





(C) 2012 Eduardo Suárez-Morales. This is an open access article distributed under the terms of the Creative Commons Attribution License 3.0 (CC-BY), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
For reference, use of the paginated PDF or printed version of this article is recommended.
During a survey of the epipelagic zooplankton carried out off the coast of the Mexican states of Jalisco and Colima, in the Eastern Tropical Pacific, female and male specimens of the poorly known calanoid copepod Pontellopsis lubbockii (Giesbrecht, 1889) were collected. Because previous descriptions and illustrations are largely incomplete and have caused some taxonomical confusion, this species is fully redescribed from specimens from the Mexican Pacific. The species has some characters that have been overlooked, but those related to the female genital double-somite are the most striking, it has two conical dorsal protuberances and a long ventral spiniform process unique of this species. The mouthparts of this species have not been hitherto described and figured, the flexible terminal setae of legs 3 and 4 is noteworthy. The male general morphology agrees in general with previous data, but new details of the leg 5 and geniculate antennule are added. Its mouthparts, with strong, serrate setae on the maxillae and maxillules, and a strong mandibular edge, suggest that this is a predator form. A dichotomous key for the identification of males and females of the species of Pontellopsis known from the Eastern Tropical Pacific is included.
Zooplankton, Mexican Pacific, taxonomy of copepods, biodiversity, pelagic crustaceans
The genus Pontellopsis Brady, 1883 currently contains up to 33 species (
Pontellopsis lubbockii has been relatively rarely taken and is known as a neritic equatorial species endemic to the Gulf of California and adjacent areas of the Eastern Tropical Pacific (ETP) (
The zooplankton samples analyzed were obtained at twelve sites sampled during 27 months between December 1995 and December 1998 on board the R/V BIP-V and “León Marino”. Samples were collected at night time by oblique, semicircular trawls at different depths (10- 115 m) with a Bongo net (0.5 mm mesh size). The zooplankton samples were fixed and preserved with 4% formaldehyde buffered with sodium borate. Copepods were sorted from the original samples and transferred to 70% ethanol with a drop of glycerine for further analysis. Voucher specimens were deposited in the Zooplankton collection of El Colegio de la Frontera Sur, Chetumal, Mexico (ECO-CHZ).
SystematicsOrder Calanoida Sars, 1903
Family Pontellidae Dana, 1853
Pontellopsis Brady, 1883
Monops Lubbock, 1853
Pseudomonops Claus, 1892
Pontellopsis lubbockii (Giesbrecht, 1889)
Eastern Tropical Pacific (3–6°N, 80–82° W), about 400 km west of the coasts of Colombia and 320 km south of the Panama coast.
Two adult females from the central Pacific of Mexico, 14 December 2010, 19.171°N, 104.912°W, coll. E. Kozak and C. Franco-Gordo, specimens undissected, vial deposited at El Colegio de la Frontera Sur, Chetumal, Mexico (ECO-CHZ-08957). One adult male, same date, site, and collector; specimen dissected, semi-permanent slides sealed with Entellan® (ECO-CHZ-08958). One adult male, 25 October, 2011, same site and collector; specimen dissected in slides sealed with Entellan® (ECO-CHZ-08959). One adult female, 24 October, 2011, 19.171°N, 104.912°W, coll. C. Franco-Gordo; specimen undissected, ethanol-preserved, vial (ECO-CHZ-08960). One adult male, 25 September, 1997, 19.033°N, 104.674°W, coll. C. Franco-Gordo; specimens undissected, ethanol-preserved, vial deposited in ECOSUR (ECO-CHZ-08961). One adult female from Californian coast, 7 October, 1904, 30.67°N, 119.59°W, Albatross cruise, Eastern Pacific Expedition, ethanol-preserved, identified by A. Fleminger (USNM-109384). One adult female from off Ecuador, South Pacific Ocean, 8 November, 1928, 01.531°N, 82.273°W, Carnegie Institution of Washington, ethanol-preserved (USNM-80382), previously examined by P. Pillai.
Body length of females range between: 2.09 and 2.17 mm (average 2.13 mm, n=5), measured from anterior cephalosome to posterior border of anal somite. Cephalosome robust, widest at level of fully separated first pedigerous somite. Pedigerous somites 4 and 5 fused; posterior corners of fifth pedigerous somite strongly developed, forming large spine-like processes (Fig. 1A, B). Processes straight, posteriorly directed, reaching about halfway along urosome. Cephalosome with rounded forehead, dorsal lenses absent. Rostrum bifid, with long, slender rostral filaments, gap between rostral rami wide (Fig. 1H), in lateral view reaching halfway of second antennular segment (Fig. 1G). Urosome with two segments: genital double somite and anal somite. Genital double-somite representing about 55% of urosome length, excluding caudal rami; somite strongly asymmetrical, with pair of dorsal protuberances arising from distal margin of somite (Fig. 1C, D). In dorsal view, right protuberance subtriangular, curved, posteriorly directed, reaching about half way along anal somite. Left process smaller, also posteriorly directed rounded tapering distally into strongly chitinized bulb-like process (Fig. 1C). Proximal margin of somite bearing lateral spine-like process on each margin, slightly asymmetrical, right one being longer. Ventral surface of genital double somite swollen, with sickle-shaped process arising anterior to genital operculum, posteriorly directed (Fig. 1D, E, F). Anal somite subrectangular, about 1.5 times wider than long, with rounded distomedial process between insertion points of caudal rami. Dorsal surface of anal somite swollen in lateral view, ornamented with rows of minute spinules. Caudal rami weakly asymmetrical, left ramus slightly larger than right, both rami bearing 6 setae: 1 inner, 3 terminal, 1 outer setae plus short, slender dorsal seta.
Antennules (Fig. 2A) symmetrical, 16-segmented. Segments armed as follows (Arabic numbers= setae; Roman numerals= spines, aes=aesthetascs): 1 (I-III) (1), 2 (IV-VII) (9+aes), 3 (VIII-X) (6, I+aes), 4 (XI-XIII) (4, II+3aes), 5 (XIV) (1, I+aes), 6 (XV-XVI) (4, I+ 2aes), 7(XVII) (1+aes), 8(XVIII) (1+aes), 9 (XIX) (1+aes), 10 (XX) (1+aes), 11 (XXI) (1+aes), 12 (XXII) (1), 13 (XXIII) (1), 14 (XXIV) (1, I), 15 (XXV) (2+aes), 16 (XXVI-XXVIII) (4+aes). Larger and longer setae on segments 2, 4, 7, 8, and 13. Modified, wide-based heavily setulated seta proximally inserted on segment 6; same segment with distally blunt, strongly chitinized spine reaching about 2/3 of way along succeeding segment 7 (Fig. 2A).
Antenna (Fig. 2B) biramous: coxa with short plumose distal seta. Basis and first endopodal segment separated, basis bearing 2 setae, one short, one long. First endopodal segment elongate, armed with two small subdistal setae. Second endopodal segment with 9 and 7 setae on proximal and distal lobes, respectively; distal lobe armed with basal outer row of spinules; exopod 6-segmented, setation formula 1, 2, 1, 1, 1, 2.
Mandible (Figs 2C–E) with wide, heavily chitinized gnathobase; mandibular palp biramous, basipod robust, subrectangular, armed with inner basipodal seta. Endopod 2-segmented, first segment armed with 3 long and one short setae; second segment with 6 terminal setae. Exopod 5-segmented, setal formula as: 1, 1, 1, 1, 2. Mandibular distal edge bearing 7 teeth: from ventral margin dentition includes one apical (a), one subapical (sa), two compound medial (med), and three basal (bas) (see Fig. 2C); medial teeth with rounded edges. Clusters of long and short spinules on base of medial teeth; dorsal end of gnathobase with tight row of setae.
Maxillule (Fig. 3A) typical of pontellids, praecoxal arthrite with 14 setal elements; coxal endite (cx end) with 3 long, robust spine-like elements on endite and 9 setae on epipodite (epi); basis with 3 and 1 setae on proximal (bend1) and distal (bend2) endites, respectively; 1st and 2nd endopod segments, each with 2 setae, incorporated into basis, distal endopod segment with 5 apical setae; exopod with 8 setae.
Maxilla (Fig. 2F) uniramous, first praecoxal endite bearing 4 setae, second with 3 setae (one of them shorter and thinner than the others); two coxal endites each bearing 3 setae. Basis with 2 setae; endopod 4-segmented, setal formula of endopod as: 2, 2, 1, 1. Basal and endopodal setae strongly serrate.
Maxilliped (Fig. 3B) uniramous, with praecoxa and coxa fused, three syncoxal endites well developed, with setal formula 2, 2, 3; endites setae strong, serrate. Inner lateral margin of third endite with rows of short setae. Basis fringed with medial row of 5-6 spinules and 2 distal setae. Endopod 4-segmented, setal formula of endopod as: 2, 1, 1, 2.
Leg 1 with 3-segmented endopod; legs 2-4 with 2-segmented endopods and 3-segmented exopods (Figs 3C-F). Coxae with plumose inner seta; basis of leg 4 with slender outer seta, medial patch of spinules on medial anterior margin of legs 3 and 4. First endopodal segment of second leg with inner rounded protuberance (arrowed in Fig. 3D). In one specimen examined, terminal exopodal spine of legs 3 and 4 modified, represented by flexible seta (Italized in setal formula) (Fig. 3G, H). Seta and spine formula (Arabic numbers=setae, Roman numerals=spines) of legs 1-4 as:
Coxa Basis Exopod Endopod
Leg 1 0-1 0-0 I-1;I-1;II, I, 4 0-1;0-2;1, 2, 3
Leg 2 0-1 0-0 I-1; I-1;III, I, 5 0-3; 2, 2, 4
Leg 3 0-1 0-0 I-1; I-1; III, 1, 5 0-3; 2, 2, 4
Leg 4 0-1 1-0 I-1; I-1;III, 1, 5 0-3; 2, 2, 3
Leg 5 (Fig. 1I, J) biramous, slightly asymmetrical; coxa and intercoxal sclerite fused. Basis subrectangular, naked. Endopod distally bifurcate, about 0.3 times as long as exopodal ramus. Exopod of both legs 1-segmented, elongate, right leg with 3 outer spiniform processes and a large distal inner process; left leg smooth except for two subdistal outer spine-like setae.
Body (Fig. 4A) robust, slightly smaller than female (1.85–2.07 mm, average: 1.98 mm, n=4). Cephalosome about 3.5 times as long as urosome (caudal rami excluded), dorsal surface of cephalosome pilose, particularly pedigerous somites 1-5. Fifth pedigerous somite with asymmetrical lateral expansions, left process spiniform, reaching posterior margin of first urosomite; right side with long curved, ventromedially directed process with small, distally curved rounded process (Fig. 4B). Urosome (Fig. 4A-C) with 5 somites. Genital double-somite strongly asymmetrical, left side with 2 sensilla on outer distal corner; right side expanded forming rounded process armed with two unequal setae (Fig. 4C). Second urosomite with pair of sensillae on right side; third urosomite as long as succeeding somite, with strong laterally-directed rod-like process on right margin, process armed with anterodistal curved row of teeth-like spinules, a short seta, and terminal rows of spinules (Fig. 4D). Anal somite symmetrical, as long as preceding somite. Caudal rami slightly asymmetrical, approximately twice as long as wide.
Right antennule (Fig. 4E–G) with 12 segments geniculate between segments 10–11, reaching middle of third pedigerous somite. Antennular segments armed as follows (Arabic numbers= setae; Roman numerals= spines, aes=aesthetascs): 1 (I-III) (1), 2 (IV-VII) (8+2aes), 3 (VIII-X) (2), 4 (?) (2), 5 (?) (2+aes), 6 (X-XIV) (5+ 2aes), 7 (XV-XVI) (4+aes), 8 (XVII) (2, I+aes), 9 (XVIII-XIX) (3+aes), 10 (XX) (1), 11(XXI-XXIII) (1, II), 12 (XXIV-XXVIII) (8+aes). Spine on segment 8 long, slightly curved; segments 9 and 10 with coarse double row of acuminate sharp teeth (Fig. 4F). Segment 11 with proximal process forming fan-like row of strong spines plus two usual stout spines adjacent to segmental margin (Fig. 4G). Anterior margin of segments 10 and 11 with usual spiniform processes parallel to segmental margin. Left antennule as in female except for shorter spiniform process on segment 6 which is also relatively shorter than in female (Fig. 5A).
Leg 5 (Figs 5B–E) asymmetrical, typical of pontellids. Left leg 5 short; coxa quadrate, basipod (bp) robust, cylindrical, naked. Exopod 3-segmented, segments 2–3 partly fused; first segment cylindrical, with subtriangular process on outer distal margin. Second exopodal segment (Fig. 5E) with medial surface covered by patch of long hair-like setae, segment with inner rounded expansion and subdistal seta on outer lateral margin; third segment with 2 unequal spines plus inner spiniform process. Right leg 5 basis with 2 unequal setae. Exopod with two segments, forming robust, widely open chela; first segment (exp1) forming thumb of chela ending in short, strong process curving inward with inner surface armed with shallow cuticular ridges and small spinules (Fig. 5C). Second exopodal segment forming distal elongate finger, tapering distally, armed with two subequal proximal setae on outer surface plus one proximal and one distal setae inserted on inner surface of segment (Fig. 5D).
Our specimens from the Mexican Pacific were identified as Pontellopsis lubbockii by the females having acute, symmetrical posterolateral corners of the fifth pedigerous somite plus an asymmetrical genital double-somite as long as the anal somite and with two dorsal protuberances. Males have a long, curved process on the right side of the fifth pedigerous somite, a laterally directed process on the third urosomite combined with a pair of long stout setae on the right margin of the genital double somite. Females of this species are easily distinguishable from its congeners by the structure and details of the genital double somite. It is unique in having two conical dorsal processes and also a ventral spine arising from the genital field. One of these processes might have been overlooked in previous descriptions (
The extremely long spiniform ventral process present in the genital double-somite of Pontellopsis lubbockii, is a unique character of this species and has not been hitherto described in or illustrated in previous works (
The mandibular dentition found in our specimens agrees with the pattern described by
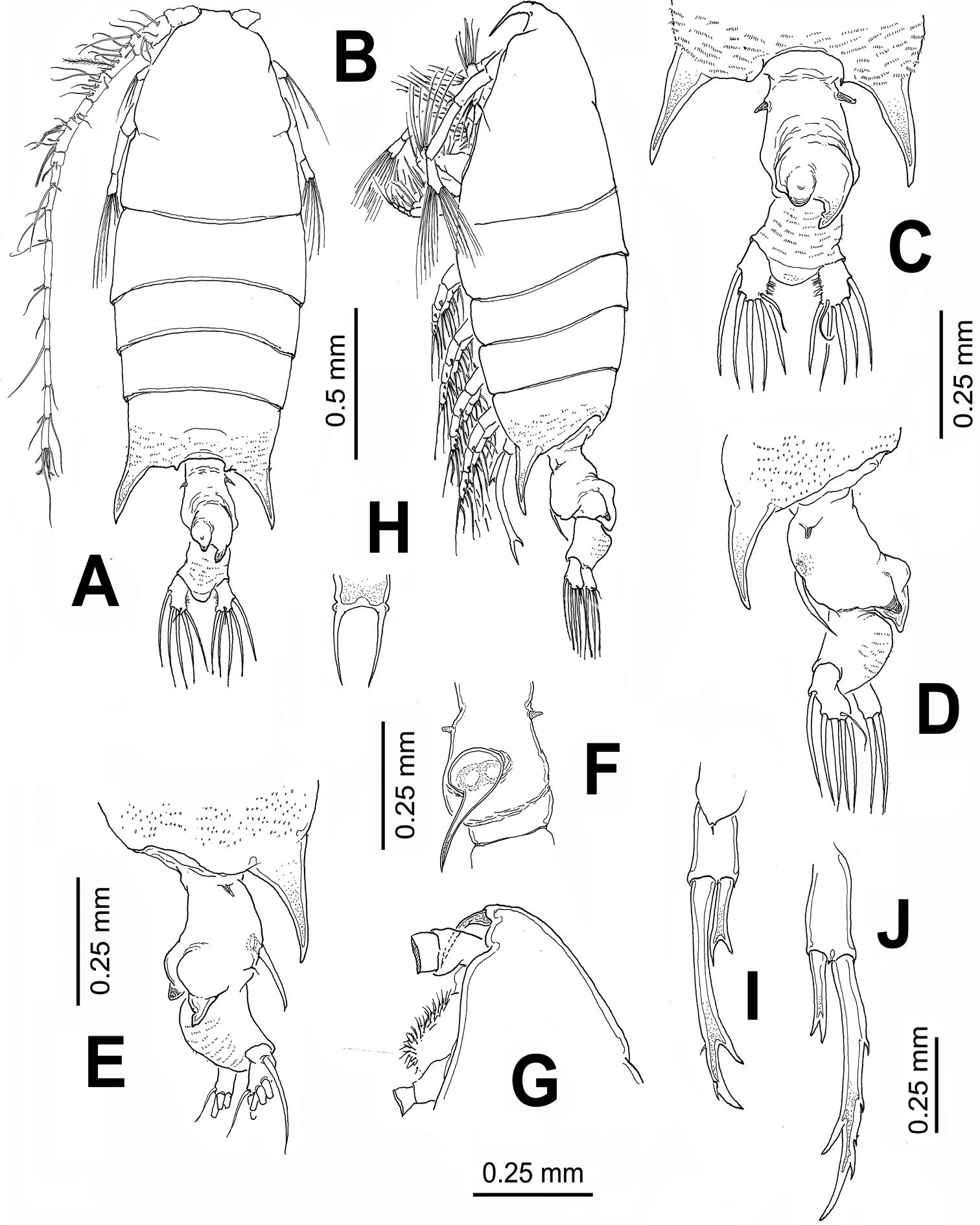
Pontellopsis lubbockii (Giesbrecht) from the Mexican Pacific. Adult female A habitus, dorsal view B same, lateral view C urosome showing details of dorsal processes of genital double-somite, ventral view D same, left lateral view E same, right lateral view F genital double-somite, ventral view G cephalic section, lateral view H rostrum, ventral view I right leg 5 J left leg 5.
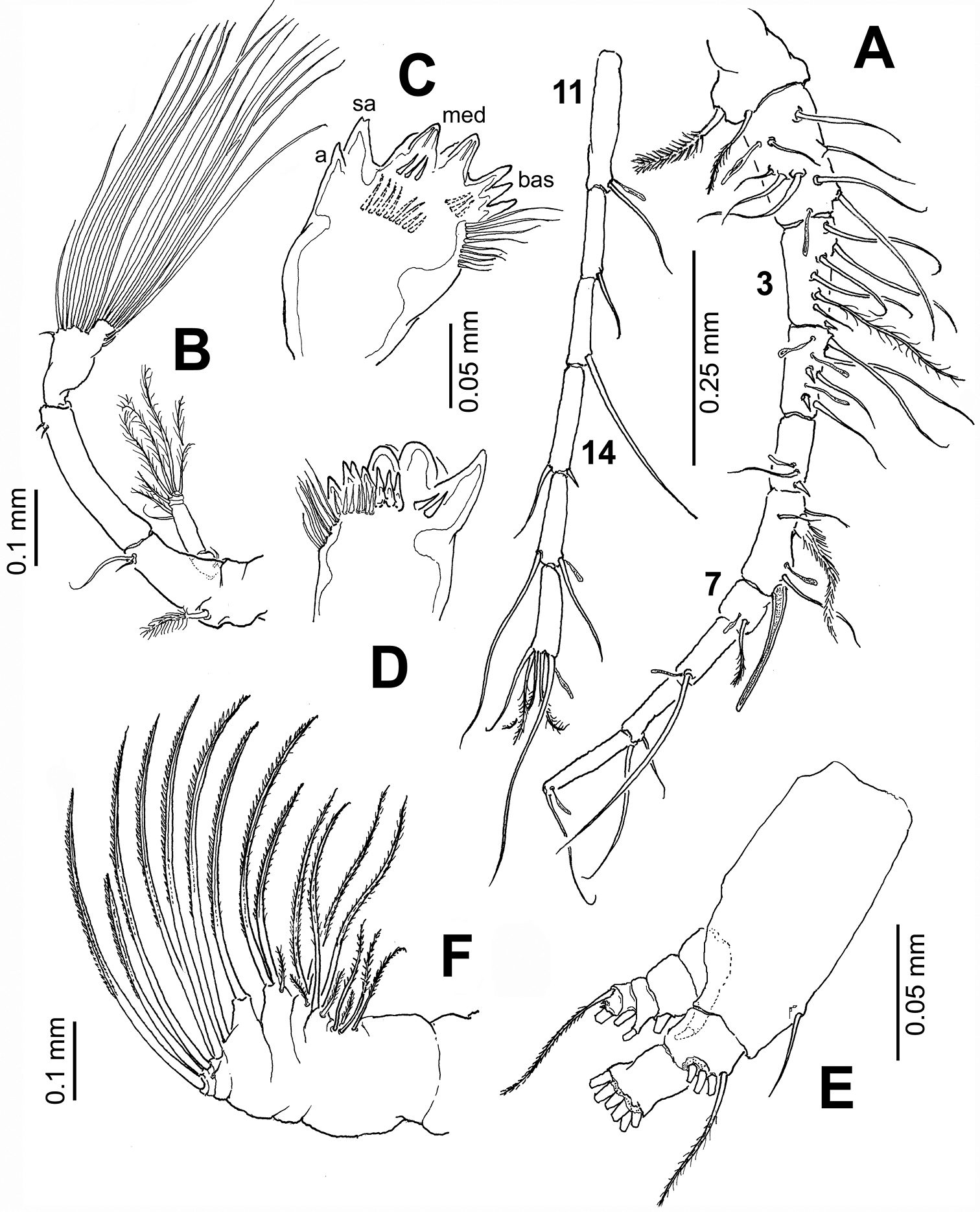
Pontellopsis lubbockii (Giesbrecht) from the Mexican Pacific. Adult female A antennule (in two sections) B antenna C mandible edge showing dentition, apical (a), subapical (sa), medial (med), and basal (bas) teeth D same, another view E mandibular palp F maxilla.
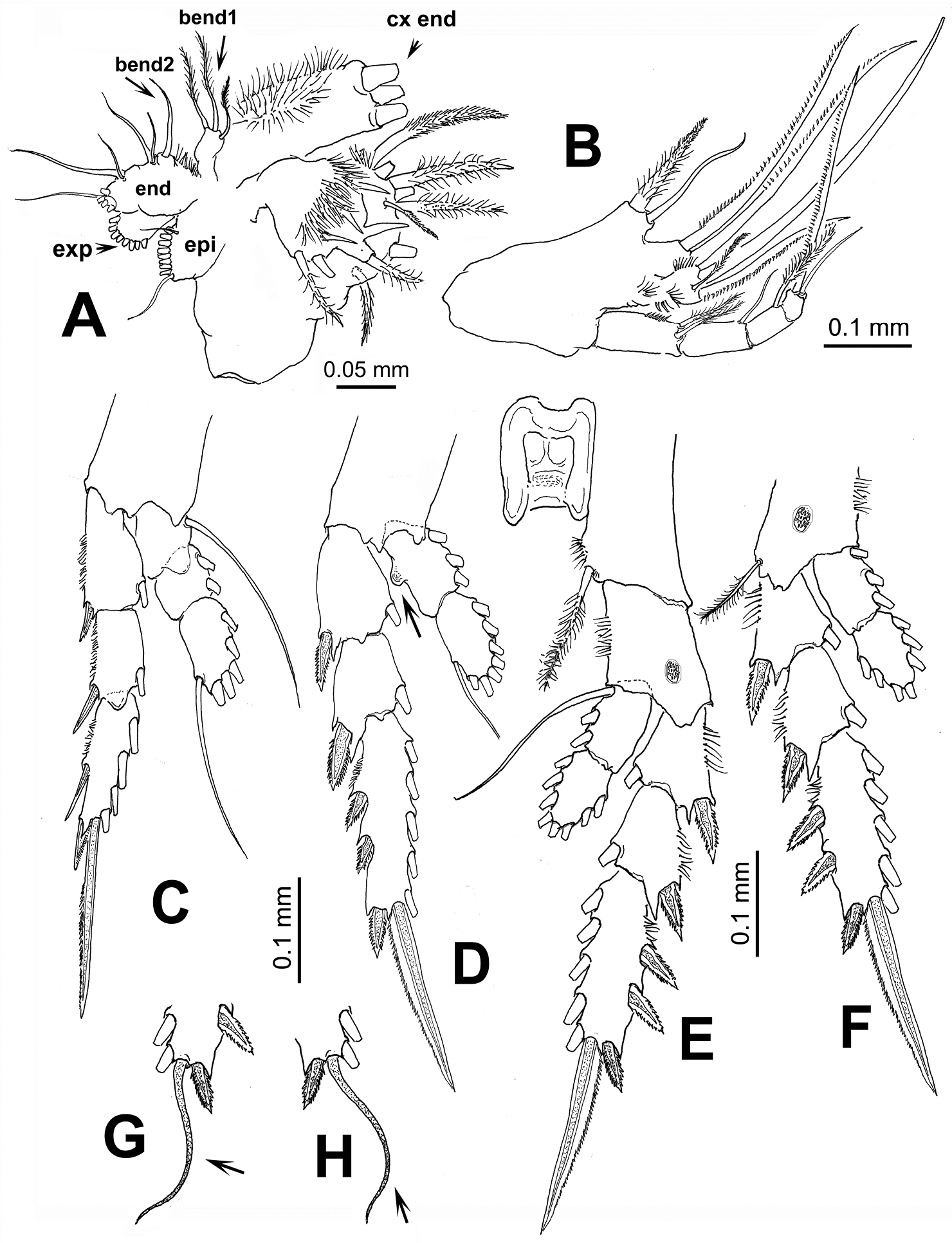
Pontellopsis lubbockii (Giesbrecht) from the Mexican Pacific. Adult female A maxillule showing armature of coxal endite (cx end distal spiniform elements cut short), proximal basal endite (bend1), distal basal endite (bend2), epipodite (epi), exopod (exp), and endopod (end) B maxilliped C leg 1 D leg 2 E eg 3 F leg 4 G variant form of leg 3 third exopodal segment with flexible terminal setal element (arrowed) H same, leg 4.
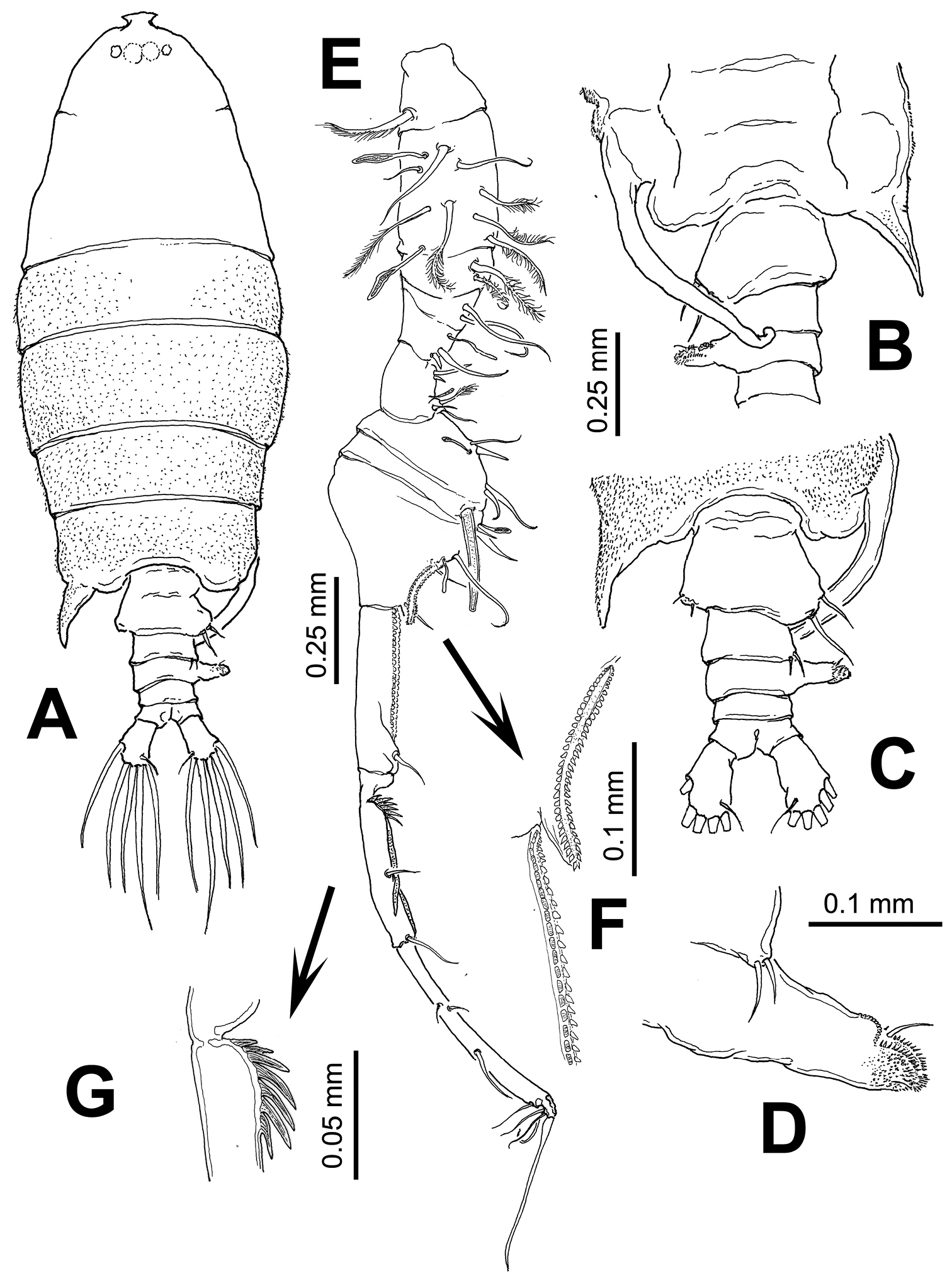
Pontellopsis lubbockii (Giesbrecht) from the Mexican Pacific. Adult male A habitus, dorsal view B urosome, ventral view C same, dorsal view D detail of process on right margin of third urosomite E geniculate antennule F detail of ornamentation on antennular segments 9 and 10 (arrowed) G detail of ornamentation of proximal part of antennular segment 11 (arrowed).
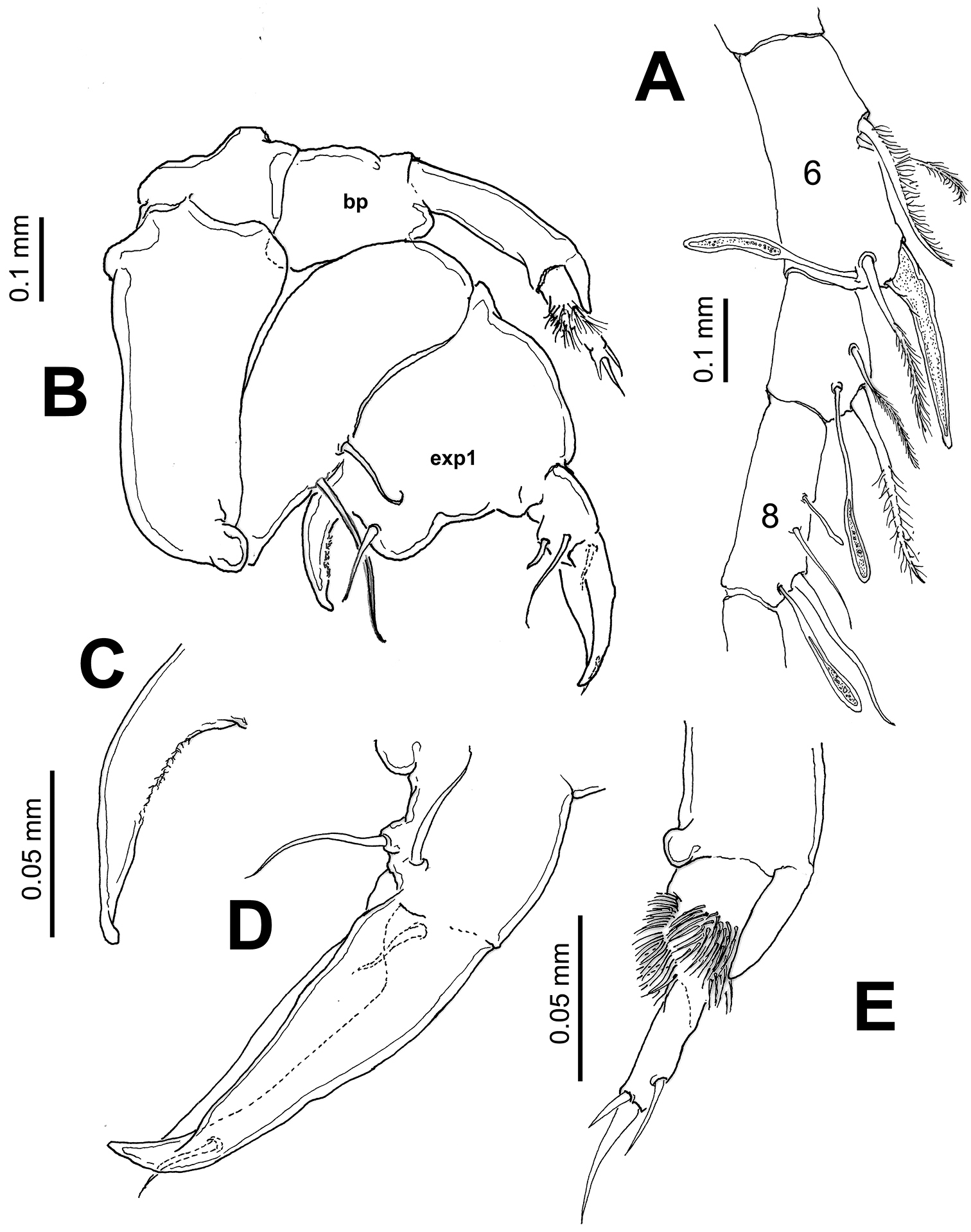
Pontellopsis lubbockii (Giesbrecht) from the Mexican Pacific. Adult male A left antennule, segments 6–8 showing spiniform process on segment 6 B. leg 5 showing basipod (bp) of left ramus and first exopodal segment of right ramus (exp1) C right leg, detail of basal thumb of chela D right leg, detail of second exopodal segment or distal finger of chela E left leg, distal segments and ornamentation.
In the Eastern Pacific, particularly in the California Current region, only a few species of Pontellopsis have been recorded: Pontellopsis occidentalis Esterly, 1906, Pontellopsis regalis (Dana, 1849), and Pontellopsis lubbockii. Pontellopsis occidentalis is regarded as endemic of southern California, the Gulf of California, and Baja California area. Pontellopsis regalis is frequently found in waters of the ETP (
| 1A | Posterolateral corners of fifth pedigerous somite with terminally rounded processes (Figs 6A, E) | 2 |
| 1B | Posterolateral corners of fifth pedigerous somite forming acute spiniform processes (Fig. 6B, C, D, F) | 3 |
| 2A | Genital double-somite elongate, with 2 acute dorsal processes of unequal size on posterior half of somite (Fig. 6E) | Pontellopsis yamadae |
| 2B | Genital double-somite with 2 unequal spiniform processes, one small, one long, in right side of posterior half of somite (Fig. 6A) | Pontellopsis tenuicauda |
| 3A | Spiniform processes of fifth pedigerous somite reaching the middle length of anal somite or beyond (Fig. 6C, D) | 4 |
| 3B | Spiniform processes of fifth pedigerous somite not as long, barely reaching the posterior margin of the genital double-somite or even shorter (Fig. 6 B, F) | 5 |
| 4A | Genital double-somite with strong, thumb-like process on left margin. Anal somite half the length of genital double-somite (Fig. 6D) | Pontellopsis villosa |
| 4B | Genital double-somite without distinct process. Anal somite as long as genital double-somite (Fig. 6C) | Pontellopsis armata |
| 5A | Genital double-somite as long as or slightly longer than anal somite, with processes or expansions on both margins or on dorsal surface | 6 |
| 5B | Genital double-somite twice as long as anal somite, with lateral process on right margin only | Pontellopsis occidentalis |
| 6A | Genital double- somite with two dorsal conical unequal protuberances | Pontellopsis lubbockii |
| 6 | BGenital double- somite with no such dorsal processes | 7 |
| 7A | Both lateral margins of genital double-somite expanded forming nearly symmetrical rounded processes, that on the right side globular; anal somite strongly produced between caudal rami (Fig. 6B) | Pontellopsis perspicax |
| 7B | Genital double-somite with asymmetrical, rounded lateral processes, anal somite not strongly produced between caudal rami (Fig. 6F) | Pontellopsis regalis |
| 1A | Posterolateral corners of fifth pedigerous somite with symmetrical or nearly symmetrical processes (Fig. 6G) | 2 |
| 1B | Posterolateral corners of fifth pedigerous somite with strongly asymmetrical processes, with long, slender, curved process on the right side (Fig. 6H) | 3 |
| 2A | Second urosomite with small lateral process on the left margin; second exopodal segment of left leg 5 cylindrical, as long as preceding segment (Fig. 6O), process on first exopodal segment of right leg 5 very short, distally blunt | Pontellopsis occidentalis |
| 2B | Second urosomite without such process on left margin; second exopodal segment of left leg 5 globose, half as long as preceding segment (Fig. 6P), process on first exopodal segment of right leg 5 short, distally acute | Pontellopsis villosa |
| 3A | Left posterolateral corner of fifth pedigerous somite forming short terminally rounded or broadly subtriangular process, not acute (Fig. 6I) | 4 |
| 3B | Left posterolateral corner of fifth pedigerous somite forming relatively long acute process (Fig. 6H) | 5 |
| 4A | Second and third urosomites with weak lateral expansions (Fig. 6I), process on first exopodal segment of right leg 5 long, distally truncate (arrow in Fig. 6L) | Pontellopsis tenuicauda |
| 4B | Second and/or third urosomites with lateral expansion on right side, process on first exopodal segment of right leg 5 long, tapering distally (Fig. 6K, M, N) | 7 |
| 5A | First urosomite symmetrical, armed with small unequal setae inserted on posterolateral margin | 6 |
| 5B | First urosomite clearly asymmetrical, with rounded process on right lateral margin; process armed with two long, stout setae | Pontellopsis lubbockii |
| 6A | Right posterolateral corner of fifth pedigerous somite long, acute, tapering distally (Fig. 6I); caudal rami as long as wide, distal segment of chela with protuberance on medial position of inner margin (arrow Fig. 6K) | Pontellopsis regalis |
| 6B | Right posterolateral corner of fifth pedigerous somite long, slender from insertion, branch-like (Fig. 6H); caudal rami twice as long as wide, distal segment of chela with low proximal expansion on inner margin | Pontellopsis armata |
| 7A | Second and third urosomites expanded laterally, process on first exopodal segment of right leg 5 shorter than second exopodal segment (Fig. 6N) | Pontellopsis yamadae |
| 7B | Only third urosomite expanded laterally. Right leg 5 with finger-like process of first exopodal segment longer than second exopodal segment (Fig. 6M) | Pontellopsis perspicax |
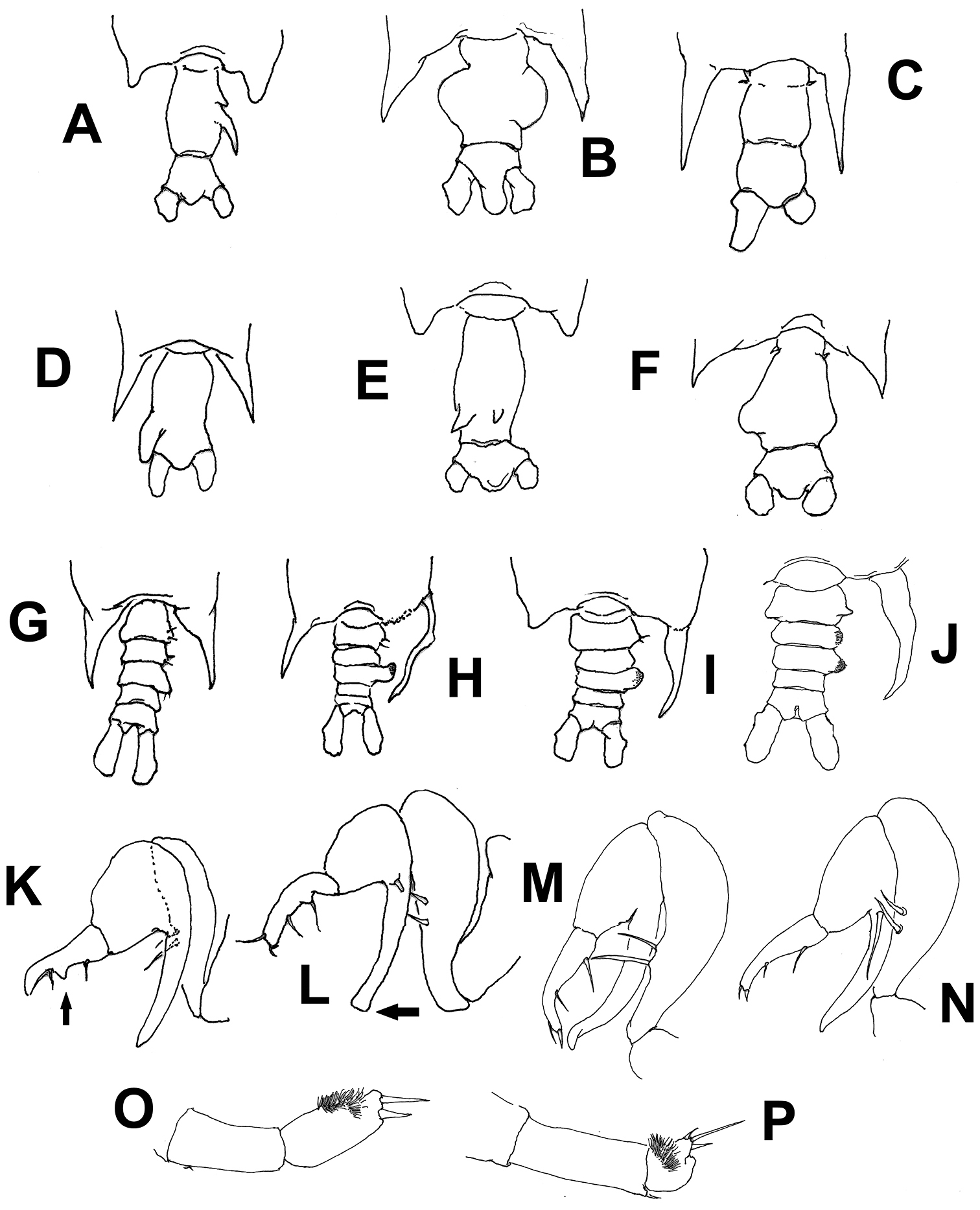
Schematic illustrations of characters used in the identification key to species of Pontellopsis from the Eastern Tropical Pacific. Explanation in key couplets. Illustrations modified from
This work is part of the doctoral thesis presented by E.K. in the postgraduate program of the Universidad de Guadalajara. Carmen Franco-Gordo and Enrique González-Domínguez (CUCSUR-Universidad de Guadalajara) kindly granted access to this material. Funding for fieldwork and general logistic support was provided by CUCSUR, Universidad de Guadalajara. We thank the crew of the R/V ‘BIP-V’ for their help and guidance during these years of sampling. Rosa Ma. Hernández deposited the specimens examined in the collection of Zooplankton at El Colegio de la Frontera Sur (ECOSUR), Chetumal, Mexico. The comments and valuable suggestions of two referees allowed us to improve a previous version of this contribution, particularly in reference to the interpretation and description of morphological details of the mouthparts. The fine editorial work and processing of the manuscript by Danielle Defaye is deeply appreciated.