





(C) 2014 Eknarin Rodcharoen. This is an open access article distributed under the terms of the Creative Commons Attribution License (CC BY 4.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
For reference, use of the paginated PDF or printed version of this article is recommended.
Citation: Rodcharoen E, Bruce NL, Pholpunthin P (2014) Cirolana songkhla, a new species of brackish-water cirolanid isopod (Crustacea, Isopoda, Cirolanidae) from the lower Gulf of Thailand. ZooKeys 375: 1–14. doi: 10.3897/zookeys.375.6573
Cirolana songkhla sp. n. was collected from brackish-water habitats including lagoons and estuaries in the coastal zone of the lower Gulf of Thailand. C. songkhla sp. n. is described and fully illustrated; C. songkhla sp. n. can be recognized by the presence of abundant chromatophores dorsally, lack of ornamentation on the posterior pereonites, pleonites and pleotelson, the number of robust setae on the uropodal and pleotelson margins (uropod exopod lateral margin with 12–14 RS, mesial margin with 5–8 RS; endopod lateral margin with 8–10 RS, mesial margin with 11–13 RS; pleotelson with 12–15 RS) and lack of setae on the endopods of pleopods 3–5. A dichotomous key of brackish Cirolana species in Thailand is given.
Isopoda, Cirolanidae, Cirolana, new species, brackish water, Thailand
The family Cirolanidae has received little attention in Thailand and South-East Asia in general, with only 13 species in eight genera known from Thailand. The genus Cirolana Leach, 1818, the largest genus in the family with 129 species (
This present report corrects Chilton’s record of Cirolana parva, describes Cirolana songkhla sp. n., and presents a key of the brackish-water species of Cirolana that occur in Thailand.
Specimens were collected by using baited traps from brackish-water habitats including the lagoon and estuary in the lower Gulf of Thailand (Figure 1). Appendages were dissected and drawn under stereo and compound microscopes with a camera lucida. Morphological characters for the description (based on
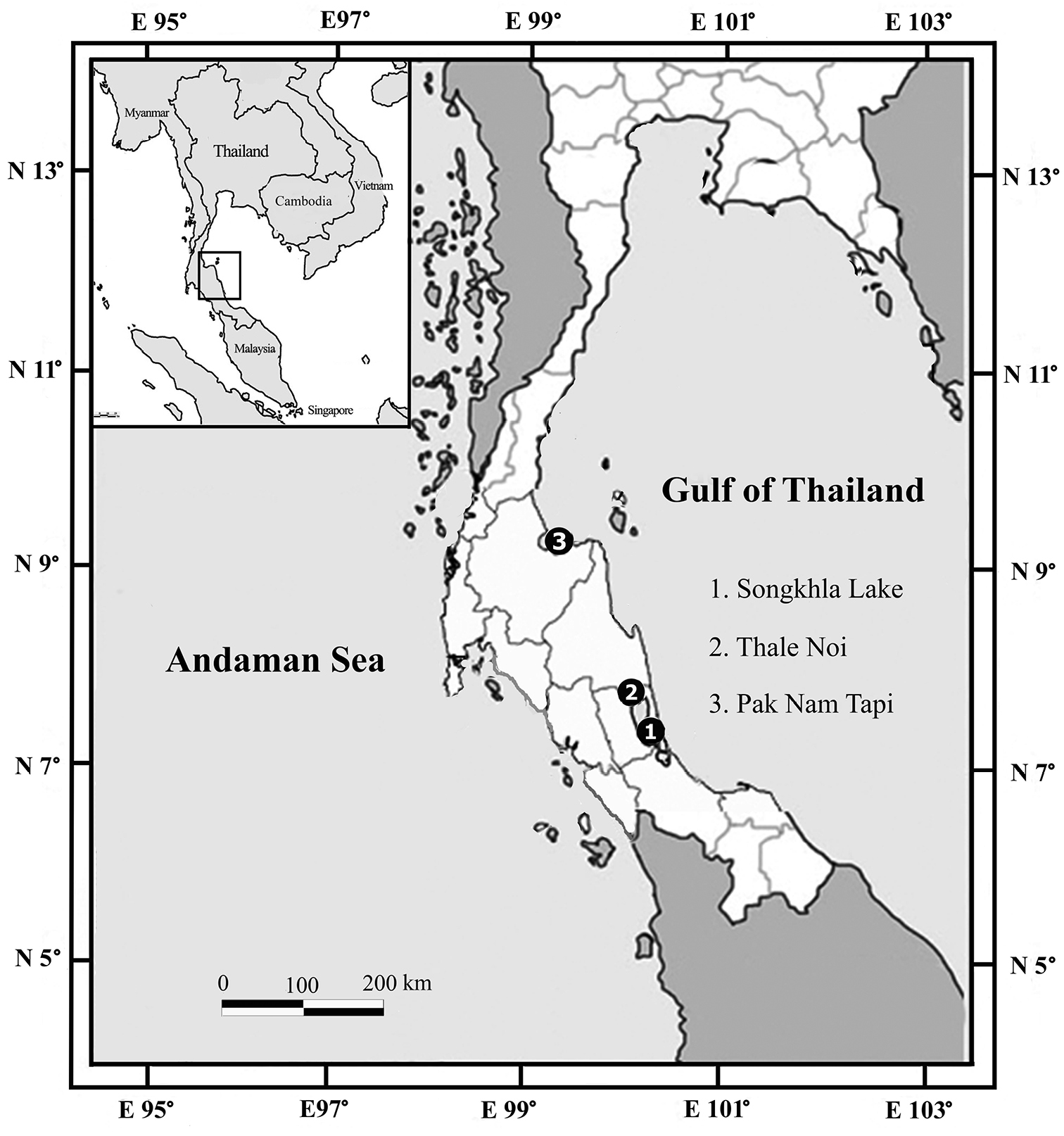
Map of sampling sites.
Abbreviations: PMS, plumose marginal setae; RS, robust seta/setae; CPS, circumplumose setae.
Cirolana is the largest genus of the Cirolanidae (
Diagnoses to the genus have been given by
http://zoobank.org/40967001-288A-433E-8B14-A411034CE8A7
http://species-id.net/wiki/Cirolana_songkhla
Figs 2–6Holotype, ♂ (13.7 mm), middle part of Songkhla Lake, Phattalung province, 07°29.09'N, 100°20.11'E, 23 October 2011, gravel bottom and associated with water plants, salinity 4 ppt, coll. E. Rodcharoen (PSUZC-CR0281-01).
Paratypes, 10 ♂ (13.8, 11.2, 12.4, 10.2, 10.3, 10.0, 13.8, 11.8, 11.4, 10.6 mm [dissected]), 1 ♀ (ovig. 8.7 mm [dissected]), further specimens unmeasured, same data as holotype (PSUZC-CR0281-02; MTQ W34265). 8 ♂ (11.1, 12.8, 12.2, 13.9, 13.2, 12.4, 12.2, 9.8 mm [dissected]), 4 ♀ (ovig. 8.6, 8.7 [dissected], 8.8, 9.0 mm), further specimens unmeasured, Klong Ban Klang, Thale Noi, Phattalung province, 07°46.44'N, 100°09.27'E, 27 May 2013 clay bottom, salinity 0.6 ppt, coll. E. Rodcharoen (PSUZC-CR0281-03; MTQ W34266).
12 ♂ (unmeasured), same data as holotype (PSUZC-CR0281-04), 38 ♂ and 3 ♀ (unmeasured), same data as paratype PSUZC-CR0281-03; MTQ W34266 (PSUZC-CR0281-05; MTQ W34267), 1 adult ♂ and 3 juvenile (unmeasured), Pak Nam Tapi (estuary), Surat Thani province, 09°10.31'N, 99°21.36'E, 30 October 2012 coll. E. Rodcharoen (PSUZC-CR0281-06).
Body 3.2 times as long as greatest width, dorsal surfaces smooth, widest at pereonite 5 and pereonite 6, lateral margins subparallel (Figure 2A). Rostral point (Figure 2C) present, folded ventrally and posteriorly, in contact with frontal lamina (Figure 2D, E). Eye colour dark brown; eyes separated by about 81% width of head (Figure 2C). Pereonite 1 and coxae 2–3 each with posteroventral angle rounded; coxae 5–7 with entire oblique carina; posterior margins of pereonites 5–7 smooth (Figure 2B). Pleon (Figure 2F) with pleonite 1 largely concealed by pereonite 7; pleonites 3–5 posterior margins smooth; posterolateral angles of pleonite 2 forming acute point, extending posteriorly to anterior of pleonite 4; pleonite 3 with posterolateral margins extending clearly beyond posterior margins of pleonites 4 and 5, narrowly rounded; posterolateral margin of pleonite 4 rounded, clearly extending beyond posterior margin of pleonite 5. Pleotelson (Figure 6E) 1.0 times as long as anterior width, dorsal surface without longitudinal carina; lateral margins convex; margins weakly serrate, posterior margin converging to small distinct caudomedial point, with 12 RS (Figure 6E, F).
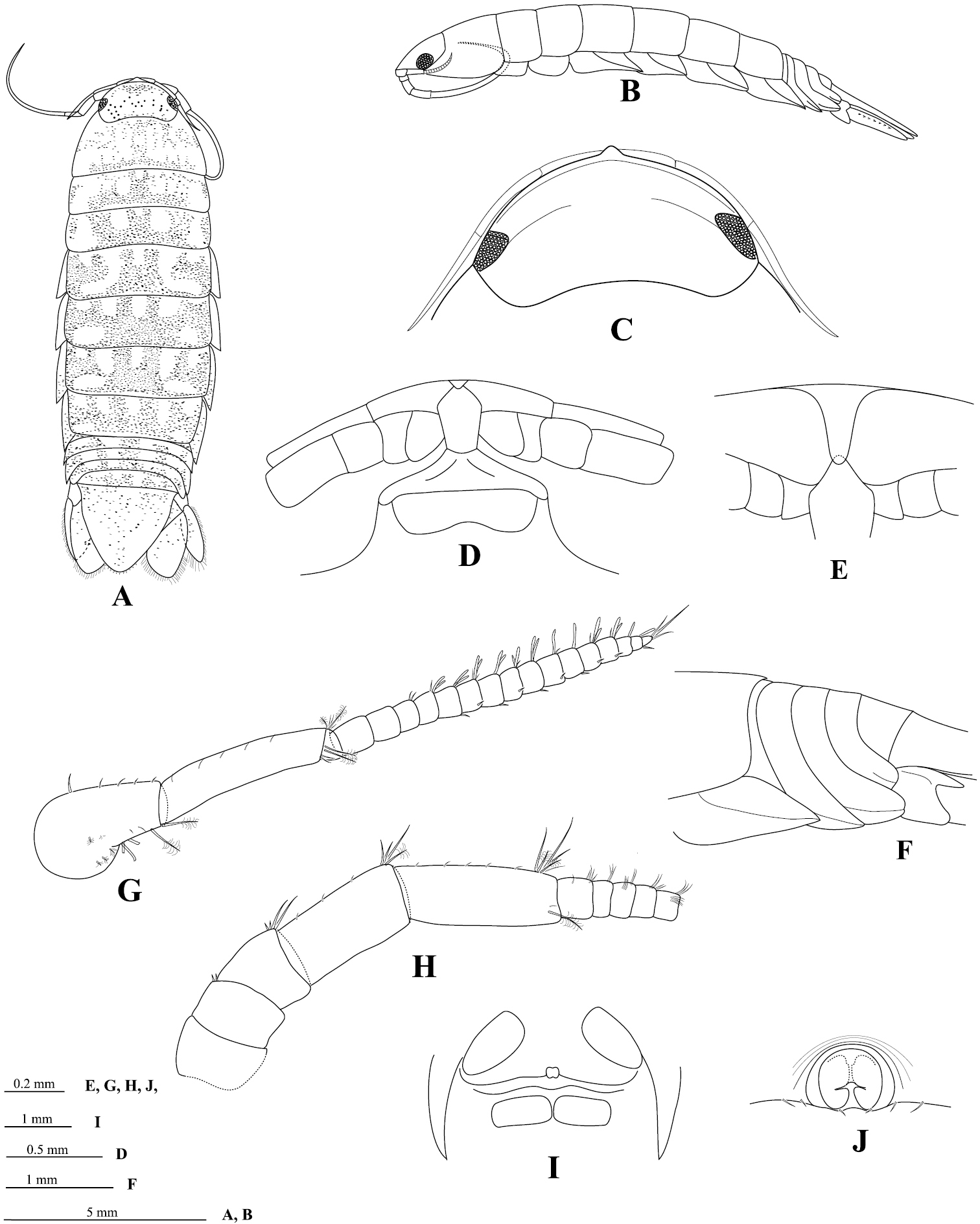
Cirolana songkhla sp. n., male holotype (PSUZC-CR0281-01) (13.7 mm) (A–F), male paratype (PSUZC-CR0281-2) (11.2 mm) (G–H), male paratype (PSUZC-CR0281-2) (13.8 mm) (I–J). A dorsal view B lateral view C head, dorsal view D frons E detail of frontal lamina F pleon G antennule H antennal peduncle I antero-ventral view of penial opening J ventral view of penial opening.
Antennule (Figure 2G) peduncle articles 1 and 2 entirely fused; articles 3 and 4 1.3 times as long as combined lengths of articles 1 and 2, article 3 5.0 times as long as wide; flagellum with 16 articles, extending to middle of pereonite 1. Antenna (Figure 2H) peduncle article 4 1.8 times as long as wide, 2.2 times as long as article 3, anterodistal angle with 3 short simple setae and 1 plumose seta; article 5 1.2 times as long as article 4, 2.4 times as long as wide, anterodistal angle with cluster of 4 short simple setae and 2 plumose setae; flagellum with 34 articles, extending to posterior of pereonite 4.
Frontal lamina (Figure 2D, E) pentagonal, 2.2 times as long as posterior width; 1.6 times as long as greatest width, lateral margins straight, diverging slightly towards anterior, anterior margin acute, with small median point.
Mandible molar process (Figure 3A) anterior margin with 19 flat teeth; with proximal cluster of long simple setae; right mandible spine row composed of 13 spines; palp article 2 with 21 distolateral setae, palp article 3 with 22 biserrate RS (Fig. 3B). Maxillule (Figure 3C) mesial lobe with 3 large circumplumose RS; lateral lobe with 13 RS (weakly serrated). Maxilla (Figure 3D) lateral lobe with 4 long setae; middle lobe with 12 long setae; mesial lobe with 4 distal plumose setae and 14 proximal plumose setae. Maxilliped palp (Figure 3E) article 1 mesial margin with 1 slender seta; article 2 mesial margin with 6 slender setae, lateral margin distally with 2 slender setae; article 3 mesial margin with 15 slender setae, lateral margin with 13 slender setae; article 4 mesial margin with 17 slender setae, lateral margin with 9 slender setae; article 5 distal margin with 18 setae, lateral margin with 6 setae; endite (Figure 3F) with 6 long CPS, and 2 coupling setae.
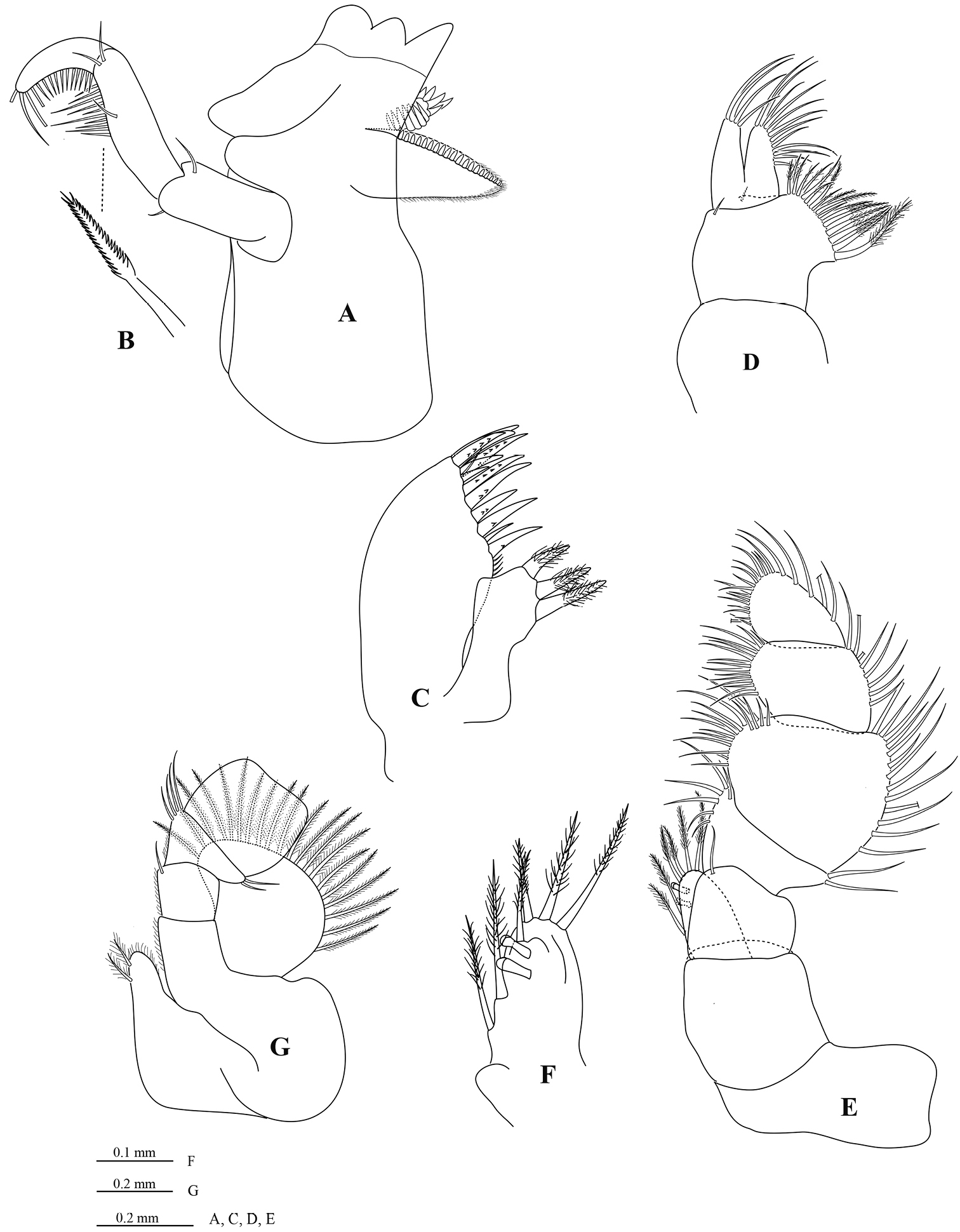
Cirolana songkhla sp. n., male paratype (PSUZC-CR0281-2) (11.2 mm) (A–F), ovigerous female paratype (PSUZC-CR0281-2) (8.7 mm) (G). A right madible B robust setae C right maxillule D right maxilla E right maxilliped F maxilliped endite G left maxiliped, basal articles.
Pereopod 1 (Figure 4A, C) basis 2.3 times as long as greatest width, inferior distal angle with cluster of 2 acute setae; ischium 0.5 times as long as basis, inferior margin with 1 acute seta, inferior distolateral margin with 2 setae (1 molariform RS and 1 acute seta), median distolateral margin with 2 acute setae, superior distal margin with 3 RS; merus inferior margin with 6 molariform RS, (set in rows of 4 and 2), superior distal angle with 3 setae (slender); carpus inferior distal margin with 2 setae (1 RS and 1 acute seta); propodus 2.0 times as long as wide, inferior margin with 2 RS; dactylus (Figure 4B) 0.5 times as long as propodus. Pereopod 2 (Figure 4D) ischium inferior margin with 2 stout, bluntly rounded RS, superior distal margin with 3 RS; merus inferior margin with 10 stout RS, set in one row, superior distal margin with 6 acute RS; carpus inferodistal angle with 3 RS (plus 1 slender seta); propodus 3.5 times as long as wide; dactylus 0.7 times as long as propodus. Pereopod 3 similar to pereopod 2. Pereopod 4 (Figure 4E) intermediate in from between pereopod 3 and pereopod 5. Pereopod 6 (Figure 4F) similar to pereopod 7. Pereopod 7 (Figure 4G) basis 3.0 times as long as greatest width, superior margin weakly convex, inferior margin with 3 palmate setae; ischium 0.8 times as long as basis, inferior margin with 8 RS (set in groups of 3, 3, 1 and 1), superior distal angle with 6 RS (5 simple, 1 biserrate), inferior distal angle with 2 RS; merus 0.5 times as long as ischium, 2.0 times as long as wide, inferior margin with 3 RS, superior distal angle with 6 RS (3 simple, 3 biserrate), inferior distal angle with 7 RS; carpus 0.7 times as long as ischium, 2.4 times as long as wide, inferior margin with 6 RS (set in groups of 2 and 4), superior distal angle with 11 RS (4 simple, 7 biserrate), inferior distal angle with 9 RS (6 simple, 3 biserrate); propodus 0.8 times as long as ischium, 5.4 times as long as wide, inferior margin with 7 RS (set in groups of 1, 2, 2 and 2), superior distal angle with 3 slender setae and 1 palm seta, inferior distal angle with 2 RS; dactylus 0.3 times as long as propodus.
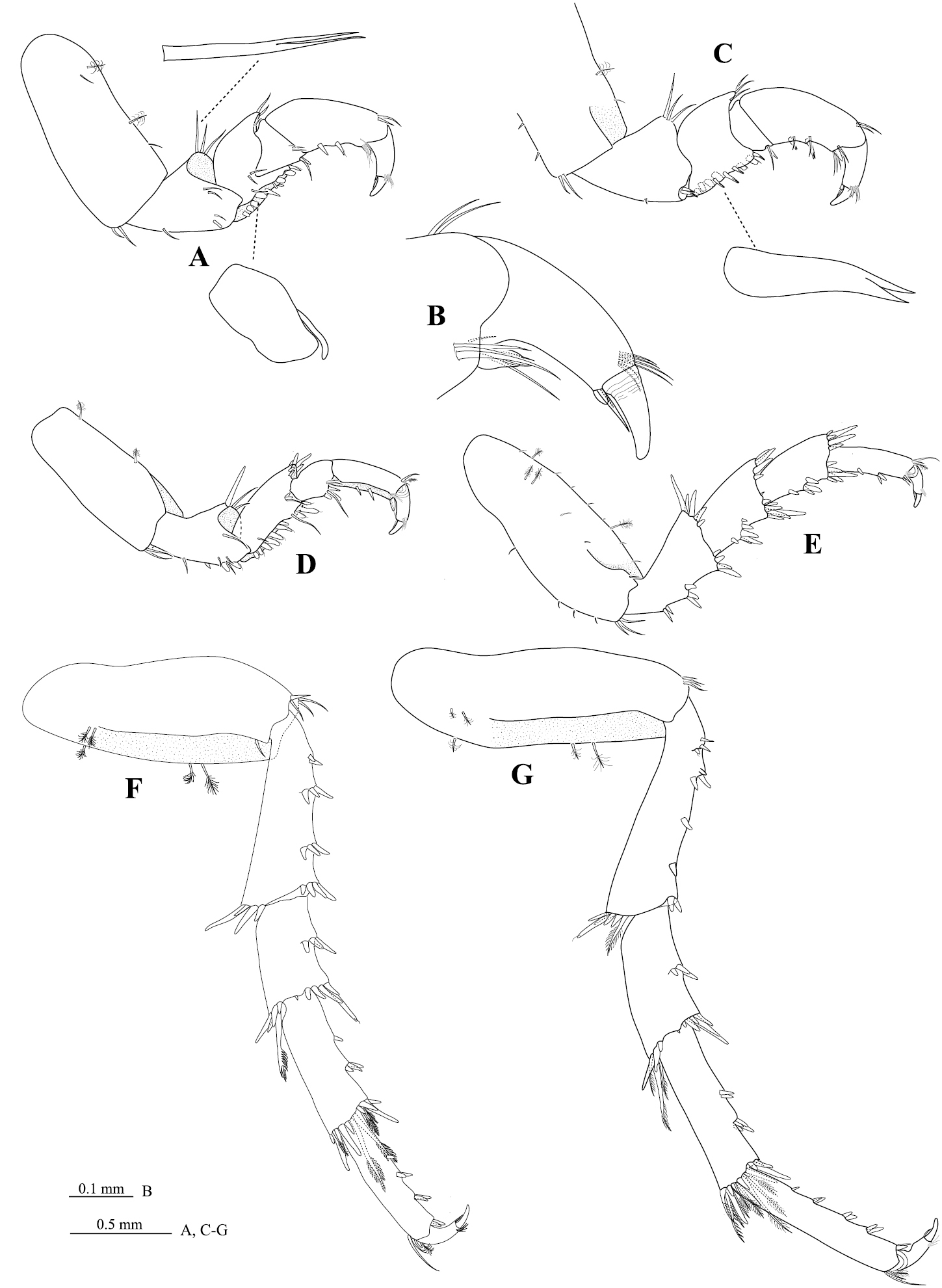
Cirolana songkhla sp. n., male paratype (PSUZC-CR0281-2) (11.2 mm). A pereopod 1 B dactylus of pereopod 1 C mesial view, pereopod 1 D pereopod 2 E pereopod 4 F pereopod 6 G pereopod 7.
Penes (Figure 2I, J) medially united low papillae.
Pleopod 1 (Figure 5A) exopod 1.3 times as long as wide, lateral margin weakly convex, distally broadly rounded, mesial margin weakly convex, with ~35 PMS from distal one-third; endopod 2.1 times as long as wide, distally broadly rounded, lateral margin sinuate, with ~16 PMS on distal margin only; peduncle 1.9 times as wide as long, mesial margin with 4 coupling hooks. Pleopod 2 (Figure 5B) exopod with ~52 PMS, endopod with ~24 PMS; appendix masculina with parallel margins, 1.0 times as long as endopod, distally notch. Pleopod 3 (Figure 5C) endopod without PMS, exopod with ~60 PMS. Pleopod 4 (Figure 5D) endopod without PMS, exopod with ~60 PMS. Pleopod 5 (Figure 5E) endopod without PSM, exopod with ~54 PMS. Pleopods 2–5 peduncle distolateral margin with prominent acute RS, 3–5 endopods with distomesial serrate scales.
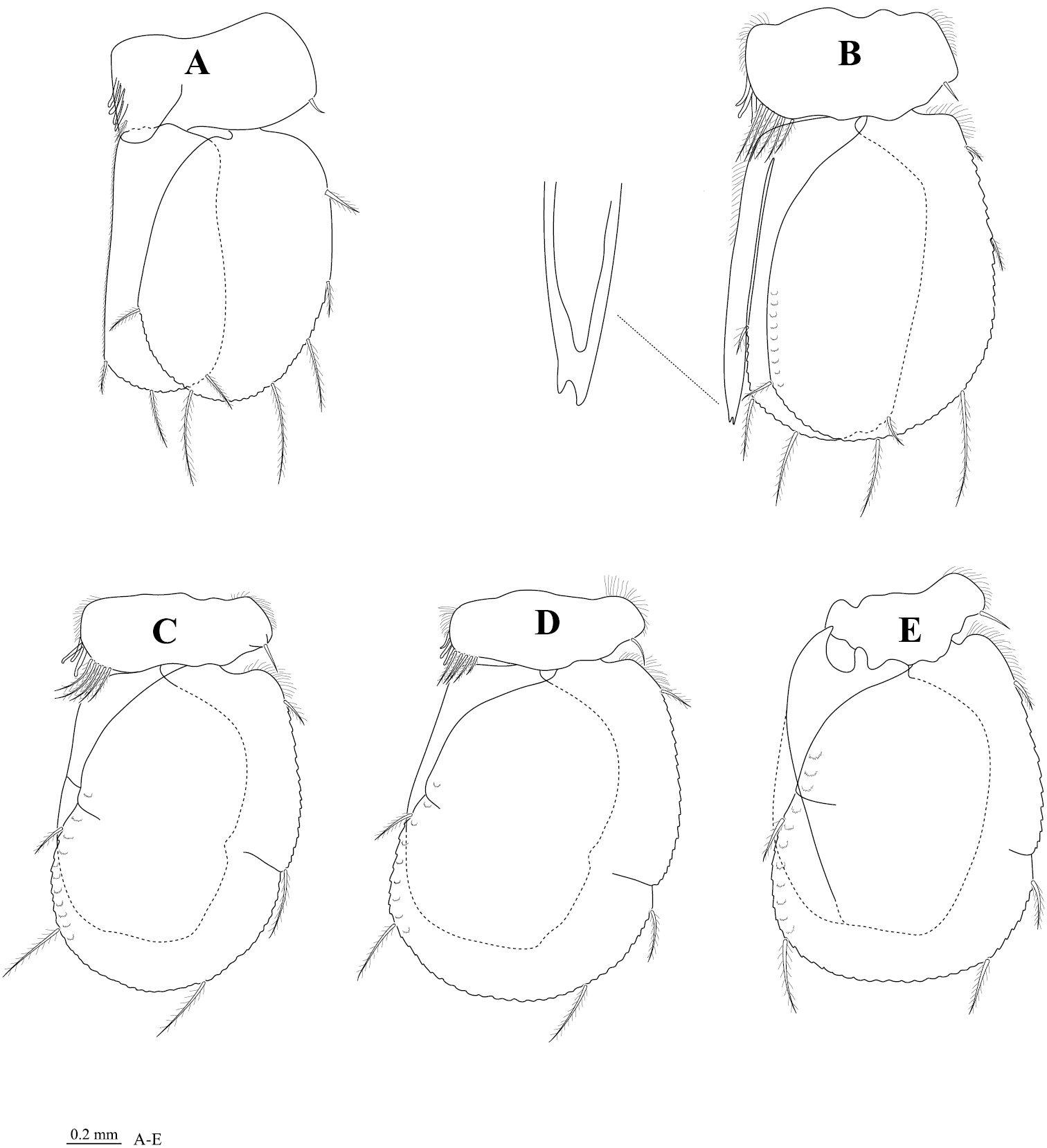
Cirolana songkhla sp. n., male paratype (PSUZC-CR0281-2) (11.2 mm) A–E pleopods 1–5 respectively.
Uropod (Figure 6A) peduncle ventrolateral margin (Figure 6D) with 2 RS, lateral margin with 1 mesial short acute RS, posterior lobe about one-half as long as endopod; rami extending beyond pleotelson, marginal setae in single tiers, apices acute. Endopod (Figure 6A) apically sub-bifid, medial process prominent (Figure 6C); lateral margin weakly convex, with 8 RS, mesial margin strongly convex, with 12 RS. Exopod (Figure 6A) not extending to end of endopod, 3.3 times as long as greatest width, apically sub-bifid, medial process prominent (Figure 6B); lateral margin weakly convex, with 13 RS; mesial margin weakly convex, with 6 RS.
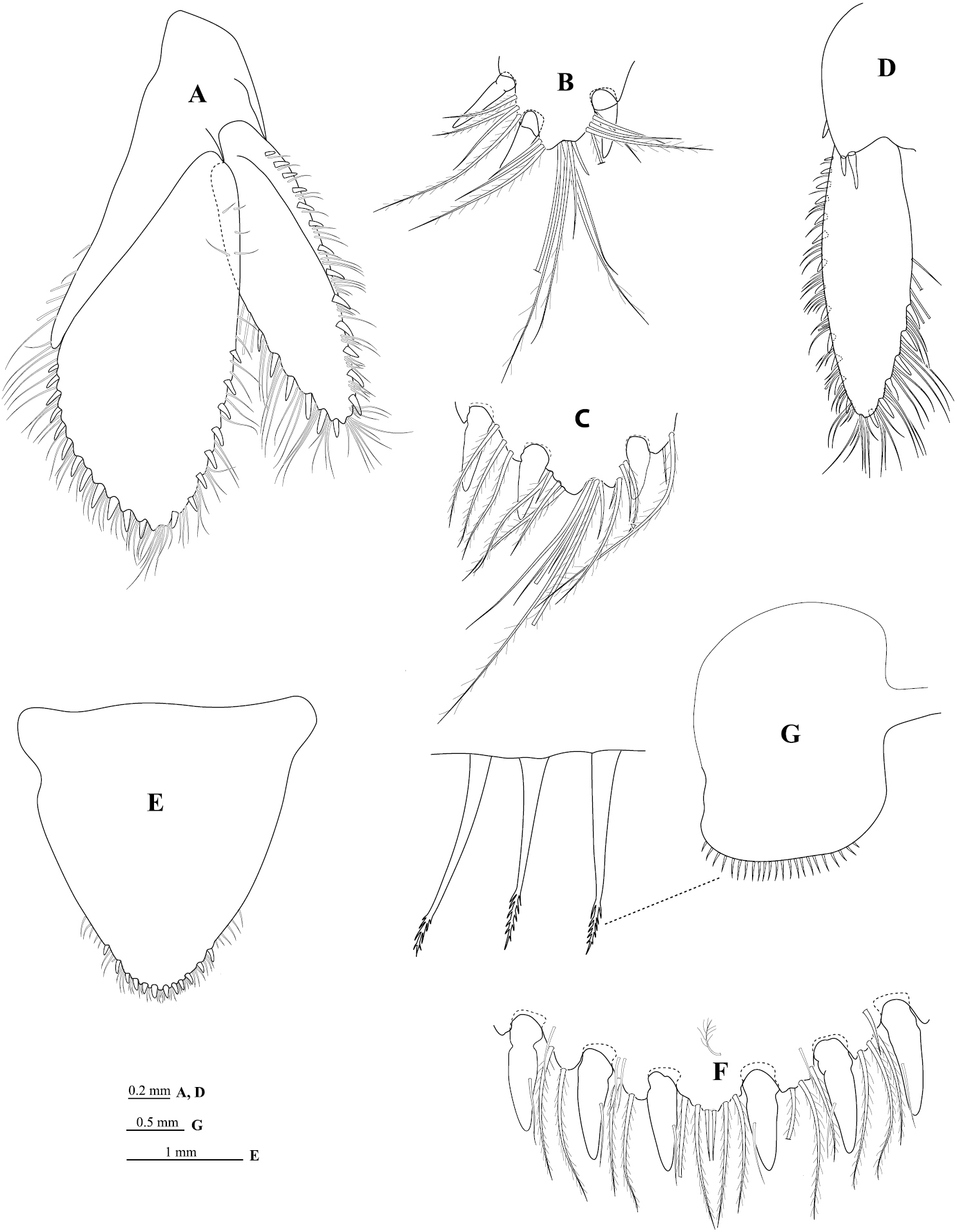
Cirolana songkhla sp. n., male paratype (PSUZC-CR0281-2) (11.2 mm) (A–F), ovigerous female (PSUZC-CR0281-2) (8.7 mm) (G). A uropod B uropod exopod apex C uropod endopod apex D uropod exopod E pleotelson F detail of pleotelson apex G oostegite 4.
Similar to male but on average smaller. Antennal flagellum slightly longer, extending to anterior of pereonite 5; maxilliped with lamina vibrans (Figure 3G); brood pouch composed of 5 pairs of oostegites (Figure 6G) arising on sternites 1–5 (as recorded for some other Cirolana species, such as Cirolana (Anopsilana) barnardi (Bruce, 1992) and Cirolana kokoru Bruce, 2004), lateral margin of oostegite 4 with ~23 slender setae.
Pleotelson (n=23 [18♂ and 5♀]) with 12–15 RS, with 14 RS (7+7) most frequent (74%), 12 (4%) and 15 (4%) occurring only once. Uropod endopod mesial margin with 11–13 RS, with 12 (52%) and 11(39%) most frequent, lateral margin with 8–10 RS, with 8 (52%) and 9 (39%) most frequent; exopod mesial margin with 5–8 RS, with 7 (39%) and 6 (35%) most frequent, lateral margin with 12–14, with 13 (74%) most frequent.
Adult males (n=19) 9.8–13.9 mm (mean 11.9 mm); ovigerous females (n=5) 8.6–9.0 mm (mean 8.8 mm).
The presence of abundant chromatophores and lack of ornamentation on the posterior pereonites, pleonites and pleotelson excludes Cirolana songkhla sp. n. from the Cirolana ‘tuberculate-group’ (see
Cirolana songkhla sp. n. is characterized by lacking plumose setae on endopods of pleopods 3–5. This character is particularly associated with brackish and freshwater cirolanid species, formerly placed in the genus and then subgenus Anopsilana (
Only Cirolana barnardi (Bruce, 1992) from tropical eastern Australia is similar to Cirolana songkhla sp. n. having in common a smooth dorsal surface, rostral point and pentagonal frontal lamina. However, the two species can be clearly distinguished by body size of Cirolana songkhla sp. n., which is larger than that of Cirolana barnardi (male average at 11.9 VS 3.9 mm, ovigerous female at 8.8 VS 4.0 mm). Furthermore, Cirolana songkhla sp. n. has more numerous robust setae on the uropodal rami and pleotelson margin than Cirolana barnardi; uropod exopod lateral margin with 12–14 RS (VS 7–10), mesial margin with 5–8 RS (VS 5–6); endopod lateral margin with 8–10 RS (VS 3–4), mesial margin with 11–13 RS (VS 5–7) and pleotelson, posterior margin converging to small distinct caudomedial point (VS posterior margin subtruncate), with 12–15 RS (VS 7–10).
Cirolana parva Hansen, 1890 has been recorded in freshwater from Thailand by
Cirolana songkhla sp. n. is named for the type locality.
| 1 | Anterior margin of head without rostral point; frontal lamina anterior margin rounded | Cirolana fluviatilis |
| – | Anterior margin of head with rostral point, folded ventrally and posteriorly, in contact with frontal lamina; frontal lamina pentagonal | 2 |
| 2 | Body dorsal surface without ornamentation; pleotelson lateral margins convex | Cirolana songkhla sp. n. |
| – | Body dorsal surface with tubercles on pereonites, pleonites and pleotelson; pleotelson lateral margins concave | Cirolana willeyi |
The first author would like to thank Miss Bongkot Wichachucherd (Seaweed and Seagrass Research Unit) and Mr. Phuripong Meksuwan (Plankton Research Unit) department of biology, Prince of Songkla University for helping to collect the samples, and we are grateful to department of biology, Prince of Songkla University for the laboratory facilities. This work was financed through Graduated School; the Higher Education Research Promotion and National Research University Project of Thailand, Office of the Higher Education Commission, Prince of Songkla University.