anteriorly on frons as shown in Figs 1, 3, 9, 11−14 [in red]. One or two presutural dorsocentral setae. Katepisternum varying from entirely covered with microtomentum to largely smooth. Male: postpronotal setae well developed | 3 |
| 3 | Genal dilation and occiput entirely covered with microtometum (Figs 1, 3, 11, 12 [in red]). Occipital microtomentum extends anteriorly on frons as shown in Figs 1, 3, 11, 12 [in red]. One presutural dorsocentral seta (Figs 1, 3, 27). Katepisternum entirely covered with thick microtomentum. Usually two upper reclinate orbital setae, the anterior one very short and weak (Figs 2, 4) (only one upper reclinate orbital seta present in a few specimens from Sardinia). Male: phallapodeme, in lateral view, with an evenly convex dorsal margin and posterior tip lobe-like (Fig. 22); one or rarely two hair-like postpronotal setae | anthropophaga (Robineau-Desvoidy) |
| – | Genal dilation entirely smooth, occiput largely without microtomentum laterally (Figs 9, 13, 14 [where microtomentum is red highlighted]). Occipital microtomentum extends anteriorly on frons as shown in Figs 9, 13, 14 [in red]. Two presutural dorsocentral setae (Fig. 9). Katepisternum almost entirely smooth. Two upper reclinate orbital setae, the anterior one distinctly shorter but somewhat stout (Fig. 10). Male: phallapodeme, in lateral view, with an almost straight dorsal margin, posterior tip narrow and hook-like (Fig. 23); two strong postpronotal setae | orientalis (Hendel) |
Centrophlebomyia anthropophaga was described by
Our study shows that specimens of Centrophlebomyia from Sardinia and central Italy are conspecific and are different from both Centrophlebomyia orientalis and Centrophlebomyia furcata, so we here propose to remove orientalis Hendel from synonymy with anthropophaga Robineau-Desvoidy.
Furthermore, considering the confused taxonomic and nomenclatural situation described above, created by
Considering that:
i) Centrophlebomyia anthropophaga’s original description is vague and insufficient, and no type specimens ever existed in collections as no actual specimen was ever collected;
ii) The name anthropophaga is nomenclaturally available according to the
iii)
iv) Robineau-Desvoidy’s original description of Centrophlebomyia anthropophaga is not inconsistent also with characters shown by the Italian specimens;
v) The Centrophlebomyia from Italy is not conspecific with Centrophlebomyia orientalis, the species dealt with by
vi) The specimens from Sardinia were already referred to under the name anthropophaga by
we select as neotype a male specimen from Sardinia chosen among those studied by
The thyreophorine genus Protothyreophora was proposed by
For these reasons, we consider the monotypic genus Protothyreophora as a junior synonym of Centrophlebomyia.
The European thyreophorine species have always been considered very rare insects and have famously also been considered as locally or even globally extinct (
It would seem that these flies may also be relatively abundant in sites where they are present (
It would also be interesting to assess the role of food sources other than large mammal carcasses in maintaining natural populations of these flies. Other studies have shown that thyreophorine species have also been collected from a bag of dead decaying snails (Centrophlebomyia anthropophaga:
We hope that the interest generated by the recent rediscovery of these flies will result in further studies on their reproductive biology and ethology, as combining the results of such studies with the already available morphological data may help explain the great polymorphism observed in adult males.
Most sincere thanks are due to J.E. O’Hara (Canadian National Collection of Insects, Ottawa, Canada) and two anonymous reviewers for their constructive comments on earlier drafts of this manuscript, to A. Minelli (Padova University, Italy), T. Pape and V. Michelsen (ZMUC) for their very useful and appreciated advice during the course of this study. Many thanks also to T. Pape (ZMUC), P. Sehnal (NHMW), A. Freidberg (TAU) and L. Rivosecchi (Rome, Italy), who allowed us to study the material under their care; to Tiziana Altea, Luciano Sammarone, Mario Posillico, Rosario Bartolo and Vincenzo Cofini (Riserva Nazionale Orientata Monte Velino, Abruzzo, Italy) who facilitated access and field work in the Reserve; to M. Mini (Rome, Italy) for help collecting the material; to Angela Cerasoli (Vienna) for assistance in examining the holotype of Centrophlebomyia orientalis. The authors also thank F. Mason (Italian National Forestry Service, Verona, Italy) and A. Vigna Taglianti (MZUR) for their constant support of our research activities in the Central Apennines.
Online media coverage of recent findings of Thyreophorina in Europe:
http://www.youtube.com/watch?v=gwqL02RstAw
http://www.youtube.com/watch?v=QW-tOF0WtXA
http://www.youtube.com/watch?v=zXgIrfwwRvg
http://www.youtube.com/watch?v=Bmqgf-TaJsQ
http://www.youtube.com/watch?v=eCm9I__G7HI
http://www.youtube.com/watch?v=p1QcpliZm3g
http://www.youtube.com/watch?v=gGqGAEUDFUU
http://www.youtube.com/watch?v=NnECVsursMs
http://www.diptera.info/forum/viewthread.php?forum_id=5&thread_id=46137
http://www.diptera.info/forum/viewthread.php?forum_id=5&thread_id=26429
http://www.diptera.info/forum/viewthread.php?forum_id=22&thread_id=12704
http://www.diptera.info/forum/viewthread.php?forum_id=5&thread_id=11348
http://www.diptera.info/forum/viewthread.php?forum_id=5&thread_id=32315
http://www.diptera.info/forum/viewthread.php?forum_id=12&thread_id=32317
http://www.diptera.info/forum/viewthread.php?forum_id=5&thread_id=26429
http://news.bbc.co.uk/earth/hi/earth_news/newsid_9008000/9008585.stm